

Photoinhibition of the Picophytoplankter Synechococcus Is Exacerbated by Ocean Acidification


Abstract
:1. Introduction
2. Materials and Methods
2.1. Cultures and Experimental Design
2.2. Carbonate Chemistry System
2.3. Growth Rates
2.4. Chlorophyll a Content and Optical Absorption Cross Section
2.5. Chlorophyll a Fluorescence
2.6. Carbon Fixation Rates
2.7. C and N Analysis
2.8. Data Analysis
3. Results
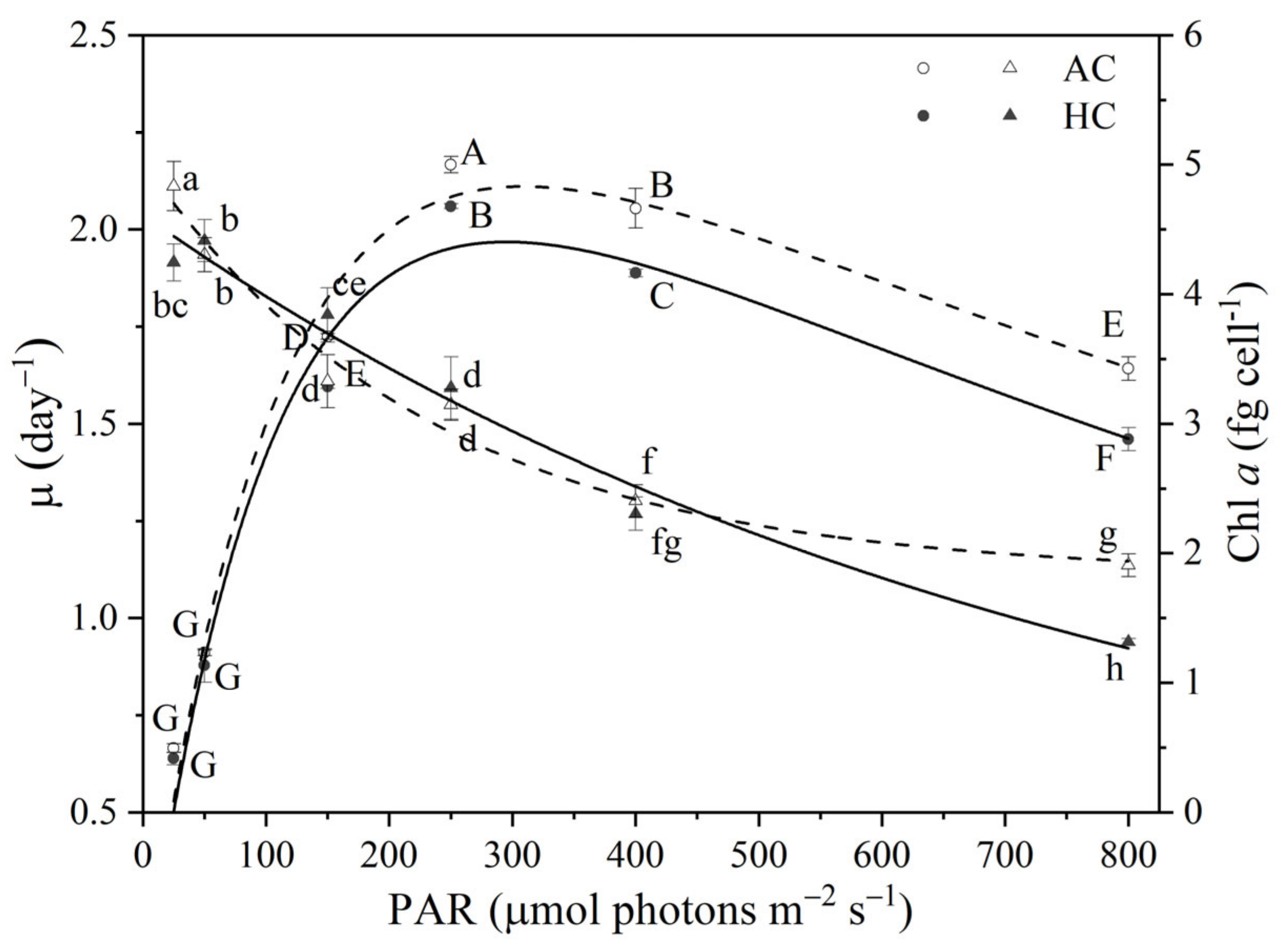
3.1. Growth and Chl a
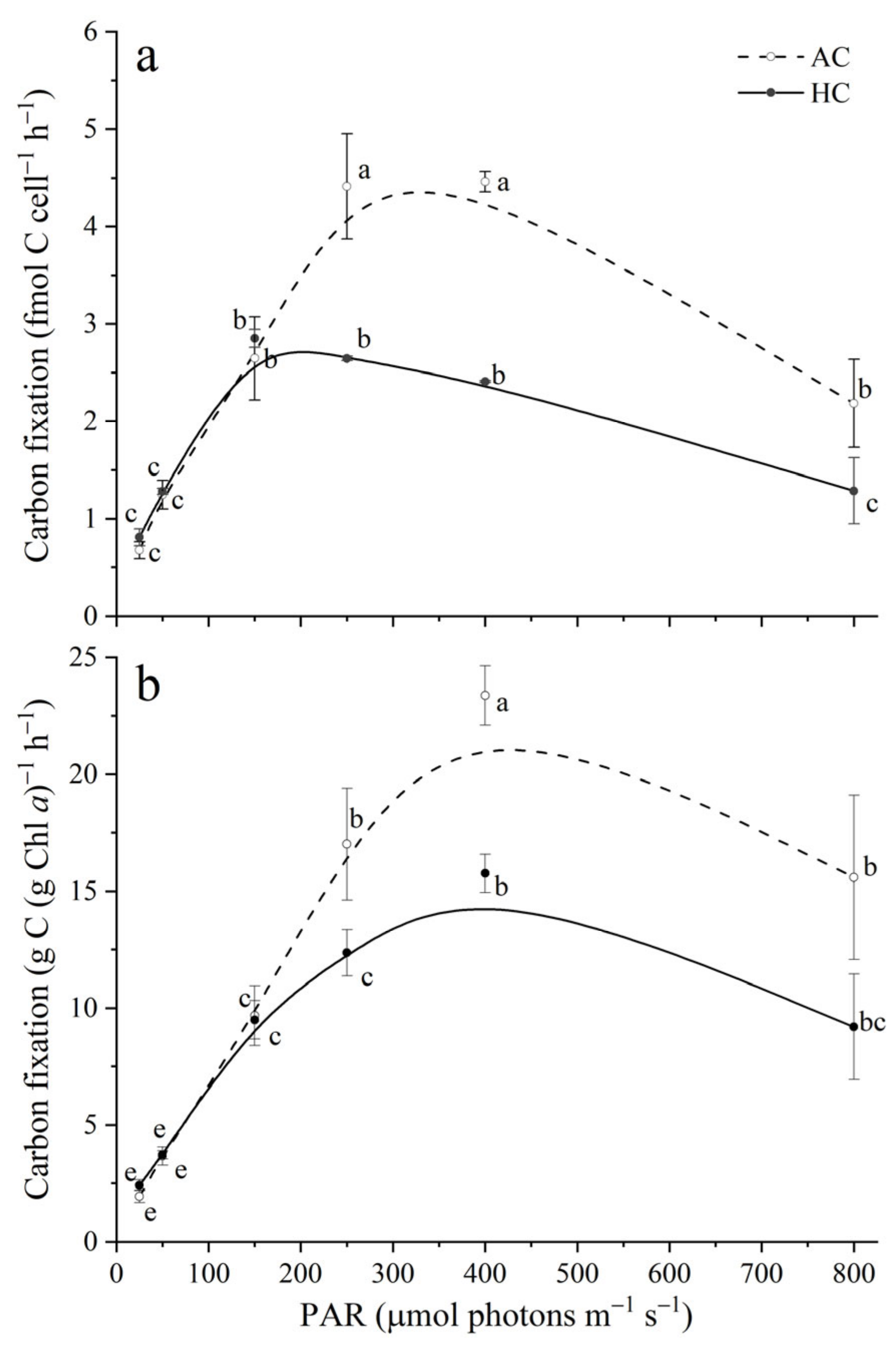
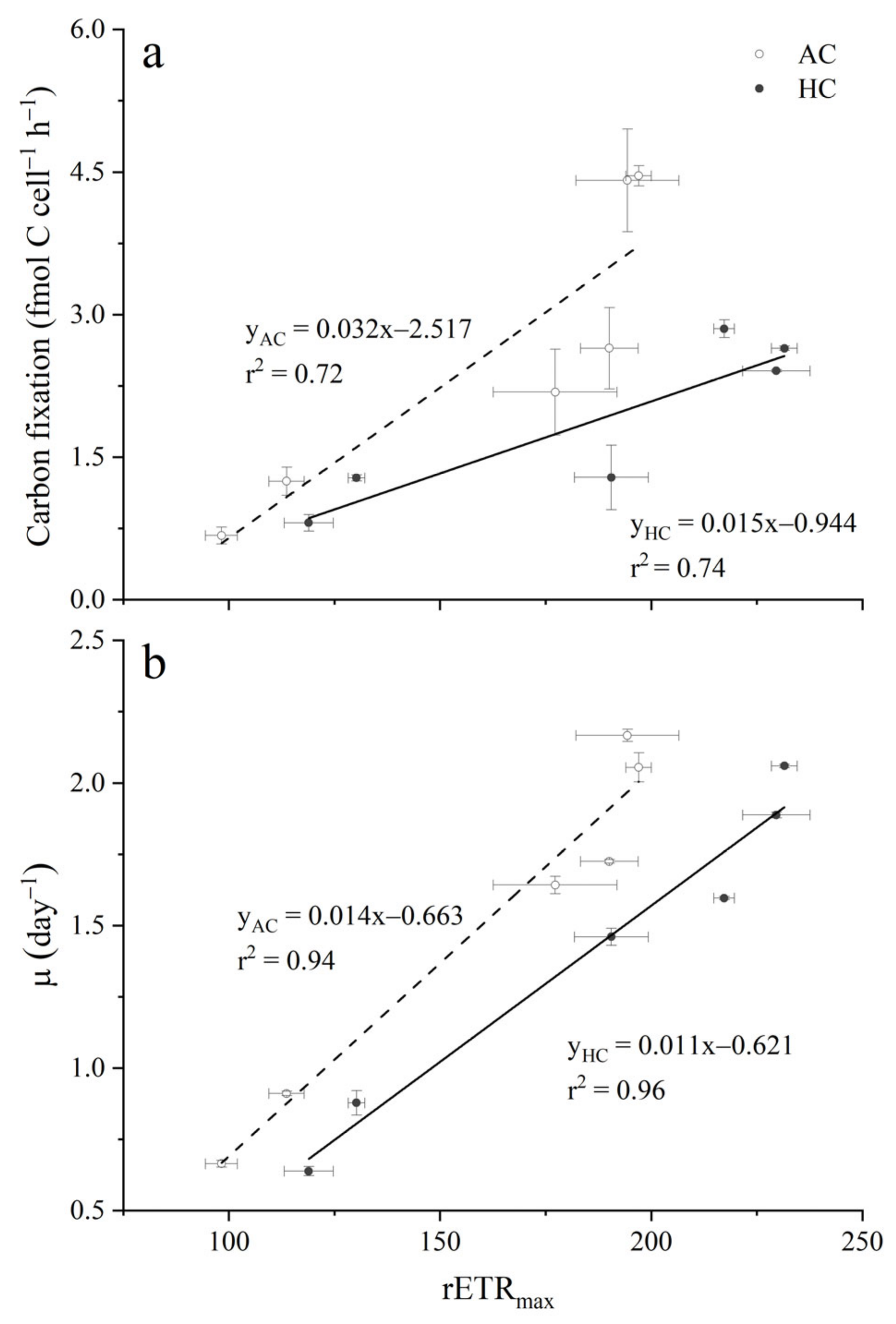
3.2. Carbon Fixation
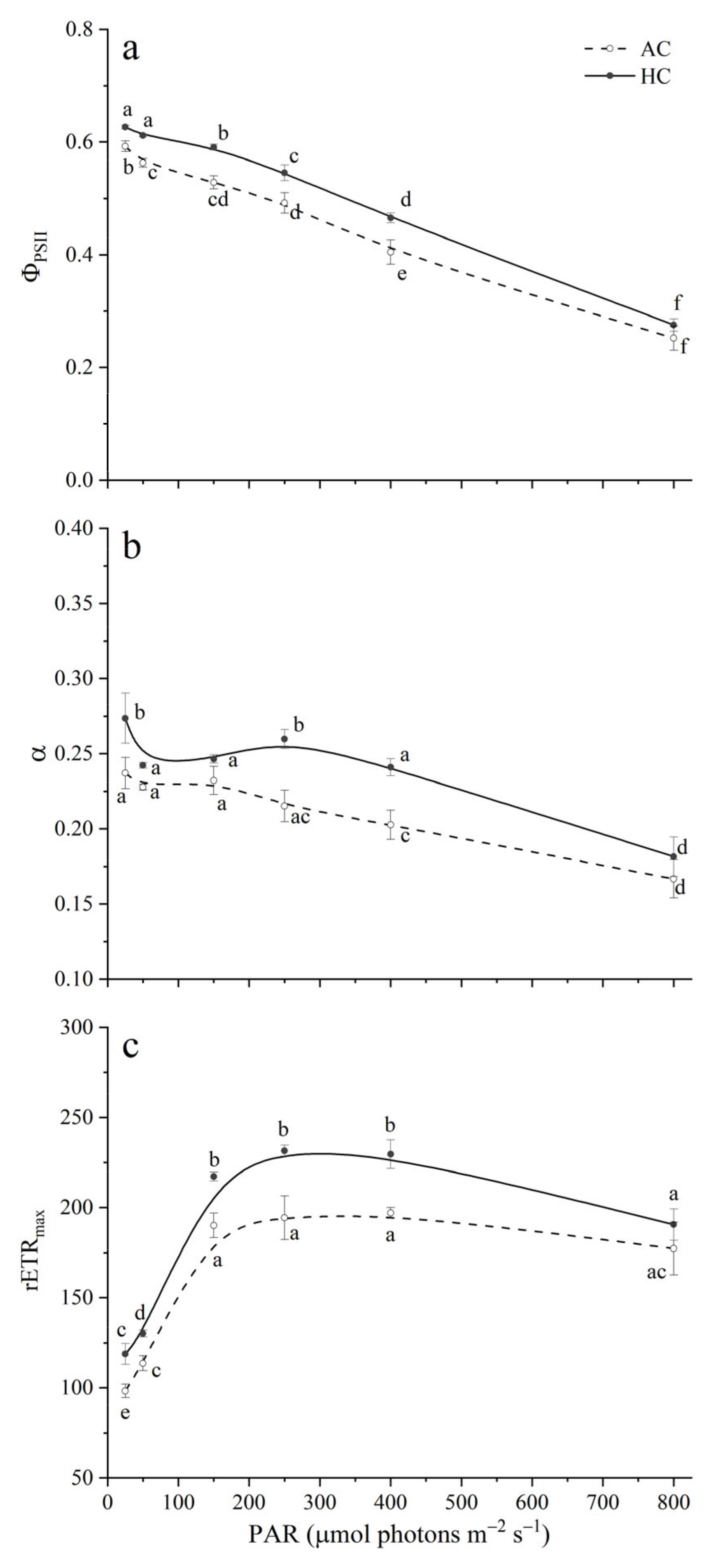
3.3. Chlorophyll a Fluorescence
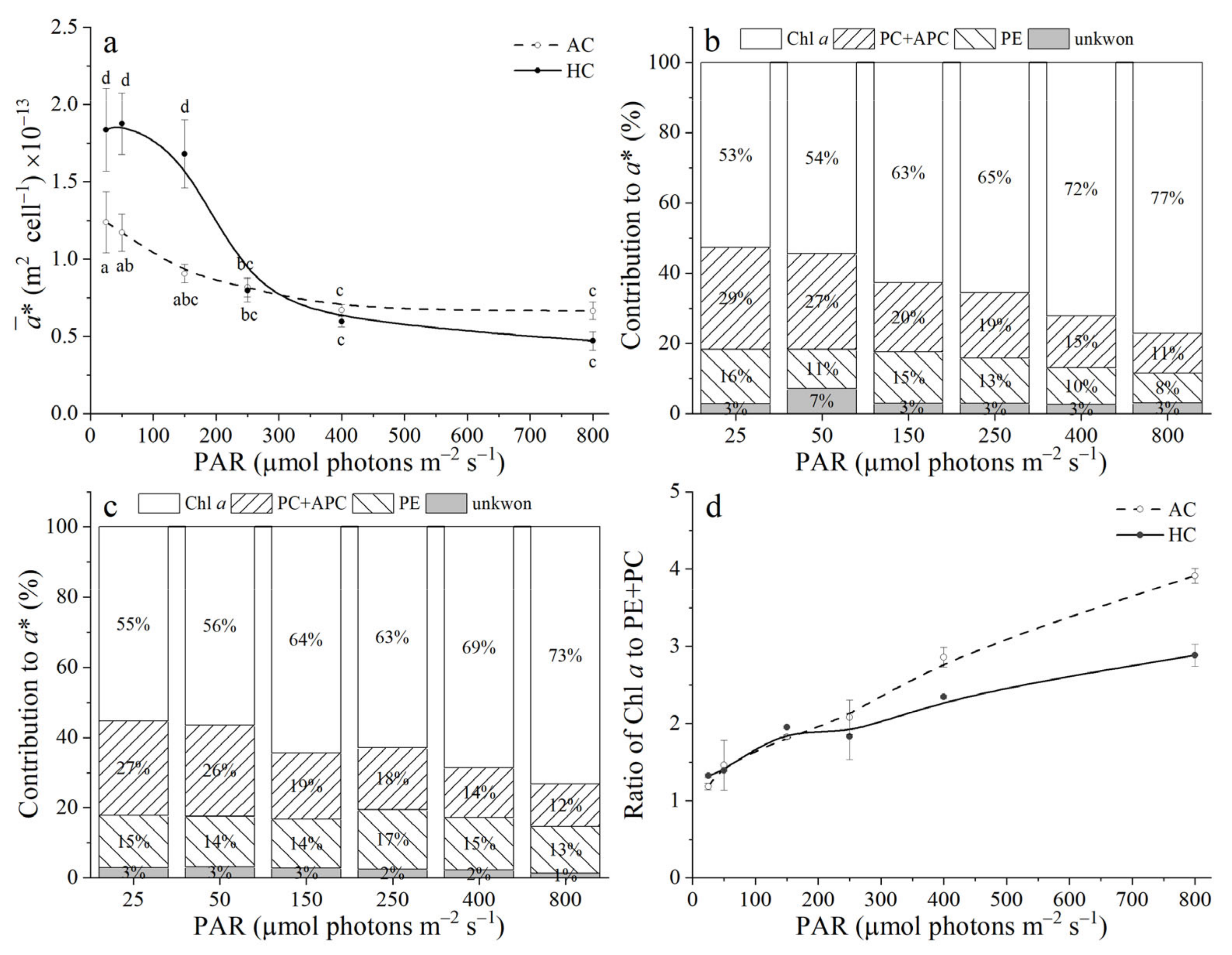
3.4. Optical Absorption Cross Section
3.5. Cellular POC Content and POC
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Flombaum, P.; Gallegos, J.L.; Gordillo, R.A.; Rincón, J.; Zabala, L.L.; Jiao, N.; Karl, D.M.; Li, W.K.; Lomas, M.W.; Veneziano, D. Present and future global distributions of the marine Cyanobacteria Prochlorococcus and Synechococcus. Proc. Natl. Acad. Sci. USA 2013, 110, 9824–9829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.; Jeon, J.; Kwak, M.S.; Kim, G.H.; Koh, I.; Rho, M. Photosynthetic functions of Synechococcus in the ocean microbiomes of diverse salinity and seasons. PLoS ONE 2018, 13, e0190266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Partensky, F.; Blanchot, J.; Vaulot, D. Differential distribution and ecology of Prochlorococcus and Synechococcus in oceanic waters: A review. Bull.-Inst. Oceanogr. Monaco-Numero Spec. 1999, 19, 457–476. [Google Scholar]
- Parli, B.V.; Bhaskar, J.T.; Jawak, S.; Jyothibabu, R.; Mishra, N. Mixotrophic plankton and Synechococcus distribution in waters around Svalbard, Norway during June 2019. Polar Sci. 2021, 30, 100697. [Google Scholar] [CrossRef]
- Fucich, D.; Chen, F. Presence of toxin-antitoxin systems in picocyanobacteria and their ecological implications. ISME J. 2020, 14, 2843–2850. [Google Scholar] [CrossRef]
- Wang, T.; Li, J.; Jing, H.; Qin, S. Picocyanobacterial Synechococcus in marine ecosystem: Insights from genetic diversity, global distribution, and potential function. Mar. Environ. Res. 2022, 177, 105622. [Google Scholar] [CrossRef]
- Guidi, L.; Chaffron, S.; Bittner, L.; Eveillard, D.; Larhlimi, A.; Roux, S.; Darzi, Y.; Audic, S.; Berline, L.; Brum, J.R.; et al. Plankton networks driving carbon export in the oligotrophic ocean. Nature 2016, 532, 465–470. [Google Scholar] [CrossRef] [Green Version]
- Howes, E.L.; Joos, F.; Eakin, C.M.; Gattuso, J.-P. An updated synthesis of the observed and projected impacts of climate change on the chemical, physical and biological processes in the oceans. Front. Mar. Sci. 2015, 2, 36. [Google Scholar] [CrossRef]
- Orr, J.C.; Fabry, V.J.; Aumont, O.; Bopp, L.; Doney, S.C.; Feely, R.A.; Gnanadesikan, A.; Gruber, N.; Ishida, A.; Joos, F. Anthropogenic ocean acidification over the twenty-first century and its impact on calcifying organisms. Nature 2005, 437, 681–686. [Google Scholar] [CrossRef] [Green Version]
- Gattuso, J.-P.; Magnan, A.; Billé, R.; Cheung, W.W.; Howes, E.L.; Joos, F.; Allemand, D.; Bopp, L.; Cooley, S.R.; Eakin, C.M. Contrasting futures for ocean and society from different anthropogenic CO2 emissions scenarios. Science 2015, 349, aac4722. [Google Scholar] [CrossRef]
- Cai, W.-J.; Hu, X.; Huang, W.-J.; Murrell, M.C.; Lehrter, J.C.; Lohrenz, S.E.; Chou, W.-C.; Zhai, W.; Hollibaugh, J.T.; Wang, Y. Acidification of subsurface coastal waters enhanced by eutrophication. Nat. Geosci. 2011, 4, 766–770. [Google Scholar] [CrossRef]
- Feely, R.A.; Alin, S.R.; Newton, J.; Sabine, C.L.; Warner, M.; Devol, A.; Krembs, C.; Maloy, C. The combined effects of ocean acidification, mixing, and respiration on pH and carbonate saturation in an urbanized estuary. Estuar. Coast. Shelf Sci. 2010, 88, 442–449. [Google Scholar] [CrossRef]
- Giordano, M.; Beardall, J.; Raven, J.A. CO2 concentrating mechanisms in algae: Mechanisms, environmental modulation, and evolution. Annu. Rev. Plant Biol. 2005, 56, 99–131. [Google Scholar] [CrossRef] [Green Version]
- Beardall, J.; Giordano, M. Ecological implications of microalgal and cyanobacterial CO2 concentrating mechanisms, and their regulation. Funct. Plant Biol. 2002, 29, 335–347. [Google Scholar] [CrossRef]
- Gao, K.; Gao, G.; Wang, Y.; Dupont, S. Impacts of ocean acidification under multiple stressors on typical organisms and ecological processes. Mar. Life Sci. Technol. 2020, 2, 279–291. [Google Scholar] [CrossRef]
- McNicholl, C.; Koch, M.; Swarzenski, P.; Oberhaensli, F.; Taylor, A.; Batista, M.G.; Metian, M. Ocean acidification effects on calcification and dissolution in tropical reef macroalgae. Coral Reefs 2020, 39, 1635–1647. [Google Scholar] [CrossRef]
- Raven, J.A.; Beardall, J.; Quigg, A. Light-Driven Oxygen Consumption in the Water-Water Cycles and Photorespiration, and Light Stimulated Mitochondrial Respiration. In Photosynthesis in Algae: Biochemical and Physiological Mechanisms; Larkum, A.W.D., Grossman, A.R., Raven, J.A., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 161–178. [Google Scholar]
- Qu, L.; Beardall, J.; Jiang, X.; Gao, K. Elevated pCO2 enhances under light but reduces in darkness the growth rate of a diatom, with implications for the fate of phytoplankton below the photic zone. Limnol. Oceanogr. 2021, 66, 3630–3642. [Google Scholar] [CrossRef]
- Gao, K.; Xu, J.; Gao, G.; Li, Y.; Hutchins, D.A.; Huang, B.; Wang, L.; Zheng, Y.; Jin, P.; Cai, X.; et al. Rising CO2 and increased light exposure synergistically reduce marine primary productivity. Nat. Clim. Chang. 2012, 2, 519–523. [Google Scholar] [CrossRef]
- Jin, P.; Ding, J.; Xing, T.; Riebesell, U.; Gao, K. High levels of solar radiation offset impacts of ocean acidification on calcifying and non-calcifying strains of Emiliania huxleyi. Mar. Ecol. Prog. Ser. 2017, 568, 47–58. [Google Scholar] [CrossRef] [Green Version]
- Ji, Y.; Gao, K. Effects of climate change factors on marine macroalgae: A review. Adv. Mar. Biol. 2021, 88, 91–136. [Google Scholar] [CrossRef]
- Fu, F.X.; Warner, M.E.; Zhang, Y.; Feng, Y.; Hutchins, D.A. Effects of Increased temperature and CO2 on photosynthesis, growth, and elemental ratios in marine Synechococcus and Prochlorococcus (cyanobacteria). J. Phycol. 2007, 43, 485–496. [Google Scholar] [CrossRef]
- Basu, S.; Mackey, K.R. Effect of Rising Temperature and Carbon Dioxide on the Growth, Photophysiology, and Elemental Ratios of Marine Synechococcus: A Multistressor Approach. Sustainability 2022, 14, 9508. [Google Scholar] [CrossRef]
- Mou, S.; Zhang, Y.; Li, G.; Li, H.; Liang, Y.; Tang, L.; Tao, J.; Xu, J.; Li, J.; Zhang, C. Effects of elevated CO2 and nitrogen supply on the growth and photosynthetic physiology of a marine cyanobacterium, Synechococcus sp. PCC7002. J. Appl. Phycol. 2017, 29, 1755–1763. [Google Scholar] [CrossRef]
- Bao, N.; Gao, K. Interactive effects of elevated CO2 concentration and light on the picophytoplankton Synechococcus. Front. Mar. Sci. 2021, 8, 634189. [Google Scholar] [CrossRef]
- Laws, E.A.; McClellan, S.A. Interactive effects of CO2, temperature, irradiance, and nutrient limitation on the growth and physiology of the marine cyanobacterium Synechococcus (Cyanophyceae). J. Phycol. 2022, 58, 703–718. [Google Scholar] [CrossRef]
- Wang, K.; Wommack, K.E.; Chen, F. Abundance and distribution of Synechococcus spp. and cyanophages in the Chesapeake Bay. Appl. Environ. Microb. 2011, 77, 7459–7468. [Google Scholar] [CrossRef] [Green Version]
- Marsan, D.; Wommack, K.E.; Ravel, J.; Chen, F. Draft genome sequence of Synechococcus sp. strain CB0101, isolated from the Chesapeake Bay estuary. Genome Announc. 2014, 2, e01111-13. [Google Scholar] [CrossRef] [Green Version]
- Chen, F.; Wang, K.; Kan, J.; Bachoon, D.S.; Lu, J.; Lau, S.; Campbell, L. Phylogenetic diversity of Synechococcus in the Chesapeake Bay revealed by Ribulose-1, 5-bisphosphate carboxylase-oxygenase (RuBisCO) large subunit gene (rbcL) sequences. Aquat. Microb. Ecol. 2004, 36, 153–164. [Google Scholar] [CrossRef] [Green Version]
- Waterbury, J.B. Biological and ecological characterization of the marine unicellular cyanobacterium Synechococcus. Can Bull. Fish Aquat. Sci. 1986, 214, 71–120. [Google Scholar]
- Lewis, E.; Wallace, D. Program Developed for CO2 System Calculations; Environmental System Science Data Infrastructure for a Virtual Ecosystem. 1998. Available online: https://www.osti.gov/servlets/purl/639712 (accessed on 8 March 2023).
- Platt, T.; Gallegos, C.; Harrison, W.G. Photoinhibition of photosynthesis in natural assemblages of marine phytoplankton. J. Mar. Res. 1980, 38, 687–701. [Google Scholar]
- Ritchie, R.J. Consistent sets of spectrophotometric chlorophyll equations for acetone, methanol and ethanol solvents. Photosynth. Res. 2006, 89, 27–41. [Google Scholar] [CrossRef]
- Subramaniam1, A.; Carpenter, E.J.; Karentz, D.; Falkowski, P.G. Bio-optical properties of the marine diazotrophic cyanobacteria Trichodesmium spp. I. Absorption and photosynthetic action spectra. Limnol. Oceanogr. 1999, 44, 608–617. [Google Scholar] [CrossRef]
- Stramski, D.; Reynolds, R.A.; Kaczmarek, S.; Uitz, J.; Zheng, G. Correction of pathlength amplification in the filter-pad technique for measurements of particulate absorption coefficient in the visible spectral region. Appl. Opt. 2015, 54, 6763–6782. [Google Scholar] [CrossRef] [Green Version]
- Küpper, H.; Seibert, S.; Parameswaran, A. Fast, sensitive, and inexpensive alternative to analytical pigment HPLC: Quantification of chlorophylls and carotenoids in crude extracts by fitting with Gauss peak spectra. Anal. Chem. 2007, 79, 7611–7627. [Google Scholar] [CrossRef]
- Genty, B.; Briantais, J.-M.; Baker, N.R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. BBA-Gen. Subjects 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Eilers, P.; Peeters, J. A model for the relationship between light intensity and the rate of photosynthesis in phytoplankton. Ecol. Model. 1988, 42, 199–215. [Google Scholar] [CrossRef]
- Rodriguez, F.; Chauton, M.; Johnsen, G.; Andresen, K.; Olsen, L.; Zapata, M. Photoacclimation in phytoplankton: Implications for biomass estimates, pigment functionality and chemotaxonomy. Mar. Biol. 2006, 148, 963–971. [Google Scholar] [CrossRef]
- Marie-Rose Vandenhecke, J.; Bastedo, J.; Cockshutt, A.M.; Campbell, D.A.; Huot, Y. Changes in the Rubisco to photosystem ratio dominates photoacclimation across phytoplankton taxa. Photosynth. Res. 2015, 124, 275–291. [Google Scholar] [CrossRef]
- Andresen, E.; Lohscheider, J.; Šetlikova, E.; Adamska, I.; Šimek, M.; Küpper, H. Acclimation of Trichodesmium erythraeum ISM101 to high and low irradiance analysed on the physiological, biophysical and biochemical level. New Phytol. 2010, 185, 173–188. [Google Scholar] [CrossRef] [Green Version]
- Walker, B.J.; Strand, D.D.; Kramer, D.M.; Cousins, A.B. The response of cyclic electron flow around photosystem I to changes in photorespiration and nitrate assimilation. Plant Physiol. 2014, 165, 453–462. [Google Scholar] [CrossRef] [Green Version]
- Fujita, Y. A study on the dynamic features of photosystem stoichiometry: Accomplishments and problems for future studies. Photosynth. Res. 1997, 53, 83–93. [Google Scholar] [CrossRef]
- Xu, J.; Gao, K. Future CO2-induced ocean acidification mediates the physiological performance of a green tide alga. Plant Physiol. 2012, 160, 1762–1769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, G.D.; Badger, M.R.; Woodger, F.J.; Long, B.M. Advances in understanding the cyanobacterial CO2-concentrating-mechanism (CCM): Functional components, Ci transporters, diversity, genetic regulation and prospects for engineering into plants. J. Exp. Bot. 2008, 59, 1441–1461. [Google Scholar] [CrossRef] [PubMed]
- Hopkinson, B.M.; Dupont, C.L.; Allen, A.E.; Morel, F.M.M. Efficiency of the CO2-concentrating mechanism of diatoms. Proc. Natl. Acad. Sci. USA 2011, 108, 3830–3837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Gao, K.; Riebesell, U. CO2-induced seawater acidification affects physiological performance of the marine diatom Phaeodactylum tricornutum. Biogeosciences 2010, 7, 2915–2923. [Google Scholar] [CrossRef] [Green Version]
- Suffrian, K.; Schulz, K.G.; Gutowska, M.; Riebesell, U.; Bleich, M. Cellular pH measurements in Emiliania huxleyi reveal pronounced membrane proton permeability. New Phytol. 2011, 190, 595–608. [Google Scholar] [CrossRef] [Green Version]
- Flynn, K.J.; Blackford, J.C.; Baird, M.E.; Raven, J.A.; Clark, D.R.; Beardall, J.; Brownlee, C.; Fabian, H.; Wheeler, G.L. Changes in pH at the exterior surface of plankton with ocean acidification. Nat. Clim. Chang. 2012, 2, 510–513. [Google Scholar] [CrossRef]
- Raven, J.A.; Giordano, M.; Beardall, J.; Maberly, S.C. Algal evolution in relation to atmospheric CO2: Carboxylases, carbon-concentrating mechanisms and carbon oxidation cycles. Philos. Trans. R. Soc. B 2012, 367, 493–507. [Google Scholar] [CrossRef] [Green Version]
- Liu, N.; Beardall, J.; Gao, K. Elevated CO2 and associated seawater chemistry do not benefit a model diatom grown with increased availability of light. Aquat. Microb. Ecol. 2017, 79, 137–147. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.-Z.; Wang, T.; Huang, R.; Yi, X.; Zhang, D.; Beardall, J.; Hutchins, D.A.; Liu, X.; Wang, X.; Deng, Z.; et al. Enhancement of diatom growth and phytoplankton productivity with reduced O2 availability is moderated by rising CO2. Commun. Biol. 2022, 5, 54. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Growth | Carbon Fixation | |||
|---|---|---|---|---|
| AC | HC | AC | HC | |
| α | 0.0246 ± 0.0006 | 0.0234 ± 0.0017 | 0.0146 ± 0.0028 | 0.0295 ± 0.0013 |
| β | 0.0010 ± 0.0001 | 0.0011 ± 0.0001 | 0.0044 ± 0.0010 | 0.0025 ± 0.0006 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, H.; Beardall, J.; Gao, K. Photoinhibition of the Picophytoplankter Synechococcus Is Exacerbated by Ocean Acidification. Water 2023, 15, 1228. https://doi.org/10.3390/w15061228
Li H, Beardall J, Gao K. Photoinhibition of the Picophytoplankter Synechococcus Is Exacerbated by Ocean Acidification. Water. 2023; 15(6):1228. https://doi.org/10.3390/w15061228
Chicago/Turabian StyleLi, He, John Beardall, and Kunshan Gao. 2023. "Photoinhibition of the Picophytoplankter Synechococcus Is Exacerbated by Ocean Acidification" Water 15, no. 6: 1228. https://doi.org/10.3390/w15061228
APA StyleLi, H., Beardall, J., & Gao, K. (2023). Photoinhibition of the Picophytoplankter Synechococcus Is Exacerbated by Ocean Acidification. Water, 15(6), 1228. https://doi.org/10.3390/w15061228