
Light Transmissivity of Tree Shelters Interacts with Site Environment and Species Ecophysiology to Determine Outplanting Performance in Mediterranean Climates


Abstract
:1. Introduction
2. Materials and Methods
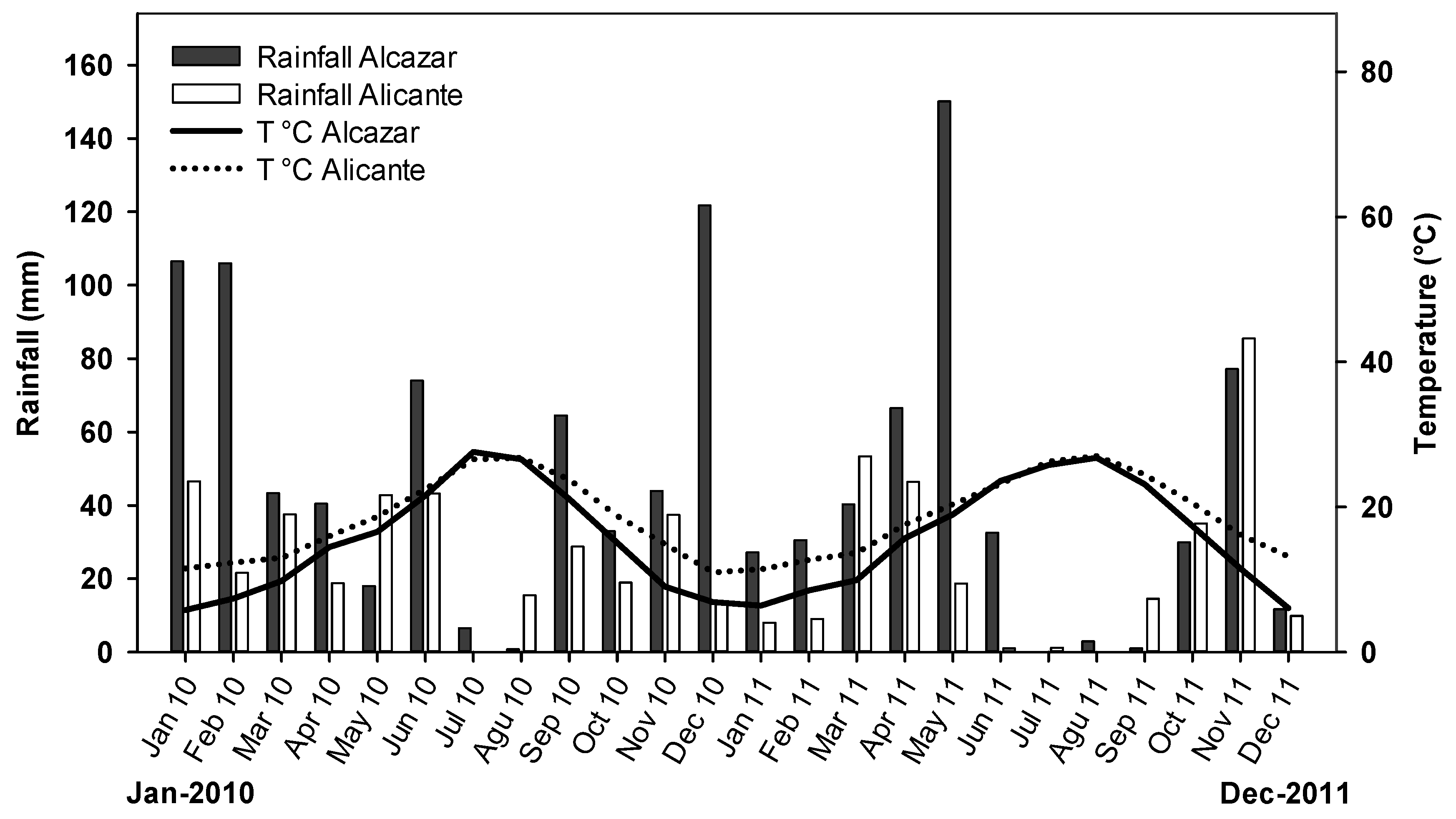
2.1. Study Sites, Plant Material and Experimental Layout
2.2. Outplanting Performance and Physiological Measurements
2.3. Statistical Analyses
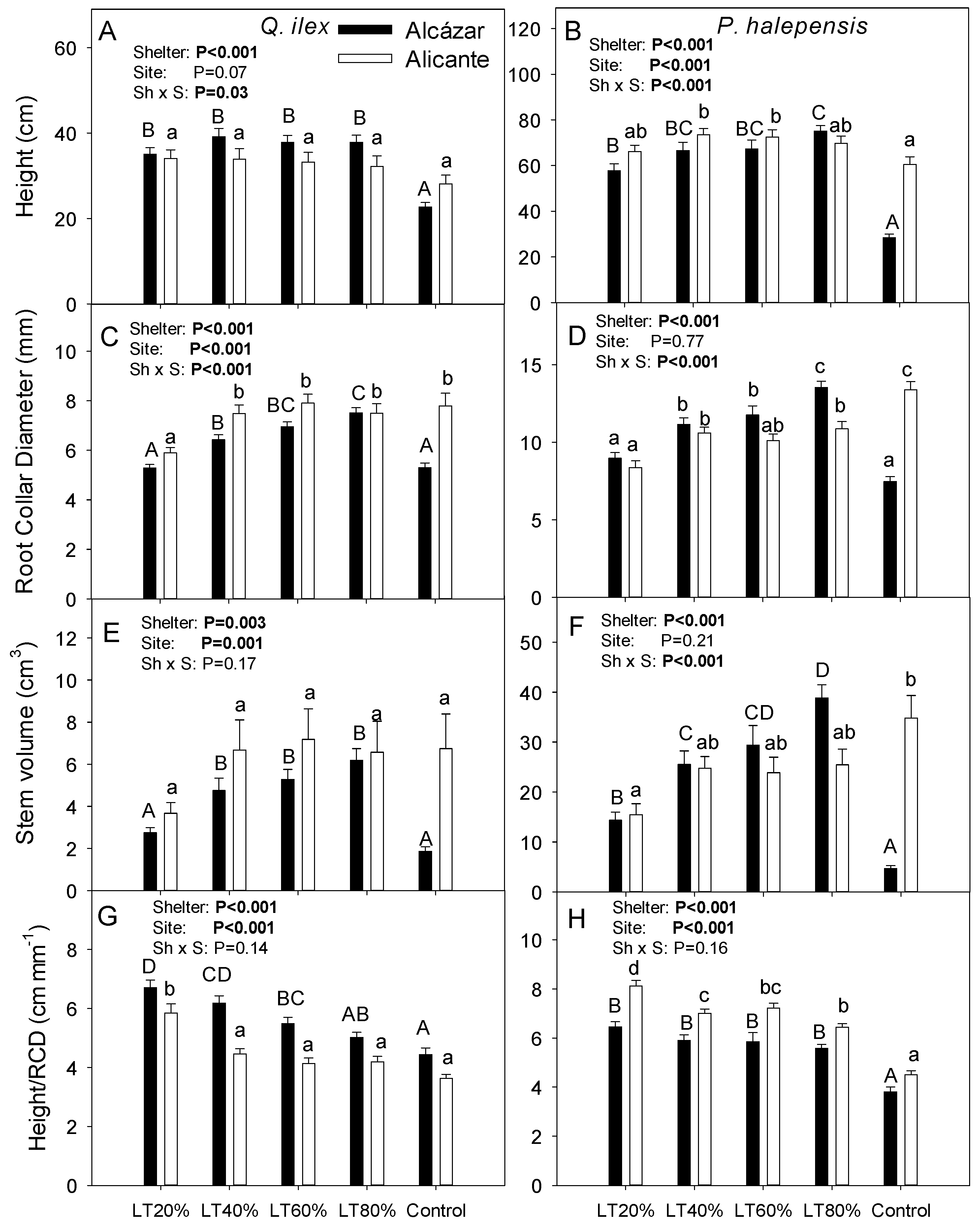
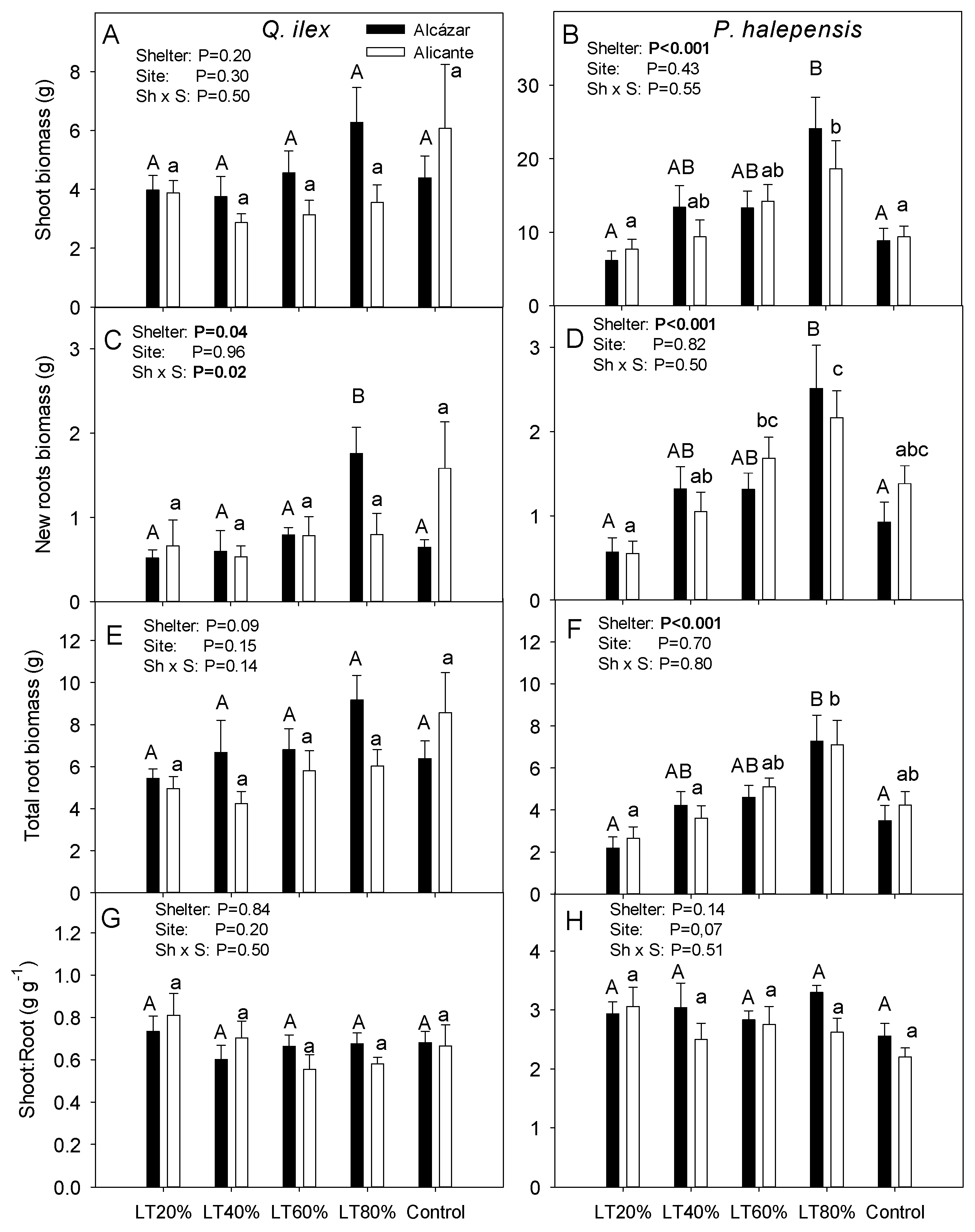
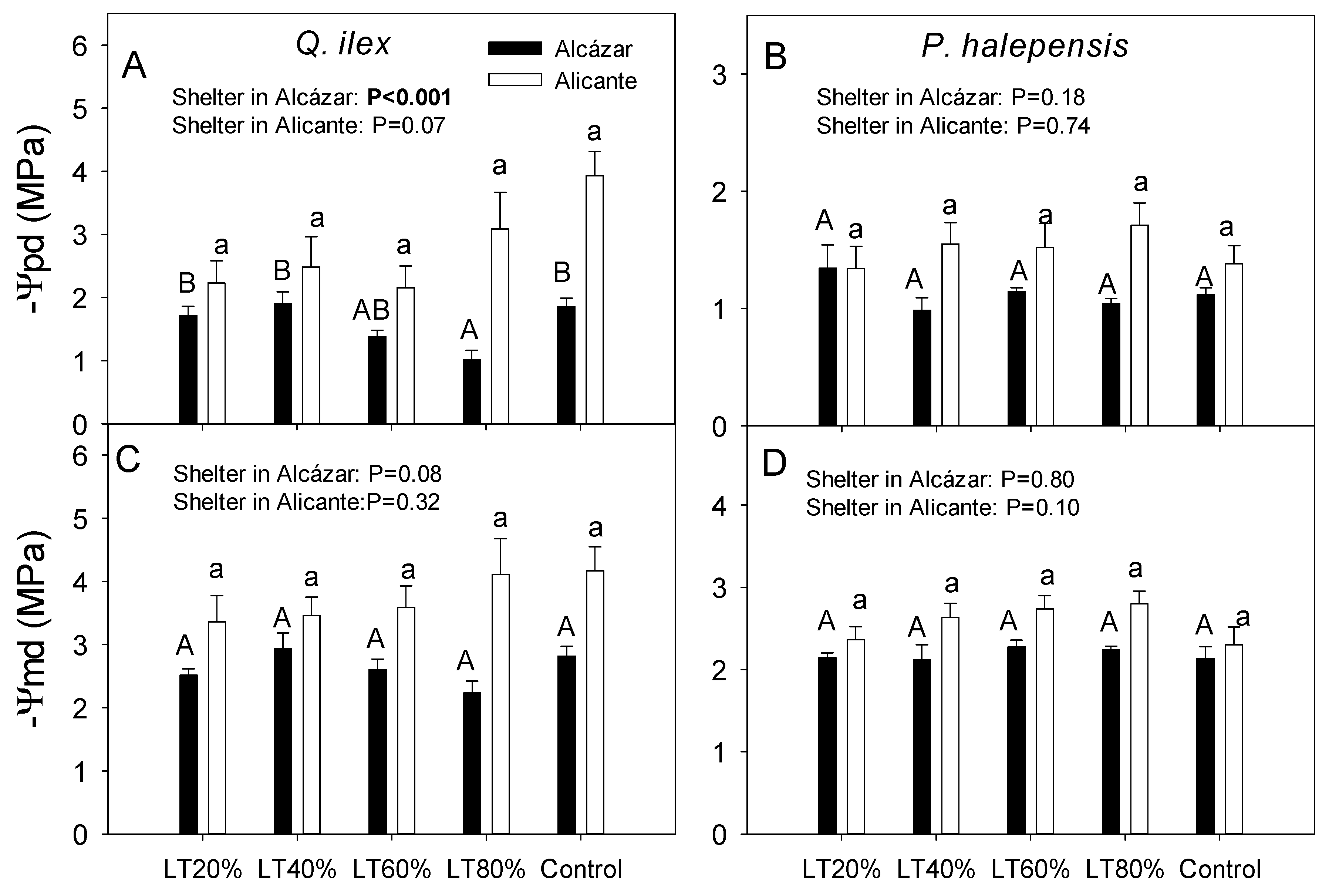
3. Results
4. Discussion
4.1. Effects of Light Transmissivity of Tree Shelters on Post-Planting Performance
4.2. Effect of Tree Shelters versus Non-Shelter Control
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Alcázar | Alicante | |
|---|---|---|
| pH | 8.6 ± 0.1 | 8.5 ± 0.2 |
| Conductivity (mS/cm) | 2.5 ± 0.2 | 0.5 ± 0.1 |
| Sand (g/100 g) | 34.5 ± 2.4 | 26.2 ± 1.0 |
| Silt (g/100 g) | 48.1 ± 1.9 | 36.3 ± 5.7 |
| Clay (q/100 g) | 17.5 ± 1.2 | 37.5 ± 6.4 |
| Oxidable organic matter (g/100 g) | 1.2 ± 0.1 | 1.7 ± 0.6 |
| Carbonates (CaCO3/100 g) | 26.6 ± 1.1 | 55.4 ± 5.7 |
| Active limestone (g/100 g) | 9.8 ± 0.2 | 13.9 ± 0.3 |
| Soluble phosphorus (mg/kg) | 9.83 ± 0.8 | 8.5 ± 2.7 |
| Soluble potassium (mg/kg) | 628.3 ± 54.9 | 396.3 ± 136.4 |
| Soluble calcium (meq/100 g) | 72.1 ± 20 | 42.7 ± 1.3 |
| Soluble magnesium (meq/100 g) | 2.8 ± 0.2 | 4.4 ± 0.4 |
| Soluble sodium (meq/100 g) | 0.06 ± 0.0 | 0.6 ± 0.0 |

References
- Cortina, J.; Amat, B.; Castillo, V.; Fuentes, D.; Maestre, F.T.; Padilla, F.M.; Rojo, L. The restoration of vegetation cover in the semi-arid Iberian southeast. J. Arid Environ. 2011, 75, 1377–1384. [Google Scholar] [CrossRef]
- Stanturf, J.A.; Mansourian, S. Forest landscape restoration. State Play R. Soc. Open Sci. 2020, 7, 201218. [Google Scholar] [CrossRef]
- Delgado-Baquerizo, M.; Maestre, F.T.; Gallardo, A.; Bowker, M.A.; Wallenstein, M.D.; Quero, J.L.; Ochoa, V.; Gozalo, B.; García-Gómez, M.; Soliveres, S.; et al. Decoupling of soil nutrient cycles as a function of aridity in global drylands. Nature 2013, 502, 672–676. [Google Scholar] [CrossRef] [PubMed]
- Calvete, C.; Estrada, R.; Angulo, E.; Cabezas-Ruiz, S. Habitat factors related to wild rabbit conservation in an agricultural landscape. Landsc. Ecol. 2004, 19, 531–542. [Google Scholar] [CrossRef]
- Castro, J.; Leverkus, A.B.; Fuster, F. A new device to foster oak forest restoration via seed sowing. New For. 2015, 46, 919–929. [Google Scholar] [CrossRef]
- Vallejo, R.V.; Smanis, A.; Chirino, E.; Fuentes, D.; Valdecantos, A.; Vilagrosa, A. Perspectives in dryland restoration: Approaches for climate change adaptation. New For. 2012, 43, 561–579. [Google Scholar] [CrossRef] [Green Version]
- Niinemets, U.; Keenan, T. Photosynthetic responses to stress in Mediterranean evergreens: Mechanisms and models. Environ. Exp. Bot. 2014, 103, 24–41. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2014: Impacts, Adaptation, and Vulnerability. In Part A: Global and Sectoral Aspects. Page Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2014. [Google Scholar]
- Valdecantos, A.; Fuentes, D.; Smanis, A.; Llovet, J.; Morcillo, L.; Bautista, S. Effectiveness of Low-Cost Planting Techniques for Improving Water Availability to Olea europaea Seedlings in Degraded Drylands. Rest. Ecol. 2014, 22, 327–335. [Google Scholar] [CrossRef]
- Acevedo, M.; Álvarez, C.; Cartes, E.; Dumroese, R.K.; González, M. Production and establishment techniques for the restoration of Nothofagus alessandrii, an endangered keystone species in a Mediterranean forest. New For. 2020, 51, 159–174. [Google Scholar] [CrossRef]
- Piñeiro, J.; Maestre, F.T.; Bartolomé, L.; Valdecantos, A. Ecotechnology as a tool for restoring degraded drylands: A meta-analysis of field experiments. Ecol. Eng. 2013, 61, 133–144. [Google Scholar] [CrossRef]
- Valenzuela, P.; Arellano, E.; Burger, J.; Pérez, M.A.; Oliet, J.A. Soil conditions and sheltering techniques improve active restoration of degraded Nothofagus pumilio forest in Southern Patagonia. For. Ecol. Manag. 2018, 424, 28–34. [Google Scholar] [CrossRef]
- Padilla, F.M.; Miranda, J.D.; Ortega, R.; Hervás, M.; Sánchez, J.; Pugnaire, F.I. Does shelter enhance early seedling survival in dry environments? A test with eight Mediterranean species. Appl. Veg. Sci. 2011, 14, 31–39. [Google Scholar] [CrossRef] [Green Version]
- Oliet, J.A.; Blasco, R.; Valenzuela, P.; Melero De Blas, M.; Puértolas, J. Should we use meshes or solid tube shelters when planting in Mediterranean semiarid environments? New For. 2019, 50, 267–282. [Google Scholar] [CrossRef]
- Oliet, J.; Navarro, R.; Contreras, O. Evaluación de la aplicación de mejoradores y tubos en repoblaciones forestales. In Manuales de Restauración Forestal; Consejería de Medio Ambiente de la Junta de Andalucía: Seville, Spain, 2003; 234p. [Google Scholar]
- Defaa, C.; Elantry, S.; El Alami, S.L.; Achour, A.; El Mousadik, A.; Msanda, F. Effects of Tree Shelters on the Survival and Growth of Argania spinosa Seedlings in Mediterranean Arid Environment. Int. J. Ecol. 2015, 2015, 124075. [Google Scholar] [CrossRef] [Green Version]
- Oliet, J.; Jacobs, D.F. Microclimatic conditions and plant morpho-physiological development within a tree shelter environment during establishment of Quercus ilex seedlings. Agric. For. Meteorol. 2007, 144, 58–72. [Google Scholar] [CrossRef]
- Bergez, J.E.; Dupraz, Z.C. Radiation and thermal microclimate in tree shelter. Agric. For. Meteorol. 2009, 149, 179–186. [Google Scholar] [CrossRef]
- Pemán, J.; Peguero-Pina, J.J.; Valladares, F.; Gil-Pelegrín, E. Evaluation of unventilated treeshelters in the context of Mediterranean climate: Insights from a study on Quercus faginea seedlings assessed with a 3D architectural plant model. Ecol. Eng. 2010, 36, 517–526. [Google Scholar] [CrossRef]
- Close, D.C.; Ruthrof, K.X.; Turner, S.; Rokich, D.P.; Dixon, K.W. Ecophysiology of Species with Distinct Leaf Morphologies: Effects of Plastic and Shadecloth Tree Guards. Rest. Ecol. 2009, 17, 33–41. [Google Scholar] [CrossRef]
- Puértolas, J.; Oliet, J.A.; Jacobs, D.F.; Benito, L.F.; Peñuelas, J.L. Is light the key factor for success of tube shelters in forest restoration plantings under Mediterranean climates? For. Ecol. Manag. 2010, 260, 610–617. [Google Scholar] [CrossRef]
- Jiménez, M.N.; Navarro, F.B.; Ripoll, M.A.; Bocio, I.; De Simón, E. Effect of shelter tubes on establishment and growth of Juniperus thurifera L. (Cupressaceae) seedlings in Mediterranean semi-arid environment. Ann. For. Sci. 2005, 62, 717–725. [Google Scholar] [CrossRef] [Green Version]
- Ballaré, C.; Pierik, R. The shade-avoidance syndrome: Multiple signals and ecological consequences. Plant Cell Environ. 2017, 40, 2530–2543. [Google Scholar] [CrossRef]
- Smith, T.; Huston, M. A theory of the spatial and temporal dynamics of plant communities. Plant Ecol. 1989, 83, 49–69. [Google Scholar] [CrossRef]
- Holmgren, M.; Gómez-Aparicio, L.; Quero, J.L.; Valladares, F. Non-linear effects of drought under shade: Reconciling physiological and ecological models in plant communities. Oecologia 2012, 169, 293–305. [Google Scholar] [CrossRef] [Green Version]
- Padilla, F.M.; Miranda, J.; Pugnaire, F.I. Early root growth plasticity in seedlings of three Mediterranean woody Species. Plant Soil 2007, 296, 103–113. [Google Scholar] [CrossRef]
- Perl-Treves, R.; Perl, A. Oxidative stress: An introduction. In Oxidative Stress in Plants; Inzé, D., van Montagu, M., Eds.; Taylor & Francis: London, UK, 2002; pp. 1–32. [Google Scholar]
- Oliet, J.A.; Vázquez De Castro, A.; Puértolas, J. Establishing Quercus ilex under Mediterranean dry conditions: Sowing recalcitrant acorns versus planting seedlings at different depths and tube shelter light transmissions. New For. 2015, 46, 869–883. [Google Scholar] [CrossRef] [Green Version]
- Vázquez De Castro, A.; Oliet, J.A.; Puértolas, J.; Jacobs, D.F. Light transmissivity of tube shelters affects root growth and biomass allocation of Quercus ilex L. and Pinus halepensis Mill. Ann. For. Sci. 2014, 71, 91–99. [Google Scholar] [CrossRef] [Green Version]
- Vadell, E.; De Miguel, S.; Pemán, J. Large-scale reforestation and afforestation policy in Spain: A historical review of its underlying ecological, socioeconomic and political dynamics. Land Use Policy 2016, 55, 37–48. [Google Scholar] [CrossRef]
- Pausas, J.G.; Blade, C.; Valdecantos, A.; Seva, J.P.; Fuentes, D.; Alloza, J.A.; Vilagrosa, A.; Bautista, S.; Cortina, J.; Vallejo, R. Pines and oaks in the restoration of Mediterranean landscapes of Spain: New perspectives for an old practice–A review. Plant Ecol. 2004, 171, 209–220. [Google Scholar] [CrossRef]
- Klein, T. The variability of stomatal sensitivity to leaf water potential across tree species indicates a continuum between isohydric and anisohydric behaviours. Func. Ecol. 2014, 28, 1313–1320. [Google Scholar] [CrossRef]
- Niinemets, U.; Valladares, F. Tolerance to Shade, Drought, and Waterlogging of Temperate Northern Hemisphere Trees and Shrubs. Ecol. Monogr. 2006, 76, 521–547. [Google Scholar] [CrossRef]
- Agencia Estatal De Meteorología, Instituto De Meteorología De Portugal. Atlas Climático Ibérico; Ministerio de Medio Ambiente, Medio Rural y Marino: Madrid, Spain, 2011; 80p.
- Agencia Estatal De Meteorología. Available online: http://agroclimap.aemet.es/ (accessed on 15 June 2021).
- Gómez-Miguel, V.D.; Badía-Villas, D. Soil Distribution and Classification. In The Soils of Spain; Gallardo, J.F., Ed.; World Soils Book Series; Springer: Berlin, Germany, 2016; pp. 11–48. [Google Scholar]
- Alía Miranda, R.; García Del Barrio, J.M.; Iglesias Sauce, S.; Mancha Núñez, J.A.; De Miguel y Del Ángel, J.; Nicolás Peragón, J.L.; Pérez Martín, F.; Sánchez De Ron, D. Regiones de Procedencia de Especies Forestales Españolas; Organismo Autónomo Parques Nacionales: Madrid, Spain, 2009; 363p. [Google Scholar]
- Puértolas, J.; Pardos, M.; Jiménez, M.D.; Aranda, I.; Pardos, J.A. Interactive responses of Quercus suber L. seedlings to light and mild water stress: Effects on morphology and gas exchange traits. Ann. For. Sci. 2008, 65, 611. [Google Scholar] [CrossRef] [Green Version]
- Korner, C. Paradigm shift in plant growth control. Curr. Opin. Plant Biol. 2015, 25, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Villar-Salvador, P.; Planelles, R.; Oliet, J.; Peñuelas-Rubira, J.; Jacobs, D.F.; González De Chavez, M. Drought tolerance and transplanting performance of holm oak (Quercus ilex L.) after drought hardening in the nursery. Tree Physiol. 2004, 24, 1147–1155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puértolas, J.; Benito, L.F.; Peñuelas, J.L. Effects of nursery shading on seedling quality and post-planting performance in two Mediterranean species with contrasting shade tolerance. New For. 2009, 38, 295–308. [Google Scholar] [CrossRef]
- Bellot, J.; Ortiz De Urbina, J.M.; Bonet, A.; Sánchez, J.R. The effects of treeshelters on the growth of Quercus coccifera L. seedlings in a semiarid environment. Forestry 2002, 75, 89–106. [Google Scholar] [CrossRef] [Green Version]
- Maseyk, K.S.; Lin, T.; Rotenberg, E.; Grünzweig, J.M.; Schwartz, A.; Yakir, D. Physiology-phenology interactions in a productive semi-arid pine forest. New Phytol. 2008, 178, 603–616. [Google Scholar] [CrossRef] [PubMed]
- Calamassi, R.; Paoletti, E.; Strati, S. Frost hardening and resistance in three Aleppo pine (Pinus halepensis Mill.) provenances. Isr. J. Plant Sci. 2001, 49, 179–186. [Google Scholar] [CrossRef]
- Climent, J.; e Silva, F.C.; Chambel, M.R.; Pardos, M.; Almeida, M.H. Freezing injury in primary and secondary needles of Mediterranean pine species of contrasting ecological niches. Ann. For. Sci. 2009, 66, 407. [Google Scholar] [CrossRef] [Green Version]
- Ne’eman, G.; Trabaud, L. (Eds.) Ecology, Biogeography and Management of Pinus halepensis and P. brutia Forest Ecosystems in the Mediterranean Basin; Backhuys Publishers: Kerkwerve, The Netherlands, 2000; 407p. [Google Scholar]
- Kjelgren, R.; Rupp, L.A. Establishment in treeshelters I: Shelters reduce growth, water use, and hardiness, but not drought avoidance. Hortscience 1997, 32, 1281–1283. [Google Scholar] [CrossRef] [Green Version]
- Coutand, C.; Dupraz, C.; Jaouen, G.; Ploquin, S.; Adam, B. Mechanical stimuli regulates the allocation of biomass in trees: Demonstration with young Prunus avium trees. Ann. Bot. 2008, 101, 1421–1432. [Google Scholar] [CrossRef]
- Javanmard, Z.; Tabari Kouchaksaraei, M.; Bahrami, H.A.; Hosseini, S.M.; Modarres Sanavi, S.A.M.; Struve, D.; Ammere, C. Soil dust effects on morphological, physiological and biochemical responses of four tree species of semiarid regions. Eur. J. For. Res 2020, 139, 333–348. [Google Scholar] [CrossRef]




| Treatment | Pinus halepensis | Quercus ilex | ||
|---|---|---|---|---|
| Alcázar | Alicante | Alcázar | Alicante | |
| First Year | ||||
| Control | 96.0 a | 100 a | 100 a | 85.7 a |
| LT80% | 98.0 a | 92.0 a | 100 a | 94.1 b |
| LT60% | 94.0 a | 100 a | 100 a | 94.0 b |
| LT40% | 94.0 a | 100 a | 100 a | 96.0 b |
| LT20% | 94.0 a | 98.0 a | 100 a | 96.0 b |
| Second Year | ||||
| Control | 81.8 a | 100 a | 100 a | 81.8 a |
| LT80% | 95.5 b | 91.1 a | 100 a | 82.6 a |
| LT60% | 85.0 b | 100 a | 100 a | 91.1 a |
| LT40% | 92.9 b | 100 a | 98.0 a | 86.7 a |
| LT20% | 92.5 b | 97.7 a | 98.0 a | 93.3 a |
| P. halepensis | Q. ilex | ||||
|---|---|---|---|---|---|
| d.f | Chi2 Test | p | Chi2 Test | p | |
| First Year | |||||
| Site | 1 | 3.7 | 0.05 | 18.5 | <0.001 |
| Shelter | 4 | 0.8 | 0.93 | 3.7 | 0.5 |
| Site × Shelter | 4 | 6.3 | 0.16 | 3.7 | 0.5 |
| Second Year | |||||
| Site | 1 | 12.0 | 0.001 | 26.6 | <0.001 |
| Shelter | 4 | 2.8 | 0.60 | 3.3 | 0.5 |
| Site × Shelter | 4 | 9.2 | 0.04 | 5.2 | 0.3 |
| Site | Pinus halepensis | Quercus ilex | ||||||
|---|---|---|---|---|---|---|---|---|
| Height (cm) | Root Collar Diameter (mm) | Height (cm) | Root Collar Diameter (mm) | |||||
| Alcázar | Alicante | Alcázar | Alicante | Alcázar | Alicante | Alcázar | Alicante | |
| First Year | ||||||||
| Control | 14.3 ± 0.9 a | 9.5 ± 0.6 a | 3.0 ± 0.1 a | 3.1 ± 0.2 c | 7.7 ± 0.6 a | 4.0 ± 0.6 a | 1.1 ± 0.0 a | 0.8 ± 0.1 b |
| LT80% | 25.2 ± 1.4 b | 15.5 ± 1.2 b | 4.2 ± 0.2 b | 2.8 ± 0.2 bc | 14.9 ± 1.0 b | 5.9 ± 0.8 ab | 1.9 ± 0.1 b | 1.0 ± 0.1 c |
| LT60% | 25.1 ± 1.3 b | 17.0 ± 1.0 bc | 4.2 ± 0.2 b | 2.7 ± 0.1 bc | 13.5 ± 0.9 b | 7.0 ± 1.0 b | 1.7 ± 0.1 b | 1.0 ± 0.1 c |
| LT40% | 24.9 ± 1.3 b | 20.7 ± 1.2 c | 3.3 ± 0.2 a | 2.6 ± 0.1 b | 13.7 ± 0.9 b | 8.0 ± 1.2 b | 1.4 ± 0.1 ab | 0.9 ± 0.1 c |
| LT20% | 22.8 ± 1.3 b | 19.8 ± 1.1 c | 2.4 ± 0.1 a | 1.8 ± 0.1 a | 13.3 ± 0.9 b | 8.7 ± 1.0 b | 1.0 ± 0.0 a | 0.5 ± 0.0 a |
| Second Year | ||||||||
| Control | 4.0 ± 1.0 a | 37.9 ± 3.0 ab | 1.8 ± 0.2 a | 7.7 ± 0.4 c | 2.8 ± 0.6 a | 8.9 ± 1.5 a | 0.6 ± 0.1 a | 2.4 ± 0.4 b |
| LT80% | 37.8 ± 2.3 c | 41.0 ± 2.5 ab | 6.9 ± 0.3 d | 5.5 ± 0.4 b | 10.9 ± 0.9 b | 11.4 ± 1.7 a | 1.9 ± 0.1 b | 2.1 ± 0.3 b |
| LT60% | 30.7 ± 1.4 bc | 43.4 ± 2.5 b | 5.0 ± 0.5 cd | 5.0 ± 0.3 b | 11.3 ± 1.0 b | 11.5 ± 1.6 a | 1.5 ± 0.1 b | 2.4 ± 0.3 b |
| LT40% | 28.7 ± 2.9 bc | 39.5 ± 2.2 ab | 5.2 ± 0.4 c | 5.4 ± 0.3 b | 12.8 ± 1.2 b | 9.8 ± 1.4 a | 1.4 ± 0.1 b | 2.0 ± 0.3 b |
| LT20% | 19.2 ± 2.3 b | 33.7 ± 1.8 a | 3.9 ± 0.3 b | 4.1 ± 0.3 a | 9.5 ± 0.7 b | 8.9 ± 1.6 a | 0.7 ± 0.1 a | 1.1 ± 0.2 a |
| Pinus halepensis | Quercus ilex | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| HG | RCDG | HG | RCDG | ||||||
| d.f | F | p | F | p | F | p | F | p | |
| First Year | |||||||||
| Site | 1 | 72.9 | <0.001 | 59.2 | <0.001 | 92.2 | <0.001 | 72.8 | <0.001 |
| Shelter | 4 | 28.8 | <0.001 | 18.8 | <0.001 | 10.3 | <0.001 | 13.9 | <0.001 |
| Site × Shelter | 4 | 2.0 | 0.09 | 6.6 | <0.001 | 2.3 | 0.05 | 1.4 | 0.2 |
| Second Year | |||||||||
| Site | 1 | 73.5 | <0.001 | 12.8 | <0.001 | 0.02 | 0.88 | 22.9 | <0.001 |
| Shelter | 4 | 14.7 | <0.001 | 10.8 | <0.001 | 22.7 | <0.001 | 11.9 | <0.001 |
| Site × Shelter | 4 | 9.4 | <0.001 | 27.9 | <0.001 | 11.0 | <0.001 | 4.5 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oliet, J.A.; Puértolas, J.; Valenzuela, P.; Vázquez de Castro, A. Light Transmissivity of Tree Shelters Interacts with Site Environment and Species Ecophysiology to Determine Outplanting Performance in Mediterranean Climates. Land 2021, 10, 753. https://doi.org/10.3390/land10070753
Oliet JA, Puértolas J, Valenzuela P, Vázquez de Castro A. Light Transmissivity of Tree Shelters Interacts with Site Environment and Species Ecophysiology to Determine Outplanting Performance in Mediterranean Climates. Land. 2021; 10(7):753. https://doi.org/10.3390/land10070753
Chicago/Turabian StyleOliet, Juan A., Jaime Puértolas, Patricio Valenzuela, and Alberto Vázquez de Castro. 2021. "Light Transmissivity of Tree Shelters Interacts with Site Environment and Species Ecophysiology to Determine Outplanting Performance in Mediterranean Climates" Land 10, no. 7: 753. https://doi.org/10.3390/land10070753
APA StyleOliet, J. A., Puértolas, J., Valenzuela, P., & Vázquez de Castro, A. (2021). Light Transmissivity of Tree Shelters Interacts with Site Environment and Species Ecophysiology to Determine Outplanting Performance in Mediterranean Climates. Land, 10(7), 753. https://doi.org/10.3390/land10070753