
The Multi-Generational Effect of Seawater Acidification on Larval Development, Reproduction, Ingestion Rate, and ATPase Activity of Tigriopus japonicus Mori, 1938




Abstract
:1. Introduction
2. Materials and Methods
2.1. Maintenance of Animals and Experimental Exposure
2.2. Experimental Set Up
2.3. Larval Development and Reproduction
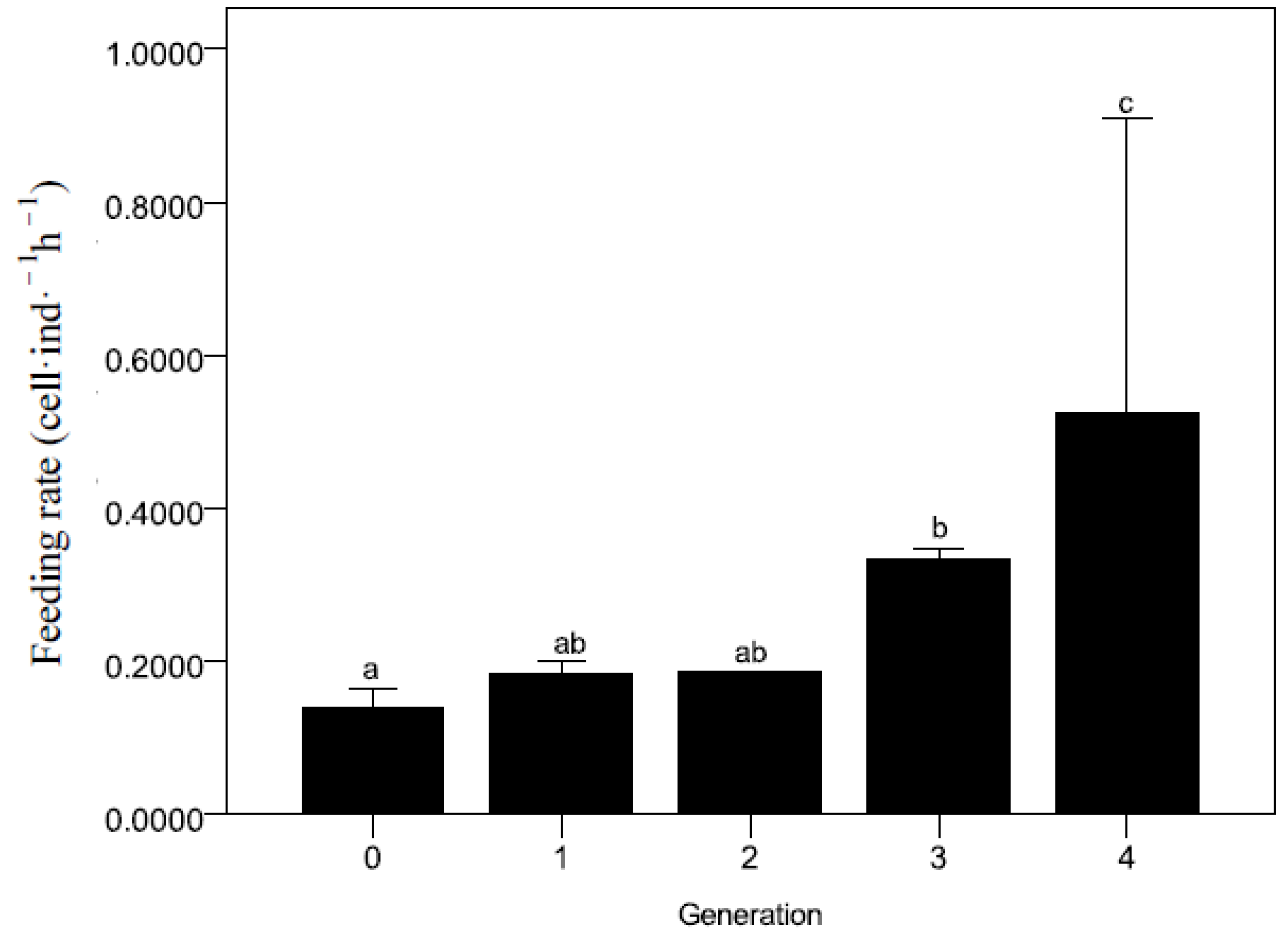
2.4. The Ingestion Rate
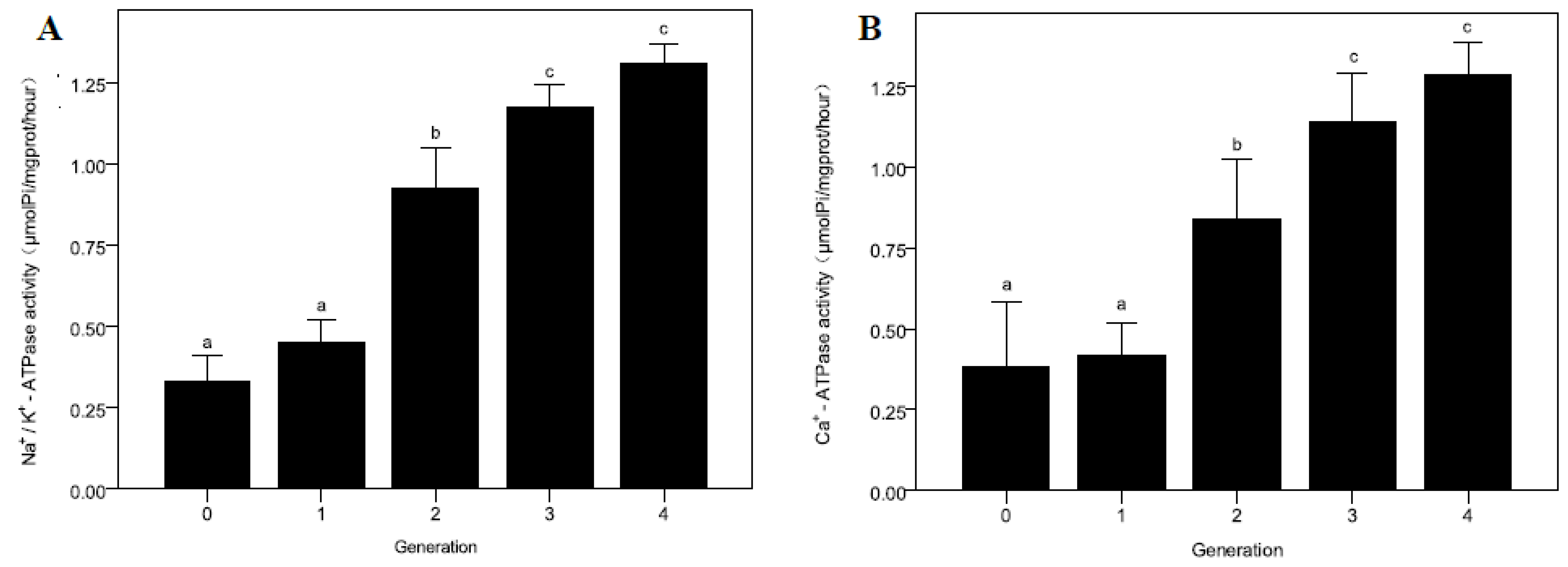
2.5. ATPase Activity
2.6. Statistical Analysis
3. Results
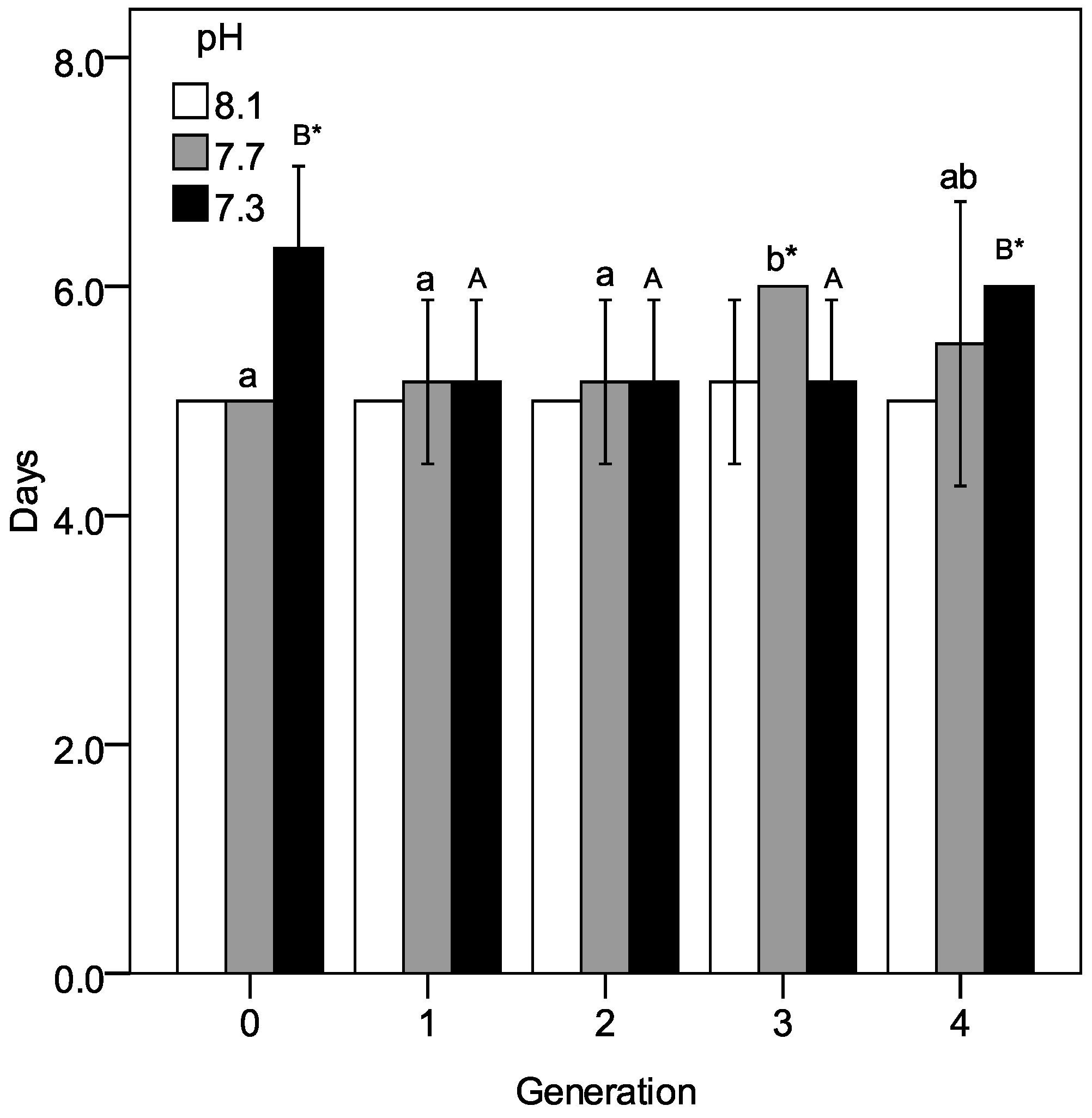
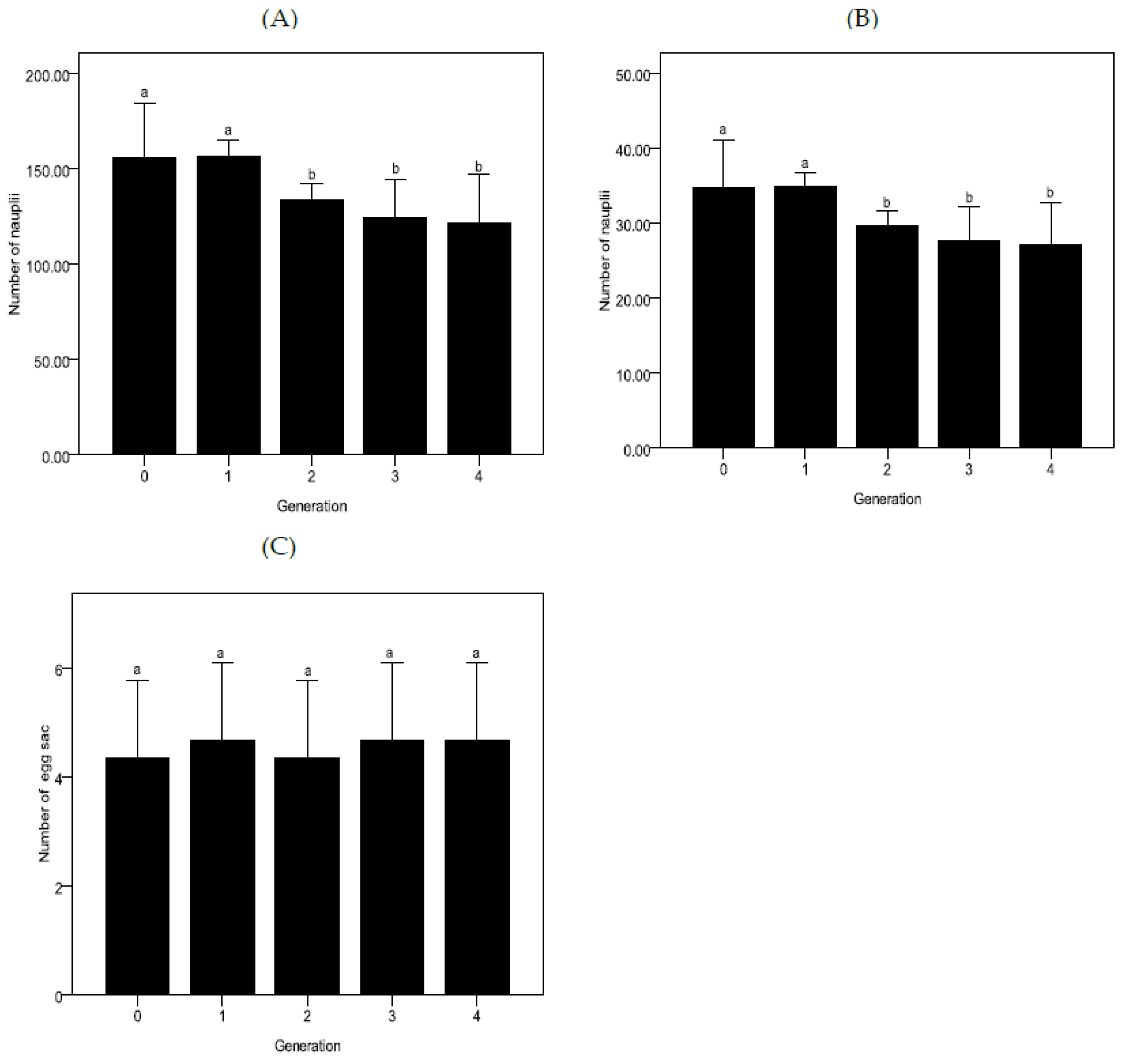
3.1. Larval Development and Production of Nauplii at Constant pH Conditions
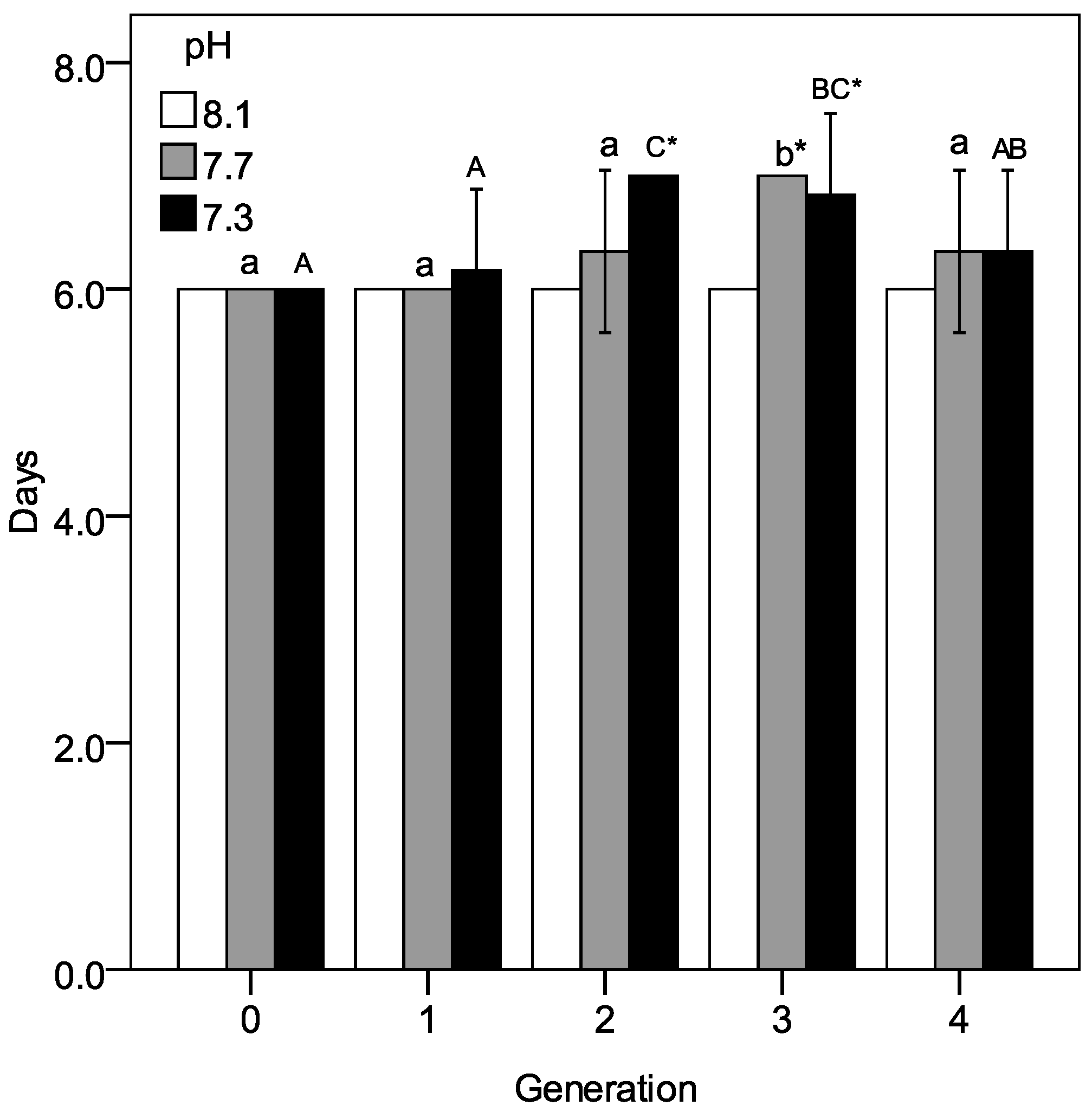
3.2. Larval Development and Production of Nauplii at Descending pH Conditions
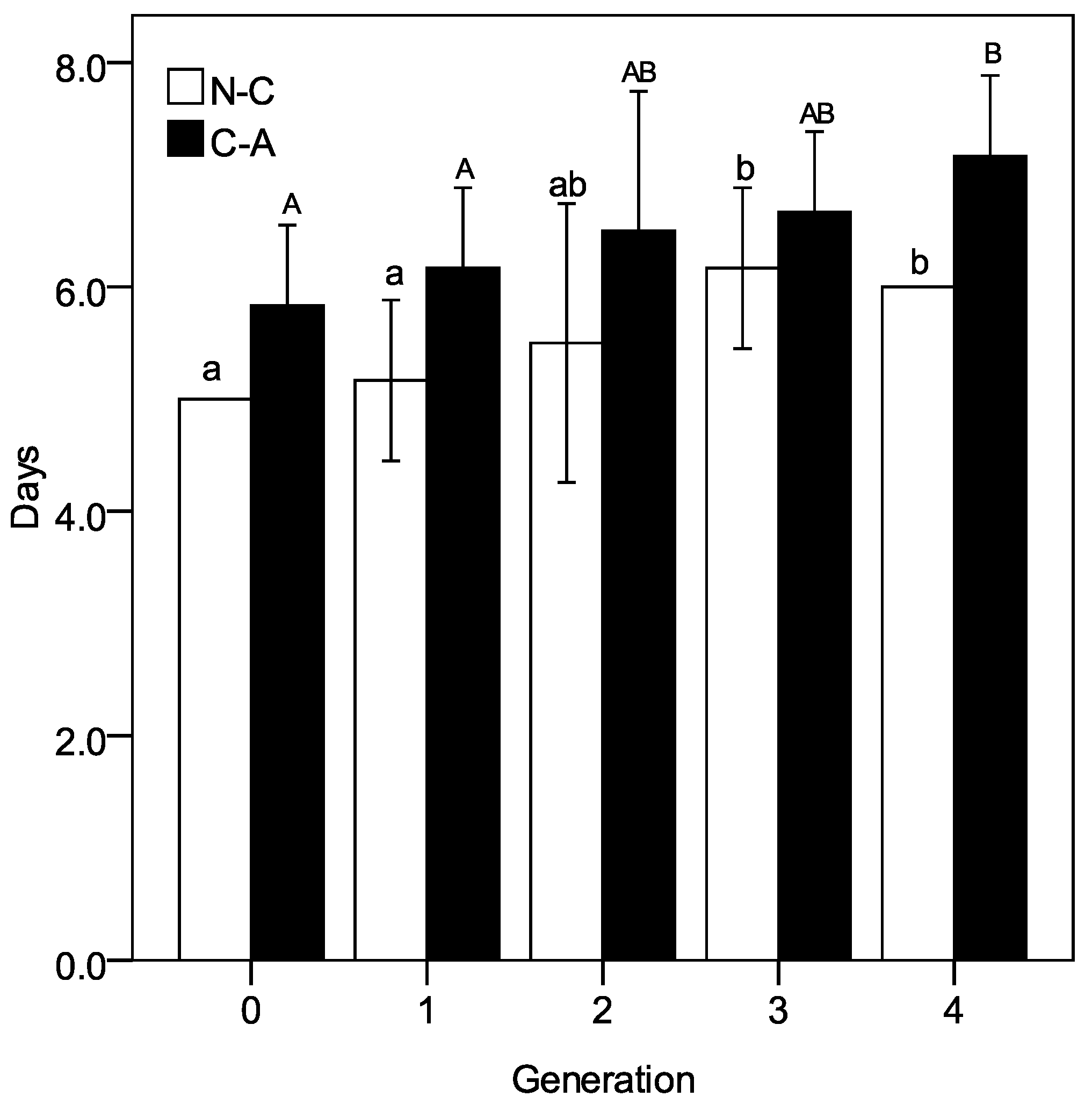
3.3. The Ingestion Rate and ATPase Activity at Descending pH Condition
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Caldeira, K.; Wickett, M.E. Oceanography: Anthropogenic carbon and ocean pH. Nature 2003, 425, 365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Church, J.; Clark, P.; Cazenave, A.; Gregory, J.; Unnikrishnan, A. The Physical Science Basis. In Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: New York, NY, USA, 2013. [Google Scholar]
- Orr, J.C.; Fabry, V.J.; Aumont, O.; Bopp, L.; Doney, S.C.; Feely, R.A.; Gnanadesikan, A.; Gruber, N.; Ishida, A.; Joos, F.; et al. Anthropogenic ocean acidification over the twenty-first century and its impact on calcifying organisms. Nature 2005, 437, 681–686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.Y.; Yin, W.L.; Zhan, Y.Y.; Jia, Y.J.; Cui, D.Y.; Zhang, W.J.; Chang, Y.Q. Comparative metabolome analysis provides new insights into increased larval mortality under seawater acidifification in the sea urchin Strongylocentrotus intermedius. Sci. Total Environ. 2020, 747, 141206. [Google Scholar] [CrossRef] [PubMed]
- Allen, S.K.; Plattner, G.K.; Nauels, A.; Xia, Y.; Stocker, T.F. Climate change 2013: The physical science basis. An overview of the working group 1 contribution to the fifth assessment report of the intergovernmental panel on climate change (ipcc). Comput. Geom. 2007, 18, 95–123. [Google Scholar]
- Feely, R.A.; Sabine, C.L.; Lee, K.; Berelson, W.; Kleypas Fabry, V.J.; Millero, F.J. Impact of anthropogenic CO2 on the CaCO3 system in the oceans. Science 2004, 305, 362–366. [Google Scholar] [CrossRef] [Green Version]
- Gargan, P.; Karlsbakk, E.; Coyne, J.; Davies, C.; Roche, W. Sea lice (Lepeophtheirus salmonis and Caligus elongatus) infestation levels on sea trout (Salmo trutta L.) around the Irish Sea, an area without salmon aquaculture. ICES J. Mar. Sci. 2016, 9, 2395–2407. [Google Scholar] [CrossRef] [Green Version]
- Kurihara, H.; Ishimatsu, A. Effects of high CO2 seawater on the copepod (Acartia tsuensis) through all life stages and subsequent generations. Mar. Pollut. Bull. 2008, 56, 1086–1090. [Google Scholar] [CrossRef] [Green Version]
- Mayor, D.J.; Everett, N.R.; Cook, K.B. End of century ocean warming and acidification effects on reproductive success in a temperate marine copepod. J. Plankton Res. 2012, 34, 258–262. [Google Scholar] [CrossRef]
- Kurihara, H.; Shimode, S.; Shirayama, Y. Effects of raised CO2 concentration on the egg production rate and early development of two marine copepods (Acartia steueri and Acartia erythraea). Mar. Pollut. Bull. 2004, 49, 721–727. [Google Scholar] [CrossRef]
- Mayor, D.J.; Matthews, C.; Cook, K.; Zuur, A.; Hay, S. CO2-induced acidification affects hatching success in Calanus finmarchicus. Mar. Ecol. Prog. Ser. 2007, 350, 91–97. [Google Scholar] [CrossRef] [Green Version]
- Pascal, P.; Fleeger, J.W.; Galvez, F.; Carman, K.R. The toxicological interaction between ocean acidity and metals in coastal meiobenthic copepods. Mar. Pollut. Bull. 2010, 60, 2201–2208. [Google Scholar] [CrossRef]
- Weydmann, A.; Søreide, J.E.; Kwasniewski, S.; Widdicombe, S. Influence of CO2-induced acidification on the reproduction of a key Arctic copepod Calanus glacialis. J. Exp. Mar. Biol. Ecol. 2012, 428, 39–42. [Google Scholar] [CrossRef]
- Lee, Y.H.; Kang, H.M.; Kim, M.S.; Wang, M.; Kim, J.H.; Jeong, C.B.; Lee, J.S. Effects of ocean acidification on life parameters and antioxidant system in the marine copepod Tigriopus japonicus. Aquat. Toxicol. 2019, 212, 186–193. [Google Scholar] [CrossRef]
- Borges, F.O.; Figueiredo, C.; Sampaio, E.; Rosa, R.; Grilo, T.F. Transgenerational deleterious effects of ocean acidification on the reproductive success of a keystone crustacean (Gammarus locusta). Mar. Environ. Res. 2018, 138, 55–64. [Google Scholar] [CrossRef]
- Pierre, D.W.; Sam, D.; Peter, T. Selection on oxidative phosphorylation and ribosomal structure as a multigenerational response to ocean acidification in the common copepod pseudocalanus acuspes. Evol. Appl. 2016, 23, 1112–1123. [Google Scholar]
- Thor, P.; Dupont, S. Transgenerational effects alleviate severe fecundity loss during ocean acidification in a ubiquitous planktonic copepod. Glob. Chang. Biol. 2015, 21, 2261–2271. [Google Scholar] [CrossRef] [Green Version]
- Doney, S.C.; Ruckelshaus, M.; Duffy, J.E.; Barry, J.P.; Chan, F.; English, C.A.; Galindo, H.M.; Grebmeier, J.M.; Hollowed, A.B.; Knowlton, N.; et al. Climate change impacts on marine ecosystems. Annu. Rev. Mar. Sci. 2012, 4, 11–37. [Google Scholar] [CrossRef] [Green Version]
- Munday, P.L.; Warner, R.R.; Monro, K.; Pandolfi, J.M.; Marshall, D.J.; Wootton, T. Predicting evolutionary responses to climate change in the sea. Ecol. Lett. 2013, 16, 1488–1500. [Google Scholar] [CrossRef]
- Glippa, O.; Jonna, E.; Kanerva, M.; Rein, A.; Vuori, k. Oxidative stress and antioxidant defense responses in acartia copepods in relation to environmental factors. PLoS One 2018, 13, e0195981. [Google Scholar] [CrossRef] [Green Version]
- Langer, J.A.F.; Meunier, C.L.; Ecker, U.; Horn, H.G.; Schwenk, K.; Boersma, M. Acclimation and adaptation of the coastal calanoid copepod Acartia tonsa to ocean acidifification: A long-term laboratory investigation. Mar. Ecol. Prog. Ser. 2019, 619, 35–51. [Google Scholar] [CrossRef]
- Lee, E.H.; Choi, S.Y.; Seo, M.H.; Lee, S.J.; Soh, H.Y. Effects of Temperature and pH on the Egg Production and Hatching Success of a Common Korean Copepod. Diversity 2020, 12, 372. [Google Scholar] [CrossRef]
- Dam, H.G.; Demayo, J.A.; Park, G.; Norton, L.; He, X.; Finiguerra, M.B.; Baumann, H.; Brennan, R.S.; Pespeni, M.H. Rapid, but limited, zooplankton adaptation to simultaneous warming and acidification. Nat. Clim. Chang. 2021, 11, 780–786. [Google Scholar] [CrossRef]
- Prazeres, M.; Uthicke, S.; Pandolfi, J.M. Ocean acidification induces biochemical and morphological changes in the calcification process of large benthic foraminifera. Proc. Biol. Sci. 2015, 282, 20142782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pörtner, H.O.; Langenbuch, M.; Reipschläger, A. Biological impact of elevated ocean CO2 concentrations: Lessons from animal physiology and earth history. J. Oceanogr. 2004, 60, 705–718. [Google Scholar] [CrossRef] [Green Version]
- Widdicombe, S.; Spicer, J.I. Predicting the impact of ocean acidification on benthic biodiversity: What can animal physiology tell us? J. Exp. Mar. Biol. Ecol. 2008, 366, 187–197. [Google Scholar] [CrossRef]
- Rasmussen, H.; Barrett, P.; Smallwood, J.; Bollag, W.; Isales, C. Calcium ion as intracellular messenger and cellular toxin. Environ. Health Perspec. 1990, 84, 17–25. [Google Scholar] [CrossRef]
- Parker, L.M.; Ross, P.M.; O’Connor, W.A.; Pörtner, H.O.; Scanes, E.; Wright, J.M. Predicting the response of molluscs to the impact of ocean acidification. Biology 2013, 2, 651–692. [Google Scholar] [CrossRef] [Green Version]
- Whiteley, N.M. Physiological and ecological responses of crustaceans to ocean acidification. Mar. Ecol. Prog. Ser. 2011, 430, 257–271. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Shi, J.; Cheung, S.G.; Shin, P.K.S.; Liu, X.; Sun, Y.; Mu, F.H. The combined effects of elevated pCO2, and food availability on Tigriopus japonicus, Mori larval development, reproduction, and superoxide dismutase activity. Mar. Pollut. Bull. 2018, 126, 623–628. [Google Scholar] [CrossRef]
- Raisuddin, S.; Kwok, K.W.; Leung, K.M.; Schlenk, D.; Lee, J.S. The copepod Tigriopus: A promising marine model organism for ecotoxicology and environmental genomics. Aquat. Toxicol. 2007, 83, 161–173. [Google Scholar] [CrossRef]
- Itô, T. The biology of a harpacticoid copepod, Tigriopus japonicus Mori (With 12 Text-figures, 1 Table and 2 Plates). J. Fac. Sci. Hokkaido Univ. 1970, 17, 474–500. [Google Scholar]
- Cao, Z.Q.; Mu, F.H.; Wei, X.H.; Sun, Y.T. Influence of CO2-induced seawater acidification on the development and lifetime reproduction of Tigriopus japonicus mori. J. Nat. Hist. 2015, 49, 2813–2826. [Google Scholar] [CrossRef]
- Sun, Y.T.; Mu, F.H.; Wei, X.H.; Cao, Z.Q.; Fu, S.S. Effect of seawater acidification on ingestion rates of Tisbe Sp. And Tigrification japonicus. Trans. Oceanol. Limnol. 2014, 03, 105–111. [Google Scholar]
- Gao, X.; Mu, C.; Li, Q. Effects of toxic dinoflflagellate Alexandrium catenella on sexual maturation and reproductive output in the pacifific oyster Crassostrea gigas. Aquat. Toxicol. 2021, 232, 105745. [Google Scholar] [CrossRef]
- Cripps, G.; Lindeque, P.; Flynn, K.J. Have we been underestimating the effects of ocean acidification in zooplankton? Glob. Chang. Biol. 2014, 20, 3377–3385. [Google Scholar] [CrossRef] [Green Version]
- Daniel, M.J.; Boyden, C.R. Diurnal variations in physico-chemical conditions within intertidal rock pools. Field Stud. 1975, 4, 161–176. [Google Scholar]
- Dupont, S.; Dorey, N.; Stumpp, M.; Melzner, F.; Thorndyke, M. Long-term and trans-life-cycle effects of exposure to ocean acidification in the green sea urchin Strongylocentrotus droebachiensis. Mar. Biol. 2013, 160, 1835–1843. [Google Scholar] [CrossRef] [Green Version]
- Pedersen, S.A.; Håkedal, O.J.; Salaberria, I.; Tagliati, A.; Gustavson, L.M.; Jenssen, B.M.; Olsen, A.J.; Altin, D. Multigenerational exposure to ocean acidification during food limitation reveals consequences for copepod scope for growth and vital rates. Int. J. Environ. Sci. Technol. 2014, 48, 12275–12284. [Google Scholar] [CrossRef] [Green Version]
- Griffith, A.W.; Gobler, C. Multigenerational Effects of Acidification on Early Life-Stage Mercenaria mercenaria (=hard clam). Am. Geophys. Union 2016, 2016, AH54A-0105. [Google Scholar]
- Kurihara, H.; Shirayama, Y. Seawater carbonate chemistry and processes during experiments with seaurchins Hemicentrotus pulcherrimus and Echinometra mathaei. Pangaea 2004. supplement to Kurihara, H.; Shirayama, Y. Effects of increased atmospheric CO2 on sea urchin early developme. Mar. Ecol. Prog. Ser. 2004, 274, 161–196. [Google Scholar] [CrossRef]
- Melzner, F.S.; Göbel, S.; Langenbuch, M.; Gutowska, M.A.; Pörtner, H.O.; Lucassen, M. Swimming performance in Atlantic Cod (Gadus morhua) following long-term (4–12 months) acclimation to elevated seawater pCO2. Aquat. Toxicol. 2009, 92, 30–37. [Google Scholar] [CrossRef] [PubMed]
- James, N.; Cameron, C.M.; Wood. Apparent H+ excretion and CO2 dynamics accompanying carapace mineralization in the blue crab (Callinectes sapidus) following moulting. J. Exp. Biol. 1985, 114, 181–196. [Google Scholar]
- Findlay, H.S.; Kendall, M.A.; Spicer, J.I.; Widdicombe, S. Future high CO2 in the intertidal may compromise adult barnacle Semibalanus balanoides survival and embryonic development rate. Mar. Ecol. Prog. Ser. 2009, 389, 193–202. [Google Scholar] [CrossRef] [Green Version]
- Ries, J.B.; Cohen, A.L.; McCorkleD, C. Marine calcifiers exhibit mixed responses to CO2-induced ocean acidification. Geology 2009, 37, 1131–1134. [Google Scholar] [CrossRef]
- Luquet, G. Biomineralizations: Insights and prospects from crustaceans. Zookeys 2012, 176, 103–121. [Google Scholar] [CrossRef] [Green Version]
- Luquet, G.; Marin, F. Biomineralisations in crustaceans: Storage strategies. Comptes Rendus Palevol 2004, 3, 515–534. [Google Scholar] [CrossRef]
- Li, W.; Gao, K.A. Marine secondary producer respires and feeds more in a high CO2 ocean. Mar. Pollut. Bull. 2012, 64, 699–703. [Google Scholar] [CrossRef]
- Nybakken, J.W.; Bertness, M.D. Marine Biology: An Ecological Approach, 6th ed.; Pearson Education, Inc.: San Francisco, CA, USA, 2004. [Google Scholar]
- Brennan, R.S.; Demayo, J.A.; Dam, H.G.; Finiguerra, M.B.; Baumann, H.; Pespeni, M.H. Loss of transcriptional plasticity but sustained adaptive capacity after adaptation to global change conditions in a marine copepod. Nat. Commun. 2022, 13, 1147. [Google Scholar] [CrossRef]
- Dupont, S.; Thorndyke, M. Ocean acidification and its impact on the early life-history stages of marine animals. In Impact of Acidification on Biological, Chemical and Physical Systems in the Mediterranean and Black Seas; CIESM Publisher: Monaco, 2008; Volume 36, pp. 89–97. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments with Constant pH | ||||
|---|---|---|---|---|
| pH 8.1 | pH 7.7 | pH 7.3 | ||
| pH | before | 8.09 (8.07–8.10) | 7.70 (7.69–7.72) | 7.31 (7.30–7.33) |
| after | 8.12 (8.10–8.14) | 7.72 (7.71–7.74) | 7.32 (7.30–7.34) | |
| S | before | 29 (±0.3) | 29 (±0.4) | 29 (±0.4) |
| after | 30 (±0.5) | 30 (±0.3) | 30 (±0.5) | |
| DO (mg/L) | before | 6.2 (±0.4) | 5.9 (±0.2) | 5.6 (±0.3) |
| after | 5.9 (±0.8) | 5.7 (±0.6) | 5.3 (±0.6) | |
| TA (mg/L) | before | 141 (±1.7) | 136 (±1.4) | 127 (±1.5) |
| after | 137 (±1.3) | 132 (±1.3) | 125 (±1.3) | |
| pCO2 (μatm) | before | 413 (±8) | 997 (±14) | 2555 (±40) |
| after | 391 (±9) | 942 (±20) | 2515 (±43) | |
| CT(mM/kg) | before | 1933 (±42) | 2026 (±34) | 2159 (±38) |
| after | 1917 (±39) | 1989 (±30) | 2126 (±30) | |
| ΩCa | before | 2.45 (±0.03) | 1.65 (±0.02) | 0.58 (±0.01) |
| after | 2.41 (±0.04) | 1.59 (±0.05) | 0.53 (±0.01) | |
| Treatments with Descending pH | ||||||
|---|---|---|---|---|---|---|
| pH 8.1 | pH 7.9 | pH 7.7 | pH 7.5 | pH 7.3 | ||
| S | before | 8.10 (8.09–8.11) | 7.91 (7.90 ± 7.93) | 7.69 (7.69 ± 7.71) | 7.50 (7.49 ± 7.52) | 7.30 (7.29 ± 7.33) |
| after | 8.13 (8.11–8.15) | 7.92 (7.91 ± 7.94) | 7.71 (7.7 ± 7.74) | 7.51 (7.51 ± 7.54) | 7.31 (7.31 ± 7.35) | |
| DO(mg/L) | before | 29 (±0.5) | 29 (±0.4) | 29 (±0.3) | 29 (±0.5) | 29 (±0.6) |
| after | 30 (±0.7) | 30 (±0.5) | 30 (±0.4) | 30 (±0.6) | 30 (±0.6) | |
| TA (mg/L) | before | 6.1 (±0.4) | 6.0 (±0.4) | 5.8 (±0.3) | 5.8 (±0.4) | 5.5 (±0.3) |
| after | 5.8 (±0.9) | 5.9 (±0.9) | 5.7 (±0.8) | 5.7 (±0.9) | 5.3 (±0.5) | |
| pCO2 (μatm) | before | 143 (±1.9) | 140 (±1.8) | 135 (±1.5) | 133 (±1.4) | 125 (±1.6) |
| after | 139 (±1.1) | 138 (±1.2) | 131 (±1.3) | 130 (±1.5) | 124 (±1.5) | |
| CT (mM/kg) | before | 403 (±5) | 588 (±8) | 987 (±15) | 1537 (±17) | 2556 (±40) |
| after | 387 (±9) | 562 (±9) | 939 (±21) | 1466 (±20) | 2510 (±45) | |
| ΩCa | before | 1930 (±44) | 2002 (±24) | 2026 (±34) | 2096 (±22) | 2159 (±38) |
| after | 1910 (±40) | 1995 (±20) | 1998 (±35) | 2045 (±24) | 2120 (±21) | |
| pH | before | 2.45 (±0.03) | 2.17 (±0.02) | 1.65 (±0.02) | 0.89 (±0.01) | 0.58 (±0.01) |
| after | 2.41 (±0.04) | 2.15 (±0.02) | 1.59 (±0.05) | 0.88 (±0.02) | 0.53 (±0.01) | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, F.; Cheung, S.G.; Shin, P.K.S.; Liu, X.; Li, Y.; Mu, F. The Multi-Generational Effect of Seawater Acidification on Larval Development, Reproduction, Ingestion Rate, and ATPase Activity of Tigriopus japonicus Mori, 1938. Water 2023, 15, 816. https://doi.org/10.3390/w15040816
Li F, Cheung SG, Shin PKS, Liu X, Li Y, Mu F. The Multi-Generational Effect of Seawater Acidification on Larval Development, Reproduction, Ingestion Rate, and ATPase Activity of Tigriopus japonicus Mori, 1938. Water. 2023; 15(4):816. https://doi.org/10.3390/w15040816
Chicago/Turabian StyleLi, Fei, S. G. Cheung, P. K. S. Shin, Xiaoshou Liu, Yanan Li, and Fanghong Mu. 2023. "The Multi-Generational Effect of Seawater Acidification on Larval Development, Reproduction, Ingestion Rate, and ATPase Activity of Tigriopus japonicus Mori, 1938" Water 15, no. 4: 816. https://doi.org/10.3390/w15040816
APA StyleLi, F., Cheung, S. G., Shin, P. K. S., Liu, X., Li, Y., & Mu, F. (2023). The Multi-Generational Effect of Seawater Acidification on Larval Development, Reproduction, Ingestion Rate, and ATPase Activity of Tigriopus japonicus Mori, 1938. Water, 15(4), 816. https://doi.org/10.3390/w15040816