
Roles of N:P Ratios on Trophic Structures and Ecological Stream Health in Lotic Ecosystems

Abstract
:1. Introduction
2. Materials and Methods
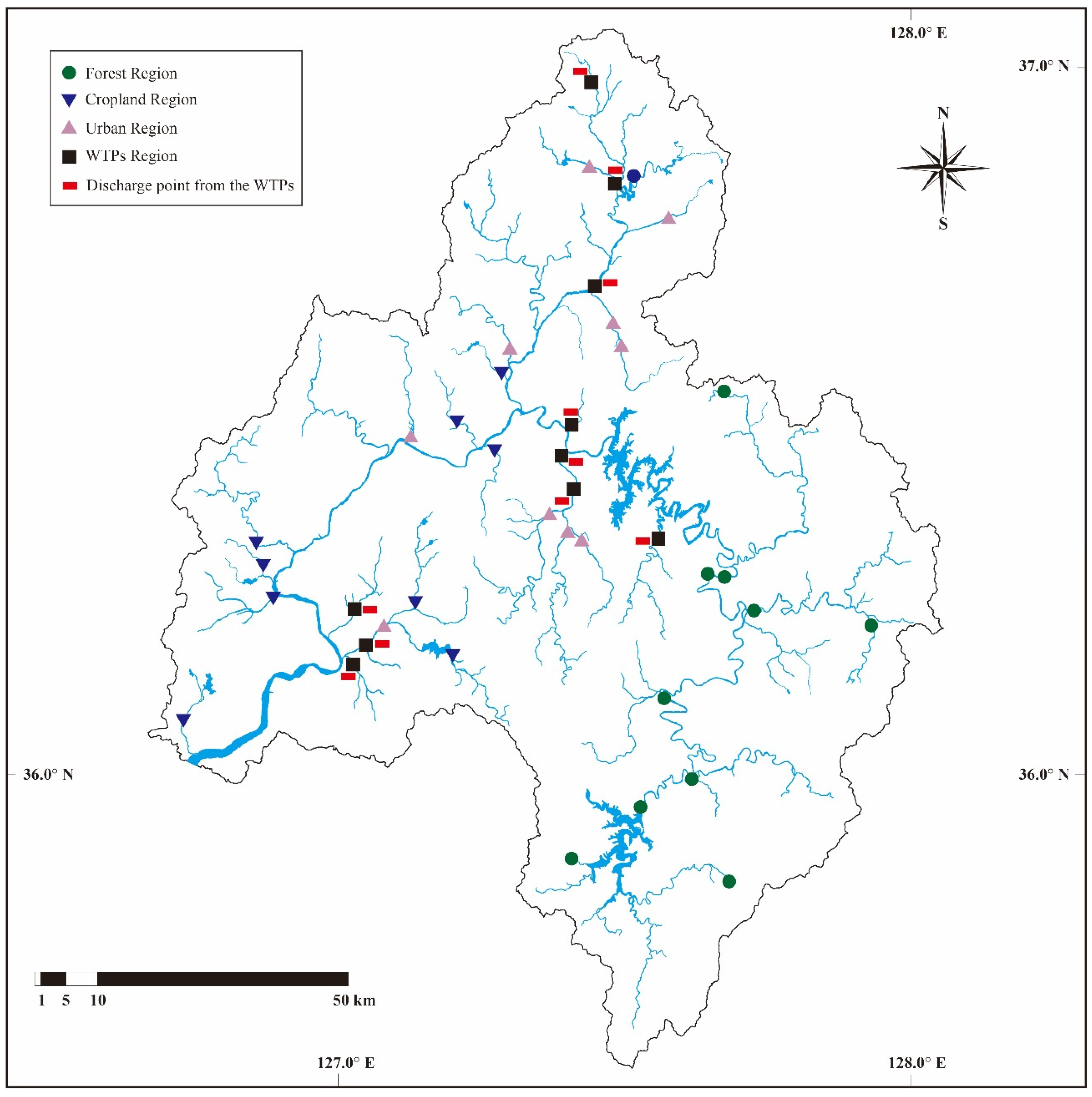
2.1. Study Area and Selection of Sampling Site

2.2. Analysis of Water Quality
2.3. Analysis of Physical Habitat Conditions
2.4. Fish Collection and Sampling Method
2.5. Analysis of Trophic Composition and Tolerance Level
2.6. Multi-Metric Fish Index of Biological Integrity (IBI) Model
2.7. Statistical Analysis
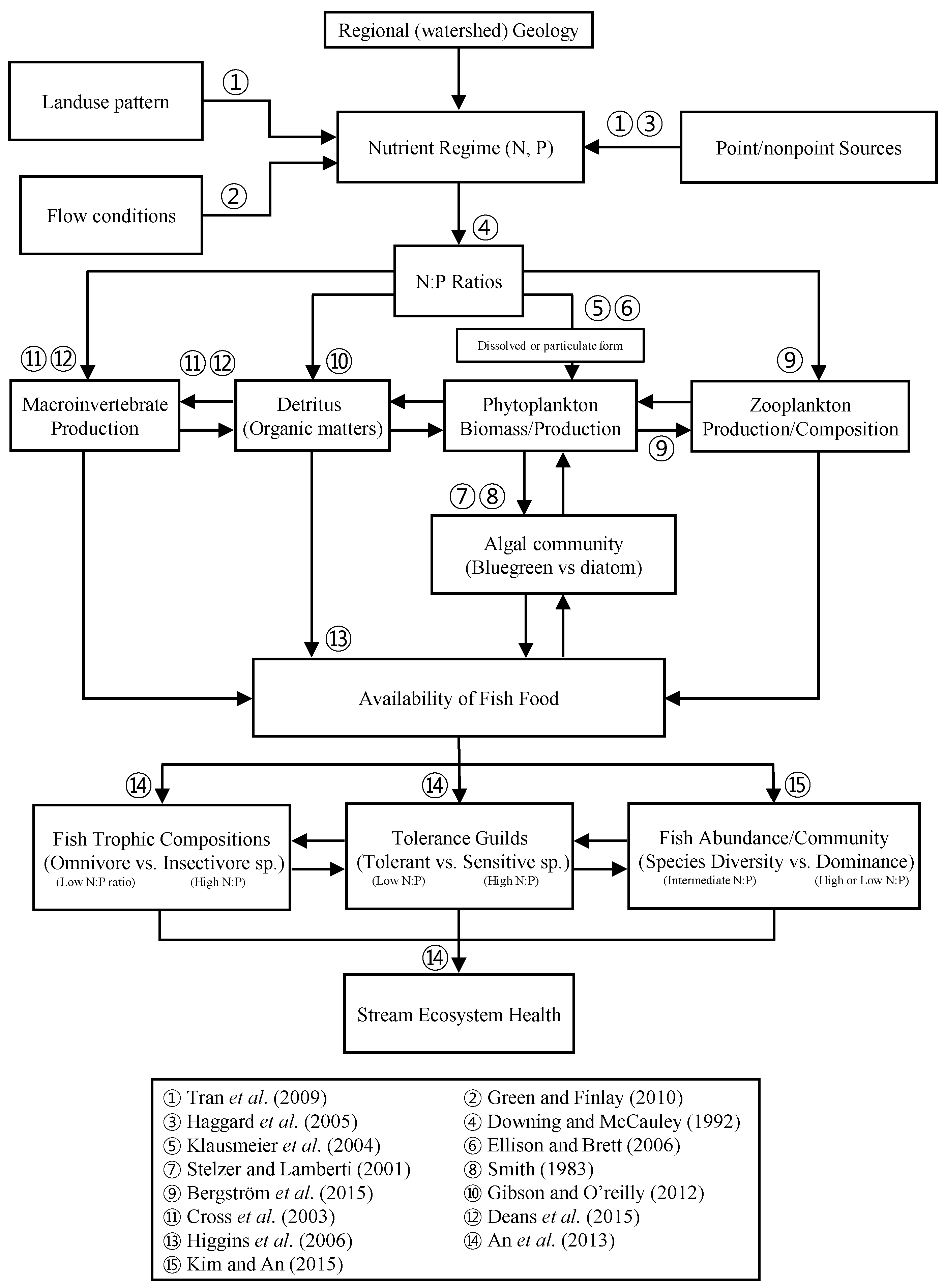
3. Results and Discussion
3.1. Influence of Land Use on Water Chemistry, Habitat Conditions, and Biological Components

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Forest Region | Cropland Region | Urban Region | WTPs Region |
|---|---|---|---|---|
| Mean ± SE | Mean ± SE | Mean ± SE | Mean ± SE | |
| Range | Range | Range | Range | |
| Total nitrogen (μg·L−1) | 1821 ± 33 a | 1740 ± 52 a | 2605 ± 65 b | 6792 ± 225 c |
| (1130–2836) | (545–3454) | (1002–5024) | (1713–14782) | |
| Total phosphorus (μg·L−1) | 26 ± 2 a | 67 ± 4 a, b | 85 ± 3 b | 462 ± 27 c |
| (4–76) | (11–164) | (21–212) | (66–1698) | |
| N:P ratios in ambient water | 143 ± 8 c | 53 ± 4 b | 46 ± 3 b | 24 ± 1 a |
| (29–388) | (5–175) | (10–110) | (5–58) | |
| Electrical Conductivity (μs·cm−1) | 136 ± 2 a | 210 ± 4 b | 263 ± 4 c | 482 ± 13 d |
| (74–212) | (133–342) | (148–383) | (237–935) | |
| BOD5 (mg·L−1) | 0.9 ± 0.02 a | 1.9 ± 0.08 b | 2.5 ± 0.10 c | 4.1 ± 0.16 d |
| (0.4–1.4) | (0.5–4.4) | (0.9–5.5) | (1.1–9.6) | |
| QHEI | 90 ± 5 b | 67 ± 5 a | 61 ± 5 a | 72 ± 6 a,b |
| (70–108) | (45–87) | (40–82) | (47–92) | |
| Sestonic CHL (μg·L−1) | 2.4 ± 0.2 a | 3.4 ± 0.3 a | 7.0 ± 0.7 a | 18.9 ± 2.4 b |
| (0.2–7.9) | (0.1–10.5) | (0.2–30.7) | (0.1–182.4) | |
| Sestonic CHL:TP | 0.15 ± 0.010 b | 0.08 ± 0.007 a | 0.10 ± 0.008 a | 0.08 ± 0.009 a |
| (0.00877–0.44200) | (0.00095–0.27789) | (0.00313–0.42713) | (0.00035–0.44106) | |
| Sestonic CHL:TN | 0.0015 ± 0.0001 a | 0.0025 ± 0.0003 a,b | 0.0039 ± 0.0005 b | 0.0039 ± 0.0004 b |
| (0.00006–0.00485) | (0.00004–0.01016) | (0.00006–0.01919) | (0.00002–0.01967) | |
| Index of biotic integrity | 32 ± 0.7 c | 23 ± 0.6 b | 19 ± 0.5 a | 20 ± 0.6 a |
| (24–38) | (16–30) | (12–24) | (14–26) |
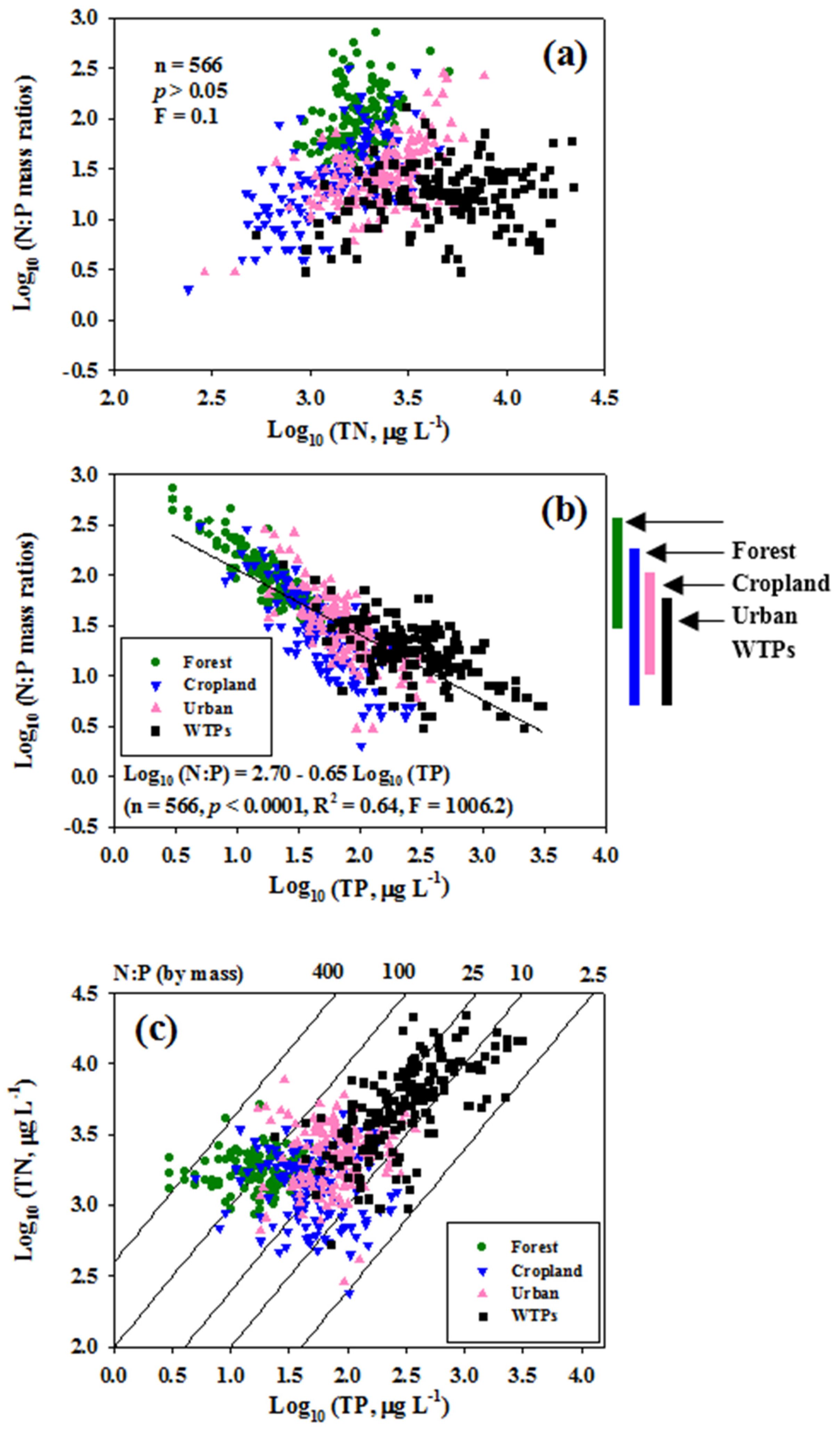
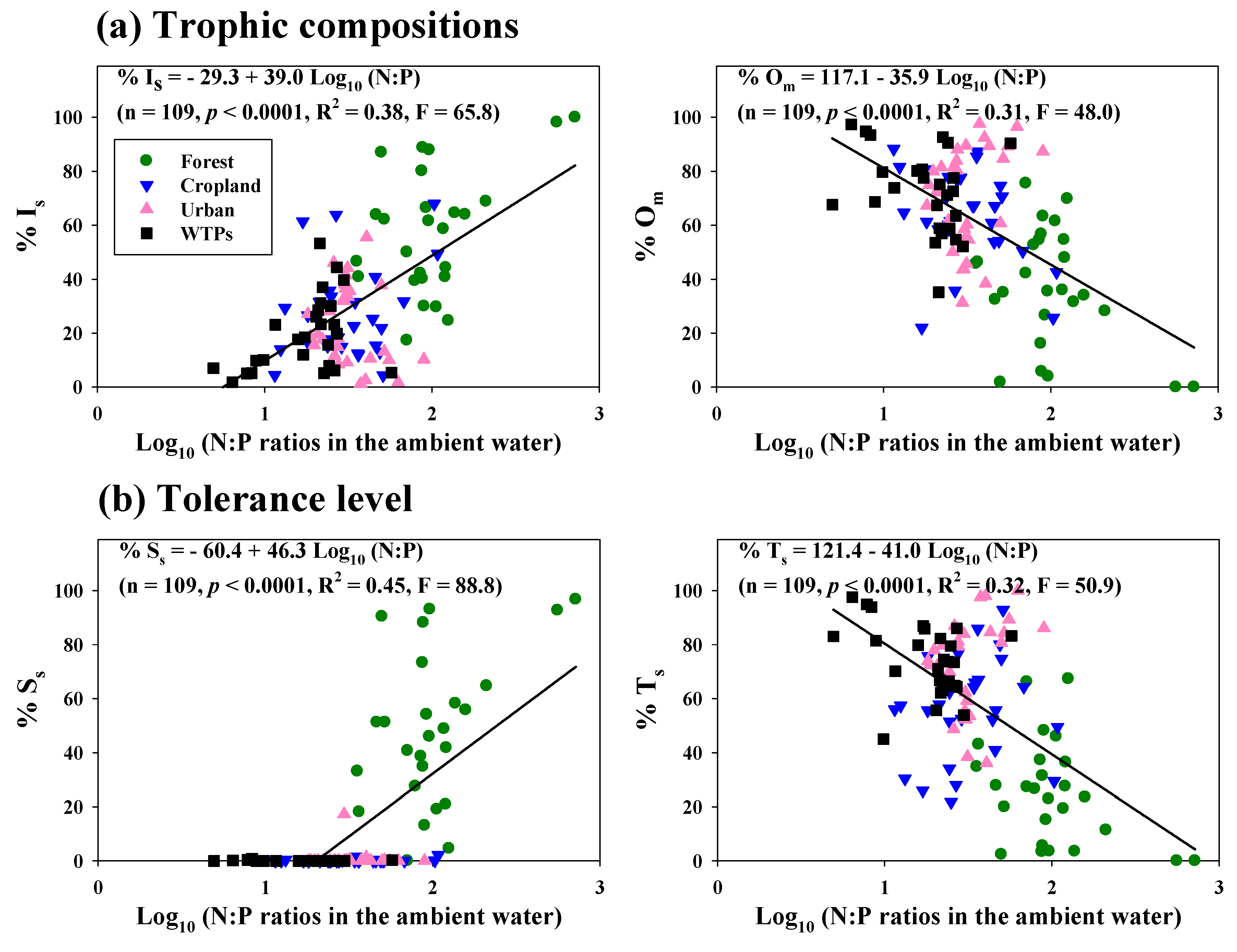
3.2. Effect of Mass Ratios of N:P on Nutrient Regimes and Their Influence on Land Use

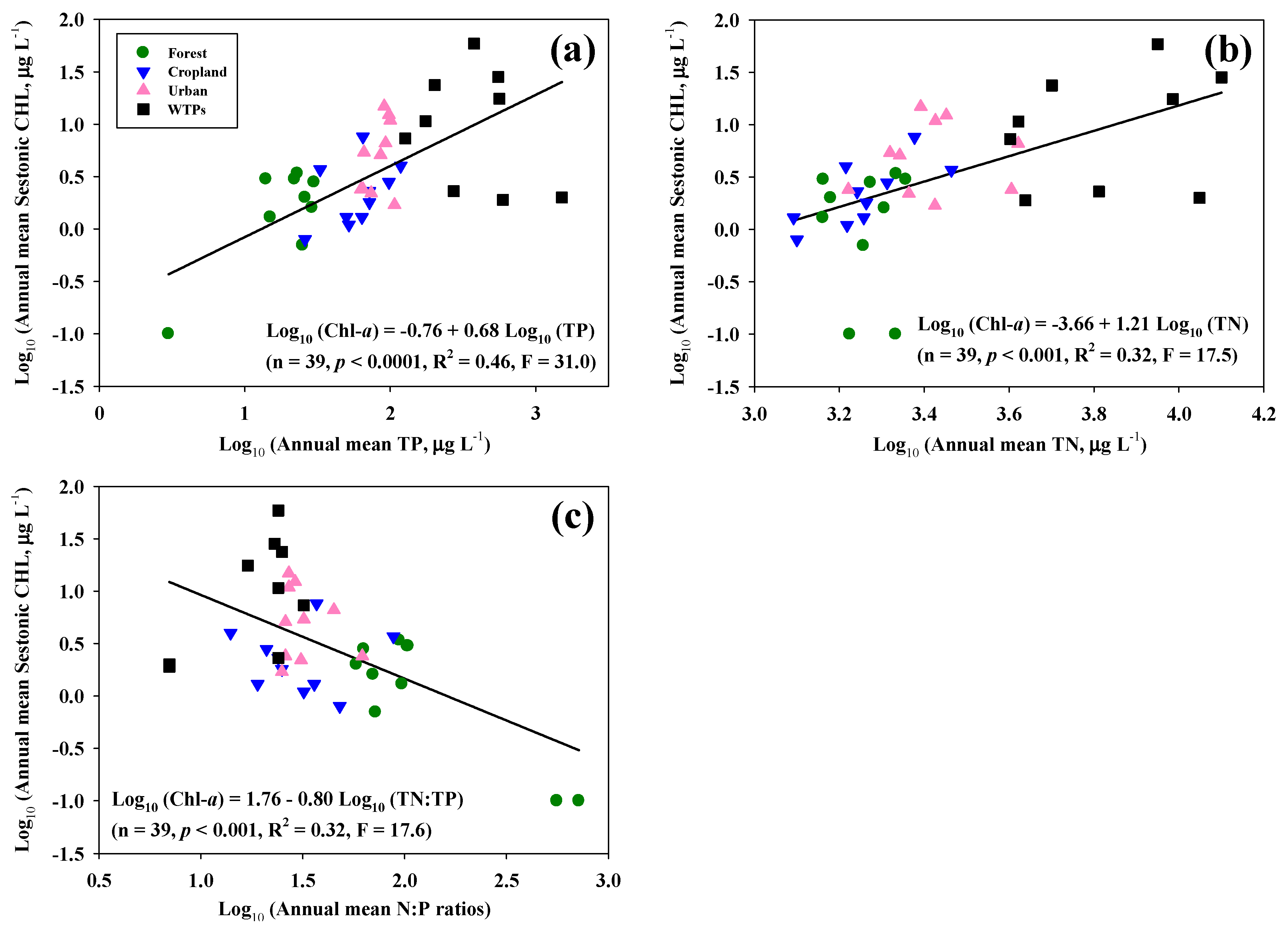
3.3. Effects of Nutrients and N:P Ratios on Sestonic CHL

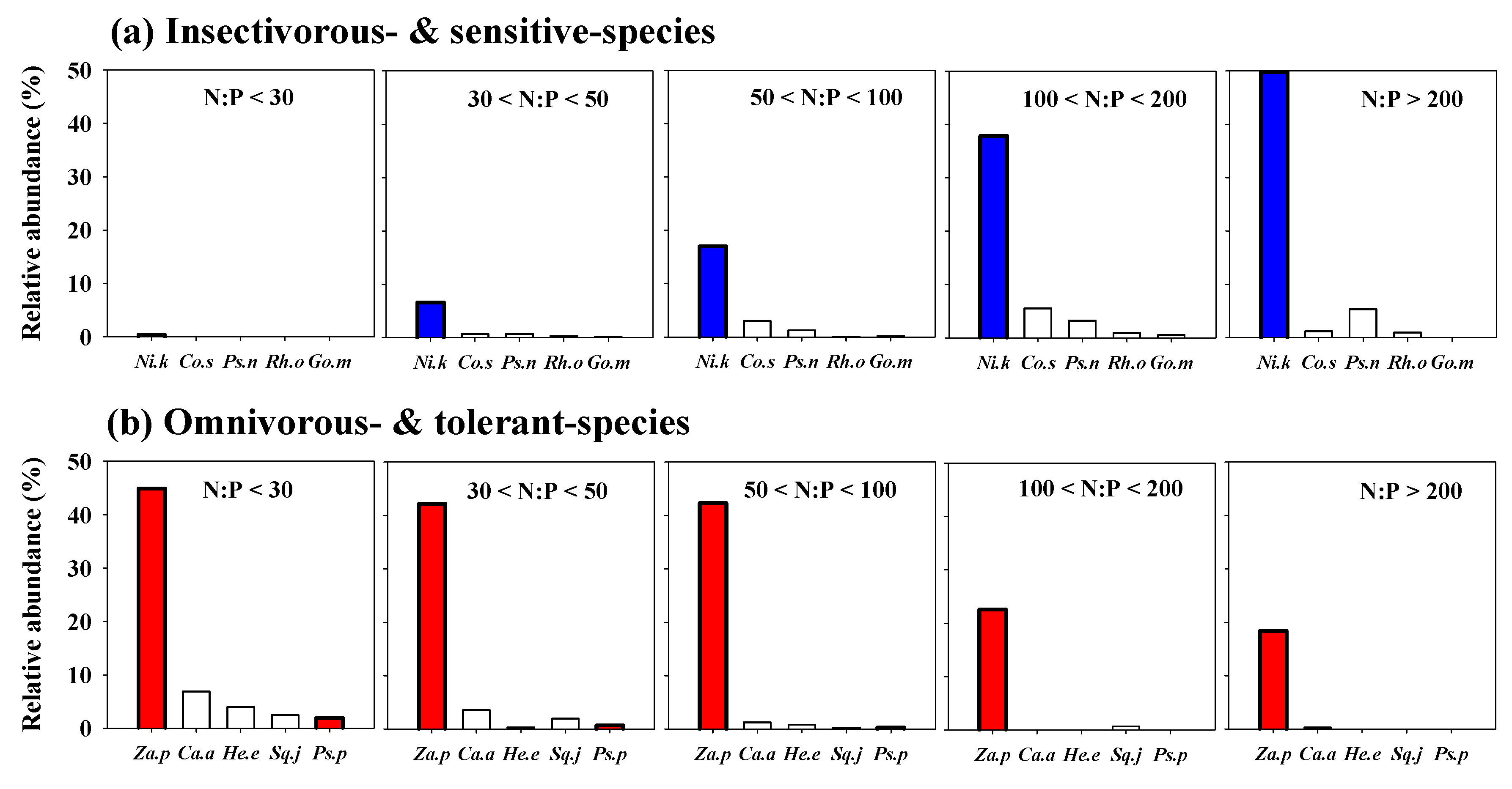
3.4. Relationships between Fish Communities and Compositions and N:P Ratios
| Type of Land | N:P Ratios in the Ambient Water | Community Group | Fish Species | Tolerance Level | Trophic Compositions | RA (%) |
|---|---|---|---|---|---|---|
| Forest Region | 143 ± 8 | Nipponocypris-Zacco community | Nipponocypris koreanus | Ss | Is | 34.5 |
| Zacco platypus | Ts | Om | 24.1 | |||
| Tanakia koreensis | In | Om | 6.0 | |||
| Coreoleuciscus splendidus | Ss | Is | 4.8 | |||
| Pungtungia herzi | In | Is | 4.2 | |||
| Other species (43) | – | – | 26.4 | |||
| Cropland region | 53 ± 4 | Zacco-Tridentiger community | Zacco platypus | Ts | Om | 38.0 |
| Tridentiger brevispinis | In | Is | 13.8 | |||
| Pseudogobio esocinus | In | Is | 6.1 | |||
| Tanakia lanceolatus | Int | Om | 5.1 | |||
| Opsarichthys uncirostris amurensis | Ts | Ca | 3.3 | |||
| Other species (51) | – | – | 33.8 | |||
| Urban Region | 46 ± 3 | Zacco-Carassius community | Zacco platypus | Ts | Om | 52.5 |
| Carassius auratus | Ts | Om | 9.5 | |||
| Pseudogobio esocinus | In | Is | 6.8 | |||
| Tanakia lanceolatus | In | Om | 5.7 | |||
| Opsarichthys uncirostris amurensis | Ts | Ca | 2.9 | |||
| Other species (41) | – | – | 22.5 | |||
| WTPs region | 24 ± 1 | Zacco-Hemiculter community | Zacco platypus | Ts | Om | 51.1 |
| Hemiculter eigenmanni | Ts | Om | 6.2 | |||
| Pseudogobio esocinus | In | Is | 6.1 | |||
| Carassius auratus | Ts | Om | 5.7 | |||
| Hemibarbus labeo | Ts | Is | 5.0 | |||
| Other species (46) | – | – | 25.9 |

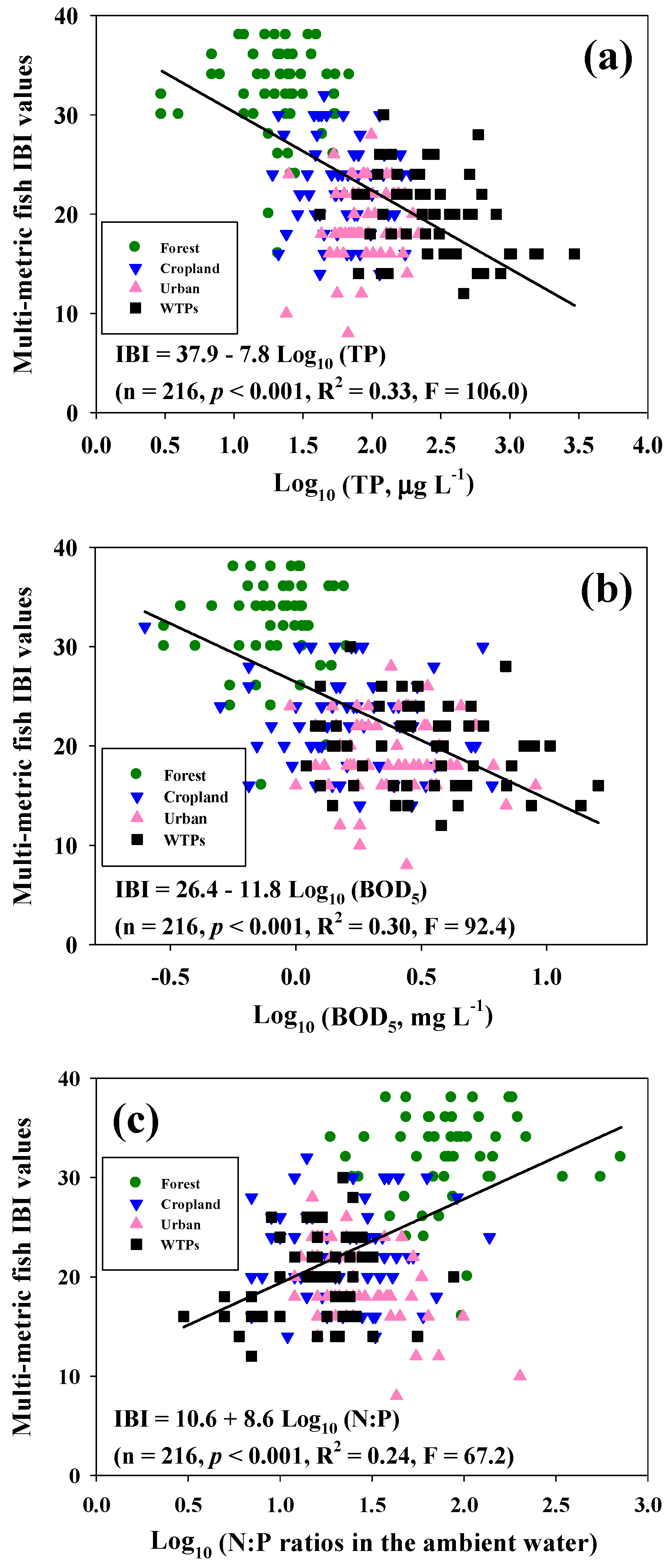
3.5. Multi-Metric IBI Model and Its Chemical Effects

| Category | Model Metric | Scoring Criteria | Forest | Cropland | Urban | WTPs | F-Value | p-Value | ||
|---|---|---|---|---|---|---|---|---|---|---|
| 5 | 3 | 1 | ||||||||
| Species richness and composition | M1: total number of native fish species | Expectations of M1 vary with stream order | 12.2 ± 4.4 b | 10.1 ± 3.2 a | 9.7 ± 3.2 a | 9.8 ± 4.6 a | 8.5 | *** | ||
| (3.8 ± 1.1) | (4.0 ± 1.2) | (3.4 ± 1.2) | (3.3 ± 1.5) | |||||||
| M2: number of riffle benthic dwelling species | Expectations of M2 vary with stream order | 2.7 ± 1.5 b | 1.2 ± 1.1 a | 0.8 ± 0.8 a | 0.9 ± 1.1 a | 38.3 | *** | |||
| (3.6 ± 1.6) | (1.7 ± 1.3) | (1.2 ± 0.6) | (1.6 ± 1.3) | |||||||
| M3: number of sensitive species | Expectations of M3 vary with stream order | 3.9 ± 1.6 b | 0.1 ± 0.3 a | 0.1 ± 0.3 a | 0.1 ± 0.4 a | 308.3 | *** | |||
| (3.7 ± 1.2) | (1.0 ± 0.0) | (1.0 ± 0.0) | (1.0 ± 0.0) | |||||||
| M4: proportion of individuals as tolerant species | <5 | 5–20 | >20 | 24.2 ± 21.4 a | 56.3 ± 21.5 b | 72.3 ± 19.6 c | 74.6 ± 18.0 c | 62.7 | *** | |
| (4.2 ± 1.1) | (2.6 ± 1.3) | (1.9 ± 1.1) | (1.8 ± 1.1) | |||||||
| Trophic composition | M5: proportion of individuals as omnivore species | <20 | 20–45 | >45 | 35.7 ± 24.3 a | 63.7 ± 19.6 b | 70.4 ± 22.8 b | 72.5 ± 20.1 b | 26.9 | *** |
| (3.6 ± 1.3) | (2.3 ± 1.2) | (2.0 ± 1.3) | (1.8 ± 1.1) | |||||||
| M6: proportion of individuals as native insectivore species | >45 | 20–45 | <20 | 59.1 ± 25.2 b | 27.2 ± 18.3 a | 23.2 ± 18.9 a | 19.5 ± 17.0 a | 36.4 | *** | |
| (4.2 ± 1.2) | (2.5 ± 1.4) | (2.3 ± 1.5) | (1.9 ± 1.4) | |||||||
| Fish abundance and condition | M7: total number of native individuals | Expectations of M7 vary with stream order | 191.1 ± 86.3 | 165.2 ± 130.3 | 170.7 ± 131.4 | 194.3 ± 146.5 | 0.9 | NS | ||
| (4.3 ± 1.0) | (4.1 ± 1.4) | (3.7 ± 1.6) | (4 ± 1.4) | |||||||
| M8: proportion of individuals as abnormalities | 0 | 0–1 | >1 | 0.0 ± 0.0 a | 0.3 ± 0.8 a | 1.2 ± 2.6 b | 0.5 ± 1.8 a,b | 5.1 | ** | |
| (5.0 ± 0.0) | (4.4 ± 1.4) | (3.7 ± 1.7) | (4.3 ± 1.2) | |||||||
| Scores (criteria of multi-metric fish IBI model) | 32.3 ± 4.6 c | 22.6 ± 4.7 b | 19.2 ± 3.9 a | 19.7 ± 4.1 a | 99.2 | *** | ||||
| Fair-excellent | Poor-fair | Poor-fair | Poor-fair | |||||||
3.6. Influence of Sestonic CHL on Trophic Compositions and River Health

3.7. Relationships among Lotic N:P Ratios, Trophic Composition and Tolerance Level

3.8. Influence of Land Use Pattern on N:P Ratios and Linkage with Trophic Compositions and Fish Tolerance
3.9. Influence of N:P Ratios on Fish Species Indicators


4. Conclusions

Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hecky, R.E.; Kilham, P. Nutrient limitation of phytoplankton in freshwater and marine environments: A review of recent evidence on the effects of enrichment. Limnol. Oceanogr. 1988, 33, 796–822. [Google Scholar] [CrossRef]
- Elser, J.J.; Marzolf, E.R.; Goldman, C.R. Phosphorus and nitrogen limitation of phytoplankton growth in the freshwaters of North America: A review and critique of experimental enrichments. Can. J. Fish. Aquat. Sci. 1990, 47, 1468–1477. [Google Scholar] [CrossRef]
- Vollenweider, R.A. Advances in defining critical loading levels for phosphorus in lake eutrophication. Mem. 1st. Ital. Idrobiol. 1976, 33, 53–83. [Google Scholar]
- Redfield, A.C. On the proportions of organic derivations in sea water and their relation to the composition of plankton. In James Johnstone Memorial Volume, 1st ed.; Daniel, R.J., Ed.; University Press of Liverpool: Liverpool, UK, 1934; pp. 177–192. [Google Scholar]
- Smith, V.H. Low nitrogen to phosphorus ratios favor dominance by blue-green algae in lake phytoplankton. J. Am. Water Resour. Assoc. 1983, 221, 669–671. [Google Scholar] [CrossRef] [PubMed]
- Frost, P.C.; Tank, S.E.; Turner, M.A.; Elser, J.J. Elemental composition of littoral invertebrates from oligotrophic and eutrophic Canadian lakes. J. N. Am. Benthol. Soc. 2003, 22, 51–62. [Google Scholar] [CrossRef]
- Cross, W.F.; Benstead, J.P.; Rosemond, A.D.; Bruce Wallace, J. Consumer-resource stoichiometry in detritus-based streams. Ecol. Lett. 2003, 6, 721–732. [Google Scholar] [CrossRef]
- An, K.-G.; Choi, J.-W.; Lee, Y.-J. Modifications of ecological trophic structures on chemical gradients in lotic ecosystems and their relations to stream ecosystem health. Anim. Cells Syst. 2013, 17, 53–62. [Google Scholar] [CrossRef]
- Kim, S-.Y.; An, K-.G. Nutrient regime, N:P ratios and suspended solids as key factors influencing fish tolerance, trophic compositions, and stream ecosystem health. J. Ecol. Environ. 2015, 38, 505–515. [Google Scholar]
- Guildford, S.J.; Hecky, R.E. Total nitrogen, total phosphorus, and nutrient limitation in lakes and oceans: Is there a common relationship? Limnol. Oceanogr. 2000, 45, 1213–1223. [Google Scholar] [CrossRef]
- Choi, J.-W.; Han, J.-H.; Park, C.-S.; Ko, D.-G.; Kang, H.-I.; Kim, J.Y.; Yun, Y.-J.; Kwon, H.-H.; An, K.-G. Nutrients and sestonic chlorophyll dynamics in Asian lotic ecosystems and ecological stream health in relation to land-use patterns and water chemistry. Ecol. Eng. 2015, 79, 15–31. [Google Scholar] [CrossRef]
- Stelzer, R.S.; Lamberti, G.A. Effects of N: P ratio and total nutrient concentration on stream periphyton community structure, biomass, and elemental composition. Limnol. Oceanogr. 2001, 46, 356–367. [Google Scholar] [CrossRef]
- Perkins, B.D.; Lohman, K.; van Nieuwenhuyse, E.; Jones, J.R. An examination of land cover and stream water quality among physiographic provinces of Missouri, U.S.A. Verh. Int. Verein. Limnol. 1998, 26, 940–947. [Google Scholar]
- Tran, C.P.; Bode, R.W.; Smith, A.J.; Kleppel, G.S. Land-use proximity as a basis for assessing stream water quality in New York State (USA). Ecol. Indic. 2010, 10, 727–733. [Google Scholar] [CrossRef]
- Loehr, R.C. Characteristics and comparative magnitude of non-point sources. J. Water Pollut. Control Fed. 1974, 46, 1849–1872. [Google Scholar]
- Carey, R.O.; Migliaccio, K.W. Contribution of wastewater treatment plant effluents to nutrient dynamics in aquatic systems: A review. Environ. Manag. 2009, 44, 205–217. [Google Scholar] [CrossRef] [PubMed]
- Haggard, B.E.; Stanley, E.H.; Storm, D.E. Nutrient retention in a point-source-enriched stream. J. N. Am. Benthol. Soc. 2005, 24, 29–47. [Google Scholar] [CrossRef]
- Ekka, S.A.; Haggard, B.E.; Matlock, M.D.; Chaubey, I. Dissolved phosphorus concentrations and sediment interactions in effluent-dominated Ozark streams. Ecol. Eng. 2006, 26, 375–391. [Google Scholar] [CrossRef]
- Tchobanoglous, G.; Burton, F.L.; Stensil, H.D. Wastewater Engineering: Treatment and Reuse, 4th ed.; McGraw-Hill: New York, NY, USA, 2003. [Google Scholar]
- Ra, J.S.; Kim, S.D.; Chang, N.I.; An, K.-G. Ecological health assessments based on whole effluent toxicity tests and the index of biological integrity in temperate streams influenced by wastewater treatment plant effluents. Environ. Toxicol. Chem. 2007, 26, 2010–2018. [Google Scholar] [CrossRef] [PubMed]
- Andersen, C.B.; Lewis, G.P.; Sargent, K.A. Influence of wastewater-treatment effluent on concentrations and fluxes of solutes in the Bush River, South Carolina, during extreme drought conditions. Environ. Geosci. 2004, 11, 28–41. [Google Scholar] [CrossRef]
- Downing, J.A.; McCauley, E. The nitrogen: Phosphorus relationship in lakes. Limnol. Oceanogr. 1992, 37, 936–945. [Google Scholar] [CrossRef]
- Redfield, A.C. The biological control of chemical factors in the environment. Am. Sci. 1958, 46, 205–221. [Google Scholar]
- Sakamoto, M. Primary production by phytoplankton community in some Japanese lakes and its dependence on lake depth. Arch. Hydrobiol. 1966, 62, 1–28. [Google Scholar]
- Forsberg, C.; Ryding, S.O. Eutrophication parameters and trophic state indices in 30 Swedish waste receiving lakes. Arch. Hydrobiol. 1980, 89, 189–207. [Google Scholar]
- Huber, W.C.; Brezonik, P.L.; Heaney, J.P.; Dickinson, R.E.; Preston, S.D.; Dwornik, D.S.; DeMaio, M.A. A Classification of Florida Lakes, 1st ed.; Water Resources Research Center, University of Florida: Tallahassee, FL, USA, 1982; pp. 1–547. [Google Scholar]
- Canfield, D.E. Prediction of chlorophyll a concentrations in Florida lakes: The importance of phosphorus and nitrogen. J. Am. Water Resour. Assoc. 1983, 19, 255–262. [Google Scholar] [CrossRef]
- Biggs, B.J. Eutrophication of streams and rivers: Dissolved nutrient-chlorophyll relationships for benthic algae. J. N. Am. Benthol. Soc. 2000, 19, 17–31. [Google Scholar] [CrossRef]
- Volk, C.; Kiffney, P. Comparison of fatty acids and elemental nutrients in periphyton, invertebrates, and cutthroat trout (oncorhynchus clarki) in conifer and alder streams of western washington state. Aquat. Ecol. 2012, 46, 85–99. [Google Scholar] [CrossRef]
- Deans, C.A.; Behmer, S.T.; Kay, A.; Voelz, N. The importance of dissolved N:P ratios on mayfly (baetis spp.) growth in high-nutrient detritus-based streams. Hydrobiologia 2014, 742, 15–26. [Google Scholar] [CrossRef]
- Smith, V.H. Predictive models for the biomass of blue-green algae in lakes. J. Am. Water Resour. Assoc. 1985, 21, 433–439. [Google Scholar] [CrossRef]
- Klausmeier, C.A.; Litchman, E.; Daufresne, T.; Levin, S.A. Optimal nitrogen-phosphorus stoichiometry of phytoplankton. Nature 2004, 429, 171–174. [Google Scholar] [CrossRef] [PubMed]
- Lindeman, R.L. The trophic-dynamic aspect of ecology. Ecology 1942, 23, 399–417. [Google Scholar] [CrossRef]
- Jeppesen, E.; Peder Jensen, J.; SØndergaard, M.; Lauridsen, T.; Landkildehus, F. Trophic structure, species richness and biodiversity in Danish lakes: Changes along a phosphorus gradient. Freshw. Biol. 2000, 45, 201–218. [Google Scholar] [CrossRef]
- Bachmann, R.W.; Jones, B.L.; Fox, D.D.; Hoyer, M.; Bull, L.A.; Canfield, D.E., Jr. Relations between trophic state indicators and fish in Florida (USA) lakes. Can. J. Fish. Aquat. Sci. 1996, 53, 842–855. [Google Scholar] [CrossRef]
- Rohlich, G.A. Fish as indices of eutrophication. In Eutrophication: Causes, Consequences, Correctives, Proceedings of a Symposium, 1st ed.; Larkin, P.A., Northcote, T.G., Eds.; National Academy of Sciences: Washington, DC, USA, 1969; Volume 1, pp. 256–273. [Google Scholar]
- Oberdorff, T.; Pont, D.; Hugueny, B.; Porcher, J.-P. Development and validation of a fish-based index for the assessment of “river health” in France. Freshw. Biol. 2002, 47, 1720–1734. [Google Scholar] [CrossRef]
- Choi, J.-W.; Kumar, H.K.; Han, J.-H.; An, K.-G. The development of a regional multimetric fish model based on biological integrity in lotic ecosystems and some factors influencing the stream health. Water Air Soil Pollut. 2011, 217, 3–24. [Google Scholar] [CrossRef]
- Frey, J.W.; Bell, A.H.; Hambrook Berkman, J.A.; Lorenz, D.L. Assessment of Nutrient Enrichment by Use of Algal-, Invertebrate-, and Fish Community Attributes in Wadeable Streams in Ecoregions Surrounding the Great Lakes; U.S. Geological Survey Scientific Investigations Report 2011–5009; U.S. Geological Survey: Reston, WV, USA, 2011; pp. 1–49.
- Noble, R.A.A.; Cowx, I.G.; Goffaux, D.; Kestemont, P. Assessing the health of European rivers using functional ecological guilds of fish communities: Standardising species classification and approaches to metric selection. Fish. Manag. Ecol. 2007, 14, 381–392. [Google Scholar] [CrossRef]
- Eaton, A.D.; Franson, M.A.H. (Eds.) Standard Methods for the Examination of Water and Wastewater; American Public Health Association: Washington, DC, USA, 2005.
- Marker, A.F.H.; Crowther, C.A.; Gunn, R.J.M. Methanol and acetone as solvents for estimating chlorophyll a and phaeopigments by spectrophotometry. Arch. Hydrobiol. Beih. Ergebn. Limnol. 1980, 14, 52–69. [Google Scholar]
- The Ministry of Environment (MOE)/National Institute of Environmental Research (NIER). Researches for Integrative Assessment Methodology of Aquatic Environments (III): Development of Aquatic Ecosystem Health Assessment and Evaluation System, 1st ed.; MOE/NIER: Incheon, Korea, 2006. [Google Scholar]
- The Ministry of Environment (MOE)/National Institute of Environmental Research (NIER). The Survey and Evaluation of Aquatic Ecosystem Health in Korea, 1st ed.; MOE/NIER: Incheon, Korea, 2008. [Google Scholar]
- Lee, J.H.; Han, J.H.; Kumar, H.K.; Choi, J.K.; Byeon, H.K.; Choi, J.; Kim, J.K.; Jang, M.H.; Park, H.K.; An, K.-G. National-level integrative ecological health assessments based on the index of biological integrity, water quality, and qualitative habitat evaluation index, in Korea rivers. Ann. Limnol. Int. J. Lim. 2011, 47, S73–S89. [Google Scholar] [CrossRef]
- Plafkin, J.L.; Barbour, M.T.; Porter, K.D.; Gross, S.K.; Hughes, R.M. Rapid Bioassessment Protocols for Use in Streams and Rivers: Benthic Macroinvertebrate and Fish; EPA/444/4-89-001; Office of Water Regulations and Standards; US EPA: Washington, DC, USA, 1989; pp. 1–34.
- Barbour, M.T.; Gerritsen, J.; Snyder, B.D.; Stribling, J.B. Rapid Bioassessment Protocols for Use in Streams and Wadeable Rivers: Periphyton, Benthic Macroinvertebrates and Fish, 2nd ed.EPA 841-B-99-002; U.S. Environmental Protection Agency, Office of Water: Washington, DC, USA, 1999; pp. 1–35.
- An, K.-G.; Park, S.S.; Shin, J.-Y. An evaluation of a river health using the index of biological integrity along with relations to chemical and habitat conditions. Environ. Int. 2002, 28, 411–420. [Google Scholar] [CrossRef]
- Ohio, E.P.A. Biological Criteria for the Protection of Aquatic Life: Volume III. Standardized Biological Field Sampling and Laboratory Method for Assessing Fish and Macroinvertebrate Communities, 2nd ed.; Ohio, E.P.A., Ed.; Columbus, OH, USA, 2015; pp. 1–64. [Google Scholar]
- U.S. EPA. Fish Field and Laboratory Methods for Evaluating the Biological Integrity of Surface Waters; EPA 600-R-92-111; Environmental Monitoring Systems Laboratory-Cincinnati office of Modeling, Monitoring Systems, and Quality Assurance Office of Research Development, U.S. EPA: Cincinnati, OH, USA, 1993; pp. 1–348.
- Kim, I.S.; Choi, Y.; Lee, C.L.; Lee, Y.J.; Kim, B.J.; Kim, J.H. Illustrated Book of Korean Fishes, 1st ed.; Kyohak: Seoul, Korea, 2005. [Google Scholar]
- Han, J.-H.; Park, C.-S.; An, J.-W.; An, K.-G.; Baek, W.-K. A Guide Book of Freshwater Fishes, 1st ed.; National Science Museum: Daejeon, Korea, 2015. [Google Scholar]
- FishBase. World Wide Web Electronic Publication. Available online: http: //www.fishbase.org (accessed on 18 May 2015).
- Sanders, R.E.; Miltner, R.J.; Yoder, C.O.; Rankin, E.T. The use of external deformities, erosion, lesions, tumors (DELT anomalies) in fish assemblages for characterizing aquatic resources: A case study of seven Ohio streams. In Assessing the Sustainability and Biological Integrity of Water Resources Using Fish Communities, 1st ed.; Simon, T.P., Ed.; CRC: Boca Raton, FL, USA, 1999; pp. 225–245. [Google Scholar]
- Karr, J.R. Assessment of biotic integrity using fish communities. Fisheries 1981, 6, 21–27. [Google Scholar] [CrossRef]
- Karr, J.R.; Fausch, K.D.; Angermeier, P.L.; Yant, P.R.; Schlosser, I.J. Assessing Biological Integrity in Running Water: A Method and Its Rationale, 1st ed.; Illinois National History Survey Special Publication 5: Champaign, IL, USA, 1986; pp. 1–28. [Google Scholar]
- An, K.-G.; Park, S.S. Indirect influence of the summer monsoon on chlorophyll-total phosphorus models in reservoirs: A case study. Ecol. Models 2002, 152, 191–203. [Google Scholar] [CrossRef]
- An, K.-G. Long-term seasonal and interannual variability of epilimnetic nutrients (N, P), chlorophyll-a, and suspended solids at the Dam site of Yongdam Reservoir and empirical models. Korean J. Limnol. 2011, 44, 214–225. [Google Scholar]
- Van Nieuwenhuyse, E.E.; Jones, J.R. Phosphorus-chlorophyll relationship in temperate streams and its variation with stream catchment area. Can. J. Fish. Aquat. Sci. 1996, 53, 99–105. [Google Scholar] [CrossRef]
- Miltner, R.J.; Rankin, E.T. Primary nutrients and the biotic integrity of rivers and streams. Freshw. Biol. 1998, 40, 145–158. [Google Scholar] [CrossRef]
- Lee, J.H.; An, K.-G. Integrative restoration assessment of an urban stream using multiple modeling approaches with physical, chemical, and biological integrity indicators. Ecol. Eng. 2014, 62, 153–167. [Google Scholar] [CrossRef]
- Robertson, D.M.; Graczyk, D.J.; Garrison, P.J.; Wang, L.; LaLiberte, G.; Bannerman, R. Nutrient Concentrations and Their Relations to the Biotic Integrity of Wadeable Streams in Wisconsin, 1st ed.; Professional Paper 1722; U.S. Geological Survey: Reston, VA, USA, 2006; pp. 1–139. [Google Scholar]
- Yeom, D.-H.; Lee, S.-A.; Kang, G.S.; Seo, J.; Lee, S.-K. Stressor identification and health assessment of fish exposed to wastewater effluents in Miho Stream, South Korea. Chemosphere 2007, 67, 2282–2292. [Google Scholar] [CrossRef] [PubMed]
- Green, M.B.; Finlay, J.C. Patterns of hydrologic control over stream water total nitrogen to total phosphorus ratios. Biogeochemistry 2010, 99, 15–30. [Google Scholar] [CrossRef]
- Ellison, M.E.; Brett, M.T. Particulate phosphorus bioavailability as a function of stream flow and land cover. Water Res. 2006, 40, 1258–1268. [Google Scholar] [CrossRef] [PubMed]
- Higgins, K.A.; Vanni, M.J.; González, M. Detritivory and the stoichiometry of nutrient cycling by a dominant fish species in lakes of varying productivity. Oikos 2006, 114, 419–430. [Google Scholar] [CrossRef]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yun, Y.-J.; An, K.-G. Roles of N:P Ratios on Trophic Structures and Ecological Stream Health in Lotic Ecosystems. Water 2016, 8, 22. https://doi.org/10.3390/w8010022
Yun Y-J, An K-G. Roles of N:P Ratios on Trophic Structures and Ecological Stream Health in Lotic Ecosystems. Water. 2016; 8(1):22. https://doi.org/10.3390/w8010022
Chicago/Turabian StyleYun, Young-Jin, and Kwang-Guk An. 2016. "Roles of N:P Ratios on Trophic Structures and Ecological Stream Health in Lotic Ecosystems" Water 8, no. 1: 22. https://doi.org/10.3390/w8010022
APA StyleYun, Y. -J., & An, K. -G. (2016). Roles of N:P Ratios on Trophic Structures and Ecological Stream Health in Lotic Ecosystems. Water, 8(1), 22. https://doi.org/10.3390/w8010022