
Chironomid-Based Modern Summer Temperature Data Set and Inference Model for the Northwest European Part of Russia

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
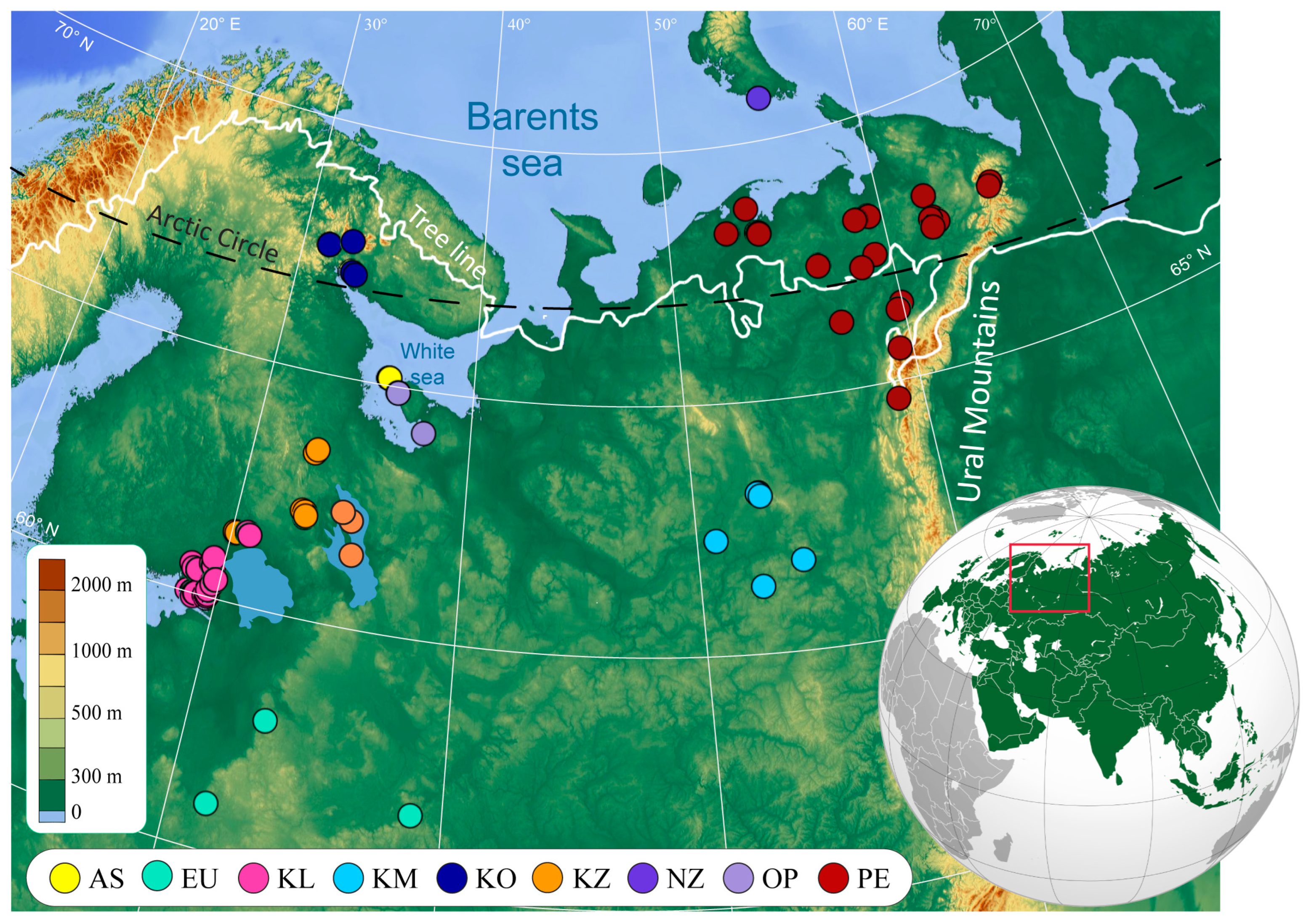
2.1. Field Methods and Derivation of Climate Variables
2.2. Chironomid Analysis
2.3. Numerical Methods
2.4. Model Development
3. Results
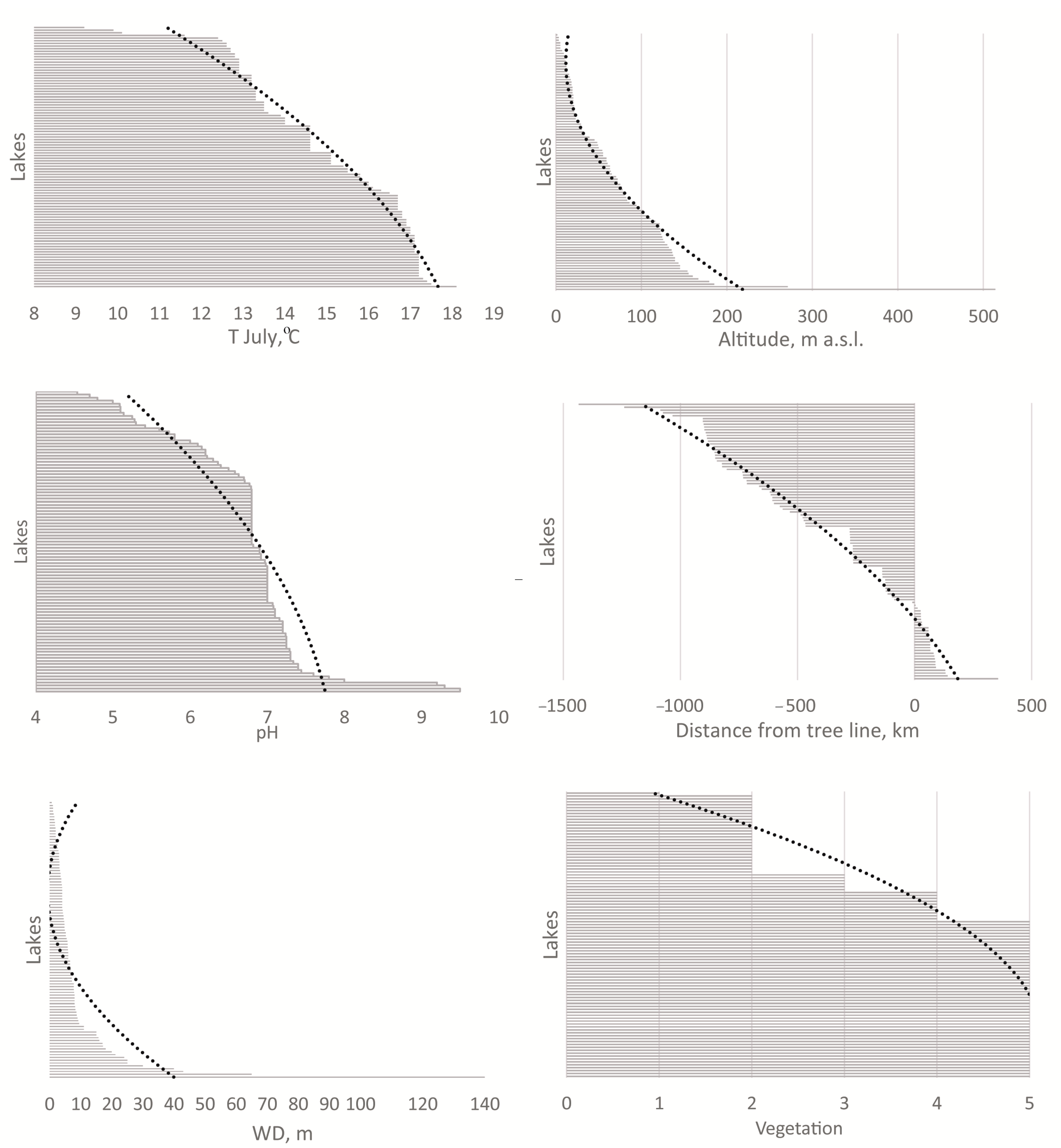
3.1. Environmental Parameters of the Lakes
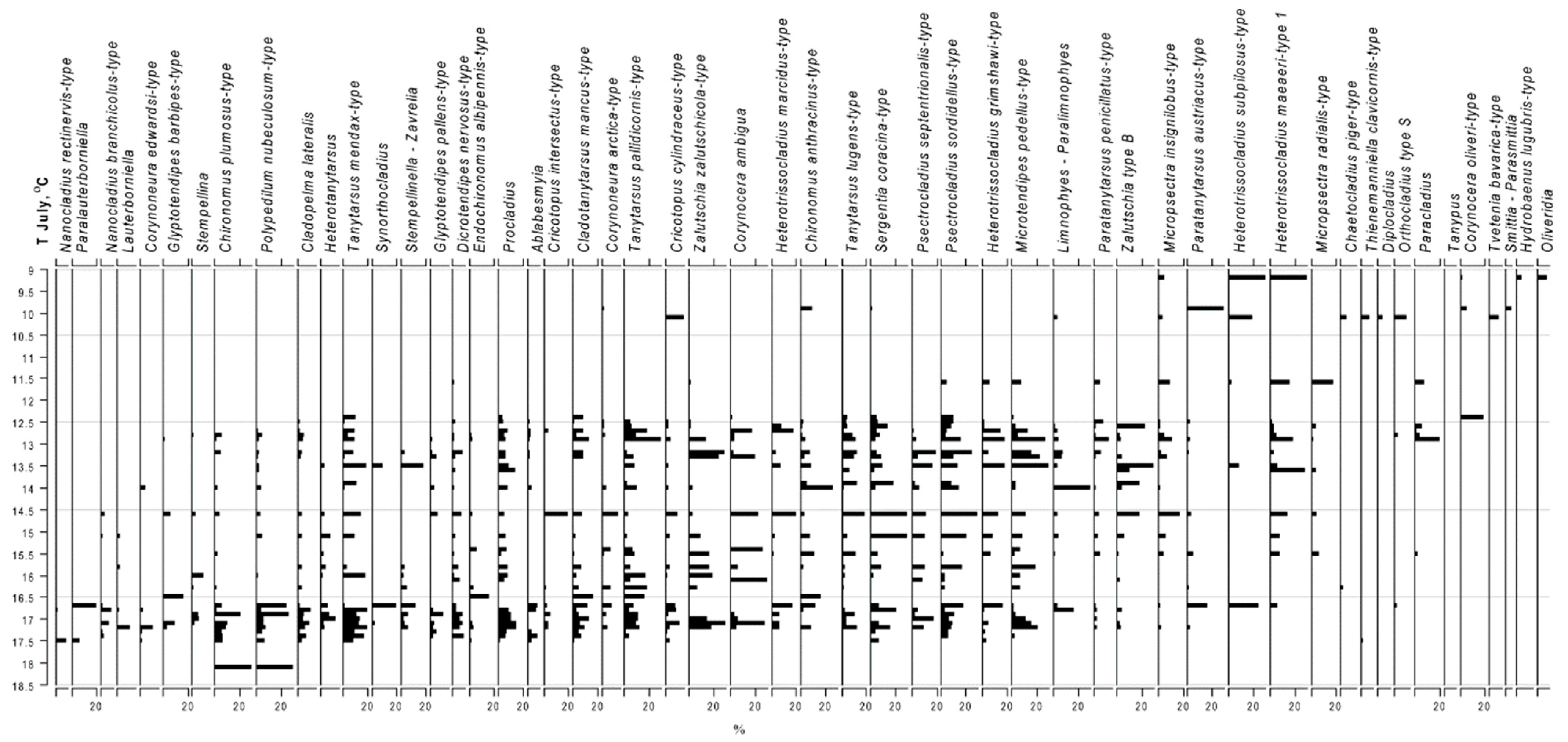
3.2. Chironomid Fauna
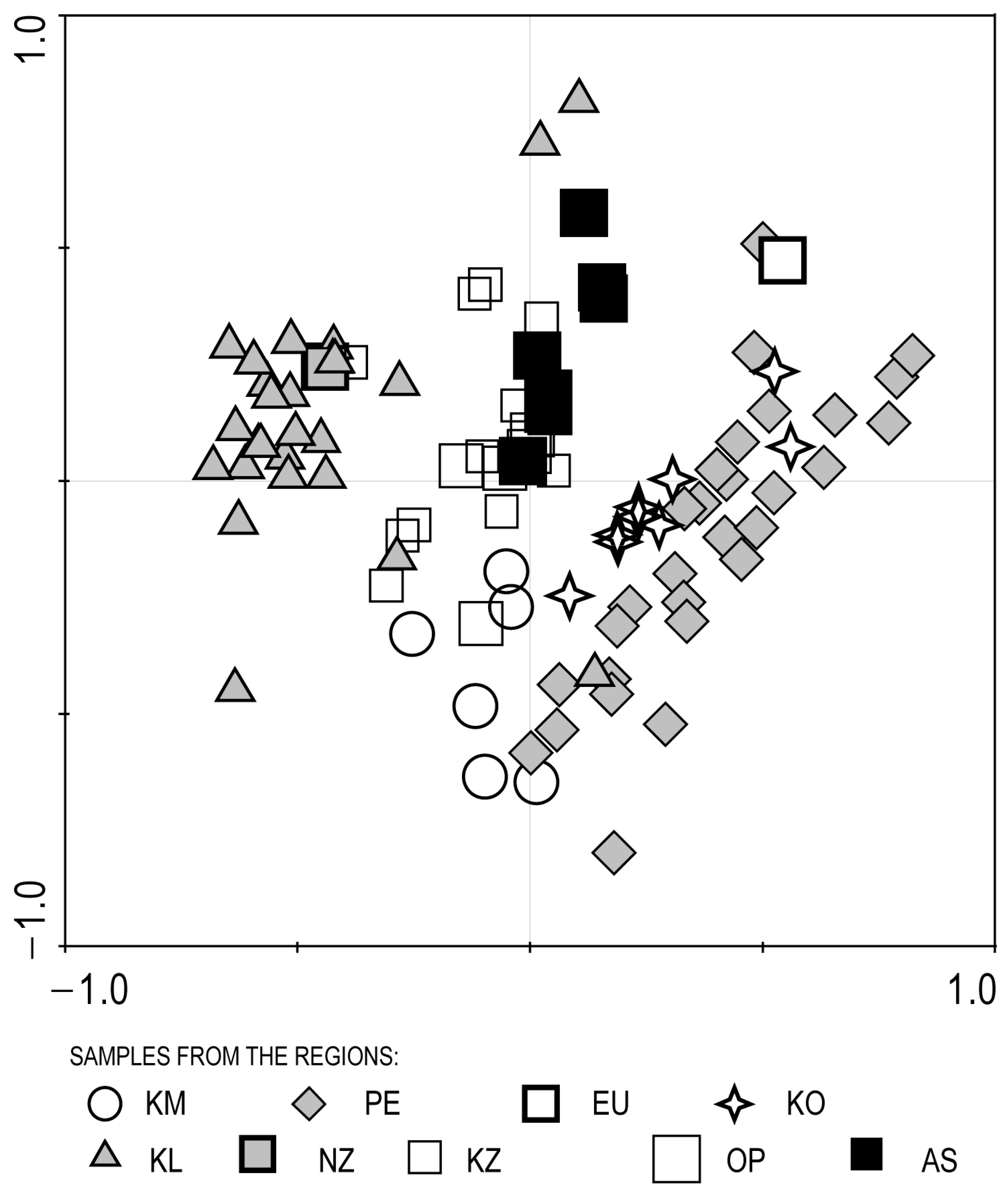
3.3. Ordination of the Set of Data
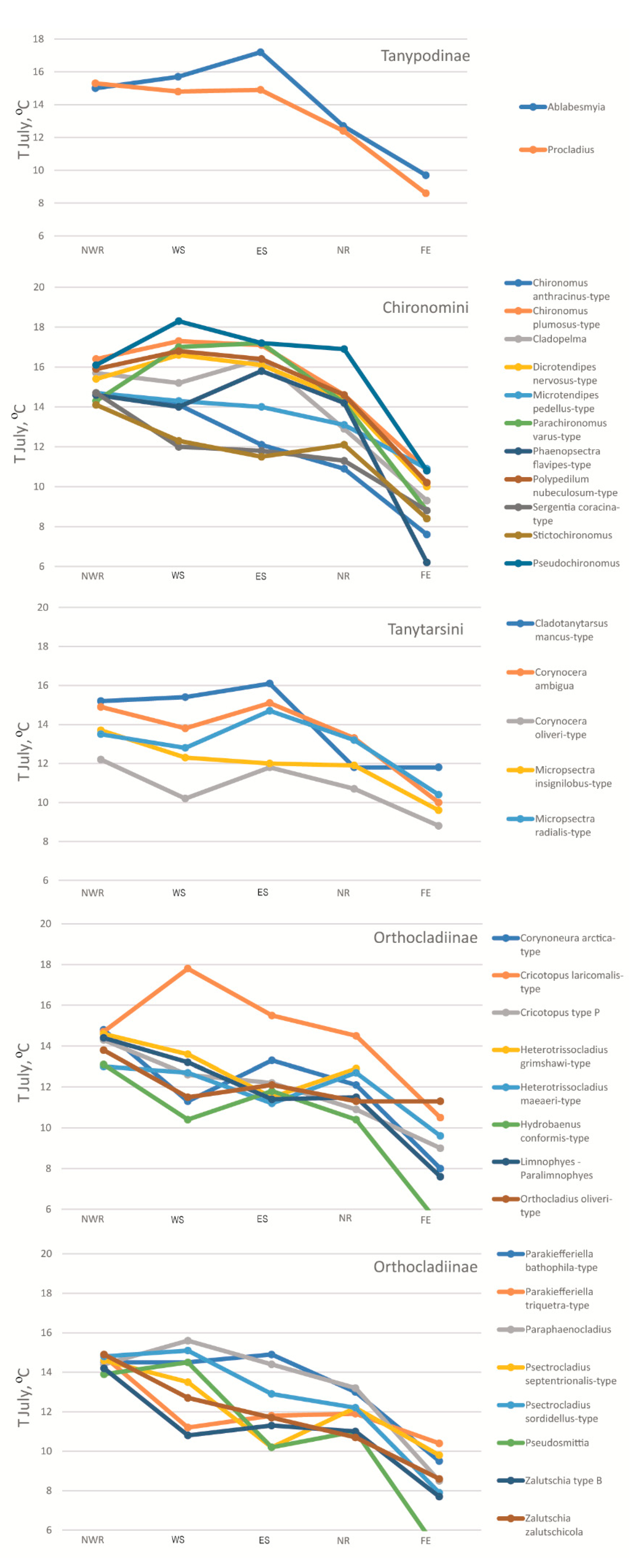
3.4. Development of Inference Models and Taxon-Specific T July Optima
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Coffman, W.; Ferrington, L. Chironomidae; Merritt, R., Cummins, K., Eds.; Kendall/Hunt Publishing Company: Dubuque, IA, USA, 1996. [Google Scholar]
- Stief, P.; Nazarova, L.; de Beer, D. Chimney construction by Chironomus riparius larvae in response to hypoxia: Microbial implications for freshwater sediments. J. N. Am. Benthol. Soc. 2005, 24, 858–871. [Google Scholar] [CrossRef]
- Hölker, F.; Vanni, M.J.; Kuiper, J.J.; Meile, C.; Grossart, H.-P.; Stief, P.; Adrian, R.; Lorke, A.; Dellwig, O.; Brand, A.; et al. Tube-dwelling invertebrates: Tiny ecosystem engineers have large effects in lake ecosystems. Ecol. Monogr. 2015, 85, 333–351. [Google Scholar] [CrossRef] [Green Version]
- Engels, S.; Medeiros, A.S.; Axford, Y.; Brooks, S.J.; Heiri, O.; Luoto, T.P.; Nazarova, L.; Porinchu, D.F.; Quinlan, R.; Self, A.E. Temperature change as a driver of spatial patterns and long-term trends in chironomid (Insecta: Diptera) diversity. Glob. Chang. Biol. 2020, 26, 1155–1169. [Google Scholar] [CrossRef] [PubMed]
- Brooks, S.J. Fossil midges (Diptera: Chironomidae) as palaeoclimatic indicators for the Eurasian region. Quat. Sci. Rev. 2006, 25, 1894–1910. [Google Scholar] [CrossRef]
- Medeiros, A.S.; Quinlan, R.; Sprules, G. The distribution of the Chironomidae (Insecta: Diptera) along multiple environmental gradients in lakes and ponds of the eastern Canadian Arctic. Can. J. Fish. Aquat. Sci. 2011, 68, 1511–1527. [Google Scholar] [CrossRef]
- Eggermont, H.; Heiri, O. The chironomid-temperature relationship: Expression in nature and palaeoenvironmental implications. Biol. Rev. Camb. Philos. Soc. 2012, 87, 430–456. [Google Scholar] [CrossRef] [PubMed]
- Fortin, M.-C.; Medeiros, A.S.; Gajewski, K.; Barley, E.M.; Larocque-Tobler, I.; Porinchu, D.F.; Wilson, S.E. Chironomid-environment relations in northern North America. J. Paleolimnol. 2015, 54, 223–237. [Google Scholar] [CrossRef]
- Barley, E.M.; Walker, I.R.; Kurek, J.; Cwynar, L.C.; Mathewes, R.W.; Gajewski, K.; Finney, B.P. A northwest North American training set: Distribution of freshwater midges in relation to air temperature and lake depth. J. Paleolimnol. 2006, 36, 295–314. [Google Scholar] [CrossRef]
- Dieffenbacher-Krall, A.; Vandergoes, M.J.; Denton, G.H. An inference model for mean summer air temperatures in the Southern Alps, New Zealand, using subfossil chironomids. Quat. Sci. Rev. 2007, 26, 2487–2504. [Google Scholar] [CrossRef]
- Rees, A.B.H.; Cwynar, L.C.; Cranston, P. Midges (Chironomidae, Ceratopogonidae, Choaboridae) as a temperature proxy: A training set from Tasmania, Australia. J. Paleolimnol. 2008, 40, 1159–1178. [Google Scholar] [CrossRef]
- Larocque, I.; Hall, R.; Grahn, E. Chironomids as indicators of climate change: A 100-lake training set from a subarctic region of northern Sweden (Lapland). J. Paleolimnol. 2001, 26, 307–322. [Google Scholar] [CrossRef]
- Heiri, O.; Lotter, A.F.; Hausmann, S.; Kienast, F. A chironomid-based Holocene summer air temperature reconstruction from the Swiss Alps. Holocene 2003, 13, 477–484. [Google Scholar] [CrossRef] [Green Version]
- Luoto, T.P. Subfossil Chironomidae (Insecta: Diptera) along a latitudinal gradient in Finland: Development of a new temperature inference model. J. Quat. Sci. 2009, 24, 150–158. [Google Scholar] [CrossRef]
- Luoto, T.; Kotrys, B.; Płóciennik, M. East European chironomid-based calibration model for past summer temperature reconstructions. Clim. Res. 2018, 77, 63–76. [Google Scholar] [CrossRef] [Green Version]
- Self, A.; Brooks, S.; Birks, H.; Nazarova, L.; Porinchu, D.; Odland, A.; Yang, H.; Jones, V. The distribution and abundance of chironomids in high-latitude Eurasian lakes with respect to temperature and continentality: Development and application of new chironomid-based climate-inference models in northern Russia. Quat. Sci. Rev. 2011, 30, 1122–1141. [Google Scholar] [CrossRef] [Green Version]
- Nazarova, L.B.; Pestryakova, L.A.; Ushnitskaya, L.A.; Hubberten, H.-W. Chironomids (Diptera: Chironomidae) in lakes of central Yakutia and their indicative potential for paleoclimatic research. Contemp. Probl. Ecol. 2008, 1, 335–345. [Google Scholar] [CrossRef] [Green Version]
- Nazarova, L.; Herzschuh, U.; Wetterich, S.; Kumke, T.; Pestryakova, L. Chironomid-based inference models for estimating mean July air temperature and water depth from lakes in Yakutia, northeastern Russia. J. Paleolimnol. 2011, 45, 57–71. [Google Scholar] [CrossRef]
- Nazarova, L.; Self, A.E.; Brooks, S.J.; van Hardenbroek, M.; Herzschuh, U.; Diekmann, B. Northern Russian chironomid-based modern summer temperature data set and inference models. Glob. Planet. Chang. 2015, 134, 10–25. [Google Scholar] [CrossRef] [Green Version]
- Nazarova, L.B.; Sapelko, T.V.; Kuznetsov, D.D.; Syrykh, L.S. Palaeoecological and Palaeoclimatical Reconstructions of Holocene According Chironomid Analysis of Lake Glubokoye Sediments. Dokl. Biol. Sci. 2015, 460, 57–60. [Google Scholar] [CrossRef] [Green Version]
- Subetto, D.A.; Nazarova, L.B.; Pestryakova, L.A.; Syrykh, L.S.; Andronikov, A.V.; Biskaborn, B.; Diekmann, B.; Kuznetsov, D.D.; Sapelko, T.V.; Grekov, I.M. Paleolimnological studies in Russian northern Eurasia: A review. Contemp. Probl. Ecol. 2017, 4, 327–335. [Google Scholar] [CrossRef]
- Syrykh, L.; Subetto, D.; Nazarova, L. Paleolimnological studies on the East European Plain and nearby regions: The PaleoLake Database. J. Paleolimnol. 2021, 65, 369–375. [Google Scholar] [CrossRef]
- Solovieva, N.; Jones, V.J.; Nazarova, L.; Brooks, S.J.; Birks, H.J.B.; Grytnes, J.-A.; Appleby, P.G.; Kauppila, T.; Kondratenok, B.; Renberg, I.; et al. Paleolimnological Evidence for Recent Climate Change in Lakes from the Northern Urals, Russia. J. Paleolimnol. 2005, 33, 463–482. [Google Scholar] [CrossRef] [Green Version]
- Nazarova, L.B.; Self, A.E.; Brooks, S.J.; Solovieva, N.; Syrykh, L.S.; Dauvalter, V.A. Chironomid fauna of the lakes from the Pechora river basin (east of European part of Russian Arctic): Ecology and reconstruction of recent ecological changes in the region. Contemp. Probl. Ecol. 2017, 4, 350–362. [Google Scholar] [CrossRef] [Green Version]
- Nazarova, L.B.; Frolova, L.A.; Palagushkina, O.V.; Rudaya, N.A.; Syrykh, L.S.; Grekov, I.M.; Solovieva, N.; Loskutova, O.A. Recent shift in biological communities: A case study from the Eastern European Russian Arctic (Bol`shezemelskaya Tundra). Polar Biol. 2021, 44, 1107–1125. [Google Scholar] [CrossRef]
- Andreev, A.; Tarasov, P.; Schwamborn, G.; Iljashuk, B.; Iljashuk, E.; Bobrov, A.; Klimanov, V.; Rachold, V. Holocene palaeoenvironmental records from Nikolay Lake, Lena Delta, Arctic Russia. Palaeogeogr. Palaeoclim. Palaeoecol. 2004, 209, 197–217. [Google Scholar] [CrossRef] [Green Version]
- Brooks, S.J.; Langdon, P.G.; Heiri, O. Using and Identifying Chironomid Larvae in Palaeoecology; QRA Technical Guide No 10; Quaternary Research Association: London, UK, 2007. [Google Scholar]
- Plikk, A.; Engels, S.; Luoto, T.P.; Nazarova, L.; Salonen, J.S.; Helmens, K.F. Chironomid-based temperature reconstruction for the Eemian Interglacial (MIS 5e) at Sokli, northeast Finland. J. Paleolimnol. 2019, 61, 355–371. [Google Scholar] [CrossRef] [Green Version]
- Płóciennik, M.; Mroczkowska, A.; Pawłowski, D.; Wieckowska-Lüth, M.; Kurzawska, A.; Rzodkiewicz, M.; Okupny, D.; Szmańda, J.; Mazurkevich, A.; Dolbunova, E.; et al. Summer temperature drives the lake ecosystem during the Late Weichselian and Holocene in Eastern Europe: A case study from East European Plain. Catena 2022, 214, 106206. [Google Scholar] [CrossRef]
- Renberg, I. The HON-Kajak sediment corer. J. Paleolimnol. 1991, 6, 167–170. [Google Scholar] [CrossRef]
- New, M.; Lister, D.; Hulme, M.; Makin, I. A high-resolution data set of surface climate over global land areas. Clim. Res. 2002, 21, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Overland, J.E.; Wang, M. The Arctic Climate Paradox: The Recent Decrease of the Arctic Oscillation. Geophys. Res. Lett. 2005, 32, L06701. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.; Ferrians, O.J., Jr.; Heginbottom, J.A.; Melnikov, E.S. Circum-Arctic Map of Permafrost and Ground Ice Conditions; National Snow and Ice Data Center/World Data Center for Glaciology; Digital Media: Boulder, CO, USA, 1998. [Google Scholar] [CrossRef]
- Esri, R. ArcGIS Desktop: Release 10.6; Environmental Systems Research Institute: Redlands, CA, USA, 2011. [Google Scholar]
- Wiederholm, T. Chironomidae of the Holarctic region. Keys and diagnoses. Part 1. Larvae. Entomol. Scand. 1983, 9 (Suppl. S19), 1–457. [Google Scholar]
- Heiri, O.; Lotter, A.F. Effect of low count sums on quantitative environmental reconstructions: An example using subfossil chironomids. J. Paleolimnol. 2001, 26, 343–350. [Google Scholar] [CrossRef]
- Quinlan, R.; Smol, J. Setting minimum head capsule abundance and taxa deletion criteria in chironomid-based inference models. J. Paleolimnol. 2001, 26, 327–342. [Google Scholar] [CrossRef]
- Biskaborn, B.K.; Nazarova, L.; Pestryakova, L.A.; Syrykh, L.; Funck, K.; Meyer, H.; Chapligin, B.; Vyse, S.; Gorodnichev, R.; Zakharov, E.; et al. Spatial distribution of environmental indicators in surface sediments of Lake Bolshoe Toko. Biogeosciences 2019, 16, 4023–4049. [Google Scholar] [CrossRef] [Green Version]
- Kumke, T.; Ksenofontova, M.; Pestryakova, L.; Nazarova, L.; Hubberten, H.-W. Limnological characteristics of lakes in the lowlands of Central Yakutia, Russia. J. Limnol. 2007, 66, 40–53. [Google Scholar] [CrossRef] [Green Version]
- Sokal, R.R.; Rohlf, F.J. Biometry: The Principles and Practice of Statistics in Biological Research; W. H. Freeman and Co.: New York, NY, USA, 1995. [Google Scholar]
- Palagushkina, O.V.; Nazarova, L.B.; Wetterich, S.; Schirrmeister, L. Diatoms of modern bottom sediments in Siberian arctic. Contemp. Probl. Ecol. 2012, 5, 413–422. [Google Scholar] [CrossRef] [Green Version]
- Lepš, J.; Šmilauer, P. Multivariate Analysis of Ecological Data Using CANOCO; Cambridge University Press: New York, NY, USA, 2003. [Google Scholar]
- Birks, H.J.B. Quantitative Palaeoenvironmental Reconstructions. In Statistical Modelling of Quaternary Science Data; Technical guide 5; Quaternary Research Association: Cambridge, UK, 1995; pp. 161–254. [Google Scholar]
- Nazarova, L.; Bleibtreu, A.; Hoff, U.; Dirksen, V.; Diekmann, B. Changes in temperature and water depth of a small mountain lake during the past 3000 years in Central Kamchatka reflected by a chironomid record. Quat. Int. 2017, 447, 46–58. [Google Scholar] [CrossRef]
- ter Braak, C.J.F. Ordination; Jongman, R.H.G., ter Braak, C.J.F., Tongeren, O.F.R., Eds.; Cambridge University Press: Cambridge, UK, 1995. [Google Scholar]
- Šmilauer, P.; Lepš, J. Multivariate Analysis of Ecological Data using CANOCO 5; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar]
- ter Braak, C.J.F. Update Notes: CANOCO Version 3.10; Agricultural Mathematics Group: Wageningen, The Netherlands, 1990. [Google Scholar]
- ter Braak, C.J.F.; Šmilauer, P. CANOCO for Windows: Software for Community Ordination (Version 4.5); Microcomputer Power: Ithaca, NY, USA, 2002. [Google Scholar]
- Gavin, D.G.; Oswald, W.; Wahl, E.R.; Williams, J.W. A statistical approach to evaluating distance metrics and analog assignments for pollen records. Quat. Res. 2003, 60, 356–367. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Raan, P.D. PAST: Palaeontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 41, 9. [Google Scholar]
- Dowsett, H.J.; Robinson, M.M. Application of the modern analogue technique (MAT) of sea surface temperature estimation to middle Pliocene North Pacific planktic foraminifer assemblages. Palaeontol. Electron. 1998, 1, 22. [Google Scholar]
- ter Braak, C.J.F.; Looman, C.W.N. Weighted Averaging, Logistic Regression and the Gaussian Response Model. Vegetatio 1986, 65, 3–11. [Google Scholar] [CrossRef]
- ter Braak, C.J.F.; Juggins, S. Weighted Averaging Partial Least Squares Regression (WA-PLS): An Improved Method for Reconstructing Environmental Variables from Species Assemblages. Hydrobiologia 1993, 269–270, 485–502. [Google Scholar] [CrossRef]
- Altman, D.G.; Bland, J.M. Measurement in Medicine: The Analysis of Method Comparison Studies. Statistician 1983, 32, 307–317. [Google Scholar] [CrossRef]
- Birks, H.J.B. Numerical tools in paleolimnology progress, potential, and problems. J Paleolimnol. 1998, 20, 307–332. [Google Scholar] [CrossRef]
- Birks, H.J.B.; Juggins, S.; Line, J.M. Lake Surface-Water Chemistry Reconstructions from Paleolimnolical Data; Mason, B., Ed.; Cambridge University Press: Cambridge, UK, 1990. [Google Scholar]
- Birks, H.J.B.; Line, J.M.; Juggins, S.; Stevenson, A.C.; ter Braak, C.J.F. Diatoms and pH Reconstruction. Philos. Trans. R. Soc. B 1990, 327, 263–278. [Google Scholar]
- Brooks, S.J.; Birks, H.J.B. Chironomid-Inferred Air Temperatures from Late-Glacial and Holocene Sites in North-West Europe: Progress and Problems. Quat. Sci. Rev. 2001, 20, 1723–1741. [Google Scholar] [CrossRef]
- Juggins, S. C2 Version 1.5 User Guide. Software for Ecological and Palaeoecological Data Analysis and Visualisation; Newcastle University: Newcastle upon Tyne, UK, 2007. [Google Scholar]
- Juggins, S. Quantitative Reconstructions in Paleolimnology: New Paradigm or Sick Science? Quat. Sci. Rev. 2013, 64, 20–32. [Google Scholar] [CrossRef]
- Syrykh, L.S.; Nazarova, L.B.; Herzschuh, U.; Subetto, D.A.; Grekov, I.M. Reconstruction of palaeoecological and palaeoclimatic conditions of the Holocene in the south of the Taimyr according to an analysis of lake sediments. Contemp. Probl. Ecol. 2017, 4, 363–369. [Google Scholar] [CrossRef] [Green Version]
- Nazarova, L.; Grebennikova, T.A.; Razjigaeva, N.G.; Ganzey, L.A.; Belyanina, N.I.; Arslanov, K.A.; Kaistrenko, V.M.; Gorbunov, A.O.; Kharlamov, A.A.; Rudaya, N.; et al. Reconstruction of Holocene environmental changes in Southern Kurils (North-Western Pacific) based on palaeolake sediment proxies from Shikotan Island. Glob. Planet. Chang. 2017, 159, 25–36. [Google Scholar] [CrossRef]
- Nazarova, L.; Syrykh, L.S.; Mayfield, R.J.; Frolova, L.A.; Ibragimova, A.G.; Grekov, I.M.; Subetto, D.A. Palaeoecological and palaeoclimatic conditions on the Karelian Isthmus (northwestern Russia) during the Holocene: Multi-Proxy Analysis of Sediments from the Lake Medvedevskoe. Quat. Res. 2020, 95, 65–83. [Google Scholar] [CrossRef] [Green Version]
- Druzhinina, O.; Kublitskii, J.; Nazarova, L.; Subetto, D.; Syrykh, L.; Arslanov, K.; Stančikaitė, M.; Vaikutienė, G.; Kul`kova, M. Palaeoenvironmental Conditions in South-Eastern Part of the Baltic Region during the Late Pleistocene—Holocene Transition (Kaliningrad District, Russia. Boreas 2020, 49, 544–561. [Google Scholar] [CrossRef]
- Mayfield, R.J.; Langdon, P.G.; Doncaster, C.P.; Dearing, J.A.; Wang, R.; Nazarova, L.B.; Medeiros, A.S.; Brooks, S.J. Metrics of structural change as indicators of chironomid community stability in high latitude lakes. Quat. Sci. Rev. 2020, 249, 106594. [Google Scholar] [CrossRef]
- MacDonald, G.M.; Szeicz, J.M.; Claricoates, J.; Dale, K.A. Response of the Central Canadian Treeline to Recent Climatic Changes. Ann. Assoc. Am. Geogr. 1998, 88, 183–208. [Google Scholar] [CrossRef]
- Macdonald, G.M.; Velichko, A.A.; Kremenetski, C.V.; Borisova, O.K.; Goleva, A.A.; Andreev, A.; Cwynar, L.C.; Riding, R.T.; Forman, S.; Edwards, T.W.; et al. Holocene Treeline History and Climate Change Across Northern Eurasia. Quat. Res. 2000, 53, 302–311. [Google Scholar] [CrossRef] [Green Version]
- Harsch, M.A.; Bader, M. Treeline form—A potential key to understanding treeline dynamics. Glob. Ecol. Biogeogr. 2011, 20, 582–596. [Google Scholar] [CrossRef]
- Self, A.; Klimaschewski, A.; Solovieva, N.; Jones, V.; Andrén, E.; Andreev, A.; Hammarlund, D.; Brooks, S. The relative influences of climate and volcanic activity on Holocene lake development inferred from a mountain lake in central Kamchatka. Glob. Planet. Chang. 2015, 134, 67–81. [Google Scholar] [CrossRef]
- Wetterich, S.; Schirrmeister, L.; Nazarova, L.; Palagushkina, O.; Bobrov, A.; Pogosyan, L.; Savelieva, L.; Syrykh, L.; Matthes, H.; Fritz, M.; et al. Holocene thermokarst and pingo development in the Kolyma Lowland (NE Siberia). Permafr. Periglac. 2018, 29, 182–198. [Google Scholar] [CrossRef]
- Porinchu, D.F.; Cwynar, L.S. Late-Quaternary History of Midge Communities and Climate from a Tundra Site near the Lower Lena River, Northeast Siberia. J. Paleolimnol. 2002, 27, 59–69. [Google Scholar] [CrossRef]
- Nazarova, L.; Sachse, D.; Fuchs, H.G.E.; Dirksen, V.; Dirksen, O.; Syrykh, L.; Razjigaeva, N.G.; Rach, O.; Diekmann, B. Holocene evolution of a proglacial lake in southern Kamchatka, Russian Far East. Boreas 2021, 50, 1011–1026. [Google Scholar] [CrossRef]
- Brodin, Y.W. The Postglacial History of Lake Flarken, Southern Sweden, Interpreted from Subfossil Insect Remains. Int. Rev. Gesamten Hydrobiol. Hydrogr. 1986, 71, 371–432. [Google Scholar] [CrossRef]
- Brundin, L. Zur Systematic Der Orthocladiinae (Dipt., Chironomidae) Report of the Institute of Freshwater Research. Drottningholm 1956, 37, 5–186. [Google Scholar]
- Quinlan, R.; Smol, J.P.; Hall, R.I. Quantitative Inferences of Past Hypolimnetic Anoxia in South-Central Ontario Lakes using Fossil Midges (Diptera: Chironomidae). Can. J. Fish. Aquat. Sci. 1998, 55, 587–596. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters * | Min. | Max. | Mean | Median | St.dev | Skew |
|---|---|---|---|---|---|---|
| Latitude, N | 56.22 | 70.59 | 64.17 | 64.89 | 3.22 | −0.26 |
| Longitude, E | 28.85 | 66.32 | 42.05 | 36.03 | 12.77 | 0.50 |
| Altitude, m a.s.l. | 0 | 514 | 74.30 | 60.00 | 71.02 | 2.76 |
| T July °C | 9.20 | 18.10 | 15.04 | 15.1 | 1.97 | −0.51 |
| Water depth, m | 0.70 | 140 | 9.74 | 5.7 | 16.39 | 5.88 |
| pH | 4.54 | 9.5 | 6.72 | 6.8 | 0.86 | −0.05 |
| Distance from treeline, km | −1434 | 356 | −390.42 | −275 | 403.29 | −0.35 |
| Water depth, log 10 | −0.15 | 2.15 | 0,77 | 0.76 | 0.40 | 0.59 |
| Altitude, log 10 | 0.18 | 2.71 | 1.67 | 1.78 | 0.50 | −0.66 |
| Lake | NZ | KM | KO | AS | OP | PE | KZ | KL | EU | Median |
|---|---|---|---|---|---|---|---|---|---|---|
| NZ | 0 | 1.39 | 1.38 | 1.37 | 1.38 | 1.35 | 1.35 | 1.28 | 1.38 | 1.37 |
| KM | 1.39 | 0 | 1.06 | 1.00 | 0.87 | 1.02 | 1.03 | 1.04 | 1.20 | 1.03 |
| KP | 1.38 | 1.06 | 0 | 0.94 | 0.77 | 0.85 | 0.96 | 0.84 | 1.28 | 0.94 |
| AS | 1.37 | 1.00 | 0.94 | 0 | 0.70 | 1.06 | 1.00 | 1.06 | 1.30 | 1.01 |
| OP | 1.38 | 0.87 | 0.77 | 0.70 | 0 | 0.98 | 1.03 | 0.95 | 1.28 | 0.95 |
| PE | 1.35 | 1.02 | 0.85 | 1.06 | 0.98 | 0 | 0.90 | 0.95 | 1.20 | 0.98 |
| KZ | 1.35 | 1.03 | 0.96 | 1.00 | 1.03 | 0.90 | 0 | 0.88 | 1.06 | 1.01 |
| KL | 1.28 | 1.04 | 0.84 | 1.06 | 0.95 | 0.95 | 0.88 | 0 | 1.18 | 0.96 |
| EU | 1.38 | 1.20 | 1.28 | 1.30 | 1.28 | 1.20 | 1.06 | 1.18 | 0 | 1.20 |
| Median | 1.37 | 1.03 | 0.94 | 1.01 | 0.95 | 0.98 | 1.01 | 0.96 | 1.20 | 1.02 |
| Variable | Variance Explained | % Total Variance Explained | F Value | Significance Leve |
|---|---|---|---|---|
| Mean July air temperature (T July) | 0.155 | 38.5 | 4.069 | 0.002 |
| Water depth (log 10) | 0.092 | 22.8 | 2.480 | 0.002 |
| Distance from tree line | 0.084 | 20.8 | 2.294 | 0.002 |
| pH | 0.062 | 15.4 | 1.969 | 0.002 |
| Altitude | 0.010 | 2.5 | 1.700 | 0.008 |
| Total variance explained | 0.403 | |||
| Total variance | 3.293 |
| Parameters | Canonical Coefficients | t-Values | Interset Correlation | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Axis 1 | Axis 2 | Axis 3 | Axis 4 | Axis 1 | Axis 2 | Axis 3 | Axis 4 | Axis 1 | Axis 2 | Axis 3 | Axis 4 | |
| T July | −0.41 | −0.03 | −0.39 | −0.23 | −5.31 | −0.46 | −6.38 | −3.53 | −0.73 | −0.02 | −0.32 | −0.07 |
| DTL | 0.39 | −0.42 | −0.32 | −0.20 | 5.29 | −5.35 | −5.18 | −2.97 | −0.18 | 0.76 | 0.22 | −0.03 |
| WD | 0.16 | 0.18 | −0.20 | −0.02 | 5.17 | 5.45 | −7.76 | −0.81 | 0.18 | 0.46 | −0.53 | 0.003 |
| pH | 0.02 | −0.11 | 0.08 | 0.24 | 0.55 | −3.26 | −2.96 | 7.97 | −0.001 | −0.24 | −0.30 | 0.55 |
| Alt | 0.01 | −0.13 | −0.05 | −0.11 | 0.35 | −3.89 | −2.05 | −4.41 | 0.13 | −0.20 | −0.180 | −0.150 |
| WAPLS Component | RMSE | r2 | Av Bias | Max Bias | r2 boot | Ave Bias boot | Max_ Bias boot | RMSEP | % Change | t-Test |
|---|---|---|---|---|---|---|---|---|---|---|
| Full model (98 lakes. 143 taxa) | ||||||||||
| 1 | 1.113 | 0.68 | 0.005 | 1.344 | 0.45 | −0.047 | 3.63 | 1.51 | … | … |
| 2 | 0.833 | 0.82 | −0.044 | 0.857 | 0.49 | −0.050 | 3.92 | 1.49 | 1.16 | 0.16 |
| 3 | 0.638 | 0.89 | 0.022 | 0.775 | 0.47 | −0.052 | 3.27 | 1.59 | −6.40 | 0.68 |
| 4 | 0.497 | 0.94 | 0.002 | 0.805 | 0.44 | −0.037 | 2.90 | 1.68 | −5.76 | 0.90 |
| 5 | 0.414 | 0.96 | −0.005 | 0.654 | 0.43 | −0.043 | 2.78 | 1.75 | −3.92 | 0.98 |
| Minus 2 lakes (96 lakes. 143 taxa) | ||||||||||
| 1 | 1.086 | 0.69 | −0.005 | 1.647 | 0.58 | −0.090 | 3.27 | 1.37 | … | … |
| 2 | 0.778 | 0.84 | −0.040 | 0.722 | 0.60 | −0.082 | 3.37 | 1.34 | 1.63 | 0.07 |
| 3 | 0.583 | 0.91 | 0.017 | 0.837 | 0.61 | −0.097 | 2.66 | 1.38 | −2.08 | 0.12 |
| 4 | 0.455 | 0.95 | −0.003 | 0.648 | 0.60 | −0.078 | 2.29 | 1.44 | −4.17 | 0.81 |
| 5 | 0.374 | 0.96 | −0.007 | 0.399 | 0.59 | −0.077 | 2.18 | 1.48 | −3.23 | 0.91 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nazarova, L.; Syrykh, L.; Grekov, I.; Sapelko, T.; Krasheninnikov, A.B.; Solovieva, N. Chironomid-Based Modern Summer Temperature Data Set and Inference Model for the Northwest European Part of Russia. Water 2023, 15, 976. https://doi.org/10.3390/w15050976
Nazarova L, Syrykh L, Grekov I, Sapelko T, Krasheninnikov AB, Solovieva N. Chironomid-Based Modern Summer Temperature Data Set and Inference Model for the Northwest European Part of Russia. Water. 2023; 15(5):976. https://doi.org/10.3390/w15050976
Chicago/Turabian StyleNazarova, Larisa, Liudmila Syrykh, Ivan Grekov, Tatiana Sapelko, Andrey B. Krasheninnikov, and Nadia Solovieva. 2023. "Chironomid-Based Modern Summer Temperature Data Set and Inference Model for the Northwest European Part of Russia" Water 15, no. 5: 976. https://doi.org/10.3390/w15050976
APA StyleNazarova, L., Syrykh, L., Grekov, I., Sapelko, T., Krasheninnikov, A. B., & Solovieva, N. (2023). Chironomid-Based Modern Summer Temperature Data Set and Inference Model for the Northwest European Part of Russia. Water, 15(5), 976. https://doi.org/10.3390/w15050976