
Paleoenvironmental Changes in the Gulf of Gaeta (Central Tyrrhenian Sea, Italy): A Perspective from Benthic Foraminifera after Dam Construction

 ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
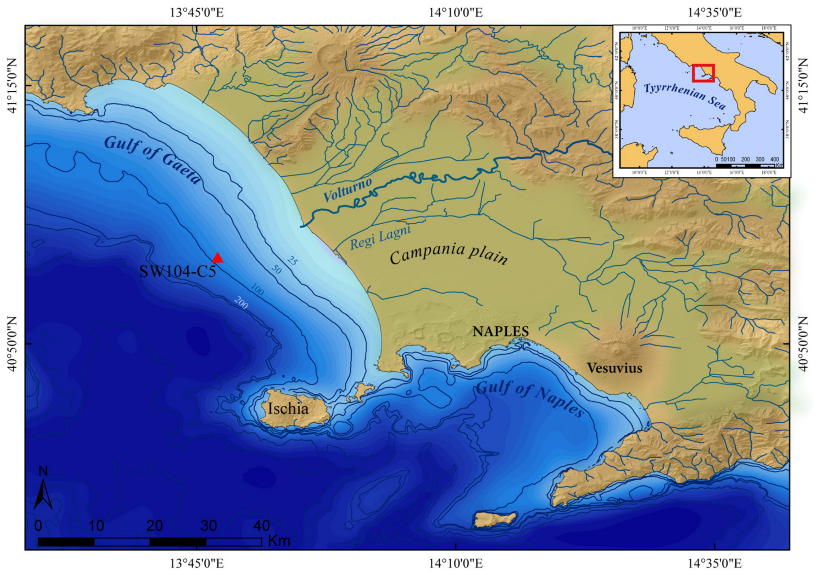
2.1. Study Area
2.2. Core Sampling and Geochronology
2.3. Benthic Foraminiferal Analysis
2.4. Statistical Analysis
3. Results
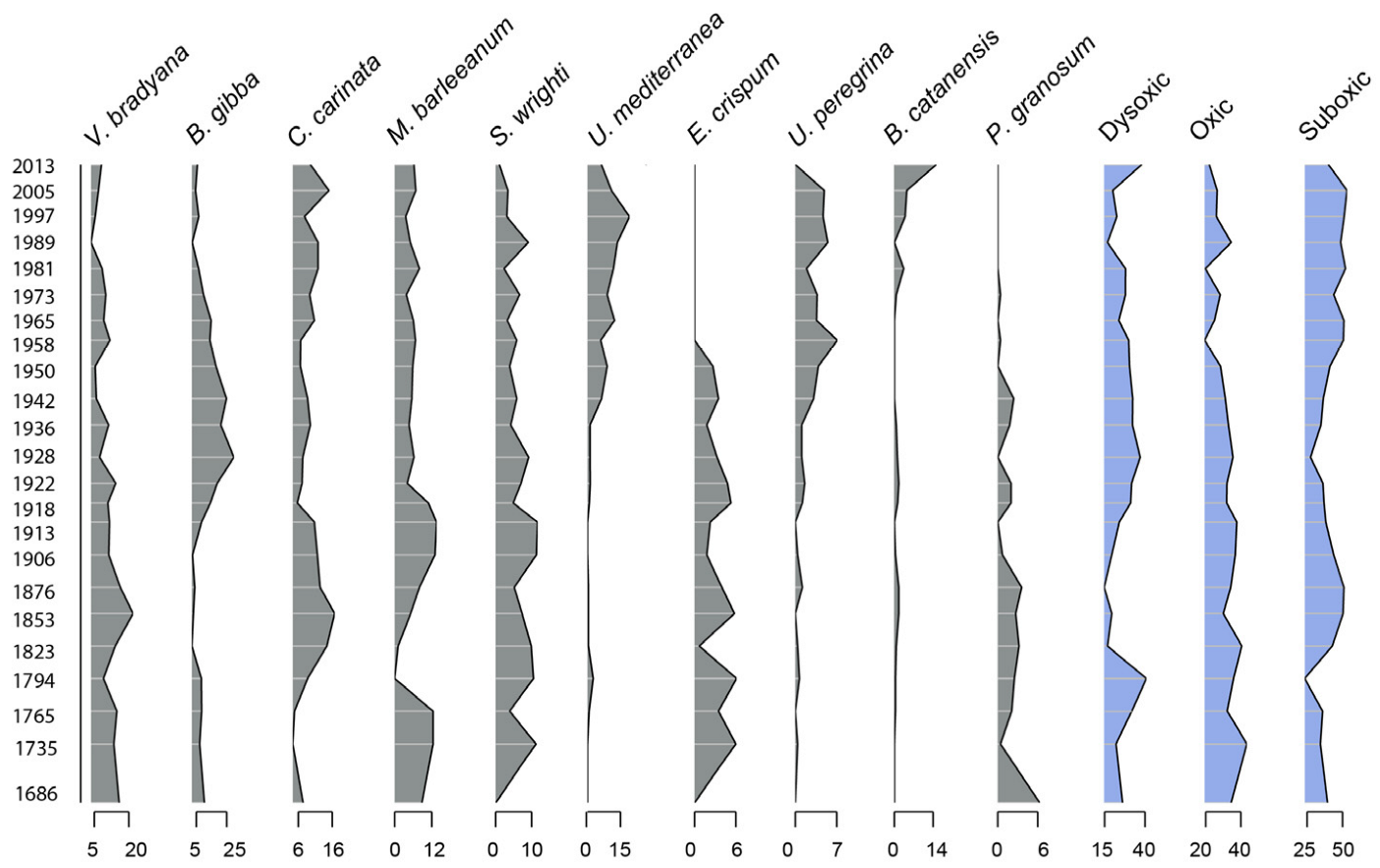
3.1. Diversity Indices and Species Abundance Variations
3.2. Changes in the Benthic Foraminiferal Composition
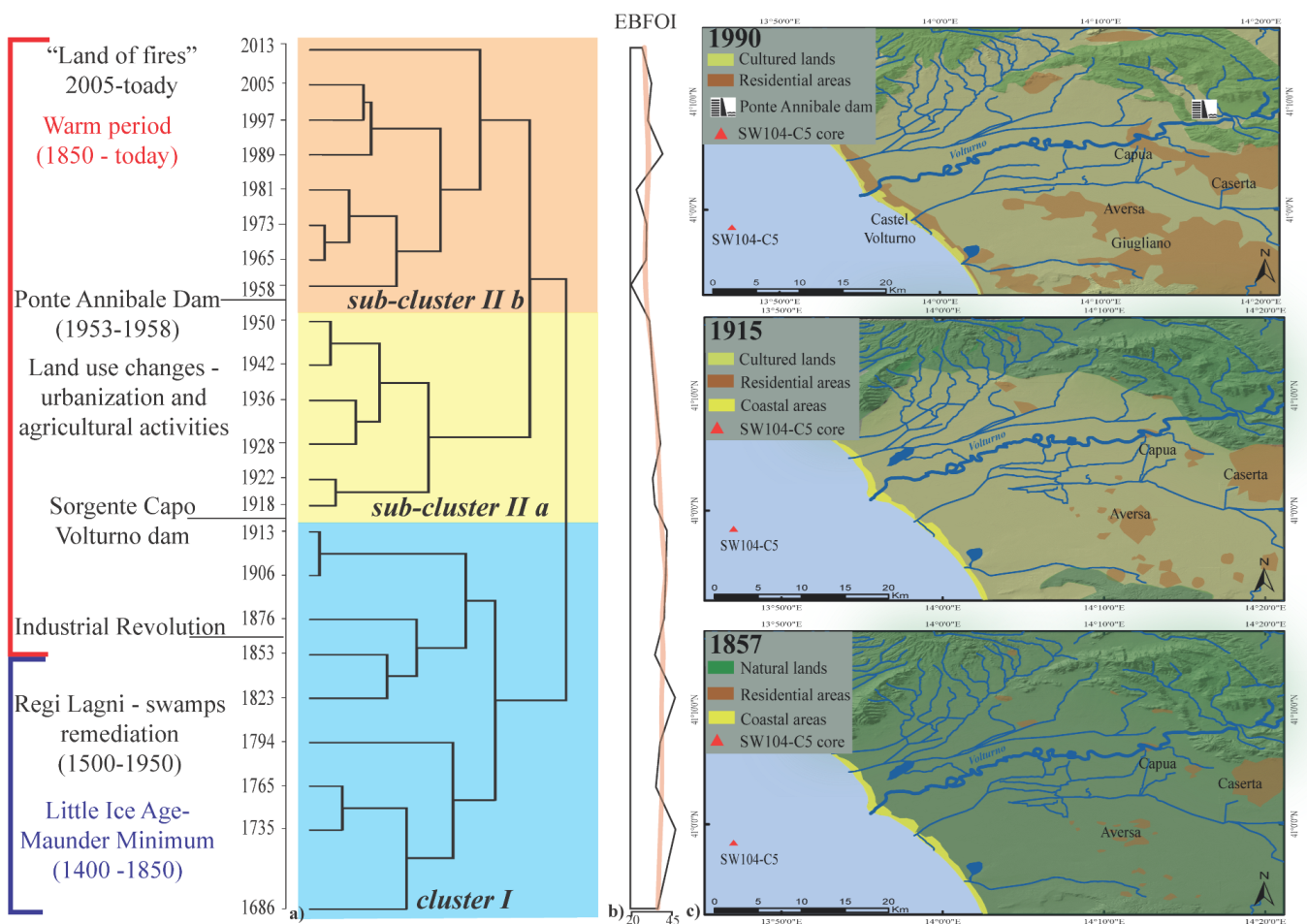
3.3. Historical Environmental Changes: The Imprint of Human Activities
4. Discussion
4.1. Natural Conditions and the Regi Lagni Reclamation
4.2. The Aftermath of the Construction of the Capo Volturno Dam
4.3. Changes Associated with the Ponte Annibale Dam
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Frontalini, F.; Semprucci, F.; Armynot du Châtelet, É.; Francescangeli, F.; Margaritelli, G.; Rettori, R.; Spagnoli, F.; Balsamo, M.; Coccioni, R. Biodiversity trends of the meiofaunal and foraminiferal assemblages of Lake Varano (Southern Italy). Proc. Biol. Soc. Wash. 2014, 127, 7–22. [Google Scholar] [CrossRef]
- Armynot du Châtelet, E.; Francescangeli, F.; Frontalini, F. Definition of benthic foraminiferal bioprovinces in transitional environments of the Eastern English Channel and the Southern North Sea. Rev. Micropaléontol. 2018, 61, 223–234. [Google Scholar] [CrossRef]
- Francescangeli, F.; Milker, Y.; Bunzel, D.; Thomas, H.; Norbisrath, M.; Schönfeld, J.; Schmiedl, G. Recent benthic foraminiferal distribution in the Elbe Estuary (North Sea, Germany): A response to environmental stressors. Est. Coast. Shelf Sci. 2021, 251, 107198. [Google Scholar] [CrossRef]
- Cavaliere, M.; Barrenechea Angeles, I.; Montresor, M.; Bucci, C.; Brocani, L.; Balassi, E.; Margiotta, F.; Francescangeli, F.; Bouchet, V.M.P.; Pawlowski, J.; et al. Assessing the ecological quality status of the highly polluted Bagnoli area (Tyrrhenian Sea, Italy) using foraminiferal eDNA metabarcoding. Sci. Total Environ. 2021, 790, 147871. [Google Scholar] [CrossRef]
- Hayward, B.W.; Wilson, K.; Morley, M.S.; Cochran, U.; Grenfell, H.R.; Sabaa, A.T.; Daymond-King, R. Microfossil record of the Holocene evolution of coastal wetlands in a technologically active region of New Zealand. Holocene 2010, 20, 405–421. [Google Scholar] [CrossRef]
- Francescangeli, F.; Portela, M.; Armynot du Chalet, E.; Billon, G.; Andersen, T.J.; Bouchet, V.M.P.; Trentesaux, A. Infilling of the Canche Estuary (eastern English Channel, France): Insight from benthic foraminifera and historical pictures. Mar. Micropal. 2018, 142, 1–12. [Google Scholar] [CrossRef]
- Alve, E.; Lepland, A.; Magnusson, J.; Backer-Owe, K. Monitoring strategies for re- establishment of ecological reference conditions: Possibilities and limitations. Mar. Pollut. Bull. 2009, 59, 297–310. [Google Scholar] [CrossRef]
- Dos, S.; de Jesus, M.; Frontalini, F.; Bouchet, V.M.P.; Yamashita, C.; Sartoretto, J.R.; Figueira, R.C.L.; de Mello, E.; Sousa, S.H. Reconstruction of the palaeo-ecological quality status in an impacted estuary using benthic foraminifera: The Santos Estuary (São Paulo state, SE Brazil). Mar. Environ. Res. 2020, 162, 105121. [Google Scholar] [CrossRef]
- Barrenechea Angeles, I.; Romero-Martínez, M.L.; Cavaliere, M.; Varrella, S.; Francescangeli, F.; Piredda, R.; Mazzocchi, M.G.; Montresor, M.; Schirone, A.; Delbono, I.; et al. Encapsulated in sediments: eDNA deciphers the ecosystem history of one of the most polluted European marine sites. Environ. Int. 2023, in press. [Google Scholar] [CrossRef]
- Francescangeli, F.; Armynot du Chatelet, E.; Billon, G.; Trentesaux, A.; Bouchet, V.M. Palaeo-ecological quality status based on foraminifera of Boulogne-sur-Mer harbour (Pas-de-Calais, Northeastern France) over the last 200 years. Mar. Environ. Res. 2016, 117, 32–43. [Google Scholar] [CrossRef]
- da Silva, L.C.; Alves, M.M.V.; Figueira, R.; Frontalini, F.; Pereira, E.; Senez-Mello, T.M.; Castelo Wellen, F.L.; Saibro, M.B.; Francescangeli, F.; Mello e Sousa, S.H.; et al. Unravelling Anthropocene Paleoenvironmental Conditions Combining Sediment and Foraminiferal Data: Proof-of-Concept in the Sepetiba Bay (SE, Brazil). Front. Ecol. Evol. 2022, 10, 852439. [Google Scholar] [CrossRef]
- Horrocks, M.; Nichol, S.L.; D’Costa, D.M.; Augustinus, P.; Jacobi, T.; Shane, P.A.; Middleton, A. A late quaternary record of natural change and human impact from Rangihoua Bay, bay of islands, northern New Zealand. J. Coast. Res. 2007, 233, 592–604. [Google Scholar] [CrossRef]
- MacDonald, D.G.; Carlson, J.; Goodman, L. On the heterogeneity of stratified-shear turbulence: Observations from a near-field river plume. J. Geophys. Res. Oceans. 2013, 118, 6223–6237. [Google Scholar] [CrossRef]
- Horner-Devine, A.R.; Hetland, R.D.; MacDonald, D.G. Mixing and transport in coastal river plumes. Annu. Rev. Fluid Mech. 2015, 47, 569–594. [Google Scholar] [CrossRef]
- Wright, L.D. Sediment transport and deposition at river mouths: A synthesis. GSA Bull. 1977, 88, 857–868. [Google Scholar] [CrossRef]
- De Pippo, T.; Donadio, C.; Pennetta, M. Morphological control on sediment dispersal along the southern Tyrrhenian coastal zones (Italy). Geol. Romana. 2003, 37, 113–121. [Google Scholar]
- Budillon, F.; Esposito, E.; Iorio, M.; Pelosi, N.; Porfido, S.; Violante, C. The geological record of storm events over the last 1000 years in the Salerno Bay (Southern Tyrrhenian Sea): New proxy evidences. Adv. Geosci. 2005, 2, 123–130. [Google Scholar] [CrossRef] [Green Version]
- Sacchi, E.; Conti, M.A.; D’Orazi Porchetti, S.; Logoluso, A.; Nicosia, U.; Perugini, G.; Petti, F.M. Aptian dinosaur footprints from the Apulia platform (Bisceglie, southern Italy) in the framework of the periadriatic ichnosites. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2009, 271, 104–116. [Google Scholar] [CrossRef]
- Lirer, F.; Sprovieri, M.; Ferraro, L.; Vallefuoco, M.; Capotondi, L.; Cascella, A.; Petrosino, P.; Insinga, D.D.; Pelosi, N.; Tamburrino, S.; et al. Integrated stratigraphy for the late Quaternary in the eastern Tyrrhenian Sea. Quat. Int. 2013, 292, 71–85. [Google Scholar] [CrossRef]
- Lirer, F.; Sprovieri, M.; Vallefuoco, M.; Ferraro, L.; Pelosi, N.; Giordano, L.; Capotondi, L. Planktonic foraminifera as bio-indicators for monitoring the climatic changes that have occurred over the past 2000 years in the southeastern Tyrrhenian Sea. Integr. Zool. 2014, 9, 542–554. [Google Scholar] [CrossRef]
- Bonomo, S.; Cascella, A.; Alberico, I.; Sorgato, S.; Pelosi, N.; Ferraro, L.; Lirer, F.; Vallefuoco, M.; Agnini, C.; Pappone, G.; et al. Reworked Coccoliths as runoff proxy for the last 400 years: The case of Gaeta Gulf (Central Tyrrhenian Sea, Central Italy). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2016, 459, 15–28. [Google Scholar] [CrossRef]
- Margaritelli, G.; Vallefuoco, M.; Di Rita, F.; Capotondi, L.; Bellucci, L.G.; Insinga, D.D.; Petrosino, P.; Bonomo, S.; Cacho, I.; Cascella, A.; et al. Marine response to climate changes during the last five millennia in the central Mediterranean Sea. Glob. Planet. Change 2016, 142, 53–72. [Google Scholar] [CrossRef]
- Di Rita, F.; Lirer, F.; Bonomo, S.; Cascella, A.; Ferraro, L.; Florindo, F.; Insinga, D.D.; Lurcock, P.C.; Margaritelli, G.; Petrosino, P.; et al. Late Holocene forest dynamics in the Gulf of Gaeta (central Mediterranean) in relation to NAO variability and human impact. Quat. Sci. Rev. 2018, 179, 137–152. [Google Scholar] [CrossRef]
- Misuraca, M.; Budillon, F.; Tonielli, R.; Di Martino, G.; Innangi, S.; Ferraro, L. Coastal Evolution, Hydrothermal Migration Pathways and Soft Deformation along the Campania Continental Shelf (Southern Tyrrhenian Sea): Insights from High-Resolution Seismic Profiles. Geosciences 2018, 8, 121. [Google Scholar] [CrossRef] [Green Version]
- D’Alisa, G.; Armiero, M.; De Rosa, S.P. Political ecology: Rethink Campania’s toxic-waste scandal. Nature 2015, 509, 427. [Google Scholar] [CrossRef] [Green Version]
- Legambiente. Rapporto Ecoma I Numeri e le Storie Della Criminalità Ambientale Legambiente: Roma, Italy. 2014. Available online: http://www.amblav.it/download/0417_ecomafia_introduzione.pdf (accessed on 31 March 2020).
- Triassi, M.; Montuori, P.; Provvisiero, D.P.; De Rosa, E.; Di Duca, F.; Sarnacchiaro, P.; Díez, S. Occurrence and spatial-temporal distribution of atrazine and its metabolites in the aquatic environment of the Volturno River estuary, southern Italy. Sci. Total Environ. 2022, 803, 149972. [Google Scholar] [CrossRef]
- Astraldi, M.; Gasparini, G.P. The seasonal characteristics of the circulation in the Tyrrhenian Sea. In Seasonal and Interannual Variability of the Western Mediterranean Sea Coastal and Estuarine Studies; La Violette, P., Ed.; American Geophysical Union: Washington, DC, USA, 1994; Volume 46, pp. 115–134. [Google Scholar]
- Radicati di Brozolo, F.; Di Girolamo, P.; Turi, B.; Oddone, M. 40Ar/39Ar and K-Ar dating of K-rich rocks from the Roccamonfina volcano, roman comagmatic region, Italy. Geoch. Cosmo. Acta 1988, 52, 1435–1441. [Google Scholar] [CrossRef]
- de Alteriis, G.; Fedi, M.; Passaro, P.; Siniscalchi, A. Magneto-seismic interpretation of subsurface volcanism in the Gaeta Gulf (Italy, Tyrrhenian Sea). Ann. Geophys. 2006, 49, 4–5. [Google Scholar] [CrossRef]
- Iermano, I.; Liguori, G.; Iudicone, D.; Nardelli, B.; Colella, S.; Zingone, A.; Saggiomo, V.; Ribera d’Alcalà, M. Filament formation and evolution in buoyant coastal waters: Observation and modelling. Prog. Oceanogr. 2012, 106, 118–137. [Google Scholar] [CrossRef]
- Bonomo, S.; Cascella, A.; Alberico, I.; Ferraro, L.; Giordano, L.; Lirer, F.; Vallefuoco, M.; Marsella, E. Coccolithophores from near the Volturno estuary (central Tyrrhenian Sea). Mar. Micropaleontol. 2014, 111, 26–37. [Google Scholar] [CrossRef]
- Moretti, M.; Sansone, E.; Spezie, G.; Vultaggio, M.; De Maio, A. Alcuni aspetti del movimento delle acque del Golfo di Napoli. Annali IUN 1977, 45–46, 207–217. [Google Scholar]
- Sorgente, R.; Di Maio, A.; Pessini, F.; Ribotti, A.; Bonomo, S.; Perilli, A.; Alberico, I.; Lirer, F.; Cascella, A.; Ferraro, L. Impact of Freshwater Inflow From the Volturno River on Coastal Circulation. Front. Mar. Sci. 2020, 7, 293. [Google Scholar] [CrossRef]
- Nava, M.L.; Giampaola, D.; Laforgia, E.; Boenzi, G. Tra il Clanis e il Sebeto: Nuovi dati sull’occupazione della piana campana tra il Neolitico e l’eta del Bronzo. Atti della XL Riun. Sci. Ist. It. di Preist. e Prot. 2007, 1, 101–126. [Google Scholar]
- Amorosi, A.; Pacifico, A.; Rossi, V.; Ruberti, D. Late Quaternary incision and deposition in an active volcanic setting: The Volturno valley fill, southern Italy. Sediment. Geol. 2012, 282, 307–320. [Google Scholar] [CrossRef]
- Rezza, C.; Albanese, S.; Lima, A.; De Vivo, B. Geochemical Characterization, Isotopic Approach and Environmental Risk Assessment in the Domizio Flegreo and Agro Aversano Area (Campania Region). Available online: https://www.researchgate.net/publication/278689393_Geochemical_characterization_isotopic_constraints_and_environmental_risk_assessment_in_the_Domizio_Flegreo_and_Agro_Aversano_area_Campania_region (accessed on 22 December 2022).
- Mancini, L.; Cero, C.D.; Gucci, P.; Venturi, L.; Carlo, M.D.; Volterra, L. Il fiume Volturno-la definizione dello stato di salute del corpo idrico. Inquinamento 1994, 10, 70–83. [Google Scholar]
- Parrella, A.; Isidori, M.; Lavorgna, M.; Dell’Aquila, A. Stato di qualità del fiume Volturno integrato da indagini di tossicità e genotossicità. Ann. Ig. 2003, 15, 147–157. [Google Scholar]
- Montuori, P.; De Rosa, E.; Sarnacchiaro, P.; Di Duca, F.; Provvisiero, D.P.; Nardone, A.; Triassi, M. Polychlorinated biphenyls and organochlorine pesticides in water and sediment from Volturno River, Southern Italy: Occurrence, distribution and risk assessment. Environ. Sci. Eu. 2020, 32, 123. [Google Scholar] [CrossRef]
- Montuori, P.; De Rosa, E.; Di Duca, F.; Provvisiero, D.P.; Sarnacchiaro, P.; Nardone, A.; Triassi, M. Estimation of Polycyclic Aromatic Hydrocarbons Pollution in Mediterranean Sea from Volturno River, Southern Italy: Distribution, Risk Assessment and Loads. Int. J. Environ. Res. Public Health. 2021, 18, 1383. [Google Scholar] [CrossRef]
- Sgarrella, F.; Moncharmont Zei, M. Benthic foraminifera of the Gulf of Naples (Italy): Systematics and autoecology. Boll. Soc. Paleontol. Ital. 1993, 32, 145–264. [Google Scholar]
- Milker, Y.; Schmiedl, G. A taxonomic guide to modern benthic shelf foraminifera of the western Mediterranean Sea. Palaeontol. Electron. 2012, 15, 1–134. [Google Scholar] [CrossRef]
- Hayward, B.W.; Le Coze, F.; Vachard, D.; Gross, O. World Foraminifera Database. Foraminiferida. 2022. Available online: https://www.marinespecies.org/aphia.php?p=taxdetails&id=22528 (accessed on 22 December 2022).
- Chao, A.; Shen, T.-J. Nonparametric estimation of Shannon’ s index of diversity when there are unseen species in sample. Environ. Ecol. Stat. 2003, 10, 429–443. [Google Scholar] [CrossRef]
- Kaiho, K. Benthic foraminiferal dissolved-oxygen index and dissolved-oxygen levels in the modern ocean. Geology 1994, 22, 719–722. [Google Scholar] [CrossRef]
- Kranner, M.; Harzhauser, M.; Beer, C.; Auer, G.; Piller, W.E. Calculating dissolved marine oxygen values based on an enhanced Benthic Foraminifera Oxygen Index. Sci. Rep. 2022, 12, 1376. [Google Scholar] [CrossRef] [PubMed]
- Jorissen, F.J.; Barmawidjaja, D.M.; Puskaric, S.; van der Zwaan, G.J. Vertical distribution of benthic foraminifera in the northern Adriatic Sea: The relation with the organic flux. Mar. Micropalentol. 1992, 19, 131–146. [Google Scholar] [CrossRef]
- Jorissen, F.; Nardelli, M.P.; Almogi-Labin, A.; Barras, C.; Bergamin, L.; Bicchi, E.; El Kateb, A.; Ferraro, L.; McGann, M.; Morigi, C.; et al. Developing Foram-AMBI for biomonitoring in the Mediterranean: Species assignments to ecological categories. Mar. Micropaleontol. 2018, 140, 33–45. [Google Scholar] [CrossRef] [Green Version]
- Altenbach, A.V.; Pflaumann, U.; Schiebel, R.; Thies, A.; Timm, S.; Trauth, M. Scaling percentages and distributional patterns of benthic foraminifera with flux rates of organic carbon. J. Foram. Res. 1999, 29, 173–185. [Google Scholar]
- Fontanier, C.; Jorissen, F.J.; Licari, L.; Alexandre, A.; Anschutz, P.; Carbonel, P. Live benthic foraminiferal faunas from the Bay of Biscay: Faunal density, composition, and microhabitats. Deep Sea Res. Part I Oc. Res. P. 2002, 49, 751–785. [Google Scholar] [CrossRef]
- Frezza, V.; Guillem, M.V.; Gaglianone, G.; Baldassarre, A.; Brandano, M. Mixed carbonate-siliclastic sediments and benthic foraminiferal assemblages from Posidonia oceanica seagrass meadows of the central Tyrrhenian continental shelf (Latium, Italy). It. J. Geosci. 2011, 130, 352–369. [Google Scholar] [CrossRef]
- Hausser, J.; Strimmer, K. Estimation of Entropy, Mutual Information and Related Quantities; R Package Version; R Fundation for Statistical Computing: Vienna, Austria, 2014. [Google Scholar]
- Oksanen, O.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, B.; Simpson, G.L.; Solymos, P.; Stevens, H.; Wagner, H.H. Vegan: Community Ecology Package; R Fundation for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- Juggins, S. Rioja: Analysis of Quaternary Science Data. R Package Version 1.0-5. 2022. Available online: https://cran.r-project.org/web/packages/rioja/rioja.pdf (accessed on 31 October 2022).
- Grimm, E.C. CONISS: A FORTRAN 77 program for stratigraphically constrained cluster analysis by the method of incremental sum of squares. Comp. Geosci. 1987, 13, 13–35. [Google Scholar] [CrossRef]
- Juggins, S. Rioja-Package. Analysis of Quaternary Science Data. 2020. Available online: https://cran.r-project.org/package=rioja. (accessed on 31 October 2022).
- Annali Delle Bonificazioni—Anno I—1858. Pianta Generale del Bacino Inferiore del Volturno con la Indicazione Delle Opere di Strade e di Canali Eseguite dal Real Governo. Available online: http://www.cesbim.it/2015/04/pianta-generale-del-bacino-inf-del-volturno-con-la-indicazione-delle-opere-di-strade-e-di-canali-eseguite-dal-real-governo-annali-delle-bonificazioni-anno-i-1858/ (accessed on 31 October 2022).
- Istituto Geografico Militare. Cartografia 1:25000; Istituto Geografico Militare: Florence, Italy, 1914. [Google Scholar]
- Satellite Imagery, Google Earth, 1990. Available online: https://earth.google.com/web/@10.75336992,34.7685858,620.74481137a,19580026.14889823d,35y,0h,0t,0r/data=Ci4SLBIgOGQ2YmFjYjU2ZDIzMTFlOThiNTM2YjMzNGRiYmRhYTAiCGxheWVyc18w (accessed on 31 October 2022).
- Ruberti, D.; Vigliotti, M.; Di Mauro, A.; Chieffi, R.; Di Natale, M. Human influence over 150 years of coastal evolution in the Volturno delta system (southern Italy). J. Coast Cons. 2018, 22, 897–917. [Google Scholar] [CrossRef]
- Ruberti, D.; Buffardi, C.; Sacchi, M.; Vigliotti, M. The late Pleistocene-Holocene changing morphology of the Volturno delta and coast (northern Campania, Italy): Geological architecture and human influence. Quat. Int. 2022, 625, 14–28. [Google Scholar] [CrossRef]
- Incarbona, A.; Ziveri, P.; Di Stefano, E.; Lirer, F.; Mortyn, G.; Patti, B.; Pelosi, N.; Sprovieri, M.; Tranchida, G.; Vallefuoco, M.; et al. Calcareous nannofossil assemblages from the Central Mediterranean Sea over the last four centuries: The impact of the Little Ice Age. Clim. Past 2010, 6, 817–866. [Google Scholar] [CrossRef] [Green Version]
- Blanc-Vernet, L. Contribution à l’étude des Foraminifères de Mèditerranée. Rec. Trav. St. Mar. Endoume 1969, 48, 5–281. [Google Scholar]
- Bergamin, L.; Di Bella, L.; Carboni, M.G. Valvulineria bradyana (Fornasini) in organic matter-enriched environment. Il Quaternario. 1999, 12, 51–56. [Google Scholar]
- Capotondi, L.; Bonomo, S.; Graiani, A.; Innangi, M.; Innangi, S.; Giglio, F.; Ravaioli, M.; Ferraro, L. Spatial Distribution of Benthic Foraminifera in the Neretva Channel (Croatia Coast): Faunal Response to Environmental Parameters. Geosciences 2022, 12, 456. [Google Scholar] [CrossRef]
- Frezza, V.; Argenti, L.; Bonifazi, A.; Chiocci, F.L.; Di Bella, L.; Ingrassia, M.; Martorelli, E. Benthic Foraminiferal Assemblages and Rhodolith Facies Evolution in Post-LGM Sediments from the Pontine Archipelago Shelf (Central Tyrrhenian Sea, Italy). Geosciences 2021, 11, 179. [Google Scholar] [CrossRef]
- Alberico, I.; Cavuoto, G.; Di Fiore, V.; Punzo, M.; Tarallo, D.; Pelosi, N.; Ferraro, L.; Marsella, E. Historical maps and satellite images as tools for shoreline variations and territorial changes assessment: The case study of Volturno Coastal Plain (Southern Italy). J. Coast. Cons. 2018, 22, 919–937. [Google Scholar] [CrossRef]
- Di Rita, F.; Lirer, F.; Margaritelli, G.; Michelangeli, F.; Magri, D. Climate and human influence on the vegetation of Tyrrhenian Italy during the last 2000 years: New insights from microcharcoal and non-pollen palynomorphs. Geogr. Fis. Din. Quaternaria. 2019, 42, 203–214. [Google Scholar] [CrossRef]
- Vallefuoco, M.; Lirer, F.; Ferraro, L.; Pelosi, N.; Capotondi, L.; Sprovieri, M.; Incarbona, A. Climatic variability and anthropogenic signatures in the Gulf of Salerno (southern-eastern Tyrrhenian Sea) during the last half millennium. R. Acc. Lincei. 2012, 23, 13–23. [Google Scholar] [CrossRef]
- Donadio, C.; Vigliotti, M.; Valente, R.; Stanislao, C.; Ivaldi, R.; Ruberti, D. Anthropic vs. natural shoreline changes along the northern Campania coast, Italy. J. Coast. Cons. 2018, 22, 939–955. [Google Scholar] [CrossRef]
- Checa, H.; Margaritelli, G.; Pena, L.D.; Frigola, J.; Cacho, I.; Rettori, R.; Lirer, F. High resolution paleo-environmental changes during the Sapropel 1 in the North Ionian Sea, central Mediterranean. Holocene 2020, 30, 1504–1515. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cavaliere, M.; Scipioni, V.; Francescangeli, F.; Ferraro, L.; Frontalini, F. Paleoenvironmental Changes in the Gulf of Gaeta (Central Tyrrhenian Sea, Italy): A Perspective from Benthic Foraminifera after Dam Construction. Water 2023, 15, 815. https://doi.org/10.3390/w15040815
Cavaliere M, Scipioni V, Francescangeli F, Ferraro L, Frontalini F. Paleoenvironmental Changes in the Gulf of Gaeta (Central Tyrrhenian Sea, Italy): A Perspective from Benthic Foraminifera after Dam Construction. Water. 2023; 15(4):815. https://doi.org/10.3390/w15040815
Chicago/Turabian StyleCavaliere, Marco, Vittoria Scipioni, Fabio Francescangeli, Luciana Ferraro, and Fabrizio Frontalini. 2023. "Paleoenvironmental Changes in the Gulf of Gaeta (Central Tyrrhenian Sea, Italy): A Perspective from Benthic Foraminifera after Dam Construction" Water 15, no. 4: 815. https://doi.org/10.3390/w15040815
APA StyleCavaliere, M., Scipioni, V., Francescangeli, F., Ferraro, L., & Frontalini, F. (2023). Paleoenvironmental Changes in the Gulf of Gaeta (Central Tyrrhenian Sea, Italy): A Perspective from Benthic Foraminifera after Dam Construction. Water, 15(4), 815. https://doi.org/10.3390/w15040815