
Spatial Distribution of Benthic Macroinvertebrate Assemblages in Relation to Environmental Variables in Korean Nationwide Streams

 , , ,
, , ,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
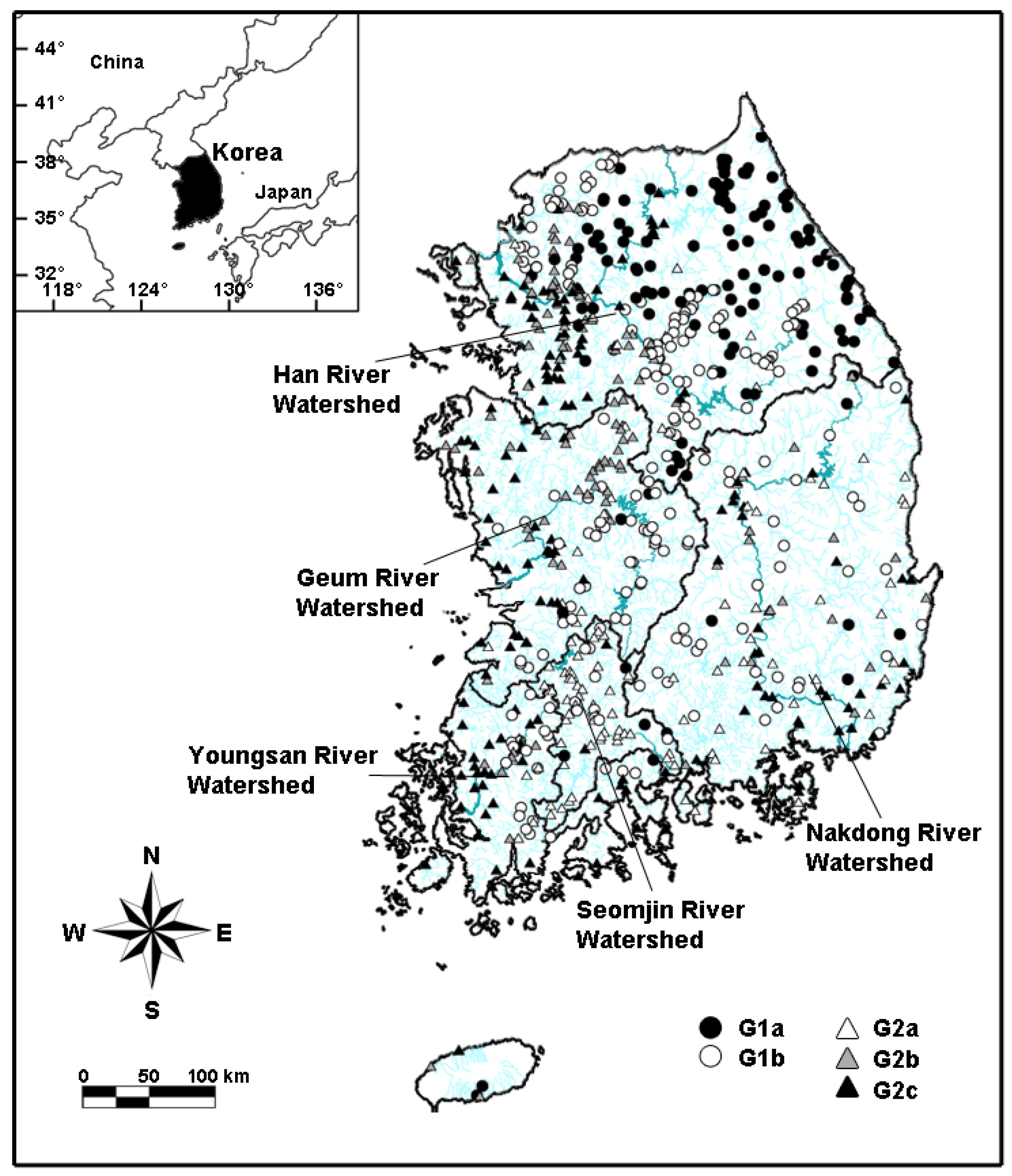
2.1. Study Area

2.2. Measurement of Environmental Variables
2.3. Sampling of Benthic Macroinvertebrate
2.4. Data Analysis
3. Results
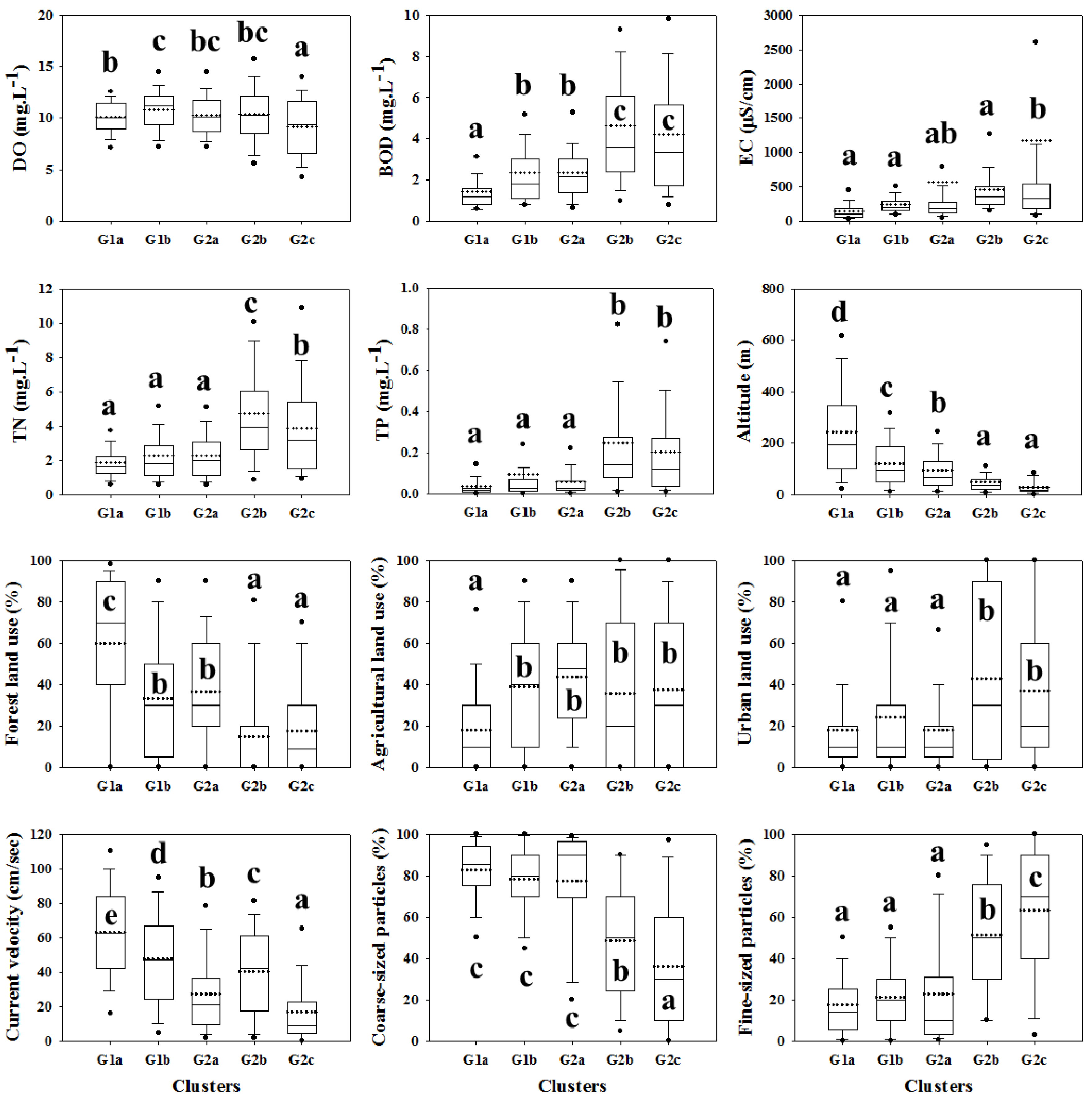
3.1. Environmental Characteristics
3.2. Benthic Macroinvertebrate Assemblages

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | HRW (n = 320) | NRW (n = 130) | GRW (n = 130) | YRW (n = 76) | SRW (n = 64) | Total (n = 720) | p (K–W) |
|---|---|---|---|---|---|---|---|
| (a) Regional and physical instream variables | |||||||
| Altitude (m) | 147.5 (151.6) d | 89.6 (119.1) b,c | 57.7 (61.2) a,b | 32.5 (35.1) a | 118.1 (79.3) c,d | 106.1 (126.2) | 0.000 |
| 1.0–721.0 | 1.0–629.0 | 0.0–278.0 | 0.0–211.0 | 1.0–335.0 | 0.0–721.0 | ||
| % Urban | 31.7 (32.0) b | 23.0 (30.8) a,b | 30.9 (35.8) b | 31.4 (30.2) b | 14.6 (12.4) a | 28.4 (31.5) | 0.000 |
| 0–100 | 0–100 | 0–100 | 0–90 | 0–80 | 0–100 | ||
| % Agriculture | 24.7 (27.5) a | 42.5 (31.0) b,c | 46.6 (38.9) c | 45.4 (30.7) c | 35.9 (20.7) b | 35.0 (31.7) | 0.000 |
| 0–100 | 0–100 | 0–100 | 0–90 | 0–80 | 0–100 | ||
| % Forest | 35.8 (33.2) b | 31.4 (27.8) b | 17.9 (30.1) a | 22.6 (24.3) a | 48.8 (24.6) c | 31.5 (31.3) | 0.000 |
| 0–100 | 0–100 | 0–100 | 0–100 | 0–95 | 0–100 | ||
| Water velocity (cm/s) | 53.8 (27.9) c | 14.7 (14.0) a | 38.1 (34.5) b | 17.7 (13.1) a | 41.9 (26.0) b | 39.0 (30.4) | 0.000 |
| 0.0–140.0 | 0.0–67.0 | 0.0–137.7 | 0.4–47.5 | 1.4–98.2 | 0.0–140.0 | ||
| % Fine particles | 31.1 (27.9) b | 36.5 (36.8) b,c | 43.6 (34.5) c | 54.7 (31.3) d | 10.8 (22.9) a | 35.0 (32.6) | 0.000 |
| 0.0–100.0 | 0.0–100.0 | 0.0–100.0 | 0.0–100.0 | 0.0–100.0 | 0.0–100.0 | ||
| % Coarse particles | 68.6 (28.1) c | 63.5 (36.8) b,c | 56.4 (34.5) b | 45.3 (31.3) a | 89.2 (22.9) d | 64.8 (32.7) | 0.000 |
| 0.0–100.0 | 0.0–100.0 | 0.0–100.0 | 0.0–100.0 | 0.0–100.0 | 0.0–100.0 | ||
| (b) Chemical variables | |||||||
| pH | 8.1 (0.8) b,c | 8.0 (0.8) a,b | 8.3 (0.9) c | 8.0 (0.7) a,b | 7.8 (0.7) a | 8.1 (0.8) | 0.003 |
| 6.5–10.1 | 6.2–10.6 | 7.0–11.1 | 6.7–10.1 | 6.7–9.6 | 6.2–11.1 | ||
| DO (mg/L) | 9.78 (2.47) a | 10.81 (2.36) b | 10.86 (3.36) b | 10.06 (1.69) a | 9.57 (1.32) a | 10.17 (2.54) | 0.000 |
| 2.42–16.10 | 2.55–17.34 | 2.74–17.86 | 6.19–15.40 | 7.11–12.69 | 2.42–17.86 | ||
| BOD (mg/L) | 3.1 (3.7) b,c | 1.9 (1.3) a | 3.7 (2.0) c | 3.5 (3.0) b,c | 2.7 (1.7) b | 3.0 (2.9) | 0.000 |
| 0.3–37.5 | 0.4–10.4 | 0.8–9.1 | 0.3–13.3 | 0.3–12.3 | 0.3–37.5 | ||
| EC (μS/cm) | 299.8 (337.6) a | 1358.7 (6067.1) b | 404.8 (349.8) a | 270.7 (402.6) a | 1284.6 (5823.4) b | 594.4 (3141.6) | 0.000 |
| 10.3–2729.0 | 19.7–44000.0 | 86.1–2780.0 | 28.5–3082.0 | 32.6–33360.0 | 10.3–44000.0 | ||
| TN (mg/L) | 3.15 (2.97) b | 2.10 (1.66) a | 3.60 (2.42) b | 3.36 (3.59) b | 2.32 (1.39) a | 2.99 (2.69) | 0.000 |
| 0.38–23.78 | 0.32–11.30 | 0.29–14.27 | 0.69–27.71 | 0.44–5.77 | 0.29–27.71 | ||
| TP (mg/L) | 0.15 (0.41) b,c | 0.07 (0.15) a,b | 0.14 (0.15) b,c | 0.21 (0.28) c | 0.05 (0.06) a | 0.13 (0.30) | 0.000 |
| 0.00–5.59 | 0.00–0.91 | 0.01–1.01 | 0.01–1.66 | 0.00–0.32 | 0.00–5.59 | ||
| Turbidity (NTU) | 9.3 (16.6) b | 8.6 (7.4) b | 13.9 (22.3) b | 7.8 (45.9) b | 2.1 (3.3) a | 9.2 (21.2) | 0.000 |
| 0.0–152.0 | 0.0–34.2 | 0.4–182.4 | 0.0–400.0 | 0.0–16.9 | 0.0–400.0 | ||
| Biological Attributes | HRW (n = 320) | NRW (n = 130) | GRW (n = 130) | YRW (n = 76) | SRW (n = 64) | Total (n = 720) | RA | p (K–W) | |
|---|---|---|---|---|---|---|---|---|---|
| (a) Assemblage attributes | |||||||||
| Taxa richness | 15.7 (9.4) a | 11.7 (7.3) b | 15.5 (10.8) a | 10.6 (6.3) b | 15.1 (8.0) a | 14.4 (9.1) | - | 0.000 | |
| EPT richness | 10.3 (8.0) c | 5.3 (5.5) a | 7.6 (8.2) b | 4.5 (4.7) a | 8.5 (5.4) b | 8.1 (7.5) | - | 0.000 | |
| Taxa abundance | 2867.5 (9791.9) b | 864.9 (1035.2) a | 3944.4 (5964.7) b | 947.0 (1054.1) a | 612.0 (548.2) a | 2297.1 (7121.0) | - | 0.000 | |
| EPT abundance | 1066.9 (1702.9) b | 303.2 (726.5) a | 1552.5 (2441.0) c | 316.4 (545.0) a | 278.6 (315.8) a | 867.4 (1647.2) | - | 0.000 | |
| Dominance index | 0.65 (0.21) b | 0.70 (0.20) b,c | 0.71 (0.21) b,c | 0.73 (0.20) c | 0.52 (0.23) a | 0.67 (0.22) | - | 0.000 | |
| Shannon diversity index | 2.33 (1.00) b | 2.08 (0.91) b | 2.09 (1.02) b | 1.79 (0.94) a | 2.63 (1.09) c | 2.21 (1.01) | - | 0.000 | |
| (b) Taxa abundance of higher taxonomic group | |||||||||
| Non-Insecta | Platyhelminthes | 18.7 (78.8) | 28.7 (196.7) | 25.5 (102.4) | 9.9 (49.2) | 17.2 (37.8) | 20.7 (109.6) | 0.01 | 0.447 |
| Nematomorpha | 0.2 (0.9) | 0.1 (0.6) | 0.1 (0.5) | 0.1 (0.8) | 0.0 (0.0) | 0.1 (0.7) | 0.00 | 0.082 | |
| Mollusca | 10.7 (39.4) a | 44.4 (151.5) c | 30.4 (54.3) b | 27.4 (49.5) b | 21.2 (30.9) a,b | 23.0 (76.5) | 0.01 | 0.000 | |
| Annelida | 603.5 (6826.2) b | 39.9 (90.8) a | 548.3 (2997.6) b | 21.0 (61.4) a | 68.1 (91.7) a | 382.7 (4731.0) | 0.20 | 0.000 | |
| Crustacea | 1.9 (12.2) a | 42.9 (214.3) a | 41.2 (243.9) a | 109.8 (560.6) b | 1.4 (7.0) a | 27.7 (230.6) | 0.01 | 0.000 | |
| Insecta | Ephemeroptera | 692.6 (1132.9) b | 316.6 (594.6) a | 565.1 (1015.1) b | 199.0 (308.7) a | 377.3 (389.0) a | 521.5 (936.0) | 0.27 | 0.000 |
| Odonata | 3.9 (12.3) a | 4.2 (12.4) a | 14.5 (37.1) b | 11.9 (29.2) b | 0.7 (2.2) a | 6.4 (21.3) | 0.00 | 0.000 | |
| Plecoptera | 6.6 (21.6) b | 5.6 (29.6) b | 4.2 (21.5) a,b | 0.1 (0.7) a | 0.7 (4.2) a | 4.8 (21.3) | 0.00 | 0.000 | |
| Hemiptera | 1.1 (9.3) a | 38.7 (207.4) b | 25.0 (129.1) a,b | 25.5 (213.5) a,b | 0.0 (0.3) a | 14.7 (125.7) | 0.01 | 0.000 | |
| Megaloptera | 2.6 (10.4) b | 1.2 (6.0) a,b | 1.6 (6.0) a,b | 0.5 (2.3) a | 1.0 (9.8) a,b | 1.8 (8.4) | 0.00 | 0.000 | |
| Coleoptera | 15.3 (97.6) a | 28.6 (80.6) a | 80.9 (200.8) c | 8.9 (31.5) a | 56.9 (94.9) b | 32.6 (119.3) | 0.02 | 0.000 | |
| Diptera | 449.3 (780.2) b | 309.8 (452.1) b | 806.2 (1853.3) c | 344.1 (712.1) b | 105.0 (164.4) a | 446.9 (1009.5) | 0.23 | 0.000 | |
| Trichoptera | 549.2 (1029.0) b | 306.6 (771.8) a | 845.3 (1564.4) c | 114.5 (296.2) a | 126.4 (295.1) a | 475.4 (1044.8) | 0.24 | 0.000 | |
| Lepidoptera | 0.0 (0.1) a | - | 0.1 (0.9) b | 0.0 (0.6) a,b | - | 0.0 (0.4) | 0.00 | 0.000 | |
| Neuroptera | - | - | - | 0.0 (0.3) | - | 0.0 (0.1) | 0.00 | 0.076 | |

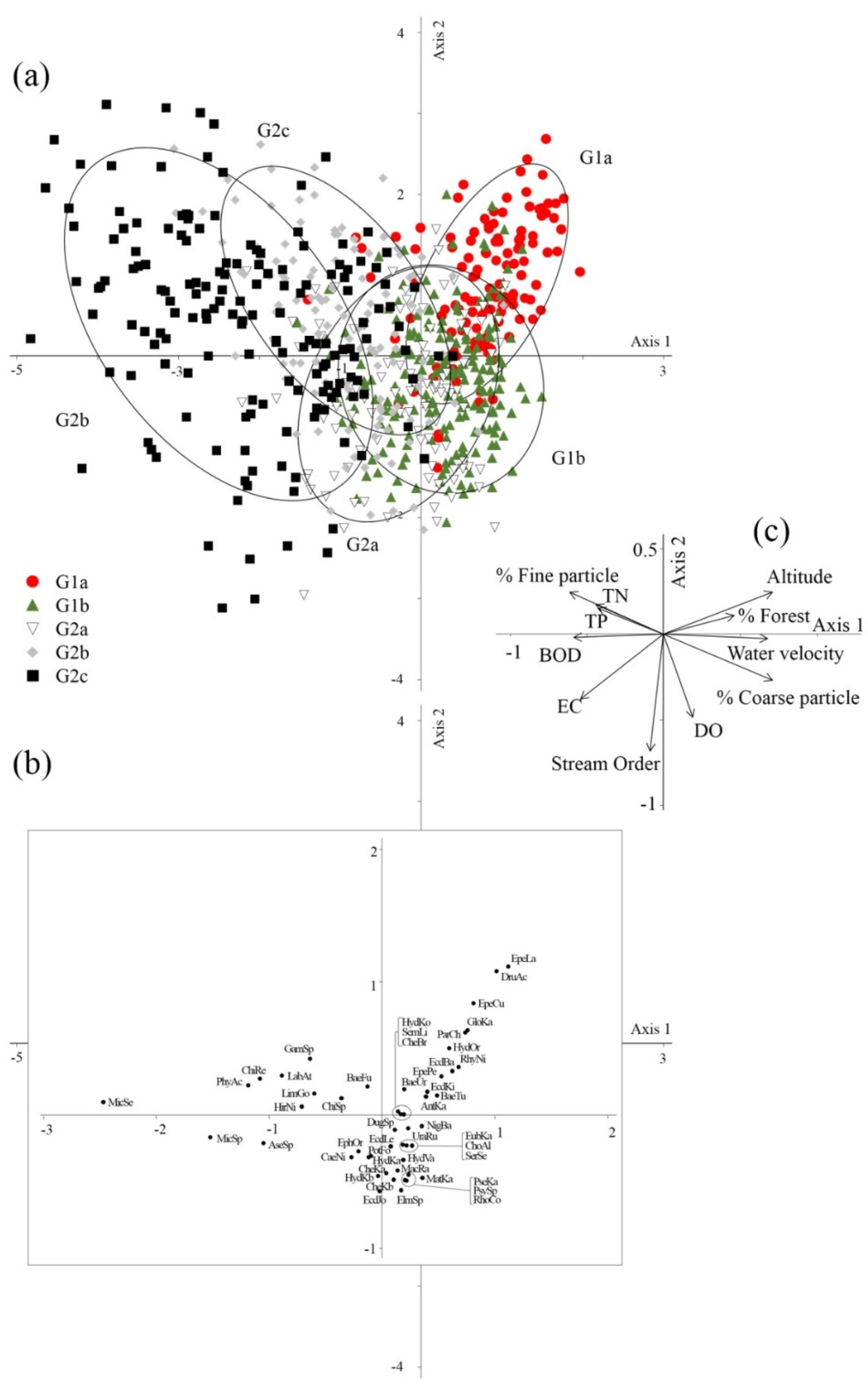
3.3. Macroinvertebrate-Based Site Classification

| Taxa | Cluster Group | p | ||||
|---|---|---|---|---|---|---|
| G1a | G1b | G2a | G2b | G2c | ||
| Rhyacophila nigrocephala Iwata | 47 | 16 | 0 | 0 | 0 | 0.001 |
| Epeorus nipponicus (Uéno) | 46 | 1 | 0 | 0 | 0 | 0.001 |
| Glossosoma KUa | 44 | 4 | 0 | 0 | 0 | 0.001 |
| Drunella aculea (Allen) | 44 | 1 | 0 | 0 | 0 | 0.001 |
| Hydropsyche orientalis Martynov | 40 | 7 | 1 | 1 | 0 | 0.001 |
| Uracanthella punctisetae (Matsumura) | 21 | 48 | 6 | 6 | 0 | 0.001 |
| Hydropsyche valvata Martynov | 6 | 46 | 2 | 9 | 0 | 0.001 |
| Cheumatopsyche brevilineata Iwata | 15 | 41 | 5 | 10 | 0 | 0.001 |
| Hydropsyche kozhantschikovi Martynov | 21 | 40 | 3 | 16 | 0 | 0.001 |
| Psychomyia sp. | 0 | 32 | 1 | 5 | 0 | 0.001 |
| Ephemera orientalis McLachlan | 2 | 20 | 41 | 5 | 2 | 0.001 |
| Ecdyonurus levis (Navás) | 5 | 28 | 40 | 1 | 0 | 0.001 |
| Ecdyonurus joernensis Bengtsson | 0 | 12 | 25 | 1 | 0 | 0.001 |
| Mataeopsephus KUa | 0 | 16 | 23 | 0 | 0 | 0.001 |
| Asellus sp. | 0 | 2 | 11 | 3 | 6 | 0.001 |
| Hirudo nipponia Whitman | 1 | 10 | 2 | 28 | 4 | 0.001 |
| Chironomini sp. | 1 | 7 | 9 | 27 | 21 | 0.001 |
| Limnodrilus gotoi Hatai | 5 | 12 | 18 | 23 | 9 | 0.001 |
| Micronecta sedula Horváth | 0 | 0 | 0 | 0 | 20 | 0.001 |
| Physa acuta Draparnaud | 2 | 3 | 2 | 12 | 16 | 0.001 |
| Micronecta sp. | 0 | 0 | 5 | 0 | 5 | 0.007 |
| Total number of significant indicator species | 12 | 21 | 8 | 3 | 3 | - |
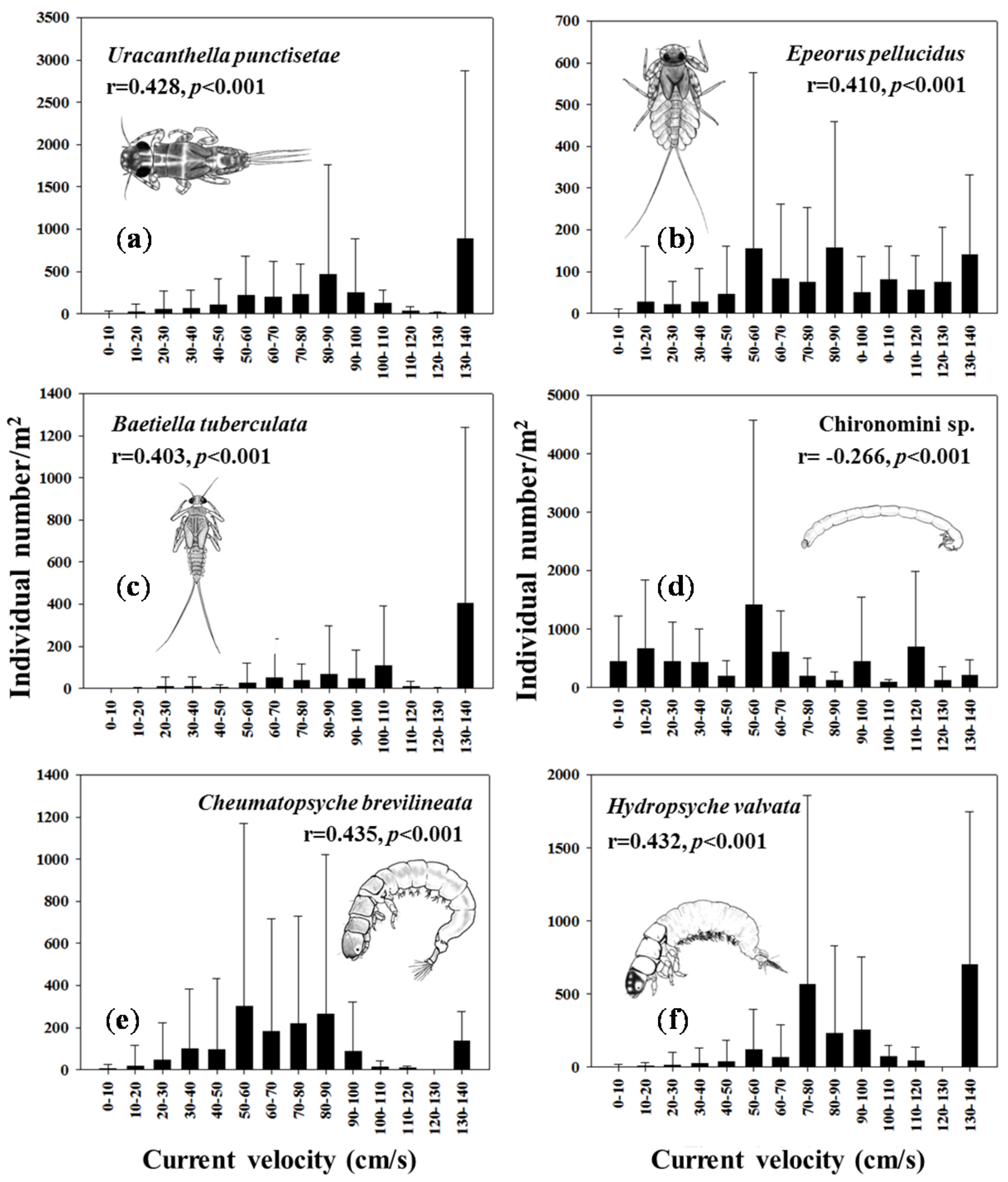
3.4. Environmental Variables Affecting Macroinvertebrate Distributions

| Variables | Axis 1 | Axis 2 | Axis 3 |
|---|---|---|---|
| (a) Correlation coefficients | |||
| Altitude | 0.793 ** | 0.236 ** | 0.013 |
| Stream order | −0.105 ** | −0.658 ** | 0.099 ** |
| % Urban | −0.218 ** | 0.134 ** | −0.317 ** |
| % Agriculture | −0.199 ** | −0.273 ** | 0.133 ** |
| % Forest | 0.504 ** | 0.055 | 0.266 ** |
| Water velocity | 0.662 ** | −0.054 | −0.510 ** |
| % Fine particles | −0.744 ** | 0.325 ** | −0.076 * |
| % Coarse particles | 0.747 ** | −0.327 ** | 0.075 * |
| pH | 0.011 | −0.236 ** | 0.007 |
| DO | 0.304 ** | −0.467 ** | −0.166 ** |
| BOD | −0.550 ** | 0.164 ** | −0.370 ** |
| EC | −0.196 ** | −0.278 ** | −0.116 ** |
| TN | −0.463 ** | 0.266 ** | −0.387 ** |
| TP | −0.373 ** | 0.229 ** | −0.314 ** |
| (b) Summary of CCA results | |||
| Eigenvalue | 0.281 | 0.101 | 0.073 |
| % variance explained in taxa data | 9.8 | 3.5 | 2.5 |
| Cumulative % variance explained | 9.8 | 13.3 | 15.9 |
| p value | 0.010 | 0.010 | 0.010 |
| Total variance | 2.869 | - | - |
4. Discussion
4.1. Macroinvertebrate Taxonomic Composition
4.2. Environmental Relationships with Macroinvertebrate Distribution
4.3. Considerations to Improve Macroinvertebrate Biomonitoring Programs
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix A
| Code | Taxon |
|---|---|
| Phylum Platyhelminthes | |
| DugSp | Dugesia sp. |
| Phylum Mollusca | |
| SemLi | Semisulcospira libertina (Gould) |
| PhyAc | Physa acuta Draparnaud |
| Phylum Annelida | |
| LimGo | Limnodrilus gotoi Hatai |
| HirNi | Hirudo nipponia Whitman |
| Phylum Arthropoda | |
| Class Crustacea | |
| AseSp | Asellus sp. |
| GamSp | Gammarus sp. |
| Class Insecta | |
| Order Ephemeroptera | |
| BaeTu | Baetiella tuberculata (Kazlauskas) |
| BaeFu | Baetis fuscatus (Linnaeus) |
| BaeUr | Baetis ursinus Kazlauskas |
| LabAt | Labiobaetis atrebatinus (Eaton) |
| NigBa | Nigrobaetis bacillus (Kluge) |
| EcdBa | Ecdyonurus bajkovae Kluge |
| EcdJo | Ecdyonurus joernensis Bengtsson |
| EcdKi | Ecdyonurus kibunensis Imanishi |
| EcdLe | Ecdyonurus levis (Navás) |
| EpeNi | Epeorus nipponicus (Uéno) |
| EpeLa | Epeorus latifolium (Uéno) |
| EpePe | Epeorus pellucidus (Brodsky) |
| ChoAl | Choroterpes altioculus Kluge |
| ParJa | Paraleptophlebia japonica (Matsumura) |
| PotFo | Potamanthus formosus Eaton |
| RhoCo | Rhoenanthus coreanus (Yoon and Bae) |
| DruAc | Drunella aculea (Allen) |
| EphOr | Ephemera orientalis McLachlan |
| SerSe | Serratella setigera (Bajkova) |
| UraPu | Uracanthella punctisetae (Matsumura) |
| CaeNi | Caenis nishinoae Malzacher |
| Order Hemiptera | |
| MicSe | Micronecta sedula Horváth |
| MicSp | Micronecta sp. |
| Order Coleoptera | |
| ElmSp | Elmidae sp. |
| EubKa | Eubrianax KUa |
| MatKa | Mataeopsephus KUa |
| PseKa | Psephenoides KUa |
| Order Diptera | |
| AntKa | Antocha KUa |
| CulSp | Culex sp. |
| ChiSp | Chironomidae spp. (non-red type) |
| ChiRe | Chironomini spp. (red-type) |
| Order Trichoptera | |
| RhyNi | Rhyacophila nigrocephala Iwata |
| HydKa | Hydroptila KUa |
| GloKa | Glossosoma KUa |
| CheBr | Cheumatopsyche brevilineata Iwata |
| CheKa | Cheumatopsyche KUa |
| CheKb | Cheumatopsyche KUb |
| HydKo | Hydropsyche kozhantschikovi Martynov |
| HydKb | Hydropsyche KUb |
| HydOr | Hydropsyche orientalis Martynov |
| HydVa | Hydropsyche valvata Martynov |
| MacRa | Macrostemum radiatum McLachlan |
| PsySp | Psychomyia sp. |
References
- Dudgeon, D.; Arthington, A.H.; Gessner, M.O.; Kawabata, Z.I.; Knowler, D.J.; Lévêque, C.; Naiman, R.J.; Prieur-Richard, A.H.; Soto, D.; Stiassny, M.L.J.; et al. Freshwater biodiversity: Importance, threats, status and conservation challenges. Biol. Rev. 2006, 81, 163–182. [Google Scholar] [CrossRef] [PubMed]
- Strayer, D.L.; Dudgeon, D. Freshwater biodiversity conservation: Recent progress and future challenges. J. N. Am. Benthol. Soc. 2010, 29, 344–358. [Google Scholar] [CrossRef]
- Li, F.; Chung, N.; Bae, M.-J.; Kwon, Y.-S.; Park, Y.-S. Relationships between stream macroinvertebrates and environmental variables at multiple spatial scales. Freshwater Biol. 2012, 57, 2107–2124. [Google Scholar] [CrossRef]
- Dudgeon, D. The ecology of tropical Asian rivers and streams in relation to biodiversity conservation. Annu. Rev. Ecol. Syst. 2000, 31, 239–263. [Google Scholar] [CrossRef]
- De Silva, S.S.; Abery, N.W.; Nguyen, T.T.T. Endemic freshwater finfish of Asia: Distribution and conservation status. Divers. Distrib. 2007, 13, 172–184. [Google Scholar] [CrossRef]
- Vörösmarty, C.J.; McIntyre, P.B.; Gessner, M.O.; Dudgeon, D.; Prusevich, A.; Green, P.; Glidden, S.; Bunn, S.E.; Sullivan, C.A.; Liermann, C.E.; et al. Global threats to human water security and river biodiversity. Nature 2010, 467, 555–561. [Google Scholar] [CrossRef] [PubMed]
- Strayer, D.L. Challenges for freshwater invertebrate conservation. J. N. Am. Benthol. Soc. 2006, 25, 271–287. [Google Scholar] [CrossRef]
- Buss, D.F.; Baptista, D.F.; Silveira, M.P.; Nessimian, J.L.; Dorvillé, L.F.M. Influence of water chemistry and environmental degradation on macroinvertebrate assemblages in a river basin in south-east Brazil. Hydrobiologia 2002, 481, 125–136. [Google Scholar] [CrossRef]
- Bott, T.L.; Brock, J.T.; Dunn, C.S.; Naiman, R.J.; Ovink, R.W.; Petersen, R.C. Benthic community metabolism in four temperate stream systems: An inter-biome comparison and evaluation of the river continuum concept. Hydrobiologia 1985, 220, 109–117. [Google Scholar] [CrossRef]
- Merz, J.R.; Ochikubo Chan, L.K. Effects of gravel augmentation on macroinvertebrate assemblages in a regulated California river. River Res. Appl. 2005, 21, 61–74. [Google Scholar] [CrossRef]
- Nelson, S.M.; Lieberman, D.M. The influence of flow and other environmental factors on benthic invertebrates in the Sacramento River, USA. Hydrobiologia 2002, 489, 117–129. [Google Scholar] [CrossRef]
- Jiang, X.M.; Xiong, J.; Qiu, J.W.; Wu, J.M.; Wang, J.W.; Xie, Z.C. Structure of macroinvertebrate communities in relation to environmental variables in a subtropical Asian river system. Int. Rev. Hydrobiol. 2010, 95, 42–57. [Google Scholar] [CrossRef]
- Vought, L.B.M.; Kullberg, A.; Petersen, R.C. Effect of riparian structure, temperature and channel morphometry on detritus processing in channelized and natural woodland streams in southern Sweden. Aquat. Conserv. Mar. Freshw. Ecosyst. 1998, 8, 273–285. [Google Scholar] [CrossRef]
- Townsend, C.R.; Hildrew, A.G.; Francis, J. Community structure in some southern English streams: The influence of physicochemical factors. Freshw. Biol. 1983, 13, 521–544. [Google Scholar] [CrossRef]
- Richards, C.; Haro, R.J.; Johnson, L.B.; Host, G.E. Catchment and reach-scale properties as indicator of macroinvertebrate species traits. Freshw. Biol. 1997, 37, 219–230. [Google Scholar] [CrossRef]
- Sandin, L. Benthic macroinvertebrates in Swedish streams: Community structure, taxon richness, and environmental relations. Ecography 2003, 26, 269–282. [Google Scholar] [CrossRef]
- Leps, M.; Tonkin, J.D.; Dahm, V.; Haase, P.; Sundermann, A. Disentangling environmental drivers of benthic invertebrate assemblages: The role of spatial scale and riverscape heterogeneity in a multiple stressor environment. Sci. Total Environ. 2015, 536, 546–556. [Google Scholar] [CrossRef] [PubMed]
- Heino, J. Biodiversity of aquatic insects: Spatial gradients and environmental correlates of assemblage-level measures at large scales. Freshw. Rev. 2009, 2, 1–29. [Google Scholar] [CrossRef]
- Shah, D.N.; Tonkin, J.D.; Haase, P.; Jähnig, S.C. Latitudinal patterns and large-scale environmental determinants of stream insect richness across Europe. Limnologica 2015, 55, 33–43. [Google Scholar] [CrossRef]
- Rossaro, B.; Pietrangelo, A. Macroinvertebrate distribution in streams: A comparison of CA ordination with biotic indices. Hydrobiologia 1993, 263, 109–118. [Google Scholar] [CrossRef]
- Rosenberg, D.M.; Resh, V.H. Freshwater Biomonitoring and Benthic Macroinvertebrates; Chapman and Hall: New York, NY, USA, 1993. [Google Scholar]
- Charvet, S.; Statzner, B.; Usseglio-Polatera, P.; Dumont, B. Traits of benthic macroinvertebrates in semi-natural French streams: An initial application to biomonitoring in Europe. Freshw. Biol. 2000, 43, 277–296. [Google Scholar] [CrossRef]
- Korea Meteorological Administration (KMA). The Meteorological Yearbook in Korea; KMA: Seoul, Korea, 2010. (in Korean) [Google Scholar]
- Jeong, K.S.; Hong, D.G.; Byeon, M.S.; Jeong, J.C.; Kim, H.G.; Kim, D.K.; Joo, G.J. Stream modification patterns in a river basin: Field survey and self-organizing map (SOM) application. Ecol. Inform. 2010, 5, 293–303. [Google Scholar] [CrossRef]
- Hwang, S.-J.; Kim, J.-Y.; Yoon, S.-A.; Kim, B.-H.; Park, M.-H.; You, K.-A.; Lee, H.-Y.; Kim, H.-S.; Kim, Y.-J.; Lee, J.; et al. Distribution of benthic diatoms in Korean rivers and streams in relation to environmental variables. Ann. Limnol. Int. J. Limnol. 2011, 47, S15–S33. [Google Scholar] [CrossRef]
- Craig, D.A. Some of what you should know about water. Bull. N. Am. Benthol. Soc. 1987, 4, 178–182. [Google Scholar]
- The Ministry of Environment/National Institute of Environmental Research (MOE/NIER). Survey and Evaluation of Aquatic Ecosystem Health in Korea; MOE/NIER: Incheon, Korea, 2012. [Google Scholar]
- American Public Health Association (APHA). Standard Methods for the Examination of Water and Wastewater, 21st ed.; APHA: Washington, DC, USA, 2001. [Google Scholar]
- McCune, B.; Grace, J.B. Analysis of Ecological Communities; MjM Software Design: Gleneden Beach, OR, USA, 2002. [Google Scholar]
- Mielke, P.W.; Berry, K.J.; Johnson, E.S. Multiresponse permutation procedures for a priori classifications. Commun. Stat. 1976, A5, 1409–1424. [Google Scholar] [CrossRef]
- Zimmerman, G.M.; Goetz, H.; Mielke, P.W. Use of an improved statistical method for group comparisons to study effects of prairie fire. Ecology 1985, 66, 606–611. [Google Scholar] [CrossRef]
- Dufrene, M.; Legendre, P. Species assemblages and indicator species: The need for flexible asymmetrical approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- Petersen, W.T.; Keister, J.E. Interannual variability in copepod community composition at a coastal station in the northern California Current: A multivariate approach. Deep Sea Res. 2003, 50, 2499–2517. [Google Scholar] [CrossRef]
- McCune, B.; Mefford, M.J. Multivariate Analysis of Ecological Data (Version 4.25); MjM Software: Gleneden Beach, OR, USA, 1999. [Google Scholar]
- Wang, X.; Cai, Q.; Tang, T.; Yang, S.; Li, F. Spatial distribution of benthic macroinvertebrates in the Erhai basin of southwestern China. J. Freshw. Ecol. 2012, 27, 89–96. [Google Scholar] [CrossRef]
- Lee, S.-W.; Hwang, S.-J.; Lee, J.-K.; Jung, D.-I.; Park, Y.-J.; Kim, J.-T. Overview and application of the National Aquatic Ecological Monitoring Program (NAEMP) in Korea. Ann. Limnol. Int. J. Limnol. 2011, 47, S3–S14. [Google Scholar] [CrossRef]
- Hoang, T.H.; Lock, K.; Chi Dang, K.; de Pauw, N.; Goethals, P.L.M. Spatial and temporal patterns of macroinvertebrate communities in the Du River Basin in Northern Vietnam. J. Freshw. Ecol. 2010, 25, 637–647. [Google Scholar] [CrossRef]
- Bae, M.-J.; Kwon, Y.; Hwang, S.-J.; Con, T.-S.; Yang, H.-J.; Kwak, I.-S.; Park, J.-H.; Ham, S.-A.; Park, Y.-S. Relationships between three major stream assemblages and their environmental factors in multiple spatial scales. Ann. Limnol. Int. J. Limnol. 2011, 47, S91–S105. [Google Scholar] [CrossRef]
- Jacobsen, D.; Cressa, C.; Mathooko, J.M.; Dudgeon, D. Macroinvertebrates: Composition, life histories and production. In Aquatic Ecosystems: Tropical Stream Ecology; Dudgeon, D., Ed.; Elsevier Science: London, UK, 2008; pp. 65–105. [Google Scholar]
- Hoang, D.H.; Bae, Y.J. Aquatic insect diversity in a tropical Vietnamese stream in comparison with that in a temperate Korean stream. Limnology 2006, 7, 45–55. [Google Scholar] [CrossRef]
- Cao, Y.; Williams, D.D.; Williams, N.E. How important rare species in aquatic community ecology and bioassessment? Limnol. Oceanogr. 1998, 43, 1403–1409. [Google Scholar] [CrossRef]
- Nijboer, R.C.; Schmidt-Kloiber, A. The effect of excluding taxa with low abundances or taxa with small distribution ranges on ecological assessment. Hydrobiologia 2004, 516, 347–363. [Google Scholar] [CrossRef]
- Braccia, A.; Voshell, J.R. Environmental factors accounting for benthic macroinvertebrate assemblage structure at the sample scale in streams subjected to a gradient of cattle grazing. Hydrobiologia 2006, 573, 55–73. [Google Scholar] [CrossRef]
- Soininen, J.; Paavola, R.; Muotka, T. Benthic diatom communities in boreal streams: Community structure in relation to environmental and spatial gradients. Ecography 2004, 27, 330–342. [Google Scholar] [CrossRef]
- Mikulyuk, A.; Sharma, S.; Egeren, S.V.; Erdmann, E.; Nault, M.E.; Hauxwell, J. The relative role of environmental, spatial, and land-use patterns in explaining aquatic macrophyte community composition. Can. J. Fish. Aquat. Sci. 2011, 68, 1778–1789. [Google Scholar] [CrossRef]
- Stomp, M.; Huisman, J.; Mittelbach, G.G.; Litchman, E.; Klausmeier, C.A. Large-scale biodiversity patterns in freshwater phytoplankton. Ecology 2011, 92, 2096–2107. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, A.M.; Quintino, V.; Sampaio, L.; Freitas, R.; Neves, R. Benthic biodiversity patterns in Ria de Aveiro, Western Portugal: Environmental-biological relationships. Estuar. Coast. Shelf Sci. 2011, 95, 338–348. [Google Scholar] [CrossRef]
- Waite, I.R.; Carpenter, K.D. Associations among fish assemblage structure and environmental variables in Willamette Basin streams, Oregon. Trans. Am. Fish. Soc. 2000, 129, 754–770. [Google Scholar] [CrossRef]
- Vannote, R.L.; Minshall, G.W.; Cummins, K.W.; Sedell, J.R.; Cushing, C.E. The river continuum concept. Can. J. Fish. Aquat. Sci. 1980, 37, 130–137. [Google Scholar] [CrossRef]
- Beauchard, O.; Gagneur, J.; Brosse, S. Macroinvertebrate richness patterns in North African streams. J. Biogeogr. 2003, 30, 1821–1833. [Google Scholar] [CrossRef]
- Pan, Y.; Hill, B.H.; Husby, P.; Hall, R.K.; Kaufmann, P.R. Relationships between environmental variables and benthic diatom assemblages in California Central Valley streams (USA). Hydrobiologia 2006, 561, 119–130. [Google Scholar] [CrossRef]
- Harding, J.S.; Young, R.G.; Hayes, J.W.; Shearer, K.A.; Stark, J.D. Changes in agricultural intensity and river health along ariver continuum. Freshw. Biol. 1999, 42, 345–357. [Google Scholar] [CrossRef]
- Boyero, L.; Bosch, J. The effect of riffle-scale environmental variability on macroinvertebrate assemblages in a tropical stream. Hydrobiologia 2004, 524, 125–132. [Google Scholar] [CrossRef]
- Potapova, M.G.; Charles, D.F. Benthic diatoms in USA rivers: Distributions along spatial and environmental gradients. J. Biogeogr. 2002, 29, 167–187. [Google Scholar] [CrossRef]
- Halwas, K.; Church, M. Channel units in small, high gradient streams on Vancouver Island, British Columbia. Geomorphology 2002, 43, 243–256. [Google Scholar] [CrossRef]
- Jun, Y.-C.; Kim, N.-Y.; Kwon, S.-J.; Han, S.-C.; Hwang, I.-C.; Park, J.-H.; Won, D.-H.; Byun, M.-S.; Kong, H.-Y.; Lee, J.-E.; et al. Effects of land use on benthic macroinvertebrate communities: Comparison of two mountain streams in Korea. Ann. Limnol. Int. J. Limnol. 2011, 47, S35–S49. [Google Scholar] [CrossRef]
- Parsons, M.; Norris, R.H. The effect of habitat-specific sampling on biological assessment of water quality using a predictive model. Freshw. Biol. 1996, 36, 416–434. [Google Scholar] [CrossRef]
- Gerth, W.J.; Herlihy, A.T. Effect of sampling different habitat types in regional macroinvertebrate bioassessment surveys. J. N. Am. Benthol. Soc. 2006, 25, 501–512. [Google Scholar] [CrossRef]
- Gilles, C.L.; Hose, G.C.; Turak, E. What do qualitative rapid assessment collections of macroinvertebrates represent? A comparison with extensive quantitative sampling. Environ. Monit. Assess. 2009, 149, 99–112. [Google Scholar] [CrossRef] [PubMed]
- Pashkevich, A.; Pavluk, T.; de Vaate, A.B. Efficiency of a standardized artificial substrate for biological monitoring of river water quality. Environ. Monit. Assess. 1996, 40, 143–156. [Google Scholar] [CrossRef] [PubMed]
- Saliu, J.K.; Ovuorie, U.R. The artificial substrate preference of invertebrates in Ogbe Creek, Lagos, Nigeria. Life Sci. J. 2007, 4, 77–81. [Google Scholar]
- Di Sabatino, A.; Cristiano, G.; Pinna, M.; Lombardo, P.; Miccoli, F.P.; Marini, G.; Vignini, P.; Cicolani, B. Structure, functional organization and biological traits of macroinvertebrate assemblages from leaf-bags and benthic samples in a third-order stream of Central Apennines (Italy). Ecol. Indic. 2014, 46, 84–91. [Google Scholar] [CrossRef]
- Vlek, H.E.; Šporka, F.; Krno, I. Influence of macroinvertebrate sample size on bioassessment of streams. Hydrobiologia 2006, 566, 523–542. [Google Scholar] [CrossRef]
- Carter, J.L.; Resh, V.H. After site selection and before data analysis: Sampling, sorting, and laboratory procedures used in stream benthic macroinvertebrate monitoring programs by USA state agencies. J. N. Am. Benthol. Soc. 2001, 20, 658–682. [Google Scholar] [CrossRef]
- Metzeling, L.; Miller, J. Evaluation of the sample size used for the rapid bioassessment of rivers using macroinvertebrates. Hydrobiologia 2001, 444, 159–170. [Google Scholar] [CrossRef]
- Kim, A.R.; Oh, M.W.; Kong, D.S. The influence of sample size on environmental assessment using benthic macroinvertebrates. J. Korean. Soc. Water Environ. 2013, 29, 790–798. [Google Scholar]
- Pinna, M.; Marini, G.; Mancinelli, G.; Basset, A. Influence of sampling effort on ecological descriptors and indicators in perturbed and unperturbed conditions: A study case using benthic macroinvertebrates in Mediterranean transitional waters. Ecol. Indic. 2014, 37, 27–39. [Google Scholar] [CrossRef]
- Vinson, M.R.; Hawkins, C.P. Effects of sampling area and subsampling procedure on comparisons of taxa richness among streams. J. N. Am. Benthol. Soc. 1996, 15, 392–399. [Google Scholar] [CrossRef]
- Marini, G.; Pinna, M.; Basset, A.; Mancinelli, G. Estimation of benthic macroinvertebrate taxonomic diversity: Testing the role of sampling effort in a Mediterranean transitional water ecosystem. Transit. Waters Bull. 2013, 7, 28–40. [Google Scholar]
- Buss, D.F.; Borges, E.L. Application of rapid bioassessment protocols (RBP) for benthic macroinvertebrates in Brazil: Comparison between sampling techniques and mesh sizes. Neotrop. Entomol. 2008, 37, 288–295. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jun, Y.-C.; Kim, N.-Y.; Kim, S.-H.; Park, Y.-S.; Kong, D.-S.; Hwang, S.-J. Spatial Distribution of Benthic Macroinvertebrate Assemblages in Relation to Environmental Variables in Korean Nationwide Streams. Water 2016, 8, 27. https://doi.org/10.3390/w8010027
Jun Y-C, Kim N-Y, Kim S-H, Park Y-S, Kong D-S, Hwang S-J. Spatial Distribution of Benthic Macroinvertebrate Assemblages in Relation to Environmental Variables in Korean Nationwide Streams. Water. 2016; 8(1):27. https://doi.org/10.3390/w8010027
Chicago/Turabian StyleJun, Yung-Chul, Nan-Young Kim, Sang-Hun Kim, Young-Seuk Park, Dong-Soo Kong, and Soon-Jin Hwang. 2016. "Spatial Distribution of Benthic Macroinvertebrate Assemblages in Relation to Environmental Variables in Korean Nationwide Streams" Water 8, no. 1: 27. https://doi.org/10.3390/w8010027
APA StyleJun, Y. -C., Kim, N. -Y., Kim, S. -H., Park, Y. -S., Kong, D. -S., & Hwang, S. -J. (2016). Spatial Distribution of Benthic Macroinvertebrate Assemblages in Relation to Environmental Variables in Korean Nationwide Streams. Water, 8(1), 27. https://doi.org/10.3390/w8010027