
Quercus rotundifolia Lam. Woodlands of the Southwestern Iberian Peninsula

 ,
, 
 ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
3. Results and Discussion
3.1. Classification of Southwestern Iberian Peninsula Holm Oak Communities
3.2. Description of Holm Oak Communities
3.3. Syntaxonomical Scheme
3.4. Considerations of the Typical Sequence of Vegetation Stages Occurring in the Holm Oak Vegetation Series
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
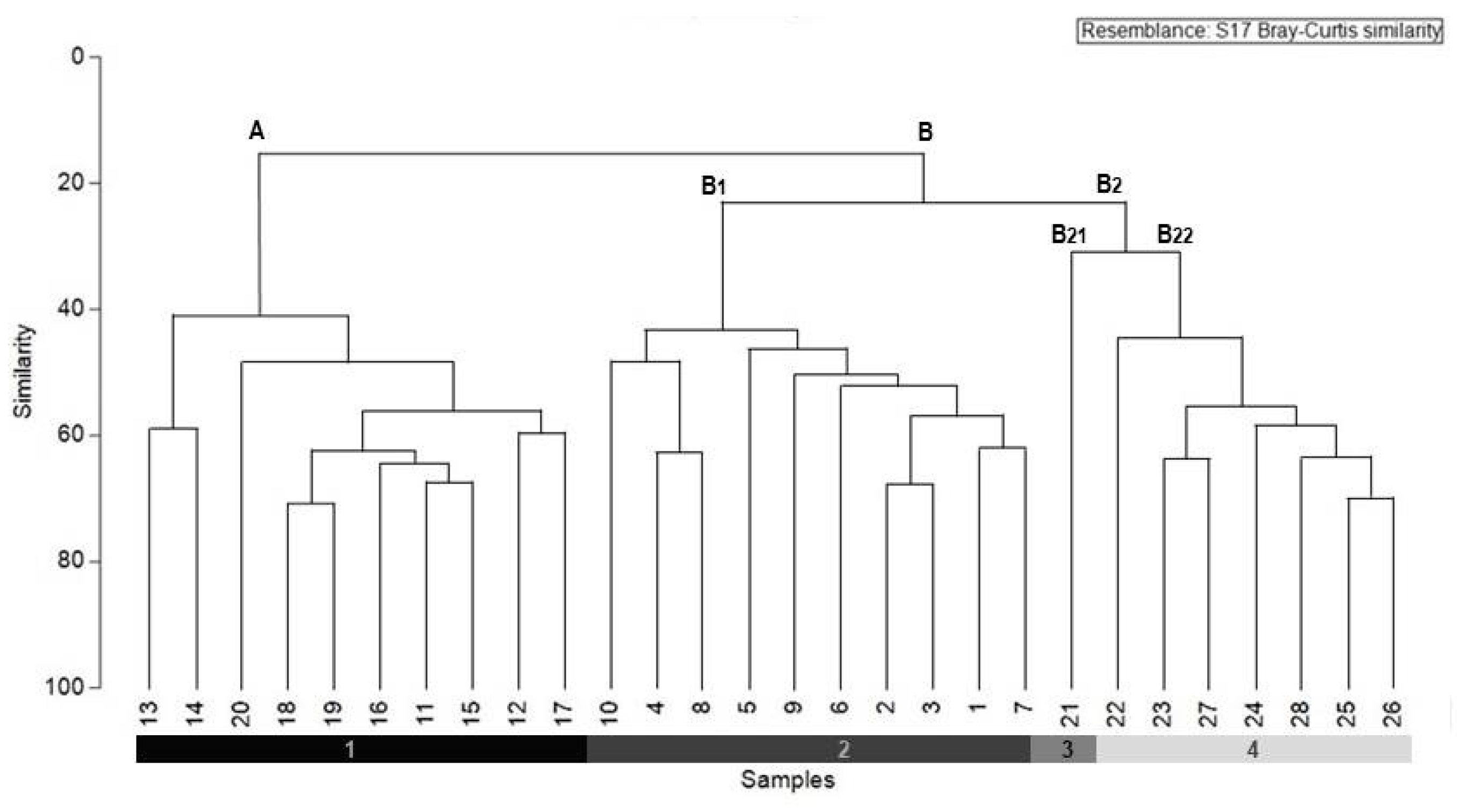{kind=link}
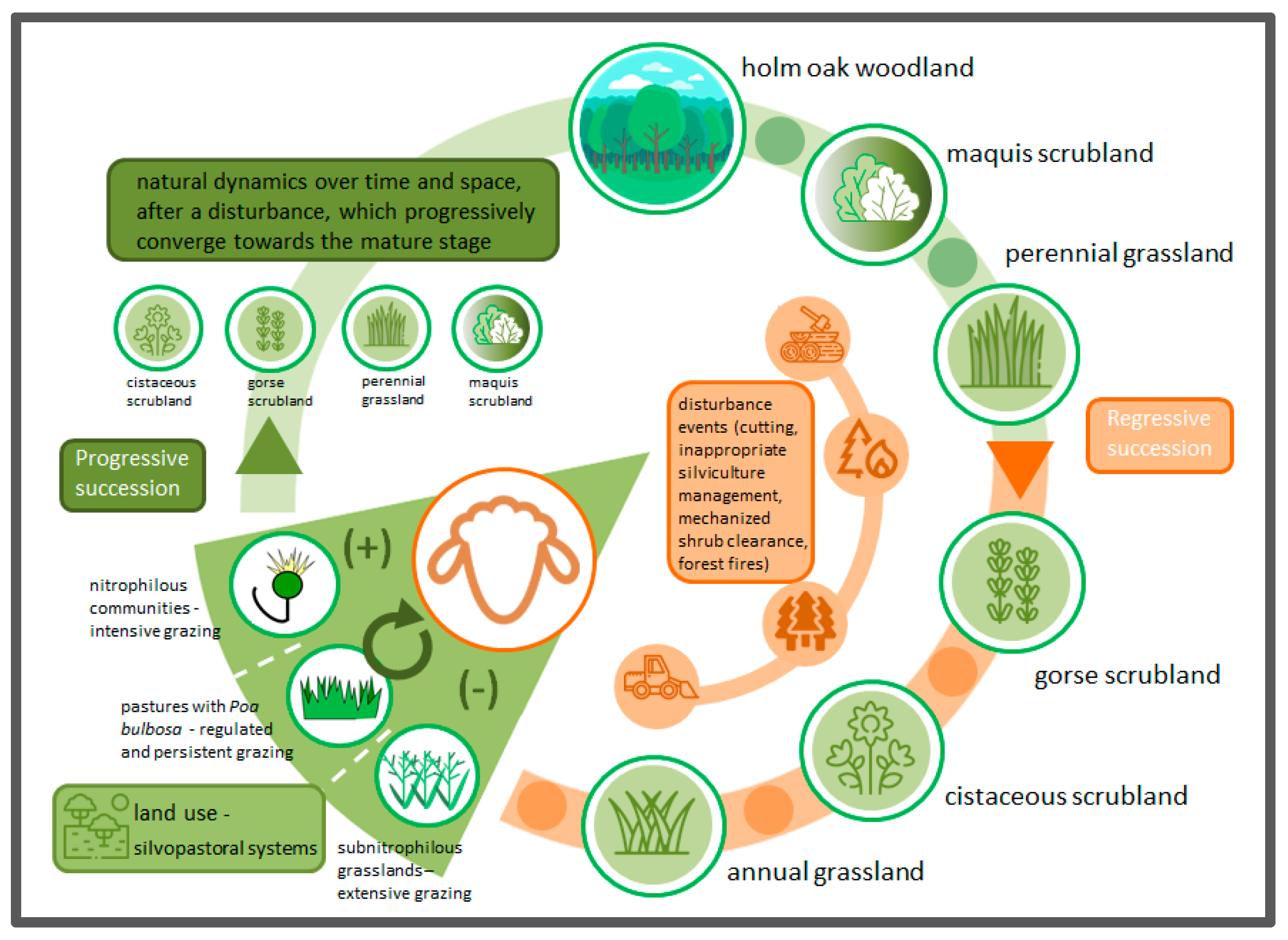{kind=link}
| Association no. | 1 | 2 | 3 | 4 | P |
|---|---|---|---|---|---|
| Characteristic | |||||
| Ulex argenteus | V | - | - | - | 10 |
| Scilla monophyllos | IV | - | - | - | 5 |
| Hyacinthoides hispanica | II | - | - | - | 3 |
| Luzula forsteri subsp. baeticum | II | - | - | - | 3 |
| Avenella stricta | II | - | - | - | 4 |
| Carex hallerana | I | - | - | - | 2 |
| Senecio lopezii | + | - | - | - | 1 |
| Phillyrea media | + | - | - | - | 1 |
| Osyris lanceolata | + | - | - | - | 2 |
| Erica arborea | V | - | - | I | 11 |
| Quercus rotundifolia | V | V | 4 | V | 28 |
| Pistacia lentiscus | V | V | 2 | V | 17 |
| Rhamnus alaternus | IV | II | 2 | II | 13 |
| Rubia peregrina | III | IV | 3 | IV | 13 |
| Olea europaea var. sylvestris | III | II | 2 | V | 17 |
| Arbutus unedo | V | IV | - | IV | 20 |
| Quercus coccifera | V | II | - | V | 21 |
| Smilax aspera var. altissima | IV | III | 1 | - | 6 |
| Phlomis purpurea | IV | I | 1 | - | 14 |
| Phillyrea angustifolia | III | V | - | III | 10 |
| Viburnum tinus | II | + | - | III | 5 |
| Daphne gnidium | I | - | 1 | III | 17 |
| Lonicera implexa | II | - | - | IV | 10 |
| Ruscus aculeatus | II | - | - | III | 8 |
| Asplenium onopteris | II | - | - | I | 3 |
| Pyrus bourgaeana | + | V | - | I | 12 |
| Rhamnus oleoides | + | - | 1 | III | 6 |
| Myrtus communis | I | V | - | - | 12 |
| Arum italicum subsp. neglectum | I | - | - | I | 2 |
| Quercus suber | + | III | - | - | 7 |
| Phillyrea latifolia | + | - | - | III | 3 |
| Arisarum vulgare subsp. simorrhinum | + | - | - | II | 2 |
| Elaeoselinum foetidum | + | - | - | II | 2 |
| Vincetoxicum nigrum | - | III | - | - | 5 |
| Moehringia pentandra | - | + | - | - | 1 |
| Asparagus acutifolius | - | V | 1 | V | 16 |
| Anemone palmata | - | III | - | I | 6 |
| Jasminum fruticans | - | II | - | V | 10 |
| Osyris alba | - | II | - | III | 7 |
| Chamaerops humilis | - | II | + | - | 4 |
| Pistacia terebinthus | - | I | - | V | 9 |
| Aristolochia baetica | - | - | 3 | - | 1 |
| Ulex baeticus subsp. scaber | - | - | 1 | - | 1 |
| Calicotome villosa | - | - | 1 | - | 1 |
| Ceratonia siliqua | - | - | 1 | - | 1 |
| Asparagus aphyllus | - | - | 1 | - | 1 |
| Rosa sempervirens | - | - | + | - | 1 |
| Clematis flammula | - | - | + | - | 1 |
| Rhamnus laderoi | - | - | - | V | 7 |
| Teucrium fruticans | - | - | - | III | 4 |
| Smilax aspera var. aspera | - | - | - | III | 10 |
| Thapsia nitida | - | - | - | III | 3 |
| Bupleurum fruticosum | - | - | - | III | 3 |
| Paeonia broteroi | - | - | - | I | 2 |
| Coronilla juncea | - | - | - | II | 2 |
| Lonicera etrusca | - | - | - | II | 2 |
| Asparagus albus | - | - | - | I | 1 |
| Coronilla glauca | - | - | - | I | 1 |
| Juniperus oxycedrus subsp. lagunae | - | - | - | I | 1 |
| Companions | |||||
| Sedum forsterianum | V | - | - | - | 9 |
| Lavandula viridis | V | - | - | - | 8 |
| Dactylis hispanica subsp. lusitanica | V | - | - | - | 9 |
| Picris spinifera | IV | - | - | - | 7 |
| Thapsia villosa | IV | - | - | - | 7 |
| Rumex induratus | IV | - | - | - | 6 |
| Tamus communis | IV | - | - | - | 5 |
| Umbilicus rupestris | III | - | - | - | 6 |
| Arrhenatherum album var. erianthum | III | - | - | - | 4 |
| Lithodora lusitanica | III | - | - | - | 5 |
| Phagnalon saxatile | III | - | - | - | 5 |
| Cynara algarbiensis | II | - | - | - | 4 |
| Asphodelus aestivus | II | - | - | - | 6 |
| Genista triacanthos | II | - | - | - | 4 |
| Erophaca baetica | II | - | - | - | 3 |
| Lavandula luisieri | II | - | - | - | 3 |
| Digitalis purpurea | II | - | - | - | 3 |
| Cistus populifolius | II | - | - | - | 2 |
| Lavandula x alportelensis | II | - | - | - | 2 |
| Cistus x hybridus | II | - | - | - | 2 |
| Anthyllis vulneraria subsp. maura | II | - | - | - | 3 |
| Sanguisorba rupicola | II | - | - | - | 4 |
| Magydaris panacifolia | II | - | - | - | 3 |
| Celtica gigantea | II | - | - | - | 2 |
| Helichrysum stoechas | II | - | - | - | 2 |
| Erica australis | II | - | - | - | 3 |
| Geranium purpureum | II | II | 1 | - | 8 |
| Saxifraga granulata | II | I | - | - | 6 |
| Brachypodium phoenicoides | II | I | - | - | 4 |
| Cistus salviifolius | III | V | - | - | 16 |
| Cistus ladanifer | III | III | - | - | 11 |
| Pulicaria odora | I | III | - | - | 7 |
| Aristolochia paucinervis | I | II | - | - | 3 |
| Hymenocarpos lotoides | I | II | - | - | 3 |
| Campanula rapunculus | + | II | - | - | 4 |
| Cistus monspeliensis | - | V | - | - | 9 |
| Briza maxima | - | V | - | - | 9 |
| Agrostis castellana | - | V | - | - | 8 |
| Tuberaria guttata | - | V | - | - | 9 |
| Cistus crispus | - | V | - | - | 9 |
| Vulpia bromoides | - | V | - | - | 9 |
| Trifolium angustifolium | - | IV | - | - | 7 |
| Eryngium tenue | - | IV | - | - | 8 |
| Gastridium ventricosum | - | IV | - | - | 7 |
| Tolpis barbata | - | IV | - | - | 7 |
| Trifolium scabrum | - | IV | - | - | 7 |
| Cynosurus echinatus | - | IV | - | - | 8 |
| Brachypodium distachyon | - | IV | - | - | 7 |
| Rumex acetosella | - | IV | - | - | 8 |
| Trifolium glomeratum | - | IV | - | - | 7 |
| Bellis annua | - | IV | - | - | 8 |
| Lavandula stoechas | - | III | - | - | 6 |
| Agrostis pourretii | - | III | - | - | 6 |
| Calamintha nepeta | - | III | - | - | 4 |
| Cynara humilis | - | III | - | - | 6 |
| Parentucellia viscosa | - | III | - | - | 5 |
| Taeniatherum caput-medusae | - | III | - | - | 6 |
| Scilla autumnalis | - | III | - | - | 6 |
| Leucojum autumnale | - | III | - | - | 5 |
| Aira cupaniana | - | III | - | - | 5 |
| Aira caryophyllea | - | III | - | - | 5 |
| Molineriella minuta | - | III | - | - | 6 |
| Dactylis glomerata | - | III | - | - | 6 |
| Teucrium haenseleri | - | II | - | - | 4 |
| Cerastium brachypetalum | - | II | - | - | 4 |
| Silene rubella subsp. segetalis | - | II | - | - | 3 |
| Briza minor | - | II | - | - | 2 |
| Dittrichia viscosa | - | II | - | - | 3 |
| Erica scoparia | - | II | - | - | 3 |
| Thymus mastichina | - | II | - | - | 3 |
| Ulex australis subsp. australis | - | II | - | - | 4 |
| Cytisus scoparius subsp. scoparius | - | II | - | - | 4 |
| Margotia gummifera | - | II | - | - | 3 |
| Lathyrus latifolius | - | II | - | - | 3 |
| Coronilla dura | - | II | - | - | 4 |
| Carlina racemosa | - | II | - | - | 4 |
| Pulicaria paludosa | - | II | - | - | 4 |
| Gaudinia fragilis | - | II | - | - | 3 |
| Gladiolus reuteri | - | II | - | - | 4 |
| Crataegus monogyna | - | III | 2 | - | 7 |
| Retama sphaerocarpa | - | III | + | - | 10 |
| Cephalantera longifolia | - | II | - | II | 6 |
| Melica ciliata subsp. magnolii | - | II | - | I | 5 |
| Piptatherum miliaceum | - | - | 2 | - | 1 |
| Cistus albidus | - | - | 1 | - | 1 |
| Thymbra capitata | - | - | + | - | 1 |
| Rubus ulmifolius | - | - | + | - | 1 |
| Vinca difformis | - | - | + | - | 1 |
| Ptilostemon hispanicus | - | - | + | - | 1 |
| Urginea maritima | - | - | - | II | 2 |
| Cytisus striatus subsp. eriocarpus | - | - | - | II | 2 |
| Cytisus scoparius subsp. bourgaei | - | - | - | I | 1 |
| Ballota hirsuta | - | - | - | I | 1 |
| Rosa pouzinii | - | - | - | I | 1 |
| Rosa canina | - | - | - | I | 1 |
| Nepeta tuberosa | - | - | - | I | 1 |
| Holm Oak Association | Biogeographic Units or Territories | Bioclimatic Units (Thermotypes and Ombrotypes) | Substratum Affinity | Characteristics and Main Differentials |
|---|---|---|---|---|
| Ulici argentei-Quercetum rotundifoliae | Monchique Sierran District | Thermomediterranean sub-humid | Silicicolous (schists and greywackes substrates) | Ulex argenteus, Scilla monophyllos, Avenella stricta, Lavandula viridis, Cynara algarbiensis. |
| Myrto communis-Quercetum rotundifoliae | Mediterranean-Iberoatlantic | thermomediterranean dry | Silicicolous (schists and greywackes substrates) | Myrtus communis, Chamaerops humilis, Lavandula sampaioana |
| Rhamno oleoidis-Quercetum rotundifoliae | Bética Province, reaching the Algarve and Monchique Sector, Cádiz and Sado Subprovince | thermomediterranean, dry to subhumid | Calcicolous (limestone and calco-dolomitic substrates) | Ulex baeticus subsp. scaber, Thymbra capitata, Aristolochia baetica, Ptilostemon hispanicus |
| Rhamno laderoi-Quercetum rotundifoliae | Lusitania and Extremadura Subprovince | upper thermomediterranean or lower mesomediterranean, subhumid | Calcicolous (calcareous, calco-dolomitic and ultramafic or serpentine substrates) | Rhamnus lycioides subsp. laderoi, Teucrium fruticans |
| Relevé no. | 1 | 2 | 3 | 4 | 5 | 6 | 7 * | 8 | 9 | 10 | PRESENCES |
| Surface (m2) | 200 | 250 | 150 | 400 | 300 | 200 | 300 | 150 | 150 | 150 | |
| Altitude (m) | 170 | 135 | 155 | 165 | 260 | 165 | 160 | 390 | 375 | 385 | |
| Cover rate (%) | 90 | 80 | 70 | 85 | 95 | 80 | 100 | 90 | 95 | 70 | |
| Orientation | O | SE | SE | O | NE | NE | O | SE | NE | SE | |
| Slope (%) | 35 | 20 | 25 | 25 | 30 | 20 | 30 | 10 | 30 | 5 | |
| Altura média (m) | 6 | 6 | 5 | 5 | 8 | 6 | 8 | 4 | 7 | 3 | |
| No. of species | 30 | 27 | 32 | 39 | 19 | 38 | 35 | 35 | 24 | 15 | |
| Characteristic of association and higher units | |||||||||||
| Quercus rotundifolia | 5 | 4 | 3 | 4 | 5 | 4 | 5 | 4 | 4 | 3 | V |
| Erica arborea | 1 | 2 | 2 | 1 | + | + | 1 | + | 2 | + | V |
| Quercus coccifera | + | + | 1 | 1 | 2 | 1 | 1 | + | + | 1 | V |
| Ulex argenteus | 1 | + | 2 | 2 | - | 1 | + | 1 | + | 2 | V |
| Pistacia lentiscus | + | 1 | 2 | + | - | 1 | + | + | 1 | + | V |
| Arbutus unedo | 1 | + | 1 | 1 | 1 | - | 1 | 1 | - | 1 | IV |
| Phlomis purpurea | - | 1 | 1 | 1 | - | + | + | 1 | - | + | IV |
| Rhamnus alaternus | + | 1 | 1 | - | - | 1 | + | - | + | - | III |
| Smilax aspera var. altissima | 1 | 1 | 1 | - | 1 | + | 2 | - | - | - | III |
| Phillyrea angustifolia | 1 | - | - | + | - | - | 1 | 2 | - | 1 | III |
| Olea europaea var. sylvestris | - | + | + | + | - | + | - | + | - | - | III |
| Scilla monophyllos | 1 | 1 | + | - | + | - | 2 | - | - | - | III |
| Rubia peregrina | + | + | - | - | 1 | - | 1 | - | - | - | II |
| Lonicera implexa | - | - | + | + | - | - | + | + | - | - | II |
| Avenella stricta | - | - | - | + | + | - | + | + | - | - | II |
| Ruscus aculeatus | - | - | - | + | - | 1 | + | - | + | - | II |
| Hyacinthoides hispanica | - | - | - | - | 1 | + | - | - | + | - | II |
| Luzula forsteri subsp. baeticum | - | - | - | - | + | + | + | - | - | - | II |
| Asplenium onopteris | - | - | - | - | - | 2 | - | - | + | - | I |
| Daphne gnidium | - | - | + | - | - | - | - | + | - | - | I |
| Osyris lanceolata | - | - | - | - | - | - | - | + | - | + | I |
| Myrtus communis | - | - | - | + | - | - | - | - | + | - | I |
| Carex hallerana | - | - | - | + | - | + | - | - | - | - | I |
| Companions | |||||||||||
| Dactylis hispanica subsp. lusitanica | + | + | + | 1 | - | 1 | + | + | + | + | V |
| Sedum forsterianum | + | 1 | 1 | 1 | - | 1 | + | + | + | - | IV |
| Lavandula viridis | 1 | 1 | - | + | + | 1 | + | + | 2 | - | IV |
| Picris spinifera | - | 1 | 1 | 1 | + | + | + | + | - | - | IV |
| Cistus salviifolius | + | + | + | + | - | - | - | + | + | + | IV |
| Thapsia villosa | + | + | + | 1 | - | 1 | + | - | 2 | - | IV |
| Umbilicus rupestris | - | - | - | + | + | - | + | + | + | + | III |
| Phagnalon saxatile | + | + | + | + | - | - | - | - | + | - | III |
| Rumex induratus | + | + | + | + | - | + | + | - | - | - | III |
| Tamus communis | 1 | + | - | - | 2 | 2 | - | - | 1 | - | III |
| Lithodora lusitanica | - | + | + | + | - | - | + | + | - | - | III |
| Cistus ladanifer | + | - | + | 1 | - | + | - | - | - | + | III |
| Arrhenatherum album var. erianthum | - | - | + | + | - | + | - | + | - | - | II |
| Cynara algarbiensis | + | - | 1 | - | - | + | + | - | - | - | II |
| Saxifraga granulata | - | - | - | - | + | 1 | + | - | + | - | II |
| Genista triacanthos | - | + | - | 1 | - | + | - | + | - | - | II |
| Sanguisorba rupicola | + | + | - | + | - | - | + | - | - | - | II |
| Erica australis | + | - | - | 1 | - | - | - | + | - | - | II |
| Geranium purpureum | + | - | - | - | + | 1 | + | - | - | - | II |
| Brachypodium phoenicoides | + | - | + | - | - | + | - | - | - | - | II |
| Erophaca baetica | + | - | - | - | - | + | - | + | - | - | II |
| Lavandula luisieri | - | - | 1 | - | - | - | - | 1 | + | - | II |
| Digitalis purpurea | + | - | - | - | - | + | + | - | - | - | II |
| Anthyllis vulneraria subsp. maura | - | - | - | + | - | + | + | - | - | - | II |
| Magydaris panacifolia | - | + | - | + | - | - | + | - | - | - | II |
| Genista hirsuta | - | - | - | - | - | - | - | 1 | + | + | II |
| Cistus populifolius | + | - | - | - | - | - | + | - | - | - | I |
| Lavandula x alportelensis | - | - | - | 1 | - | - | - | 2 | - | - | I |
| Cistus x hybridus | - | + | + | - | - | - | - | - | - | - | I |
| Celtica gigantea | - | - | - | + | - | + | - | - | - | - | I |
| Helichrysum stoechas | - | - | - | + | - | - | - | + | - | - | I |
| Selaginella denticulata | - | - | - | - | + | - | - | - | + | - | I |
| Thymelaea villosa | - | - | - | - | - | - | - | + | - | + | I |
| Clinopodium vulgare subsp. arundanum | - | - | - | - | + | + | - | - | - | - | I |
| Pulicaria odora | + | - | - | + | - | - | - | - | - | - | I |
| Carlina corymbosa | - | + | - | - | - | - | - | - | - | - | I |
| Hyparrhenia sinaica | - | - | - | + | - | - | - | + | - | - | I |
References
- Martínez, S.R. Mapa de Series, Geoseries y Geopermaseries de Vegetación de España (Memoria Del Mapa de Vegetación Potencial de España). Parte 1. Itinera Geobot. 2007, 17, 5–436. [Google Scholar]
- Rivas-Martínez, S.; Penas, Á.; Díaz González, T.E.; Cantó, P.; del Río, S.; Costa, J.C.; Herrero, L.; Molero, J. Biogeographic Units of the Iberian Peninsula and Baelaric Islands to District Level. A Concise Synopsis. In The Vegetation of the Iberian Peninsula; Loidi, J., Ed.; Plant and Vegetation; Springer International Publishing: Cham, Switzerland, 2017; Volume 1, pp. 131–188. ISBN 978-3-319-54784-8. [Google Scholar]
- Loidi, J. Dynamism in Vegetation. Vegetation Changes on a Short Time Scale. In The Vegetation of the Iberian Peninsula; Springer: Cham, Switzerland, 2017; pp. 81–99. [Google Scholar] [CrossRef]
- Piñar Fuentes, J.C.; Cano-Ortiz, A.; Musarella, C.M.; Pinto-Gomes, C.; Spampinato, G.; Cano, E. Rupicolous habitats of interest for conservation in the central-southern Iberian Peninsula. Plant Sociol. 2017, 54, 29–42. [Google Scholar] [CrossRef]
- Piñar Fuentes, J.C.; Cano-Ortiz, A.; Musarella, C.M.; Quinto Canas, R.; Pinto Gomes, C.J.; Spampinato, G.; del Río, S.; Cano, E. Bioclimatology, Structure, and Conservation Perspectives of Quercus Pyrenaica, Acer opalus subsp. granatensis, and Corylus avellana Deciduous Forests on Mediterranean Bioclimate in the South-Central Part of the Iberian Peninsula. Sustainability 2019, 11, 6500. [Google Scholar] [CrossRef] [Green Version]
- Braun-Blanquet, J. Vegetationsskizzen Aus Dem Baskenland Mit Ausblicken Auf Das Weitere Ibero-Atlantikum II Teil. Vegetatio 1967, 14, 1–126. [Google Scholar] [CrossRef]
- Géhu, J.M.; Rivas-Martínez, S. Notions Fondadamentales de Phytosociologie. In Berichte der internationalen Symposien. Syntaxonomie IV-V Int. Vaduz; Dierschke, H., Ed.; Rinteln, Germany, 1981; pp. 5–33. [Google Scholar]
- Rivas-Martínez, S. Avances En Geobotánica. In Discurso de Apertura Del Curso Académico de La Real Academia Nacional de Farmacia Del Año 2005; Real Acad. Nacional de Farmacia: Madrid, Spain, 2005. [Google Scholar]
- Biondi, E. Phytosociology Today: Methodological and Conceptual Evolution. Plant Biosyst.-Int. J. Deal. All Asp. Plant Biol. 2011, 145, 19–29. [Google Scholar] [CrossRef]
- Braun-Blanquet, J. Fitosociología. Bases Para El Estudio de Las Comunidades Vegetales; H. Blume: Madrid, Spain, 1979. [Google Scholar]
- Rivas-Martínez, S.; Fernández-González, F.; Loidi, J.; Lousã, M.; Penas, Á. Syntaxonomical Checklist of Vascular Plant Communities of Spain and Portugal to Association Level. Itinera Geobot. 2001, 14, 5–341. [Google Scholar]
- Rivas-Martínez, S.; Díaz, T.E.; Fernández-González, F.; Izco, J.; Loidi, J.; Lousã, M.; Penas, Á. Vascular Plant Communities of Spain and Portugal: Addenda to the Syntaxonomical Checklist of 2001. Itinera Geobot. 2002, 15, 5–922. [Google Scholar]
- Rivas Martínez, S.; Penas, Á.; Díaz González, T.E.; Ladero, M.; Asensi, A.; Díez Garretas, B.; Molero Mesa, J.; Valle Tendero, F.; Cano, E.; Costa Talens, M.; et al. Mapa de series, geoseries y geopermaseries de vegetación de España (Memoria del mapa de vegetación potencial de España). Parte II. Itinera Geobot. 2011, 18, 5–800. [Google Scholar]
- Costa, J.C.; Neto, C.; Aguiar, C.; Capelo, J.; Espírito-Santo, M.D.; Honrado, J.; Pinto-Gomes, C.; Monteiro-Henriques, T.; Sequeira, M.; Lousã, M. Vascular Plant Communities in Portugal (Continental, Azores and Madeira). Glob. Geobot. 2012, 2, 1–180. [Google Scholar]
- Mucina, L.; Bültmann, H.; Dierßen, K.; Theurillat, J.-P.; Raus, T.; Čarni, A.; Šumberová, K.; Willner, W.; Dengler, J.; García, R.G.; et al. Vegetation of Europe: Hierarchical floristic classification system of vascular plant, bryophyte, lichen, and algal communities. Appl. Veg. Sci. 2016, 19, 3–264. [Google Scholar] [CrossRef]
- Coutinho, A. Flora de Portugal (Plantas Vasculares), 2nd ed.; Bertrand: Lisboa, Portugal, 1939. [Google Scholar]
- Franco, J. Nova Flora de Portugal (Continente e Açores); Escolar: Lisboa, Portugal, 1971; Volume I. [Google Scholar]
- Franco, J.; Rocha-Afonso, M. Nova Flora de Portugal (Continente e Açores); Escolar: Lisboa, Portugal, 1994; Volume III. [Google Scholar]
- Castroviejo, S. Flora Iberica; Real Jardín Botánico; CSIC: Madrid, Spain, 1986. [Google Scholar]
- Valdés, B.; Talavera, S.; Fernández-Galiano, E. Flora Vascular de Andalucía Occidental; Ketres: Barcelona, Spain, 1987. [Google Scholar]
- Sequeira, M.; Espírito-Santo, D.; Aguiar, C.; Capelo, C.; Honrado, J.J. (Eds.) Checklist da Flora de Portugal Continental, Açores e Madeira; ALFA: Lisboa, Portugal, 2011. [Google Scholar]
- Rivas-Martínez, S.; Penas, Á.; del Río, S.; Díaz González, T.E.; Rivas-Sáenz, S. Bioclimatology of the Iberian Peninsula and the Balearic Islands. In The Vegetation of the Iberian Peninsula; Loidi, J., Ed.; Plant and Vegetation; Springer International Publishing: Cham, Switzerland, 2017; Volume 1, pp. 29–80. ISBN 978-3-319-54784-8. [Google Scholar]
- Rivas Goday, S.; Borja, J.; Esteve, F.; Galiano, E.F.; Rigual, A.; Rivas-Martínez, S. Contribución al Estudio de La Quercetea Ilicis Hispánica. An. Inst. Bot. Cavanilles 1959, 285–406. [Google Scholar]
- Quinto-Canas, R. Flora y Vegetación de La Serra Do Caldeirão. Ph.D. Thesis, Facultad Ciencias Experimentales, Universidad de Jaén, Jaén, Spain, 2014. [Google Scholar]
- Theurillat, J.P.; Willner, W.; Fernández-González, F.; Bültmann, H.; Čarni, A.; Gigante, D.; Mucina, L.; Weber, H. International Code of Phytosociological Nomenclature. 4th edition. Appl. Veg. Sci. 2020, 1–62. [Google Scholar] [CrossRef] [Green Version]
- Clarke, K.R. Non-Parametric Multivariate Analyses of Changes in Community Structure. Aust. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Clarke, K.R.; Gorley, R.N. Primer v6 User Manual/Tutorial: Software for PRIMER-E (Version 6.1.5); PRIMER-E: Plymouth, UK, 2006. [Google Scholar]
- Van der Maarel, E. Transformation of Cover Abundance Values in Phytosociology and Its Effects on Community Similarity. Vegetation 1979, 97–114. [Google Scholar] [CrossRef]
- Quinto-Canas, R.; Mendes, P.; Meireles, C.; Musarella, C.; Pinto-Gomes, C. The Agrostion castellanae Rivas Goday 1957 Corr. Rivas Goday & Rivas- Martínez 1963 Alliance in the Southwestern Iberian Peninsula. Plant Sociol. 2018, 55, 21–29. [Google Scholar] [CrossRef]
- Paiva-Ferreira, R.; Pinto-Gomes, C. Flora e Vegetação do Barrocal Algarvio, Tavira-Portimão; Comissão de Coordenação e Desenvolvimento Regional do Algarve: Faro, Portugal, 2005. [Google Scholar]
- Molero, J.; Marfil, J.M. Betic and Southwest Andalusia. In The Vegetation of the Iberian Peninsula; Loidi, J., Ed.; Plant and Vegetation; Springer International Publishing: Cham, Switzerland, 2017; Volume 2, pp. 143–247. ISBN 978-3-319-54867-8. [Google Scholar]
- Quinto-Canas, R.; Vila-Viçosa, C.; Paiva-Ferreira, R.; Cano-Ortiz, A.; Pinto-Gomes, C. The Algarve Climatophilous Vegetation —Portugal: A Base Document to the Planning, Management and Nature Conservation. Acta Bot. Gall. 2012, 159, 289–298. [Google Scholar] [CrossRef]
- Aubard, V.; Paulo, J.A.; Silva, J.M.N. Long-Term Monitoring of Cork and Holm Oak Stands Productivity in Portugal with Landsat Imagery. Remote Sens. 2019, 11, 525. [Google Scholar] [CrossRef] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quinto Canas, R.; Cano-Ortiz, A.; Musarella, C.M.; del Río, S.; Raposo, M.; Fuentes, J.C.P.; Gomes, C.P. Quercus rotundifolia Lam. Woodlands of the Southwestern Iberian Peninsula. Land 2021, 10, 268. https://doi.org/10.3390/land10030268
Quinto Canas R, Cano-Ortiz A, Musarella CM, del Río S, Raposo M, Fuentes JCP, Gomes CP. Quercus rotundifolia Lam. Woodlands of the Southwestern Iberian Peninsula. Land. 2021; 10(3):268. https://doi.org/10.3390/land10030268
Chicago/Turabian StyleQuinto Canas, Ricardo, Ana Cano-Ortiz, Carmelo Maria Musarella, Sara del Río, Mauro Raposo, José Carlos Piñar Fuentes, and Carlos Pinto Gomes. 2021. "Quercus rotundifolia Lam. Woodlands of the Southwestern Iberian Peninsula" Land 10, no. 3: 268. https://doi.org/10.3390/land10030268
APA StyleQuinto Canas, R., Cano-Ortiz, A., Musarella, C. M., del Río, S., Raposo, M., Fuentes, J. C. P., & Gomes, C. P. (2021). Quercus rotundifolia Lam. Woodlands of the Southwestern Iberian Peninsula. Land, 10(3), 268. https://doi.org/10.3390/land10030268