
Millennial-Scale Carbon Storage in Natural Pine Forests of the North Carolina Lower Coastal Plain: Effects of Artificial Drainage in a Time of Rapid Sea Level Rise

 ,
,  , and
, and 
Abstract
:1. Introduction
2. Methods
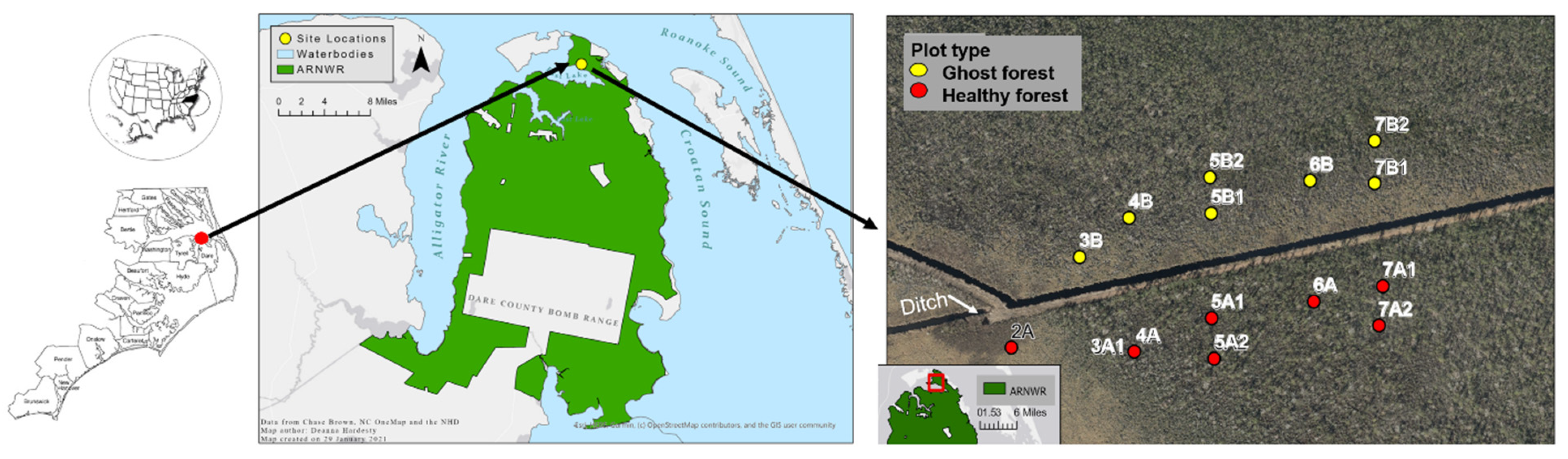
2.1. Study Site
2.2. Overstory Tree Biomass and C Content
2.3. Mid-Story and Understory Biomass and C Content
2.4. Soil Sampling
2.5. Belowground Wood Sampling
2.6. Data Analysis
3. Results
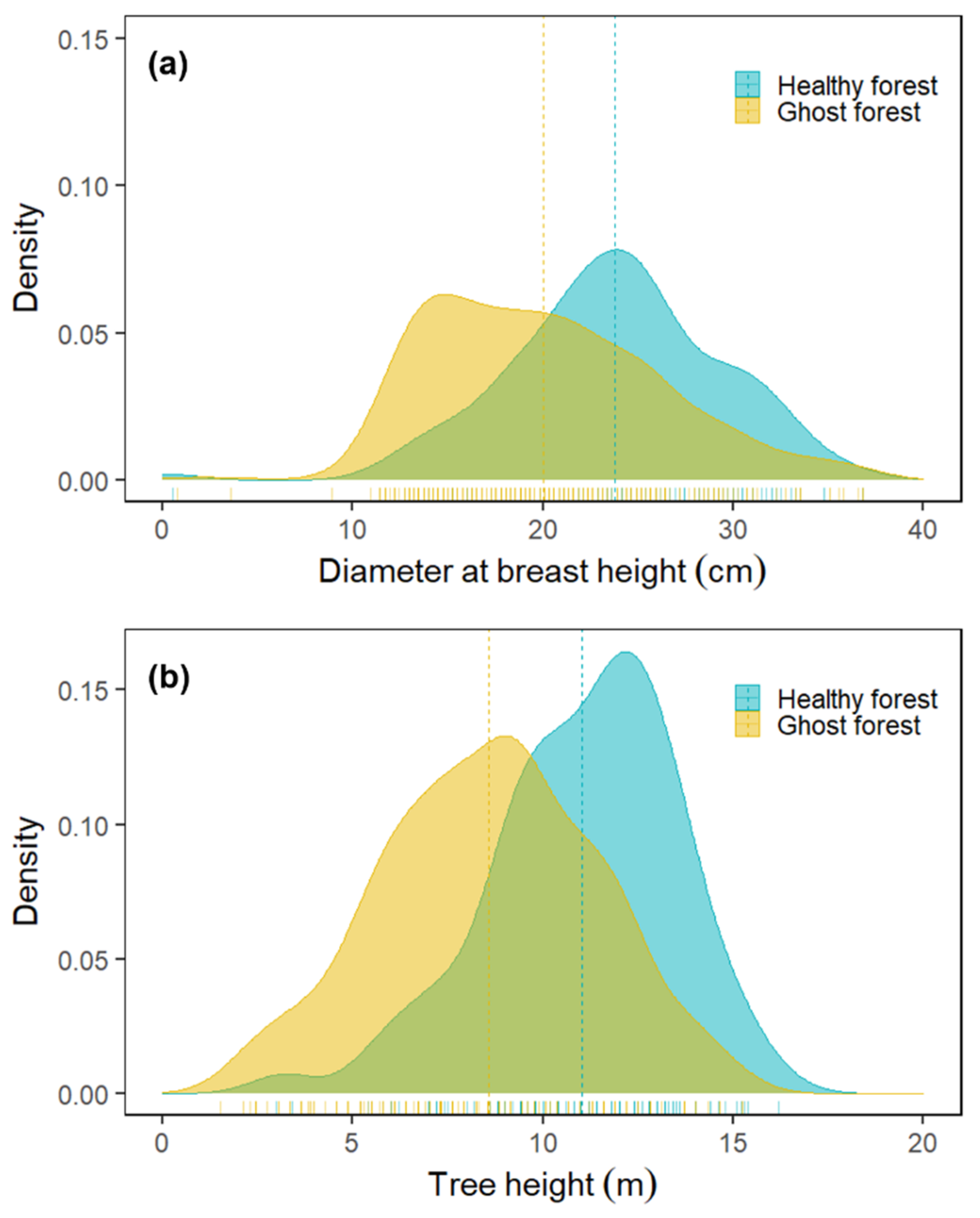
3.1. Overstory Species Composition, Tree Density, Mortality and Diameter
3.2. Overstory, Mid-Story and Understory Biomass
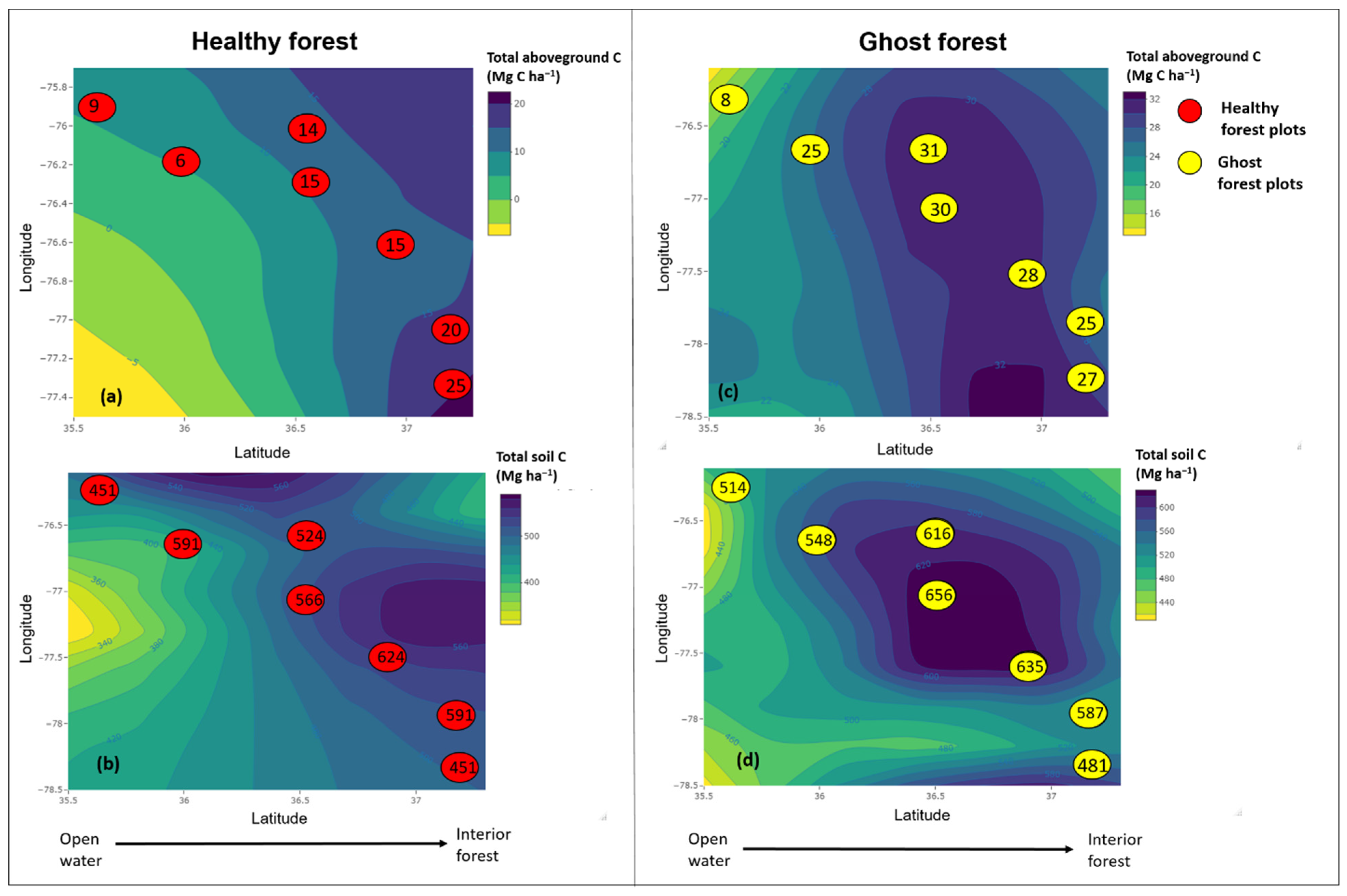
3.3. Ecosystem Carbon Stocks in Vegetation and Soils
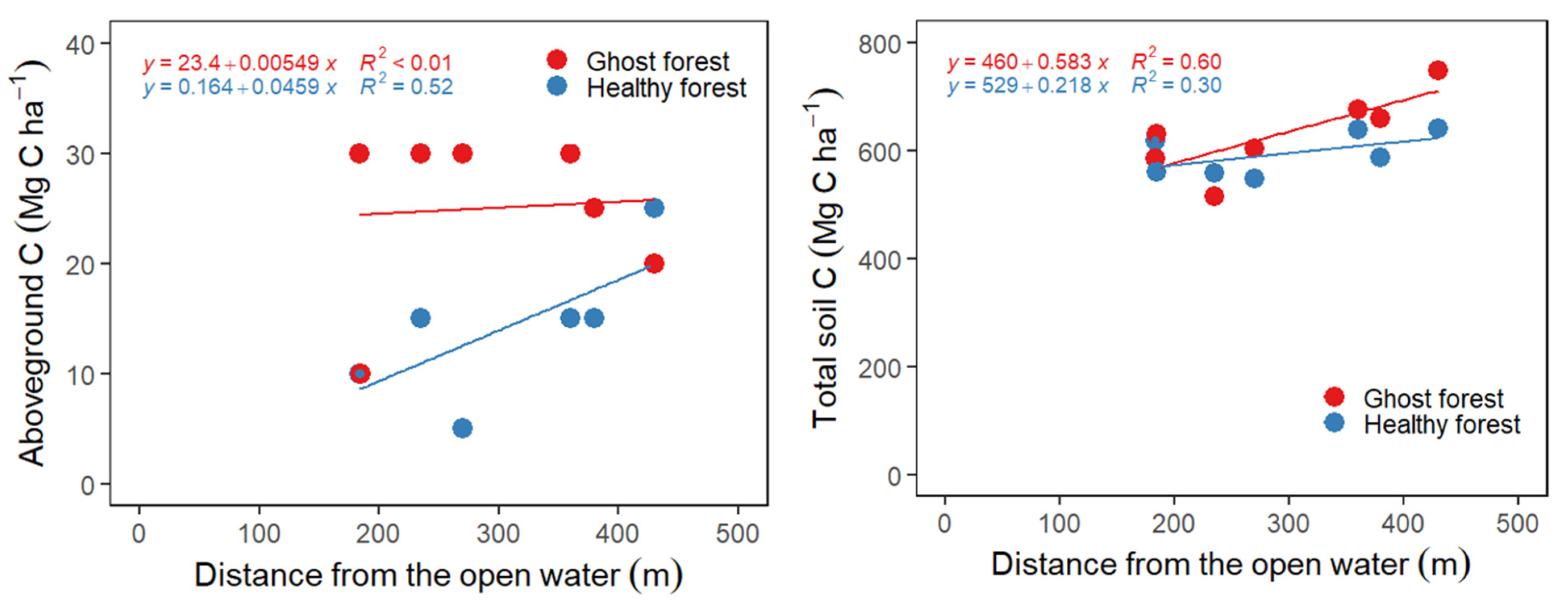
3.4. Change in Overstory C Stocks with Distance from Open Water
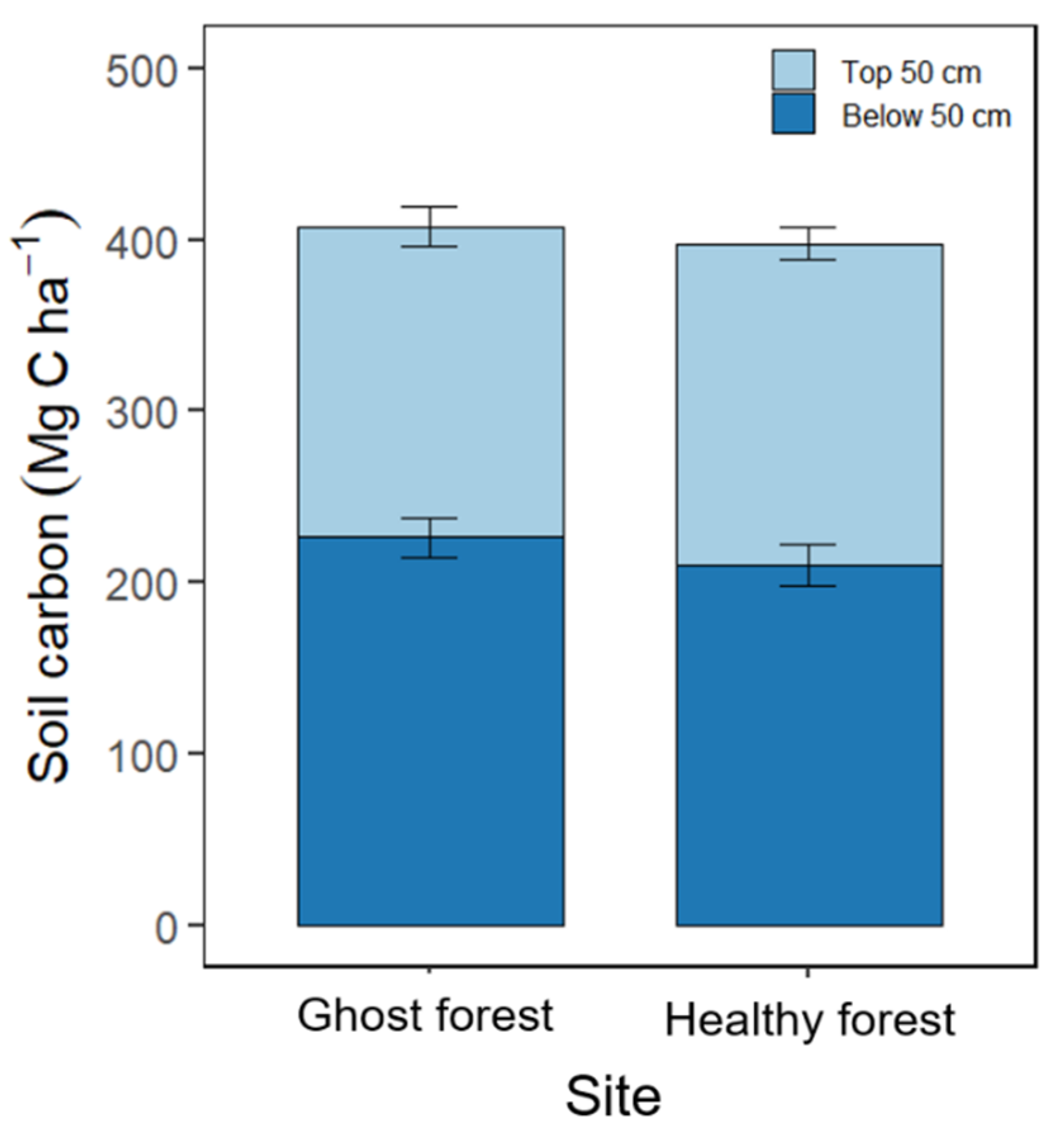
3.5. Soil Carbon Stocks
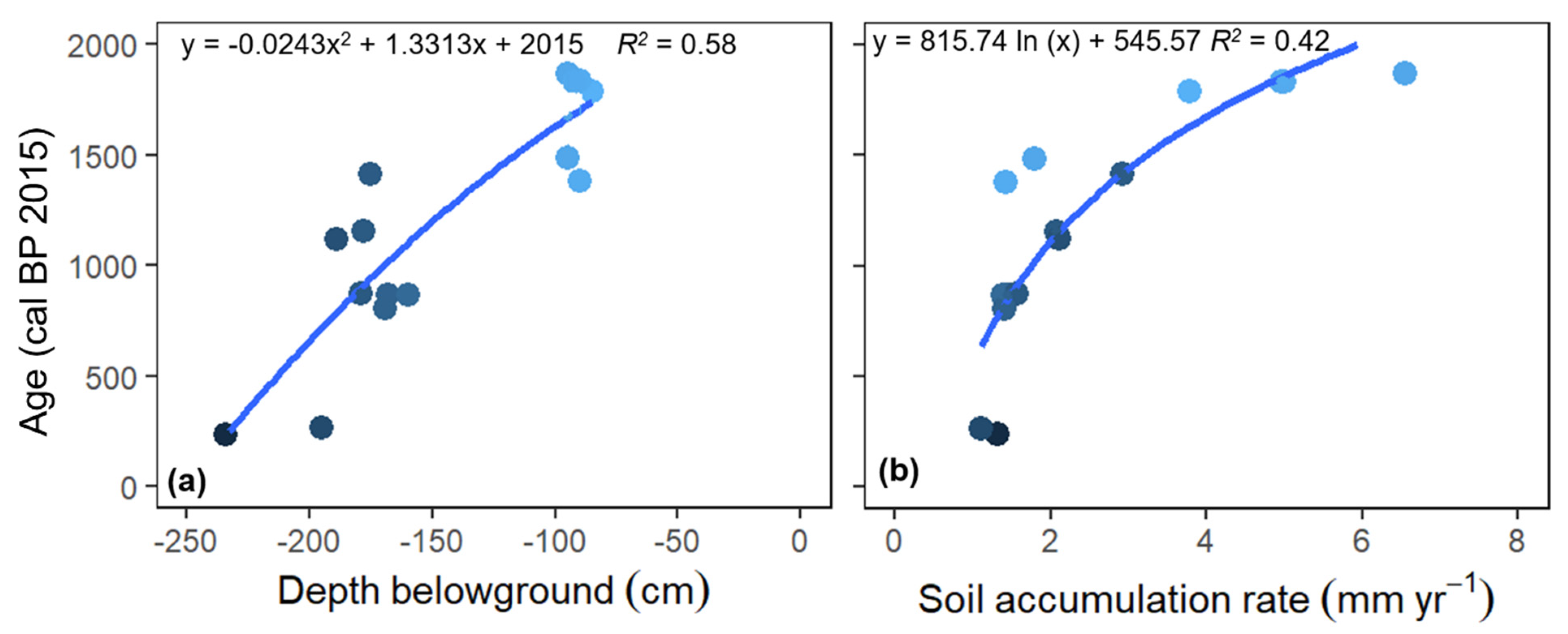
3.6. Wood Sample Carbon Dating and Soil Accumulation
4. Discussion
4.1. Species Composition and Forest Mortality
4.2. Aboveground Biomass and Carbon Content of Vegetation
4.3. Soil Carbon
4.4. Soil Accumulation over Time
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mitra, S.; Wassmann, R.; Vlek, P.L.G. An Appraisal of Global Wetland Area and Its Organic Carbon Stock. Curr. Sci. 2005, 88, 25–35. [Google Scholar]
- Cierjacks, A.; Kleinschmit, B.; Babinsky, M.; Kleinschroth, F.; Markert, A.; Menzel, M.; Ziechmann, U.; Schiller, T.; Graf, M.; Lang, F. Carbon Stocks of Soil and Vegetation on Danubian Floodplains. J. Plant Nutr. Soil Sci. 2010, 173, 644–653. [Google Scholar] [CrossRef]
- Casey, W.P.; Ewel, K.C. Patterns of Succession in Forested Depressional Wetlands in North Florida, USA. Wetlands 2006, 26, 147–160. [Google Scholar] [CrossRef]
- Nahlik, A.M.; Fennessy, M.S. Carbon Storage in US Wetlands. Nat. Commun. 2016, 7, 13835. [Google Scholar] [CrossRef] [Green Version]
- Day, J.W.; Christian, R.R.; Boesch, D.M.; Yáñez-Arancibia, A.; Morris, J.; Twilley, R.R.; Naylor, L.; Schaffner, L.; Stevenson, C. Consequences of Climate Change on the Ecogeomorphology of Coastal Wetlands. Estuaries Coasts 2008, 31, 477–491. [Google Scholar] [CrossRef]
- Horton, B.P.; Rahmstorf, S.; Engelhart, S.E.; Kemp, A.C. Expert Assessment of Sea-Level Rise by AD 2100 and AD 2300. Quat. Sci. Rev. 2014, 84, 1–6. [Google Scholar] [CrossRef]
- Cormier, N.; Krauss, K.W.; Conner, W.H. Periodicity in Stem Growth and Litterfall in Tidal Freshwater Forested Wetlands: Influence of Salinity and Drought on Nitrogen Recycling. Estuaries Coasts 2013, 36, 533–546. [Google Scholar] [CrossRef]
- Ensign, S.H.; Hupp, C.R.; Noe, G.B.; Krauss, K.W.; Stagg, C.L. Sediment Accretion in Tidal Freshwater Forests and Oligohaline Marshes of the Waccamaw and Savannah Rivers, USA. Estuaries Coasts 2014, 37, 1107–1119. [Google Scholar] [CrossRef]
- Truus, L. Estimation of Above-Ground Biomass of Wetlands. In Biomass and Remote Sensing of Biomass; IntechOpen: London, UK, 2011. [Google Scholar] [CrossRef] [Green Version]
- Stagg, C.L.; Schoolmaster, D.R.; Piazza, S.C.; Snedden, G.; Steyer, G.D.; Fischenich, C.J.; McComas, R.W. A Landscape-Scale Assessment of Above- and Belowground Primary Production in Coastal Wetlands: Implications for Climate Change-Induced Community Shifts. Estuaries Coasts 2017, 40, 856–879. [Google Scholar] [CrossRef]
- Rasmussen, C.; Southard, R.J.; Horwath, W.R. Litter Type and Soil Minerals Control Temperate Forest Soil Carbon Response to Climate Change. Glob. Chang. Biol. 2008, 14, 2064–2080. [Google Scholar] [CrossRef]
- Kirwan, M.L.; Gedan, K.B. Sea-Level Driven Land Conversion and the Formation of Ghost Forests. Nat. Clim. Chang. 2019, 9, 450–457. [Google Scholar] [CrossRef] [Green Version]
- Hopkinson, C.S.; Lugo, A.E.; Alber, M.; Covich, A.P.; Van Bloem, S.J. Forecasting Effects of Sea-Level Rise and Windstorms on Coastal and Inland Ecosystems. Front. Ecol. Environ. 2008, 6, 255–263. [Google Scholar] [CrossRef] [Green Version]
- Neumann, B.; Vafeidis, A.T.; Zimmermann, J.; Nicholls, R.J. Future Coastal Population Growth and Exposure to Sea-Level Rise and Coastal Flooding—A Global Assessment. PLoS ONE 2015, 10, e0118571. [Google Scholar] [CrossRef] [Green Version]
- White, E.; Kaplan, D. Restore or Retreat? Saltwater Intrusion and Water Management in Coastal Wetlands. Ecosyst. Health Sustain. 2017, 3, e01258. [Google Scholar] [CrossRef] [Green Version]
- Elsey-Quirk, T.; Seliskar, D.M.; Sommerfield, C.K.; Gallagher, J.L. Salt Marsh Carbon Pool Distribution in a Mid-Atlantic Lagoon, USA: Sea Level Rise Implications. Wetlands 2011, 31, 87–99. [Google Scholar] [CrossRef]
- Dreyer, G.D.; Niering, W.A. Bulletin No. 34: Tidal Marshes of Long Island Sound: Ecology, History and Restoration. Human Impacts on Tidal Wetlands: History and Regulations; Connecticut College Arboretum: New London, CT, USA, 1995; ISBN 1878899058. [Google Scholar]
- Bryant, M.R.; Jerome, P.; Andrew, J.; Hamilton, S. Alligator River National Wildlife Refuge Comprehensive Conservation Plan; Fish and Wildlife Service, US Department of the Interior: Atlanta, GA, USA, 2008. [Google Scholar]
- McKee, K.L. Biophysical Controls on Accretion and Elevation Change in Caribbean Mangrove Ecosystems. Estuar. Coast. Shelf Sci. 2011, 91, 475–483. [Google Scholar] [CrossRef]
- Morris, J.T.; Sundareshwar, P.V.; Nietch, C.T.; Kjerfve, B.; Cahoon, D.R. Responses of Coastal Wetlands to Rising Sea Level. Ecology 2002, 83, 2869–2877. [Google Scholar] [CrossRef]
- Kirwan, M.L.; Guntenspergen, G.R. Response of Plant Productivity to Experimental Flooding in a Stable and a Submerging Marsh. Ecosystems 2015, 18, 903–913. [Google Scholar] [CrossRef]
- Pennings, S.C.; Grant, M.B.; Bertness, M.D. Plant Zonation in Low-Latitude Salt Marshes: Disentangling the Roles of Flooding, Salinity and Competition. J. Ecol. 2005, 93, 159–167. [Google Scholar] [CrossRef]
- Holmquist, J.R.; Windham-Myers, L.; Bliss, N.; Crooks, S.; Morris, J.T.; Megonigal, J.P.; Troxler, T.; Weller, D.; Callaway, J.; Drexler, J.; et al. Accuracy and Precision of Tidal Wetland Soil Carbon Mapping in the Conterminous United States. Sci. Rep. 2018, 8, 9478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bridgham, S.D.; Megonigal, J.P.; Keller, J.K.; Bliss, N.B.; Trettin, C. The Carbon Balance of North American Wetlands. Wetlands 2006, 26, 889–916. [Google Scholar] [CrossRef]
- McTigue, N.; Davis, J.; Rodriguez, A.B.; McKee, B.; Atencio, A.; Currin, C. Sea Level Rise Explains Changing Carbon Accumulation Rates in a Salt Marsh Over the Past Two Millennia. J. Geophys. Res. Biogeosci. 2019, 124, 2945–2957. [Google Scholar] [CrossRef]
- Riggs, S.R.; Ames, D. Drowning the North Carolina Coast: Sea-Level Rise and Estuarine Dynamics; NC Sea Grant: Raleigh, NC, USA, 2003. [Google Scholar]
- Riggs, S.R.; Ames, D.V.; Culver, S.J.; Mallinson, D.J. Review Reviewed Work(s): The Battle for North Carolina’s Coast: Evolutionary History, Present Crisis, and Vision for the Future. Southeast. Geogr. 2012, 52, 242–244. [Google Scholar] [CrossRef]
- Minick, K.J.; Kelley, A.M.; Miao, G.; Li, X.; Noormets, A.; Mitra, B.; King, J.S. Microtopography Alters Hydrology, Phenol Oxidase Activity and Nutrient Availability in Organic Soils of a Coastal Freshwater Forested Wetland. Wetlands 2019, 39, 263–273. [Google Scholar] [CrossRef]
- Burns, R.M.; Honkala, B.H. (Eds.) Bramlett DL Pond pine (Pinus serotinaMichx). In The Silvics of North America—Agricultural Handbook 654; USDA Forest Service: Washington, DC, USA, 1990. [Google Scholar]
- FIA. Phase 3Field Guide—Down Woody Materials, Section 25; Down Woody Materials; USDA: Kansas City, MO, USA, 2007. [Google Scholar]
- Jenkins, J.C.; Chojnacky, D.C.; Heath, L.S.; Birdsey, R.A. Comprehensive Database of Diameter-Based Biomass Resgressions for North American Tree Species; United States Department of Agriculture, Forest Service, Northeastern Research Station: Kansas City, MO, USA, 2003. [Google Scholar]
- Preston, R.J.; Braham, R.R. North American Trees; Iowa State Press: Ames, IA, USA, 2002. [Google Scholar]
- Hajdas, I.; Hendriks, L.; Fontana, A.; Monegato, G. Evaluation of Preparation Methods in Radiocarbon Dating of Old Wood. Radiocarbon 2017, 59, 727–737. [Google Scholar] [CrossRef]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. [Google Scholar]
- Sievert, C. Interactive Web-Based Data Visualization with R, Plotly, and Shiny; Chapman and Hall/CRC: Boca Raton, FL, USA, 2020; ISBN 9781138331457. [Google Scholar]
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the Tidyverse. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef]
- Wickham, H. Reshaping Data with the Reshape Package. J. Stat. Softw. 2007, 12, 1–20. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Ilomets, M.; Truus, L.; Pajula, R.; Sepp, K. Species Composition and Structure of Vascular Plants and Bryophytes on the Water Level Gradient within a Calcareous Fen North Estonia. Est. J. Ecol. 2010, 59, 19–38. [Google Scholar] [CrossRef] [Green Version]
- Wilcox, D.A.; Nichols, S.J. The Effects of Water-Level Fluctuations on Vegetation in a Lake Huron Wetland. Wetlands 2008, 28, 487–501. [Google Scholar] [CrossRef]
- Aguilos, M.; Mitra, B.; Noormets, A.; Minick, K.; Prajapati, P.; Gavazzi, M.; Sun, G.; McNulty, S.; Li, X.; Domec, J.C.; et al. Long-Term Carbon Flux and Balance in Managed and Natural Coastal Forested Wetlands of the Southeastern USA. Agric. For. Meteorol. 2020, 288–289, 108022. [Google Scholar] [CrossRef]
- Craine, S.I.; Orians, C.M. Pitch Pine (Pinus Rigida Mill.) Invasion of Cape Cod Pond Shores Alters Abiotic Environment and Inhibits Indigenous Herbaceous Species. Biol. Conserv. 2004, 116, 181–189. [Google Scholar] [CrossRef]
- Williams, K.; Ewel, K.C.; Stumpf, R.P.; Putz, F.E.; Workman, T.W. Sea-Level Rise and Coastal Forest Retreat on the West Coast of Florida, USA. Ecology 1999, 80, 2045–2063. [Google Scholar] [CrossRef]
- Begin, Y. The Effects of Shoreline Transgression on Woody Plants, Upper St Lawrence Estuary, Quebec. J. Coast. Res. 1990, 6, 815–827. [Google Scholar]
- Pezeshki, S.R.; Delaune, R.D.; Patrick, W.H. Flooding and Saltwater Intrusion: Potential Effects on Survival and Productivity of Wetland Forests along the U.S. Gulf Coast. For. Ecol. Manag. 1990, 33–34, 287–301. [Google Scholar] [CrossRef]
- Lennard-Barrett, E.G. The Interaction between Waterlogging and Salinity in Higher Plants: Causes, Consequences and Implications. Plant Soil 2003, 253, 35–54. [Google Scholar] [CrossRef]
- Conner, W.H. The Effect of Salinity and Waterlogging on Growth and Survival of Baldcypress and Chinese Tallow Seedlings; Coastal Education & Research Foundation, Inc.: Lawrence, KS, USA, 1994; Volume 10, pp. 1045–1049. Available online: https://www.Jstor.Org/Stable/4298295 (accessed on 15 August 2021).
- Desantis, L.R.G.; Bhotika, S.; Williams, K.; Putz, F.E. Sea-Level Rise and Drought Interactions Accelerate Forest Decline on the Gulf Coast of Florida, USA. Glob. Chang. Biol. 2007, 13, 2349–2360. [Google Scholar] [CrossRef]
- Poulter, B.; Goodall, J.L.; Halpin, P.N. Applications of Network Analysis for Adaptive Management of Artificial Drainage Systems in Landscapes Vulnerable to Sea Level Rise. J. Hydrol. 2008, 357, 207–217. [Google Scholar] [CrossRef]
- Mendoza-Ponce, A.; Galicia, L. Aboveground and Belowground Biomass and Carbon Pools in Highland Temperate Forest Landscape in Central Mexico. Forestry 2010, 83, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Ile, O.J.; Aguilos, M.; Morkoc, S.; Minick, K.; Domec, J.-C.; King, J.S. Productivity of Low-Input Short-Rotation Coppice American Sycamore (Platanus Occidentalis L.) Grown at Different Planting Densities as a Bioenergy Feedstock over Two Rotation Cycles. Biomass Bioenergy 2021, 146, 105983. [Google Scholar] [CrossRef]
- Aguilos, M.; Sun, G.; Noormets, A.; Domec, J.C.; McNulty, S.; Gavazzi, M.; Prajapati, P.; Minick, K.J.; Mitra, B.; King, J. Ecosystem Productivity and Evapotranspiration Are Tightly Coupled in Loblolly Pine (Pinus Taeda L.) Plantations along the Coastal Plain of the Southeastern U.S. Forests 2021, 12, 1123. [Google Scholar] [CrossRef]
- Aguilos, M.; Sun, G.; Noormets, A.; Domec, J.C.; McNulty, S.; Gavazzi, M.; Minick, K.; Mitra, B.; Prajapati, P.; Yang, Y.; et al. Effects of Land-Use Change and Drought on Decadal Evapotranspiration and Water Balance of Natural and Managed Forested Wetlands along the Southeastern US Lower Coastal Plain. Agric. For. Meteorol. 2021, 303, 108381. [Google Scholar] [CrossRef]
- Minick, K.J.; Mitra, B.; Li, X.; Fischer, M.; Aguilos, M.; Prajapati, P.; Noormets, A.; King, J.S. Wetland Microtopography Alters Response of Potential Net CO2 and CH4 Production to Temperature and Moisture: Evidence from a Laboratory Experiment. Geoderma 2021, 402, 115367. [Google Scholar] [CrossRef]
- Ketcheson, S.J.; Price, J.S.; Carey, S.K.; Petrone, R.M.; Mendoza, C.A.; Devito, K.J. Constructing Fen Peatlands in Post-Mining Oil Sands Landscapes: Challenges and Opportunities from a Hydrological Perspective. Earth-Sci. Rev. 2016, 161, 130–139. [Google Scholar] [CrossRef]
- Lamers, L.P.M.; van Diggelen, J.M.H.; Op Den Camp, H.J.M.; Visser, E.J.W.; Lucassen, E.C.H.E.T.; Vile, M.A.; Jetten, M.S.M.; Smolders, A.J.P.; Roelofs, J.G.M. Microbial Transformations of Nitrogen, Sulfur, and Iron Dictate Vegetation Composition in Wetlands: A Review. Front. Microbiol. 2012, 3, 156. [Google Scholar] [CrossRef] [Green Version]
- Harpenslager, S.F.; van den Elzen, E.; Kox, M.A.R.; Smolders, A.J.P.; Ettwig, K.F.; Lamers, L.P.M. Rewetting Former Agricultural Peatlands: Topsoil Removal as a Prerequisite to Avoid Strong Nutrient and Greenhouse Gas Emissions. Ecol. Eng. 2015, 84, 159–168. [Google Scholar] [CrossRef]
- Sarneel, J.M.; Geurts, J.J.M.; Beltman, B.; Lamers, L.P.M.; Nijzink, M.M.; Soons, M.B.; Verhoeven, J.T.A. The Effect of Nutrient Enrichment of Either the Bank or the Surface Water on Shoreline Vegetation and Decomposition. Ecosystems 2010, 13, 1275–1286. [Google Scholar] [CrossRef] [Green Version]
- Dee, S.M.; Ahn, C. Plant Tissue Nutrients as a Descriptor of Plant Productivity of Created Mitigation Wetlands. Ecol. Indic. 2014, 45, 68–74. [Google Scholar] [CrossRef]
- Overbeek, C.C.; Harpenslager, S.F.; van Zuidam, J.P.; van Loon, E.E.; Lamers, L.P.M.; Soons, M.B.; Admiraal, W.; Verhoeven, J.T.A.; Smolders, A.J.P.; Roelofs, J.G.M.; et al. Drivers of Vegetation Development, Biomass Production and the Initiation of Peat Formation in a Newly Constructed Wetland. Ecosystems 2020, 23, 1019–1036. [Google Scholar] [CrossRef] [Green Version]
- Suchar, V.A.; Crookston, N.L. Understory Cover and Biomass Indices Predictions for Forest Ecosystems of the Northwestern United States. Ecol. Indic. 2010, 10, 602–609. [Google Scholar] [CrossRef]
- Kumar, P.; Chen, H.Y.H.; Thomas, S.C.; Shahi, C. Linking Resource Availability and Heterogeneity to Understorey Species Diversity through Succession in Boreal Forest of Canada. J. Ecol. 2018, 106, 1266–1276. [Google Scholar] [CrossRef]
- Litton, C.M.; Ryan, M.G.; Knight, D.H. Effects of Tree Density and Stand Age on Carbon Allocation Patterns in Postfire Lodgepole Pine. Ecol. Appl. 2004, 14, 460–475. [Google Scholar] [CrossRef] [Green Version]
- Ricker, M.C.; Lockaby, B.G. Soil Organic Carbon Stocks in a Large Eutrophic Floodplain Forest of the Southeastern Atlantic Coastal Plain, USA. Wetlands 2015, 35, 291–301. [Google Scholar] [CrossRef]
- Davis, A.A.; Stolt, M.H.; Compton, J.E. Spatial Distribution of Soil Carbon in Southern New England Hardwood Forest Landscapes. Soil Sci. Soc. Am. J. 2004, 68, 895–903. [Google Scholar] [CrossRef]
- Gleason, R.A.; Laubhan, M.K.; Euliss, N.H. Ecosystems Services Derived from Wetland Conservation Practices in the United States Prairie Pothole Region with an emphasis on the U.S. Department Of Agriculture Conservation Reserve and Wetlands Reserve Programs; U.S. Geological Survey Professional Paper 1745; U.S. Geological Survey: Liston, VA, USA, 2008; 58p, ISBN 9781411320178. [Google Scholar]
- Johnston, C.A. Wetland Losses Due to Row Crop Expansion in the Dakota Prairie Pothole Region. Wetlands 2013, 33, 175–182. [Google Scholar] [CrossRef]
- Mudd, S.M.; Howell, S.M.; Morris, J.T. Impact of Dynamic Feedbacks between Sedimentation, Sea-Level Rise, and Biomass Production on near-Surface Marsh Stratigraphy and Carbon Accumulation. Estuar. Coast. Shelf Sci. 2009, 82, 377–389. [Google Scholar] [CrossRef]
- Kirwan, M.L.; Temmerman, S.; Skeehan, E.E.; Guntenspergen, G.R.; Fagherazzi, S. Overestimation of Marsh Vulnerability to Sea Level Rise. Nat. Clim. Chang. 2016, 6, 253–260. [Google Scholar] [CrossRef]
- Crosby, S.C.; Sax, D.F.; Palmer, M.E.; Booth, H.S.; Deegan, L.A.; Bertness, M.D.; Leslie, H.M. Salt Marsh Persistence Is Threatened by Predicted Sea-Level Rise. Estuar. Coast. Shelf Sci. 2016, 181, 93–99. [Google Scholar] [CrossRef] [Green Version]
- Kirwan, M.L.; Megonigal, J.P. Tidal Wetland Stability in the Face of Human Impacts and Sea-Level Rise. Nature 2013, 504, 53–60. [Google Scholar] [CrossRef]
- Kemp, A.C.; Kegel, J.J.; Culver, S.J.; Barber, D.C.; Mallinson, D.J.; Leorri, E.; Bernhardt, C.E.; Cahill, N.; Riggs, S.R.; Woodson, A.L.; et al. Extended Late Holocene Relative Sea-Level Histories for North Carolina, USA. Quat. Sci. Rev. 2017, 160, 13–30. [Google Scholar] [CrossRef] [Green Version]
- Kopp, R.E. Does the Mid-Atlantic United States Sea Level Acceleration Hot Spot Reflect Ocean Dynamic Variability? Geophys. Res. Lett. 2013, 40, 3981–3985. [Google Scholar] [CrossRef]
- Schuerch, M.; Spencer, T.; Temmerman, S.; Kirwan, M.L.; Wolff, C.; Lincke, D.; McOwen, C.J.; Pickering, M.D.; Reef, R.; Vafeidis, A.T.; et al. Future Response of Global Coastal Wetlands to Sea-Level Rise. Nature 2018, 561, 231–234. [Google Scholar] [CrossRef]
- Johnson, B.J.; Moore, K.A.; Lehmann, C.; Bohlen, C.; Brown, T.A. Middle to Late Holocene Fluctuations of C3 and C4 Vegetation in a Northern New England Salt Marsh, Sprague Marsh, Phippsburg Maine. Org. Geochem. 2007, 38, 394–403. [Google Scholar] [CrossRef] [Green Version]
- Brevik, E.C.; Homburg, J.A. A 5000 Year Record of Carbon Sequestration from a Coastal Lagoon and Wetland Complex, Southern California, USA. Catena 2004, 57, 221–232. [Google Scholar] [CrossRef]
- Drexler, J.Z. Peat Formation Processes through the Millennia in Tidal Marshes of the Sacramento-San Joaquin Delta, California, USA. Estuaries Coasts 2011, 34, 900–911. [Google Scholar] [CrossRef]
- Telford, R.J.; Heegaard, E.; Birks, H.J.B. All Age-Depth Models Are Wrong: But How Badly? Quat. Sci. Rev. 2004, 23, 1–5. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Live Biomass Regressions | ||
|---|---|---|
| Species | Allometric Equation | Notes |
| Pond pine (P. serotina) | y = exp(2.0171 + 2.3373 × ln(dbh)) | Equation for P. rigida |
| Red maple (A. rubrum) | y = 2.52363 × (dbh2)1.9648 | |
| Sweetgum (L.sty raciflua) | y = 1.822108 × (dbh2)1.2635 | |
| Bay (Persea spp.) | y = −13.388 + 6.82 × (dbh2) | |
| FIA decay class | Qualification | C remaining (%) |
| 0 | Alive | 100 |
| 1 | Dead, small branches attached, knife does not penetrate | 97 |
| 2 | Dead, no small branches, has large branches, little penetration | 97 |
| 3 | Dead, no large branches, height intact, more penetration | 86 |
| 4 | Dead, top broken, easy penetration, highly decayed | 53 |
| Location | General Species Composition | No. of Trees Per ha−1 | * Decay Class of Standing Dead Trees | ||
|---|---|---|---|---|---|
| Healthy/No Decay | Standing Dead/Dying | Total (Healthy + Standing Dead) | |||
| Ghost forest | Swamp bay | 176 | 210 | 388 | 2 |
| Maple | 12 | 24 | 36 | 1 | |
| Pitch Pine | 85 | 178 | 263 | 2 | |
| Total | 273 a | 412 a | 687 a | ||
| Healthy forest | Maple | 63 | 2 | 65 | 1 |
| Pitch Pine | 148 | 42 | 190 | 1 | |
| Sweetgum | 10 | 0 | 10 | ||
| Total | 220 b | 44 b | 265 b | ||
| Species | Overstory Biomass (Mg ha−1) | Mid-Story Biomass (Mg ha−1) | Under-Story Biomass (Mg ha−1) | ||||
|---|---|---|---|---|---|---|---|
| Alive | Standing Dead | Total BIOMASS (Alive + Dead) | |||||
| Ghost forest | Swamp bay | 16.35 | 10.08 | 26.43 | |||
| Maple | 2.69 | 1.42 | 4.11 | ||||
| Pine | 11.87 | 13.57 | 25.43 | ||||
| Total | 30.91 a ± 6.96 | 25.06 a ± 6.26 | 55.97 a ± 12.61 | 3.65 a ± 2.70 | 8.32 a ± 4.1 | ||
| Healthy forest | Maple | 8.45 | 0.23 | 8.68 | |||
| Pine | 15.42 | 2.96 | 18.38 | ||||
| Sweetgum | 0.93 | 0.00 | 0.93 | ||||
| Total | 24.81 b ± 7.25 | 3.19 b ± 1.65 | 27.99 b ± 8.75 | 14.06 b ± 7.30 | 7.41 a ± 1.28 | ||
| Soil Depth (cm) | Age (Apprx. Calendar Year BP 2015) | Soil Accumulation Rate (mm/Year) |
|---|---|---|
| −85 | 1790 | 3.78 |
| −90 | 1380 | 1.42 |
| −90 | 1835 | 5.00 |
| −92 | 1830 | 4.97 |
| −95 | 1485 | 1.79 |
| −95 | 1870 | 6.55 |
| −160 | 865 | 1.39 |
| −168 | 865 | 1.46 |
| −169 | 805 | 1.40 |
| −175 | 1415 | 2.92 |
| −178 | 1155 | 2.07 |
| −179 | 875 | 1.57 |
| −189 | 1120 | 2.11 |
| −190 | 1775 | 7.92 |
| −195 | 265 | 1.11 |
| −234 | 235 | 1.31 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aguilos, M.; Brown, C.; Minick, K.; Fischer, M.; Ile, O.J.; Hardesty, D.; Kerrigan, M.; Noormets, A.; King, J. Millennial-Scale Carbon Storage in Natural Pine Forests of the North Carolina Lower Coastal Plain: Effects of Artificial Drainage in a Time of Rapid Sea Level Rise. Land 2021, 10, 1294. https://doi.org/10.3390/land10121294
Aguilos M, Brown C, Minick K, Fischer M, Ile OJ, Hardesty D, Kerrigan M, Noormets A, King J. Millennial-Scale Carbon Storage in Natural Pine Forests of the North Carolina Lower Coastal Plain: Effects of Artificial Drainage in a Time of Rapid Sea Level Rise. Land. 2021; 10(12):1294. https://doi.org/10.3390/land10121294
Chicago/Turabian StyleAguilos, Maricar, Charlton Brown, Kevan Minick, Milan Fischer, Omoyemeh J. Ile, Deanna Hardesty, Maccoy Kerrigan, Asko Noormets, and John King. 2021. "Millennial-Scale Carbon Storage in Natural Pine Forests of the North Carolina Lower Coastal Plain: Effects of Artificial Drainage in a Time of Rapid Sea Level Rise" Land 10, no. 12: 1294. https://doi.org/10.3390/land10121294
APA StyleAguilos, M., Brown, C., Minick, K., Fischer, M., Ile, O. J., Hardesty, D., Kerrigan, M., Noormets, A., & King, J. (2021). Millennial-Scale Carbon Storage in Natural Pine Forests of the North Carolina Lower Coastal Plain: Effects of Artificial Drainage in a Time of Rapid Sea Level Rise. Land, 10(12), 1294. https://doi.org/10.3390/land10121294