
Recovery of a Population of Slimy Sculpin (Uranidea cognata) after an Autumn Fish Kill in the Headwaters of a Minnesota Trout Stream


Abstract
:1. Introduction
2. Methods
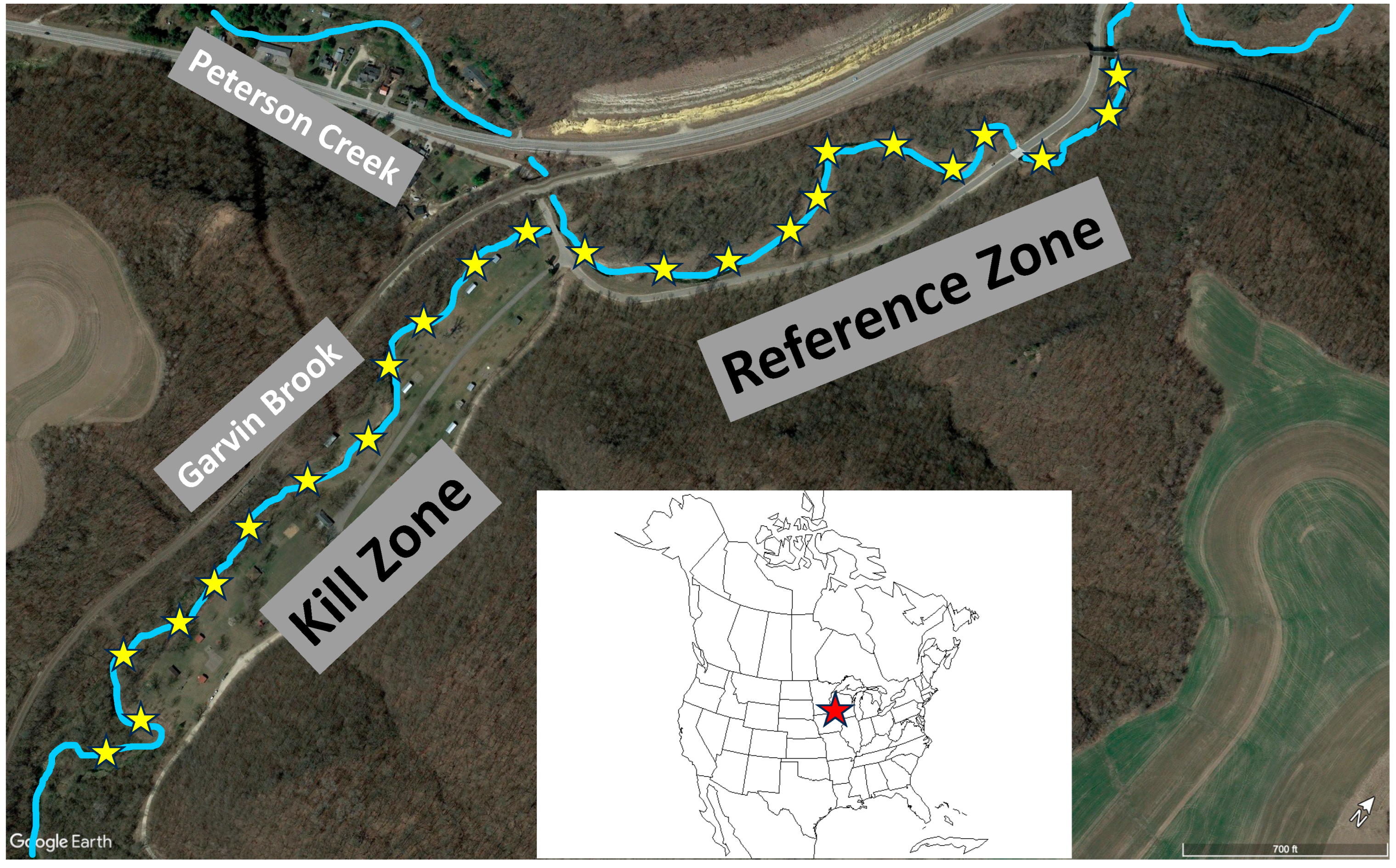
2.1. Study Site
2.2. Fieldwork
2.3. Data Analyses
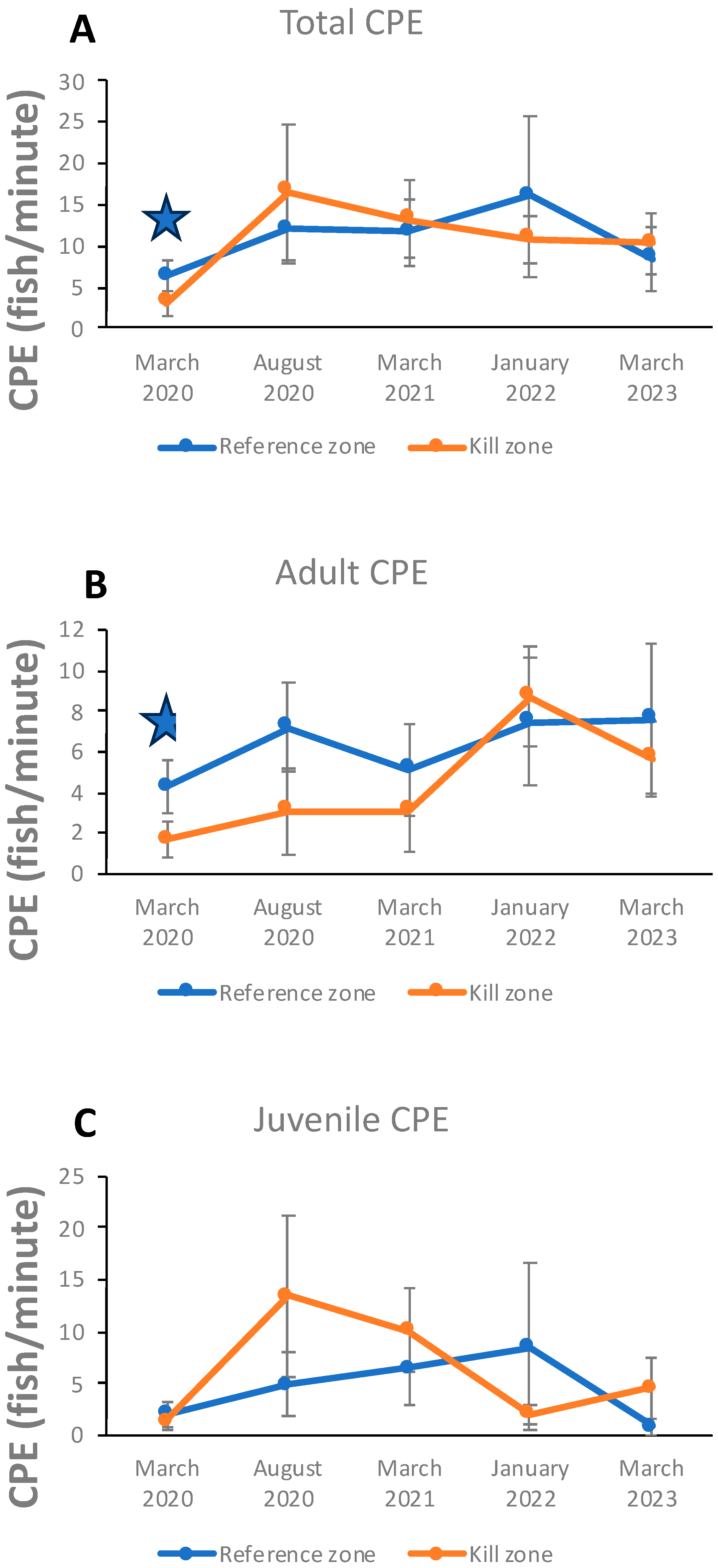
3. Results
3.1. Survey Effort
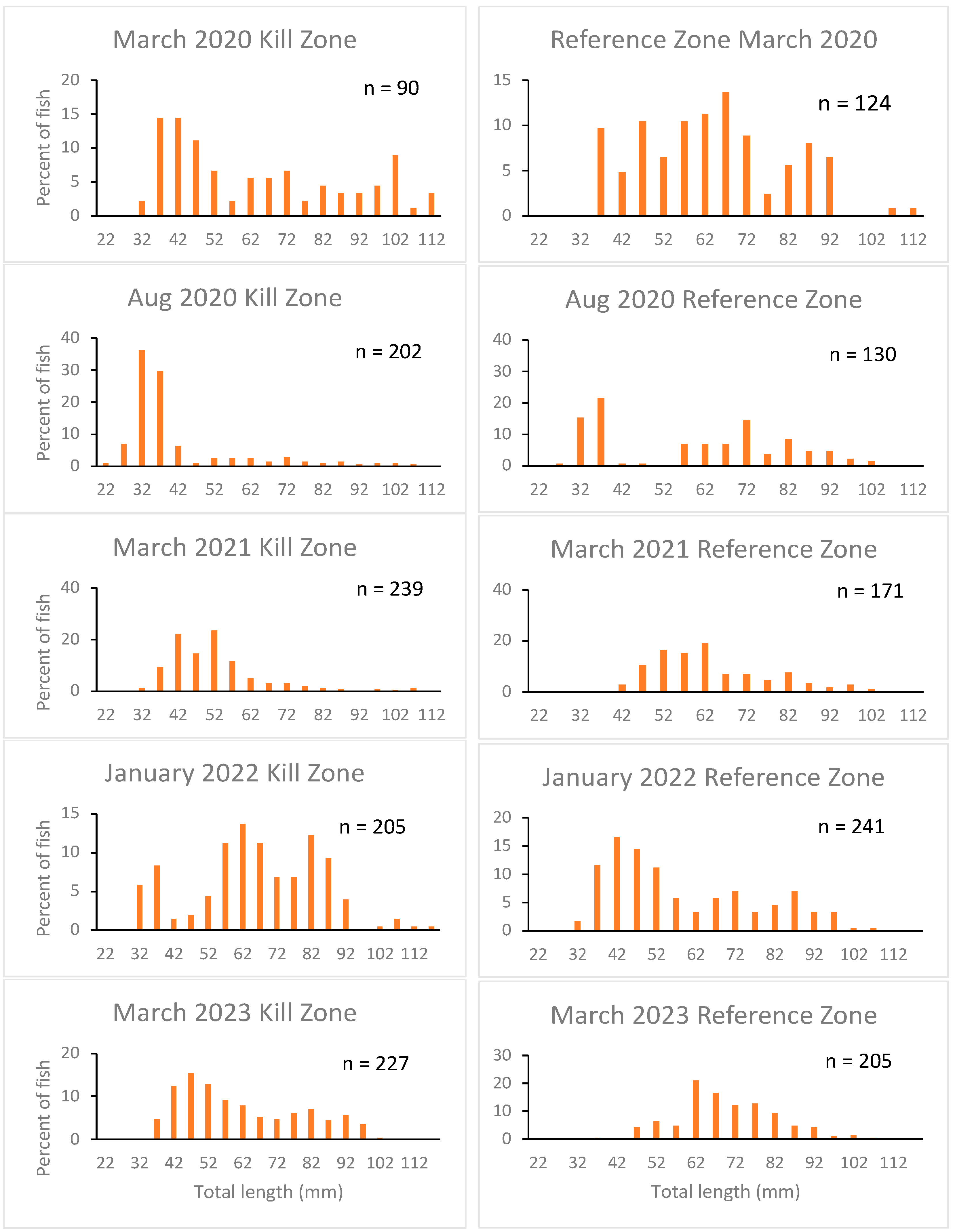
3.2. Sculpin Age Structure
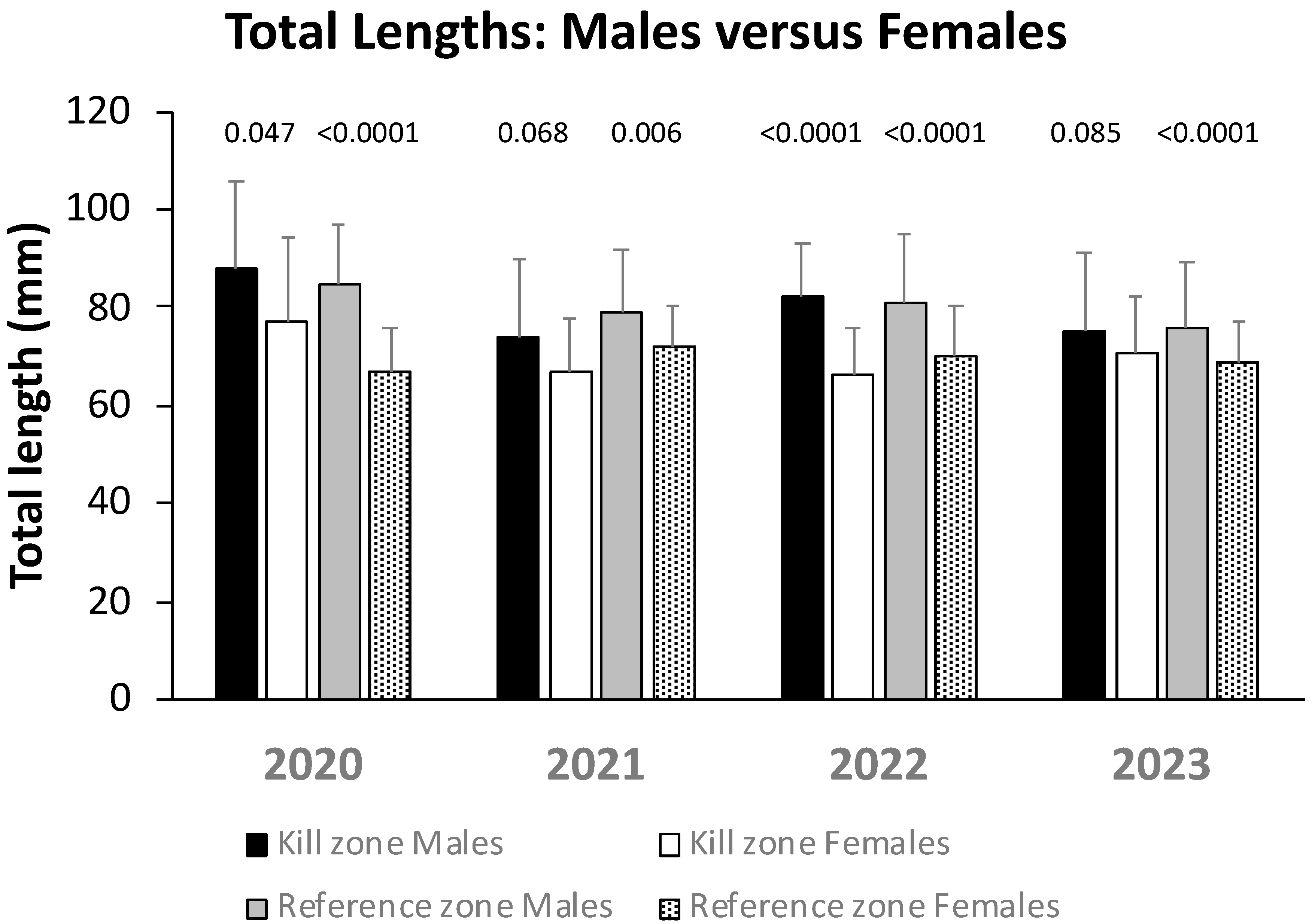
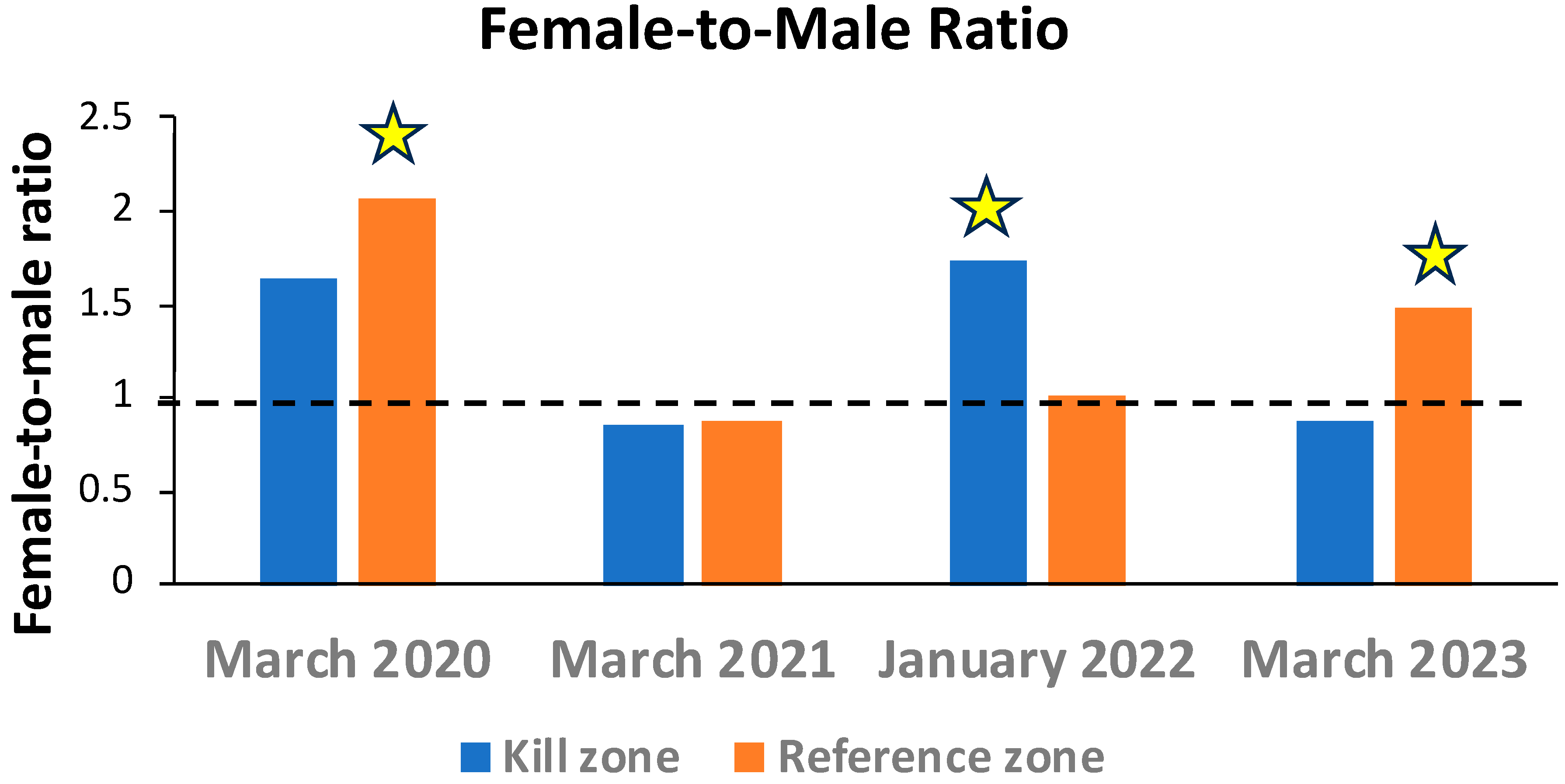
3.3. Adult Sculpin Sex Ratios and Size Differences
3.4. Sculpin CPE Relative Abundances
4. Discussion
5. Conclusions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Thronson, A.; Quigg, A. Fifty-five years of fish kills in coastal Texas. Estuaries Coasts 2008, 31, 802–813. [Google Scholar] [CrossRef]
- La, V.T.; Cooke, S.J. Advancing the science and practice of fish kill investigations. Rev. Fish. Sci. 2011, 19, 21–33. [Google Scholar] [CrossRef]
- Phelps, N.B.D.; Bueno, I.; Poo-Muñoz, D.A.; Knowles, S.J.; Massarani, S.; Rettkowski, R.; Shen, L.; Rantala, H.; Phelps, P.L.F.; Escobar, L.E. Retrospective and predictive investigation of fish kill events. J. Aquat. Anim. Health 2019, 31, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Meade, R. Fish and invertebrate recolonization in a Missouri prairie stream after an acute pollution event. N. Am. J. Fish. Manag. 2004, 24, 7–19. [Google Scholar] [CrossRef]
- Schnaser, A.C.; Mundahl, N.D. Recovery of a headwater stream population of brown trout after a fish kill in southeastern Minnesota, USA. Environ. Biol. Fish. 2022, 105, 1179–1192. [Google Scholar] [CrossRef]
- Hunt, L. South Branch Whitewater River Unified Fish Kill Response; Minnesota Department of Agriculture: Saint Paul, MN, USA, 2015; Available online: https://files.dnr.state.mn.us/areas/fisheries/lanesboro/unified-fish-kill-response12-22-15.pdf (accessed on 21 December 2021).
- Roell, M.J.; Orth, D.J. The roles of predation, competition, and exploitation in the trophic dynamics of a warmwater stream: A model synthesis, analysis, and application. Hydrobiologia 1994, 291, 157–178. [Google Scholar] [CrossRef]
- Fitzgerald, D.G.; Kott, E.; Lanno, R.P.; Dixon, D.G. A quarter century of change in the fish communities of three small streams modified by anthropogenic activities. J. Aquat. Ecosyst. Stress Recovery 1997, 6, 111–127. [Google Scholar] [CrossRef]
- Cooke, S.J.; Bunt, C.M.; Hamilton, S.J.; Jennings, C.A.; Pearson, M.P.; Cooperman, M.S.; Markle, D.F. Threats, conservation strategies, and prognosis for suckers (Catostomidae) in North America: Insights from regional case studies of a diverse family of non-game fishes. Biol. Conserv. 2005, 121, 317–331. [Google Scholar] [CrossRef]
- Peterson, J.T.; Bayley, A.B. Colonization rates of fishes in experimentally defaunated warmwater streams. Trans. Am. Fish. Soc. 1993, 122, 199–207. [Google Scholar] [CrossRef]
- Winona Daily News. DNR: Whitewater Trout Stream That Saw Fish Kill Is Recovering; Winona Daily News: Winona, MN, USA, 2016; Available online: https://www.winonadailynews.com/news/local/dnr-whitewater-trout-stream-that-saw-fish-kill-is-recovering/article_3bf19ec9-ec08-58d2-80f0-2bbd6be1a15c.html (accessed on 21 December 2021).
- Freeman, M.; Elkins, D.; Maholland, P.; Butler, Z.; Kleinhans, M.; Skaggs, J.; Stowe, E.; Straight, C.; Wegner, S. Slow recovery of headwater-stream fishes following a catastrophic poisoning event. J. Fish Wildl. Manag. 2021, 12, 362–372. [Google Scholar] [CrossRef]
- Mundahl, N.D.; Ingersoll, C.G. 1989 Home range, movements, and density of the central stoneroller, Campostoma anomalum, in a small Ohio stream. Environ. Biol. Fish. 1989, 24, 307–311. [Google Scholar] [CrossRef]
- Rasmussen, J.E.; Belk, M.C. Individual movement of stream fishes: Linking ecological drivers with evolutionary processes. Rev. Fish. Sci. Aquac. 2017, 25, 70–83. [Google Scholar] [CrossRef]
- De Fries, L.; Camana, M.; Hartz, S.M.; Becker, F.G. Heterogenous movement by a small non-migratory stream fish. Environ. Biol. Fish. 2022, 105, 1873–1885. [Google Scholar] [CrossRef]
- Mundahl, N.D.; Ingersoll, C.G. Early autumn movements and densities of johnny (Etheostoma nigrum) and fantail (E. flabellare) daters in a southwestern Ohio stream. Ohio J. Sci. 1983, 83, 103–108. [Google Scholar]
- Ingersoll, C.G.; Hlohowskyj, I.; Mundahl, N.D. Movements and densities of the darters Etheostoma flabellare, E. spectabile, and E. nigrum during spring spawning. J. Freshw. Ecol. 1984, 2, 345–351. [Google Scholar] [CrossRef]
- Keeler, R.A.; Breton, A.R.; Peterson, D.P.; Cunjak, R.A. Apparent survival and detection estimates for PIT-tagged slimy sculpin in five small New Brunswick streams. Trans. Am. Fish. Soc. 2007, 131, 281–292. [Google Scholar] [CrossRef]
- Gray, M.A.; Curry, R.A.; Arciszewski, T.J.; Munkittrick, K.R.; Brasfield, S.M. The biology and ecology of slimy sculpin: A recipe for effective environmental monitoring. Facets 2018, 3, 103–127. [Google Scholar] [CrossRef]
- Wallace, R.L.; McAllister, D.E.; Rankin, M. Cottus cognatus Richardson, slimy sculpin. In Atlas of North American Freshwater Fishes; Lee, D.S., Gilbert, C.R., Hocutt, C.H., Jenkins, R.E., McAllister, D.E., Stauffer, J.R., Jr., Eds.; North Carolina Biological Survey Publication #1980-12; North Carolina Museum of Natural Sciences: Raleigh, NC, USA, 1980; p. 808. [Google Scholar]
- Page, L.M.; Burr, B.M. A Field Guide to Freshwater Fishes: North America North of Mexico; Houghton Mifflin Company: Boston, MA, USA, 1991. [Google Scholar]
- Eddy, S.; Underhill, J.C. Northern Fishes, with Special Reference to the Upper Mississippi Valley; University of Minnesota Press: Minneapolis, MN, USA, 1974. [Google Scholar]
- Kelly, B.B.; Siepker, M.J.; Weber, M.J. 2023. Effects of non-native Salmo trutta and multiscale habitat factors on native fishes in the Driftless Area. Can. J. Fish. Aquat. Sci. 2023, 80, 942–957. [Google Scholar]
- Mundahl, N.D.; Thomas, K.N.; Mundahl, E.D. Selected habitats of slimy sculpin in coldwater tributaries of the Upper Mississippi River in Minnesota. Am. Midl. Nat. 2012, 168, 144–161. [Google Scholar] [CrossRef]
- Gray, M.A.; Munkittrick, K.R. An effects-based assessment of slimy sculpin (Cottus cognatus) populations in agricultural regions of northwestern New Brunswick. Water Qual. Res. J. Can. 2005, 40, 16–27. [Google Scholar] [CrossRef]
- Clarke, A.D.; Telmer, K.H.; Shrimpton, J.M. Movement patterns of fish revealed by otolith microchemistry: A comparison of putative migratory and resident species. Environ. Biol. Fish. 2015, 98, 1583–1597. [Google Scholar] [CrossRef]
- Weinstein, S.Y.; Coombs, J.A.; Nislow, K.H.; Riley, C.; Roy, A.H.; Whiteley, A.R. Evaluating the effects of barriers on slimy sculpin movement and population connectivity using novel sibship-based and traditional genetic metrics. Trans. Am. Fish. Soc. 2019, 148, 1117–1131. [Google Scholar] [CrossRef]
- Snook, V. Fish Kill Incident Report: Garvin Brook at Farmer’s Community Park; Minnesota Department of Natural Resources-Fisheries: Lanesboro, MN, USA, 2019. [Google Scholar]
- Burri, T.; Fier-Tucker, D.; Ganske, L.; Hunt, L.; Wieland, R. Response to 2019 Fish Kill on Garvin Brook, Winona County; Minnesota Pollution Control Agency: Saint Paul, MN, USA, 2020; Available online: https://www.pca.state.mn.us/sites/default/files/tdr-gl-22.pdf (accessed on 10 December 2023).
- Minnesota Department of Natural Resources. Garvin Brook Aquatic Management Area; Minnesota Department of Natural Resources: Saint Paul, MN, USA, 2023; Available online: https://www.dnr.state.mn.us/amas/detail_report.html?id=AMA00210 (accessed on 10 December 2023).
- Minnesota Pollution Control Agency. Agencies Conclude Investigation: Contaminated Runoff Suspected in Garvin Brook Fish Kill; Minnesota Pollution Control Agency: Saint Paul, MN, USA, 2020; Available online: https://pca.state.mn.us/news/agencies-conclude-investigation-contaminated-runoff-suspected-garvin-brook-fish-kill (accessed on 13 January 2022).
- Arciszewski, T.J.; Gray, M.; Munkittrick, K.; Baron, C. Guidance for the collection and sampling of slimy sculpin (Cottus cognatus) in northern Canadian lakes for environmental effects monitoring (EEM). In Canadian Technical Report of Fisheries and Aquatic Sciences 2909; Fisheries and Oceans Canada: Winnipeg, MB, Canada, 2010. [Google Scholar]
- Breen, M.J.; Ruetz, C.R., III; Thompson, K.J.; Kohler, S.L. Movements of mottled sculpins (Cottus bairdii) in a Michigan stream: How restricted are they? Can. J. Fish. Aquat. Sci. 2009, 66, 31–41. [Google Scholar] [CrossRef]
- Mousseau, T.A.; Collins, N.C.; Cabana, G. A comparative study of sexual selection and reproductive investment in the slimy sculpin, Cottus cognatus. Oikos 1987, 51, 156–162. [Google Scholar] [CrossRef]
- Majeski, M.J.; Cochran, P.A. Spawning season and habitat use of slimy sculpin (Cottus cognatus) in southeastern Minnesota. J. Freshw. Ecol. 2009, 24, 301–307. [Google Scholar] [CrossRef]
- Petty, T.D.; Grossman, G.D. Restricted movement by mottled sculpin (Pisces: Cottidae) in a southern Appalachian stream. Freshw. Biol. 2004, 49, 631–645. [Google Scholar] [CrossRef]
- Schmetterling, D.A.; Adams, S.B. Summer movements within the fish community of a small montane stream. N. Am. J. Fish. Manag. 2004, 24, 1163–1172. [Google Scholar] [CrossRef]
- Larimore, R.W.; Childers, W.F.; Heckrotte, C. Destruction and re-establishment of stream fish and invertebrates affected by drought. Trans. Am. Fish. Soc. 1959, 88, 261–285. [Google Scholar] [CrossRef]
- Harrel, R.C.; Davis, B.J.; Dorris, T.C. Stream order and species diversity of fishes in an intermittent Oklahoma stream. Am. Midl. Nat. 1967, 78, 428–436. [Google Scholar] [CrossRef]
- Minnesota Department of Natural Resources. Southeast Minnesota Sculpin Reintroduction Plan; Minnesota Department of Natural Resources–Fisheries: Lanesboro, MN, USA, 2003; Available online: https://files.dnr.state.mn.us/areas/fisheries/lakecity/semn_sculpin_reintroplan.pdf (accessed on 10 December 2023).
- Huff, D.D.; Miller, L.M.; Vondracek, B. Patterns of ancestry and genetic diversity in reintroduced populations of the slimy sculpin: Implications for conservation. Conserv. Genet. 2010, 11, 2379–2391. [Google Scholar] [CrossRef]
- Huff, D.D.; Miller, L.M.; Chizinski, C.J.; Vondracek, B. Mixed-source reintroductions lead to outbreeding depression in second-generation descendants of a native North American fish. Mol. Ecol. 2011, 20, 4246–4258. [Google Scholar] [CrossRef] [PubMed]
- Mundahl, N.D.; Mundahl, D.E.; Merten, E.C. Success of slimy sculpin introductions: Influence of feeding and diets. Am. Midl. Nat. 2012, 168, 162–183. [Google Scholar] [CrossRef]
- Nerbonne, B. Personal Communication; Minnesota Department of Natural Resources–Fisheries: Saint Paul, MN, USA, 2023. [Google Scholar]
- Detenbeck, N.; DeVore, P.W.; Niemi, G.J.; Lima, A. Recovery of temperate-stream fish communities from disturbance: A review of case studies and synthesis of theory. Environ. Manag. 1992, 16, 33–53. [Google Scholar] [CrossRef]
- Anderson, C.S. The structure of sculpin populations along a stream size gradient. Environ. Biol. Fish. 1985, 13, 93–102. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Date | Kill Zone | N | Reference | N | t-Value | p |
|---|---|---|---|---|---|---|
| Mar-20 | 64 (24) | 90 | 64 (17) | 124 | 0.05 | 0.959 |
| Aug-20 | 41 (17) | 202 | 59 (22) | 130 | 7.97 | <0.001 |
| Mar-21 | 52 (13) | 239 | 63 (14) | 171 | 8.55 | <0.001 |
| Jan-22 | 66 (18) | 205 | 58 (19) | 241 | 4.39 | <0.001 |
| Mar-23 | 70 (13) | 227 | 61 (18) | 205 | 5.66 | <0.001 |
| Kill Zone | Reference Zone | |||||
|---|---|---|---|---|---|---|
| Date | Males | Females | Males | Females | X2 | p |
| Mar-20 | 19 | 31 | 27 | 56 | 0.43 | 0.521 |
| Mar-21 | 29 | 25 | 40 | 35 | 0.002 | 0.967 |
| Jan-22 | 68 | 99 | 54 | 55 | 2.08 | 0.149 |
| Mar-23 | 69 | 60 | 74 | 110 | 5.38 | 0.02 |
| Total CPE | Adult CPE | Juvenile CPE | ||||
|---|---|---|---|---|---|---|
| Date | t | p | t | p | t | p |
| Mar-20 | 4.49 | 0.0002 | 5.48 | <0.0001 | 1.54 | 0.14 |
| Aug-20 | 1.66 | 0.111 | 4.65 | 0.0001 | 3.56 | 0.002 |
| Mar-21 | 0.9 | 0.375 | 2.32 | 0.03 | 2.42 | 0.024 |
| Jan-22 | 1.79 | 0.097 | 1.07 | 0.298 | 2.77 | 0.018 |
| Mar-23 | 1.18 | 0.252 | 1.57 | 0.136 | 4.27 | <0.0001 |
| Zone/Statistic | Total CPE | Adult CPE | Juvenile CPE |
|---|---|---|---|
| Kill zone | |||
| Slope | −0.226 | −0.136 | −0.089 |
| Y intercept | 4.5 | 2.524 | 1.976 |
| r2 | 0.276 | 0.295 | 0.141 |
| t-value (one-tailed) | 1.95 | 2.05 | 1.28 |
| Degrees of freedom | 10 | 10 | 10 |
| p-value | 0.04 | 0.034 | 0.115 |
| Reference zone | |||
| Slope | 0.356 | 0.162 | 0.194 |
| Y intercept | 4.329 | 3.334 | 0.995 |
| r2 | 0.339 | 0.141 | 0.249 |
| t-value (one-tailed) | 2.03 | 1.25 | 1.63 |
| Degrees of freedom | 8 | 8 | 8 |
| p-value | 0.038 | 0.142 | 0.071 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mundahl, N.D. Recovery of a Population of Slimy Sculpin (Uranidea cognata) after an Autumn Fish Kill in the Headwaters of a Minnesota Trout Stream. Water 2024, 16, 283. https://doi.org/10.3390/w16020283
Mundahl ND. Recovery of a Population of Slimy Sculpin (Uranidea cognata) after an Autumn Fish Kill in the Headwaters of a Minnesota Trout Stream. Water. 2024; 16(2):283. https://doi.org/10.3390/w16020283
Chicago/Turabian StyleMundahl, Neal D. 2024. "Recovery of a Population of Slimy Sculpin (Uranidea cognata) after an Autumn Fish Kill in the Headwaters of a Minnesota Trout Stream" Water 16, no. 2: 283. https://doi.org/10.3390/w16020283
APA StyleMundahl, N. D. (2024). Recovery of a Population of Slimy Sculpin (Uranidea cognata) after an Autumn Fish Kill in the Headwaters of a Minnesota Trout Stream. Water, 16(2), 283. https://doi.org/10.3390/w16020283