
Microbial Biomass Is More Important than Runoff Export in Predicting Soil Inorganic Nitrogen Concentrations Following Forest Conversion in Subtropical China

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Experimental Design
2.3. Soil and Fine Root Sampling, and Fine Root Biomass Estimation
2.4. Soil Physical and Chemical Property Analysis
2.5. Soil Microbial Properties Assay
2.6. Potential Soil Enzyme Activity Measurement
2.7. NH4+ and NO3− Export via Runoff
2.8. Data Analysis
3. Results
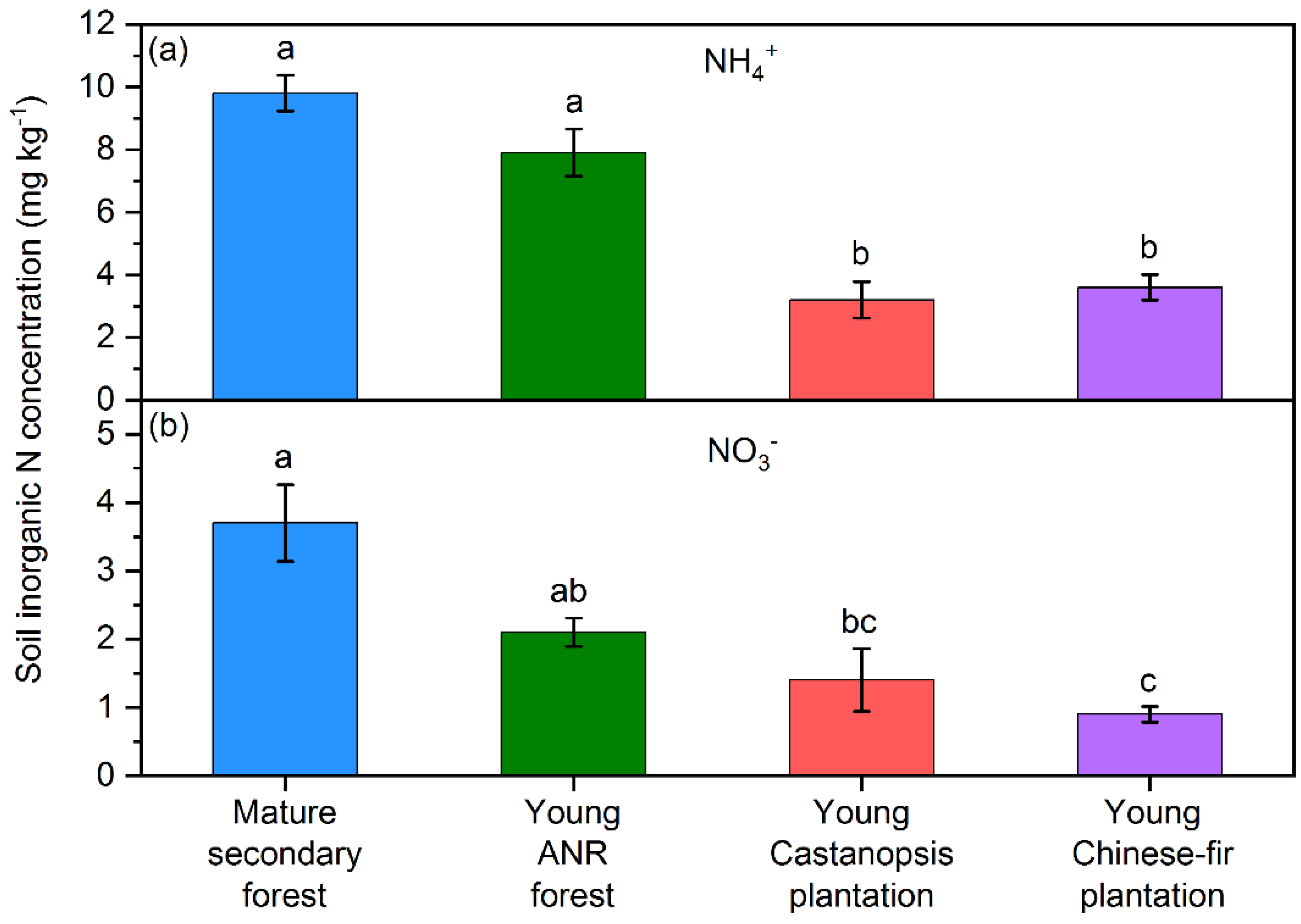
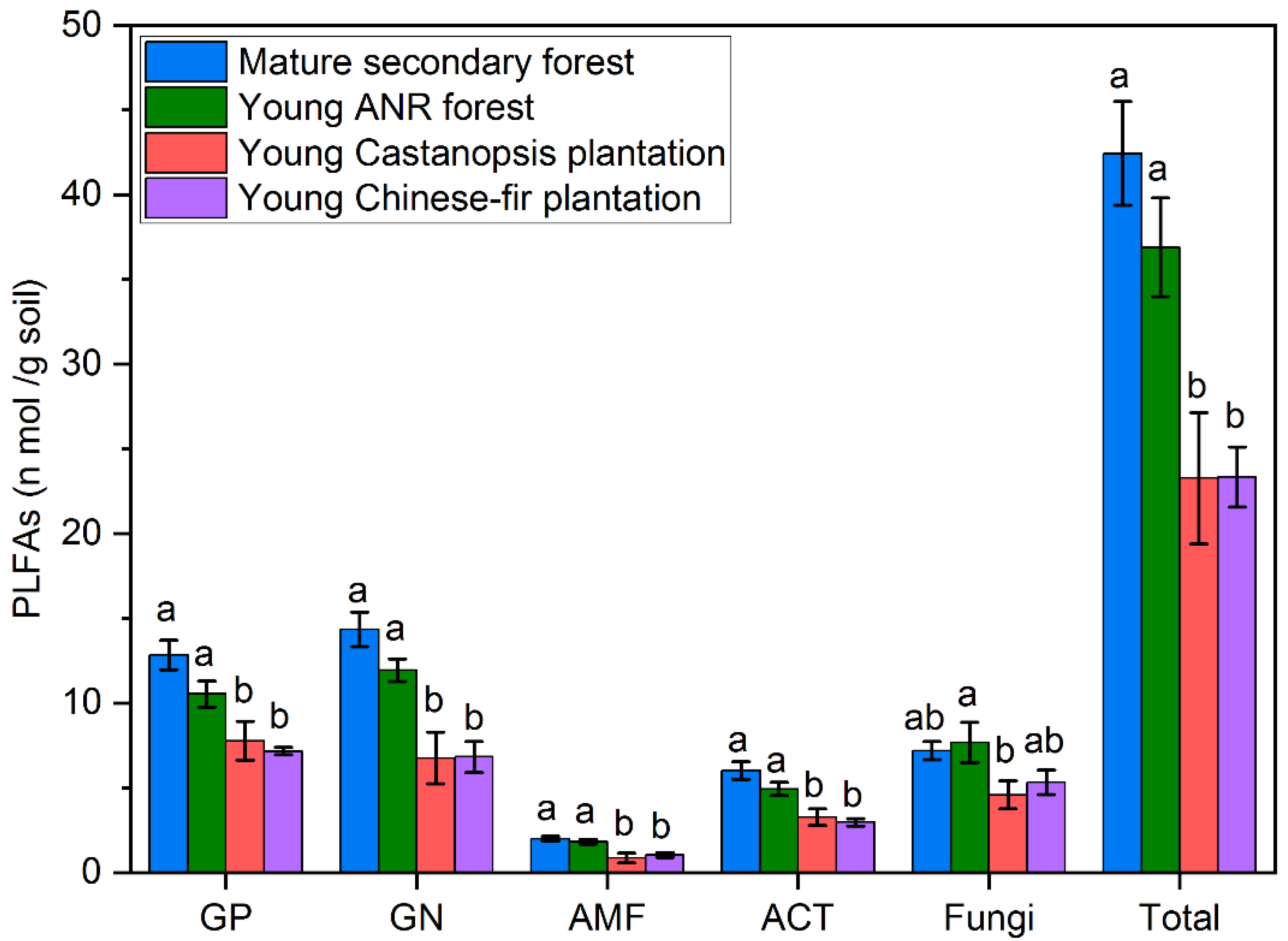
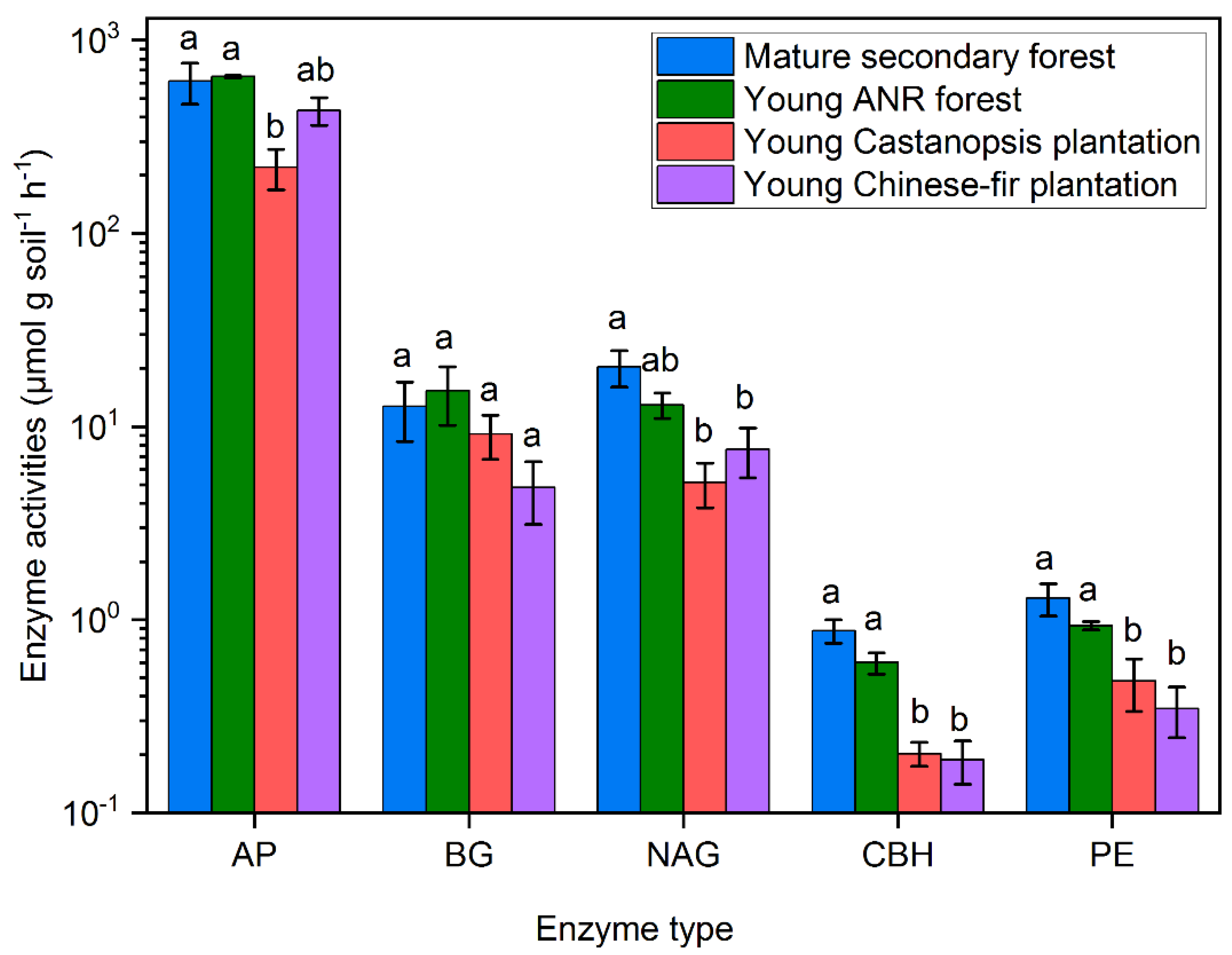
3.1. Soil Inorganic N Concentrations, Microbial Properties, and Physicochemical Properties
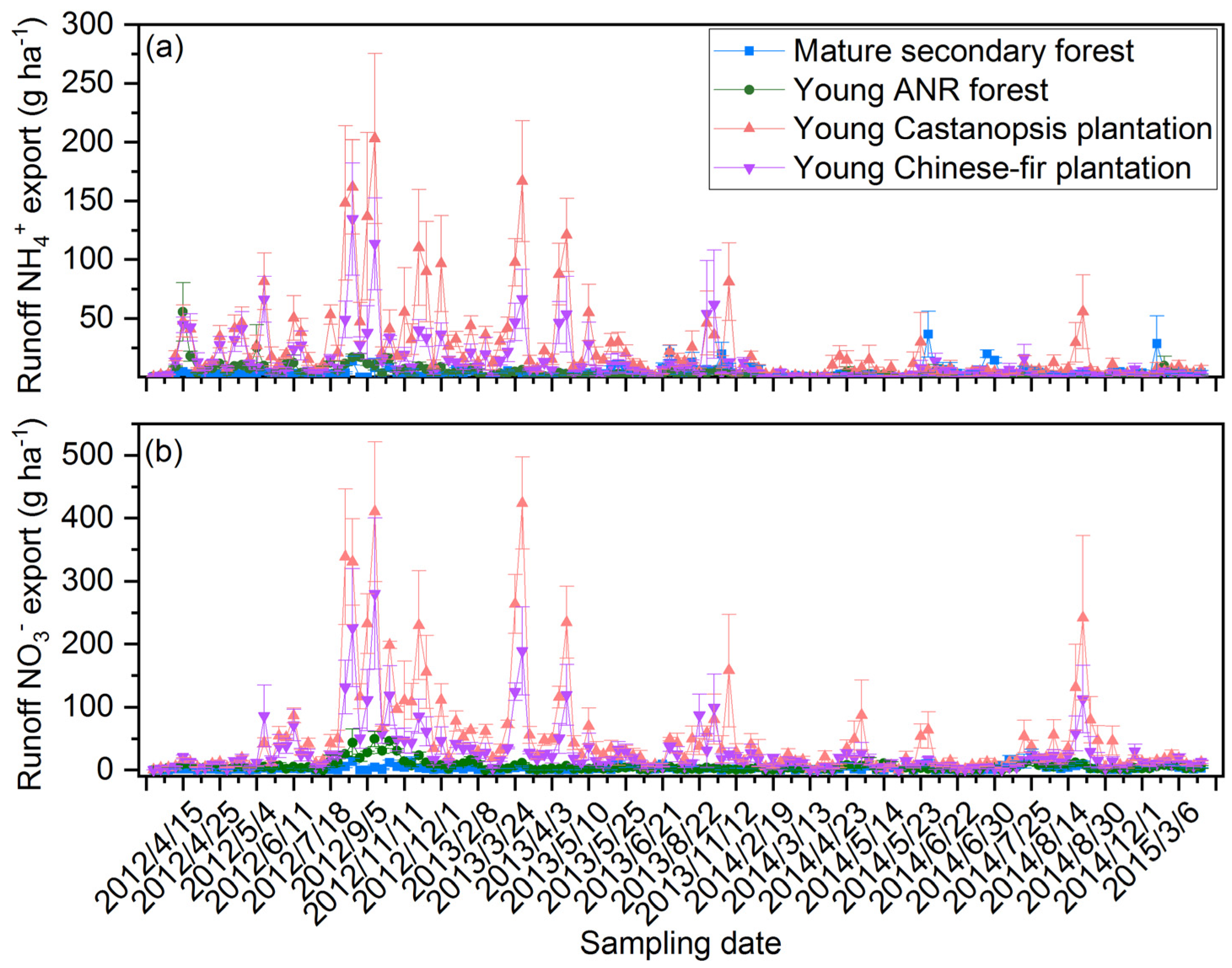
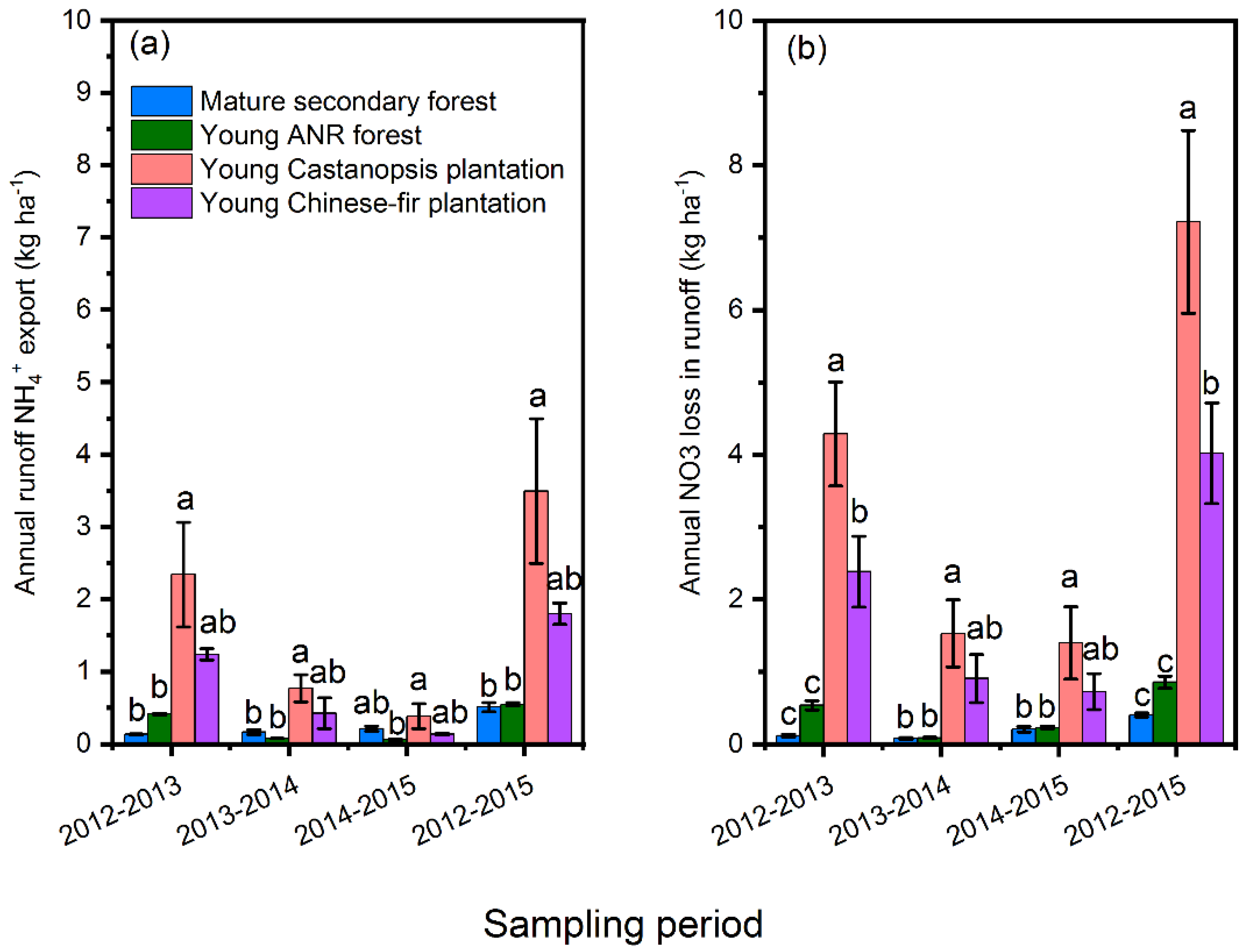
3.2. Surface Runoff NH4+ and NO3− Export
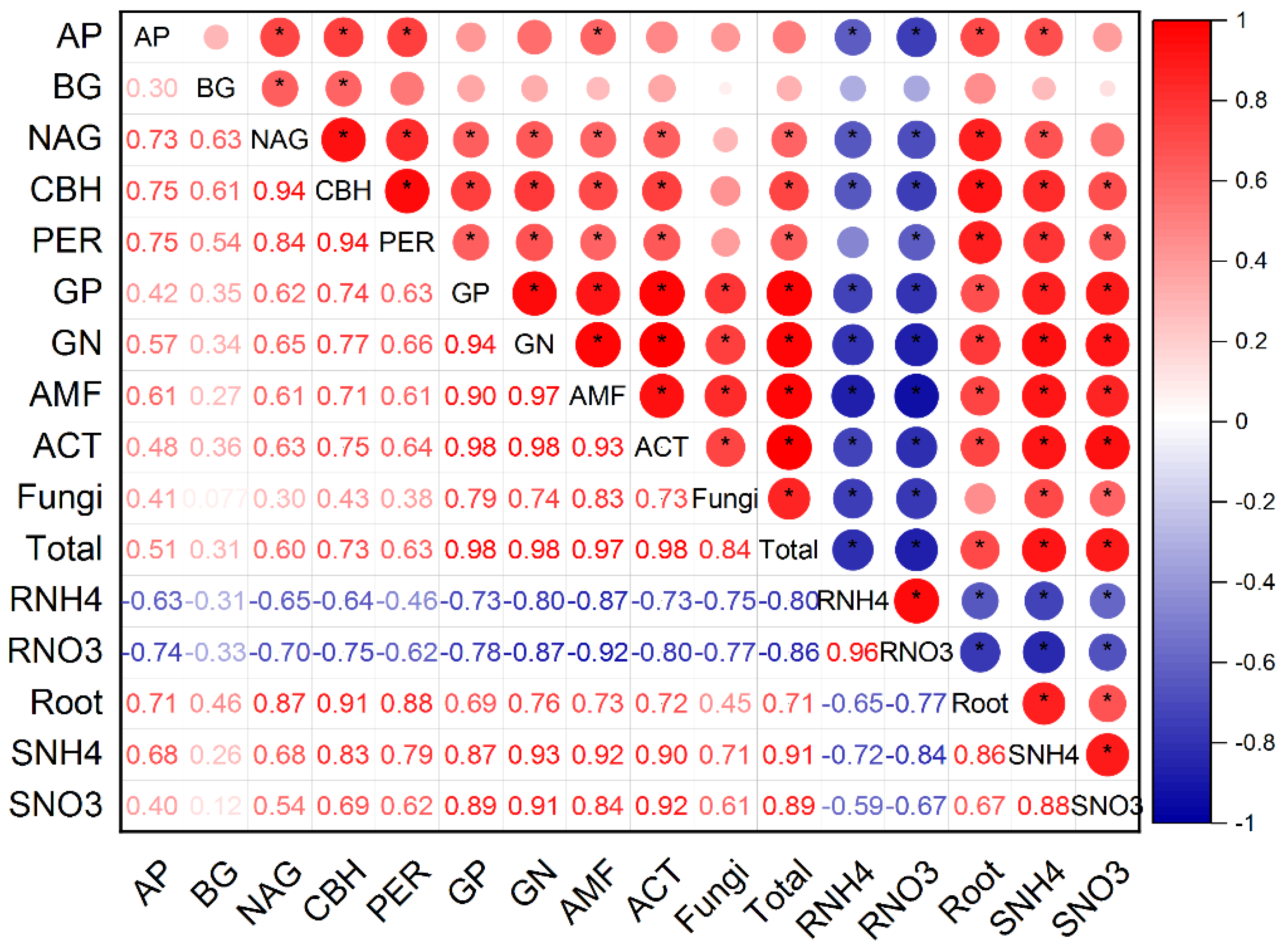
3.3. Soil Inorganic N Concentration in Relation to Soil Microbial Attributes and Surface Runoff
4. Discussion
4.1. Soil Inorganic N in Relation to Soil Microbial Attributes and Surface Runoff Inorganic N Export
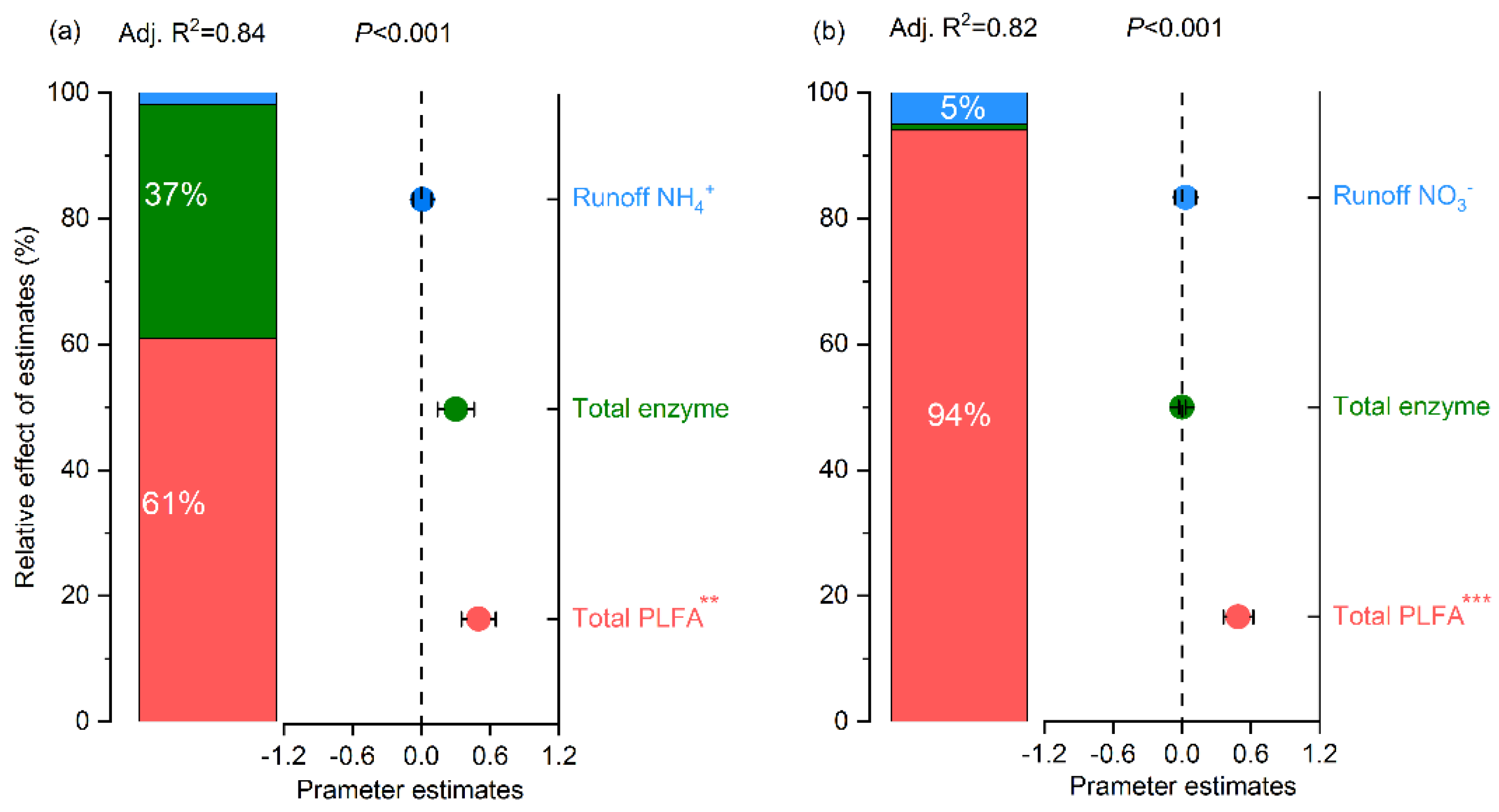
4.2. The Dominant Factors Influencing Post-Conversion Soil Inorganic N Concentrations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. Global Forest Resources Assessment 2020; 0378-1127; FAO: Rome, Italy, 2020; pp. 3–8. [Google Scholar]
- State Forestry Administration of China. China Forest Resource Report (2009–2013); State Forestry Administration of China: Beijing, China, 2014.
- Thomson, K. The State of Food and Agriculture 2008: Biofuels: Prospects, Risks and Opportunities. J. Agric. Sci. 2009, 147, 503–504. [Google Scholar] [CrossRef]
- Xu, J.; Xie, S.; Han, A.; Rao, R.; Huang, G.; Chen, X.; Zhang, L. Forest Resources in China—The 9th National Forest Inventory; National Forestry and Grassland Administration: Beijing, China, 2019.
- Balthazar, V.; Vanacker, V.; Molina, A.; Lambin, E.F. Impacts of forest cover change on ecosystem services in high Andean mountains. Ecol. Ind. 2015, 48, 63–75. [Google Scholar] [CrossRef]
- Ditt, E.H.; Mourato, S.; Ghazoul, J.; Knight, J. Forest conversion and provision of ecosystem services in the Brazilian Atlantic Forest. Land Degrad. Dev. 2010, 21, 591–603. [Google Scholar] [CrossRef]
- Walker, W.S.; Gorelik, S.R.; Baccini, A.; Aragon-Osejo, J.L.; Josse, C.; Meyer, C.; Macedo, M.N.; Augusto, C.; Rios, S.; Katan, T.; et al. The role of forest conversion, degradation, and disturbance in the carbon dynamics of Amazon indigenous territories and protected areas. Proc. Natl. Acad. Sci. USA 2020, 117, 3015–3025. [Google Scholar] [CrossRef] [Green Version]
- Guillaume, T.; Kotowska, M.M.; Hertel, D.; Knohl, A.; Krashevska, V.; Murtilaksono, K.; Scheu, S.; Kuzyakov, Y. Carbon costs and benefits of Indonesian rainforest conversion to plantations. Nat. Commun. 2018, 9, 238. [Google Scholar] [CrossRef]
- Van Straaten, O.; Corre, M.D.; Wolf, K.; Tchienkoua, M.; Cuellar, E.; Matthews, R.B.; Veldkamp, E. Conversion of lowland tropical forests to tree cash crop plantations loses up to one-half of stored soil organic carbon. Proc. Natl. Acad. Sci. USA 2015, 112, 9956–9960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Wang, S.; Yu, X. Decline of soil fertility during forest conversion of secondary forest to Chinese fir plantations in subtropical China. Land Degrad. Dev. 2011, 22, 444–452. [Google Scholar] [CrossRef]
- Corre, M.D.; Dechert, G.; Veldkamp, E. Soil Nitrogen Cycling following Montane Forest Conversion in Central Sulawesi, Indonesia. Soil Sci. Soc. Am. J. 2006, 70, 359–366. [Google Scholar] [CrossRef] [Green Version]
- Yan, E.-R.; Wang, X.-H.; Huang, J.-J.; Li, G.-Y.; Zhou, W. Decline of soil nitrogen mineralization and nitrification during forest conversion of evergreen broad-leaved forest to plantations in the subtropical area of Eastern China. Biogeochemistry 2008, 89, 239–251. [Google Scholar] [CrossRef]
- Yang, L.; Chen, S.; Li, Y.; Wang, Q.; Zhong, X.; Yang, Z.; Lin, C.; Yang, Y. Conversion of Natural Evergreen Broadleaved Forests Decreases Soil Organic Carbon but Increases the Relative Contribution of Microbial Residue in Subtropical China. Forests 2019, 10, 468. [Google Scholar] [CrossRef] [Green Version]
- Vitousek, P.M.; Howarth, R.W. Nitrogen limitation on land and in the sea: How can it occur? Biogeochemistry 1991, 13, 87–115. [Google Scholar] [CrossRef]
- Pellegrini, A.F.A.; Ahlström, A.; Hobbie, S.E.; Reich, P.B.; Nieradzik, L.P.; Staver, A.C.; Scharenbroch, B.C.; Jumpponen, A.; Anderegg, W.R.L.; Randerson, J.T.; et al. Fire frequency drives decadal changes in soil carbon and nitrogen and ecosystem productivity. Nature 2018, 553. [Google Scholar] [CrossRef] [PubMed]
- Norby, R.J.; Warren, J.M.; Iversen, C.M.; Medlyn, B.E.; McMurtrie, R.E. CO2 enhancement of forest productivity constrained by limited nitrogen availability. Proc. Natl. Acad. Sci. USA 2010, 107, 19368–19373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reich, P.B.; Hobbie, S.E.; Lee, T.; Ellsworth, D.S.; West, J.B.; Tilman, D.; Knops, J.M.H.; Naeem, S.; Trost, J. Nitrogen limitation constrains sustainability of ecosystem response to CO2. Nature 2006, 440, 922–925. [Google Scholar] [CrossRef] [PubMed]
- Ngoze, S.; Riha, S.; Lehmann, J.; Verchot, L.; Kinyangi, J.; Mbugua, D.; Pell, A. Nutrient constraints to tropical agroecosystem productivity in long-term degrading soils. Glob. Change Biol. 2008, 14, 2810–2822. [Google Scholar] [CrossRef]
- Pandey, C.B.; Singh, G.B.; Singh, S.K.; Singh, R.K. Soil nitrogen and microbial biomass carbon dynamics in native forests and derived agricultural land uses in a humid tropical climate of India. Plant Soil 2010, 333, 453–467. [Google Scholar] [CrossRef]
- Reiners, W.A.; Bouwman, A.F.; Parsons, W.F.J.; Keller, M. Tropical Rain Forest Conversion to Pasture: Changes in Vegetation and Soil Properties. Ecol. Appl. 1994, 4, 363–377. [Google Scholar] [CrossRef]
- Greaver, T.L.; Clark, C.M.; Compton, J.E.; Vallano, D.; Talhelm, A.F.; Weaver, C.P.; Band, L.E.; Baron, J.S.; Davidson, E.A.; Tague, C.L.; et al. Key ecological responses to nitrogen are altered by climate change. Nat. Clim. Change 2016, 6, 836–843. [Google Scholar] [CrossRef]
- Byers, A.-K.; Condron, L.; Donavan, T.; O’Callaghan, M.; Patuawa, T.; Waipara, N.; Black, A. Soil microbial diversity in adjacent forest systems—Contrasting native, old growth kauri (Agathis australis) forest with exotic pine (Pinus radiata) plantation forest. FEMS Microbiol. Ecol. 2020, 96. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Zou, Y.; Di Gioia, D.; Singh, B.K.; Li, Q. Conversion to agroforestry and monoculture plantation is detrimental to the soil carbon and nitrogen cycles and microbial communities of a rainforest. Soil Biol. Biochem. 2020, 147, 107849. [Google Scholar] [CrossRef]
- Wood, S.A.; Gilbert, J.A.; Leff, J.W.; Fierer, N.; D’Angelo, H.; Bateman, C.; Gedallovich, S.M.; Gillikin, C.M.; Gradoville, M.R.; Mansor, P.; et al. Consequences of tropical forest conversion to oil palm on soil bacterial community and network structure. Soil Biol. Biochem. 2017, 112, 258–268. [Google Scholar] [CrossRef]
- You, Y.; Xiang, W.; Ouyang, S.; Zhao, Z.; Chen, L.; Zeng, Y.; Lei, P.; Deng, X.; Wang, J.; Wang, K. Hydrological fluxes of dissolved organic carbon and total dissolved nitrogen in subtropical forests at three restoration stages in southern China. J. Hydrol. 2020, 583, 124656. [Google Scholar] [CrossRef]
- Zhang, Q.; Shamsi, I.H.; Wang, J.; Song, Q.; Xue, Q.; Yu, Y.; Lin, X.; Hussain, S. Surface runoff and nitrogen (N) loss in a bamboo (Phyllostachys pubescens) forest under different fertilization regimes. Environ. Sci. Pollut. Res. 2013, 20, 4681–4688. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Chen, F.; Ouyang, Z.; Tu, N.; Xu, W.; Wang, X.; Miao, H.; Li, X.; Tian, Y. Impacts of reforestation approaches on runoff control in the hilly red soil region of Southern China. J. Hydrol. 2008, 356, 174–184. [Google Scholar] [CrossRef]
- Boyrahmadi, M.; Raiesi, F. Plant roots and species moderate the salinity effect on microbial respiration, biomass, and enzyme activities in a sandy clay soil. Biol. Fertil. Soils 2018, 54, 509–521. [Google Scholar] [CrossRef]
- Wang, L.; Pang, X.; Li, N.; Qi, K.; Huang, J.; Yin, C. Effects of vegetation type, fine and coarse roots on soil microbial communities and enzyme activities in eastern Tibetan plateau. Catena 2020, 194, 104694. [Google Scholar] [CrossRef]
- Ren, C.; Chen, J.; Deng, J.; Zhao, F.; Han, X.; Yang, G.; Tong, X.; Feng, Y.; Shelton, S.; Ren, G. Response of microbial diversity to C:N:P stoichiometry in fine root and microbial biomass following afforestation. Biol. Fertil. Soils 2017, 53, 457–468. [Google Scholar] [CrossRef]
- Tripathi, N.; Singh, R.S. Influence of different land uses on soil nitrogen transformations after conversion from an Indian dry tropical forest. Catena 2009, 77, 216–223. [Google Scholar] [CrossRef]
- Guo, J.; Yang, Z.; Lin, C.; Liu, X.; Chen, G.; Yang, Y. Conversion of a natural evergreen broadleaved forest into coniferous plantations in a subtropical area: Effects on composition of soil microbial communities and soil respiration. Biol. Fertil. Soils 2016, 52, 799–809. [Google Scholar] [CrossRef]
- Henneron, L.; Kardol, P.; Wardle, D.A.; Cros, C.; Fontaine, S. Rhizosphere control of soil nitrogen cycling: A key component of plant economic strategies. New Phytol. 2020, 228, 1269–1282. [Google Scholar] [CrossRef]
- Kuzyakov, Y.; Blagodatskaya, E. Microbial hotspots and hot moments in soil: Concept & review. Soil Biol. Biochem. 2015, 83, 184–199. [Google Scholar] [CrossRef]
- Liu, M.; Adl, S.; Cui, X.; Tian, Y.; Xu, X.; Kuzyakov, Y. In situ methods of plant-microbial interactions for nitrogen in rhizosphere. Rhizosphere 2020, 13, 100186. [Google Scholar] [CrossRef]
- Brzostek, E.R.; Greco, A.; Drake, J.E.; Finzi, A.C. Root carbon inputs to the rhizosphere stimulate extracellular enzyme activity and increase nitrogen availability in temperate forest soils. Biogeochemistry 2013, 115, 65–76. [Google Scholar] [CrossRef]
- Phillips, R.P.; Finzi, A.C.; Bernhardt, E.S. Enhanced root exudation induces microbial feedbacks to N cycling in a pine forest under long-term CO2 fumigation. Ecol. Lett. 2011, 14, 187–194. [Google Scholar] [CrossRef]
- Yin, H.; Li, Y.; Xiao, J.; Xu, Z.; Cheng, X.; Liu, Q. Enhanced root exudation stimulates soil nitrogen transformations in a subalpine coniferous forest under experimental warming. Glob. Change Biol. 2013, 19, 2158–2167. [Google Scholar] [CrossRef]
- Wang, Q.; Xiao, J.; Ding, J.; Zou, T.; Zhang, Z.; Liu, Q.; Yin, H. Differences in root exudate inputs and rhizosphere effects on soil N transformation between deciduous and evergreen trees. Plant Soil 2019, 458, 277–289. [Google Scholar] [CrossRef]
- Allen, K.; Corre, M.D.; Tjoa, A.; Veldkamp, E. Soil nitrogen-cycling responses to conversion of lowland forests to oil palm and rubber plantations in Sumatra, Indonesia. PLoS ONE 2015, 10, e0133325. [Google Scholar] [CrossRef] [Green Version]
- Kara, Ö.; Bolat, İ. Soil microbial biomass C and N changes in relation to forest conversion in the Northwestern Turkey. Land Degrad. Dev. 2008, 19, 421–428. [Google Scholar] [CrossRef]
- Cai, X.; Lin, Z.; Penttinen, P.; Li, Y.; Li, Y.; Luo, Y.; Yue, T.; Jiang, P.; Fu, W. Effects of conversion from a natural evergreen broadleaf forest to a Moso bamboo plantation on the soil nutrient pools, microbial biomass and enzyme activities in a subtropical area. For. Ecol. Manag. 2018, 422, 161–171. [Google Scholar] [CrossRef]
- Lin, Z.; Li, Y.; Tang, C.; Luo, Y.; Fu, W.; Cai, X.; Li, Y.; Yue, T.; Jiang, P.; Hu, S. Converting natural evergreen broadleaf forests to intensively managed moso bamboo plantations affects the pool size and stability of soil organic carbon and enzyme activities. Biol. Fertil. Soils 2018, 54, 467–480. [Google Scholar] [CrossRef]
- Meng, M.; Chen, H.Y.H.; Lin, J.; Liu, X.; Guo, X.; Yuan, Y.; Zhang, J. Long term forest conversion affected soil nanoscale pores in subtropical China. Catena 2020, 185, 104289. [Google Scholar] [CrossRef]
- Gower, S.T.; McMurtrie, R.E.; Murty, D. Aboveground net primary production decline with stand age: Potential causes. Trends. Ecol. Evol. 1996, 11, 378–382. [Google Scholar] [CrossRef]
- Tang, J.; Luyssaert, S.; Richardson, A.D.; Kutsch, W.; Janssens, I.A. Steeper declines in forest photosynthesis than respiration explain age-driven decreases in forest growth. Proc. Natl. Acad. Sci. USA 2014, 111, 8856–8860. [Google Scholar] [CrossRef] [Green Version]
- Muñoz-Villers, L.; McDonnell, J.J.H.; Sciences, E.S. Land use change effects on runoff generation in a humid tropical montane cloud forest region. Hydrol. Earth Syst. Sci. 2013, 17, 3543. [Google Scholar] [CrossRef] [Green Version]
- Oyarzun, C.; Aracena, C.; Rutherford, P.; Godoy, R.; Deschrijver, A. Effects of land use conversion from native forests to exotic plantations on nitrogen and phosphorus retention in catchments of southern Chile. Water Air Soil Pollut. 2007, 179, 341–350. [Google Scholar] [CrossRef]
- Zhu, X.; Liu, W.; Jiang, X.J.; Wang, P.; Li, W. Effects of land-use changes on runoff and sediment yield: Implications for soil conservation and forest management in Xishuangbanna, Southwest China. Land Degrad. Dev. 2018, 29, 2962–2974. [Google Scholar] [CrossRef]
- Perakis, S.S.; Hedin, L.O. Nitrogen loss from unpolluted South American forests mainly via dissolved organic compounds. Nature 2002, 415, 416–419. [Google Scholar] [CrossRef]
- Lewis, W.M., Jr. Nitrogen and Phosphorus Runoff Losses from a Nutrient-Poor Tropical Moist Forest. Ecology 1986, 67, 1275–1282. [Google Scholar] [CrossRef]
- Fang, Y.; Koba, K.; Makabe, A.; Takahashi, C.; Zhu, W.; Hayashi, T.; Hokari, A.A.; Urakawa, R.; Bai, E.; Houlton, B.Z. Microbial denitrification dominates nitrate losses from forest ecosystems. Proc. Natl. Acad. Sci. USA 2015, 112, 1470–1474. [Google Scholar] [CrossRef] [Green Version]
- Luo, X.; Hou, E.; Chen, J.; Li, J.; Zhang, L.; Zang, X.; Wen, D. Dynamics of carbon, nitrogen, and phosphorus stocks and stoichiometry resulting from conversion of primary broadleaf forest to plantation and secondary forest in subtropical China. Catena 2020, 193, 104606. [Google Scholar] [CrossRef]
- Medorio-García, H.P.; Alarcón, E.; Flores-Esteves, N.; Montaño, N.M.; Perroni, Y. Soil carbon, nitrogen and phosphorus dynamics in sugarcane plantations converted from tropical dry forest. Appl. Soil Ecol. 2020, 154, 103600. [Google Scholar] [CrossRef]
- Ross, D.J.; Tate, K.R.; Scott, N.A.; Feltham, C.W. Land-use change: Effects on soil carbon, nitrogen and phosphorus pools and fluxes in three adjacent ecosystems. Soil Biol. Biochem. 1999, 31, 803–813. [Google Scholar] [CrossRef]
- Xu, C.; Yang, Z.; Qian, W.; Chen, S.; Liu, X.; Lin, W.; Xiong, D.; Jiang, M.; Chang, C.-T.; Huang, J.-C.; et al. Runoff and soil erosion responses to rainfall and vegetation cover under various afforestation management regimes in subtropical montane forest. Land Degrad. Dev. 2019, 30, 1711–1724. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, L.; Yang, Z.; Xu, C.; Xie, J.; Chen, G.; Lin, C.; Guo, J.; Liu, X.; Xiong, D. Large ecosystem service benefits of assisted natural regeneration. J. Geophys. Res. Biogeosci. 2018, 123, 676–687. [Google Scholar] [CrossRef]
- Ostonen, I.; Lõhmus, K.; Pajuste, K. Fine root biomass, production and its proportion of NPP in a fertile middle-aged Norway spruce forest: Comparison of soil core and ingrowth core methods. For. Ecol. Manag. 2005, 212, 264–277. [Google Scholar] [CrossRef]
- Huang, Z.; Wan, X.; He, Z.; Yu, Z.; Wang, M.; Hu, Z.; Yang, Y. Soil microbial biomass, community composition and soil nitrogen cycling in relation to tree species in subtropical China. Soil Biol. Biochem. 2013, 62, 68–75. [Google Scholar] [CrossRef]
- Bååth, E.; Anderson, T.H. Comparison of soil fungal/bacterial ratios in a pH gradient using physiological and PLFA-based techniques. Soil Biol. Biochem. 2003, 35, 955–963. [Google Scholar] [CrossRef]
- Frostegård, Å.; Bååth, E.; Tunlio, A. Shifts in the structure of soil microbial communities in limed forests as revealed by phospholipid fatty acid analysis. Soil Biol. Biochem. 1993, 25, 723–730. [Google Scholar] [CrossRef]
- Frostegård, A.; Bååth, E. The use of phospholipid fatty acid analysis to estimate bacterial and fungal biomass in soil. Biol. Fertil. Soils 1996, 22, 59–65. [Google Scholar] [CrossRef]
- Näsholm, T.; Högberg, P.; Franklin, O.; Metcalfe, D.; Keel, S.G.; Campbell, C.; Hurry, V.; Linder, S.; Högberg, M.N. Are ectomycorrhizal fungi alleviating or aggravating nitrogen limitation of tree growth in boreal forests? New Phytol. 2013, 198, 214–221. [Google Scholar] [CrossRef]
- Saiya-Cork, K.R.; Sinsabaugh, R.L.; Zak, D.R. The effects of long term nitrogen deposition on extracellular enzyme activity in an Acer saccharum forest soil. Soil Biol. Biochem. 2002, 34, 1309–1315. [Google Scholar] [CrossRef]
- Liu, X.; Lin, T.-C.; Yang, Z.; Vadeboncoeur, M.A.; Lin, C.; Xiong, D.; Lin, W.; Chen, G.; Xie, J.; Li, Y. Increased litter in subtropical forests boosts soil respiration in natural forests but not plantations of Castanopsis carlesii. Plant Soil 2017, 418, 141–151. [Google Scholar] [CrossRef]
- Sun, D.; Zhang, W.; Lin, Y.; Liu, Z.; Shen, W.; Zhou, L.; Rao, X.; Liu, S.; Cai, X.-a.; He, D.; et al. Soil erosion and water retention varies with plantation type and age. For. Ecol. Manag. 2018, 422, 1–10. [Google Scholar] [CrossRef]
- Xu, C.; Lin, T.-c.; Yang, Z.; Liu, X.; Xiong, D.; Chen, S.; Wu, F.; Yang, Y. Forest conversion effects on soil organic carbon are regulated by soil aggregate stability and not by recalcitrance: Evidence from a reforestation experiment. Catena, 2022; under revision. [Google Scholar]
- Crouzeilles, R.; Ferreira, M.S.; Chazdon, R.L.; Lindenmayer, D.B.; Sansevero, J.B.B.; Monteiro, L.; Iribarrem, A.; Latawiec, A.E.; Strassburg, B.B.N. Ecological restoration success is higher for natural regeneration than for active restoration in tropical forests. Sci. Adv. 2017, 3, e1701345. [Google Scholar] [CrossRef] [Green Version]
- Uebel, K.; Wilson, K.A.; Shoo, L.P. Assisted natural regeneration accelerates recovery of highly disturbed rainforest. Ecol. Manag. Restor. 2017, 18, 231–238. [Google Scholar] [CrossRef]
- Dennis, P.G.; Miller, A.J.; Hirsch, P.R. Are root exudates more important than other sources of rhizodeposits in structuring rhizosphere bacterial communities? FEMS Microbiol. Ecol. 2010, 72, 313–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haichar, F.e.Z.; Marol, C.; Berge, O.; Rangel-Castro, J.I.; Prosser, J.I.; Balesdent, J.; Heulin, T.; Achouak, W. Plant host habitat and root exudates shape soil bacterial community structure. ISME J. 2008, 2, 1221–1230. [Google Scholar] [CrossRef]
- Kaiser, C.; Kilburn, M.R.; Clode, P.L.; Fuchslueger, L.; Koranda, M.; Cliff, J.B.; Solaiman, Z.M.; Murphy, D.V. Exploring the transfer of recent plant photosynthates to soil microbes: Mycorrhizal pathway vs. direct root exudation. New Phytol. 2015, 205, 1537–1551. [Google Scholar] [CrossRef]
- Paterson, E.; Gebbing, T.; Abel, C.; Sim, A.; Telfer, G. Rhizodeposition shapes rhizosphere microbial community structure in organic soil. New Phytol. 2007, 173, 600–610. [Google Scholar] [CrossRef]
- Cheng, Y.; Wang, J.; Chang, S.X.; Cai, Z.; Müller, C.; Zhang, J. Nitrogen deposition affects both net and gross soil nitrogen transformations in forest ecosystems: A review. Environ. Pollut. 2019, 244, 608–616. [Google Scholar] [CrossRef]
- Schlesinger, W.H. On the fate of anthropogenic nitrogen. Proc. Natl. Acad. Sci. USA 2009, 106, 203–208. [Google Scholar] [CrossRef] [Green Version]
- Isobe, K.; Ikutani, J.; Fang, Y.; Yoh, M.; Mo, J.; Suwa, Y.; Yoshida, M.; Senoo, K.; Otsuka, S.; Koba, K. Highly abundant acidophilic ammonia-oxidizing archaea causes high rates of nitrification and nitrate leaching in nitrogen-saturated forest soils. Soil Biol. Biochem. 2018, 122, 220–227. [Google Scholar] [CrossRef]
- Garten, C.T., Jr.; Ashwood, T.L. Landscape level differences in soil carbon and nitrogen: Implications for soil carbon sequestration. Glob. Biogeochem. Cycles 2002, 16, 149–162. [Google Scholar] [CrossRef]
- Maeda, Y.; Tashiro, N.; Enoki, T.; Urakawa, R.; Hishi, T. Effects of species replacement on the relationship between net primary production and soil nitrogen availability along a topographical gradient: Comparison of belowground allocation and nitrogen use efficiency between natural forests and plantations. For. Ecol. Manag. 2018, 422, 214–222. [Google Scholar] [CrossRef]
- Tateno, R.; Hishi, T.; Takeda, H. Above- and belowground biomass and net primary production in a cool-temperate deciduous forest in relation to topographical changes in soil nitrogen. For. Ecol. Manag. 2004, 193, 297–306. [Google Scholar] [CrossRef]
- Pang, D.; Cao, J.; Dan, X.; Guan, Y.; Peng, X.; Cui, M.; Wu, X.; Zhou, J. Recovery approach affects soil quality in fragile karst ecosystems of southwest China: Implications for vegetation restoration. Ecol. Eng. 2018, 123, 151–160. [Google Scholar] [CrossRef]
- Wang, Y.; Zheng, H.; Chen, F.; Yang, Y.; Zeng, J.; Van Nostrand, J.D.; Zhou, J.; Ouyang, Z. Artificial reforestation produces less diverse soil nitrogen-cycling genes than natural restoration. Ecosphere 2019, 10, e02562. [Google Scholar] [CrossRef]
- Madulid, D.A.; Tandang, D.N.; Esperanza Agoo, E.M.G. Assisted Natural Regeneration and Biodiversity in the Philippines; Food and Agriculture Office of the United Nations-Regional Office for the Asia and Pacific; RAP Publication: Bangkok, Thailand, 2010. [Google Scholar]
- Li, Z.; Tian, D.; Wang, B.; Wang, J.; Wang, S.; Chen, H.Y.H.; Xu, X.; Wang, C.; He, N.; Niu, S. Microbes drive global soil nitrogen mineralization and availability. Glob. Change Biol. 2018, 25, 1078–1088. [Google Scholar] [CrossRef]
- Tian, L.; Dell, E.; Shi, W. Chemical composition of dissolved organic matter in agroecosystems: Correlations with soil enzyme activity and carbon and nitrogen mineralization. Appl. Soil Ecol. 2010, 46, 426–435. [Google Scholar] [CrossRef]
- Moghimian, N.; Hosseini, S.M.; Kooch, Y.; Darki, B.Z. Impacts of changes in land use/cover on soil microbial and enzyme activities. Catena 2017, 157, 407–414. [Google Scholar] [CrossRef]
- Wang, Q.; Xiao, F.; He, T.; Wang, S. Responses of labile soil organic carbon and enzyme activity in mineral soils to forest conversion in the subtropics. Ann. For. Sci. 2013, 70, 579–587. [Google Scholar] [CrossRef] [Green Version]
- Xu, G.; Long, Z.; Ren, P.; Ren, C.; Cao, Y.; Huang, Y.; Hu, S. Differential responses of soil hydrolytic and oxidative enzyme activities to the natural forest conversion. Sci. Total Environ. 2020, 716, 136414. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.-S.; Yang, Z.-J.; Gao, R.; Xie, J.-S.; Guo, J.-F.; Huang, Z.-Q.; Yang, Y.-S. Carbon storage in a chronosequence of Chinese fir plantations in southern China. For. Ecol. Manag. 2013, 300, 68–76. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mature Secondary Forest | Young ANR Forest | Young Castanopsis Plantation | Young Chinese-Fir Plantation | |
|---|---|---|---|---|
| Bulk density (g cm−3) | 1.1 (0.03) b | 1.1 (0.03) b | 1.1 (0.06) ab | 1.2 (0.05) a |
| Fine root biomass (g m−2) | 436.4 (56.4) a | 334.0 (26.9) b | 186.7 (38.5) c | 244.2 (21.3) c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, C.; Lin, T.-C.; Huang, J.-C.; Yang, Z.; Liu, X.; Xiong, D.; Chen, S.; Wang, M.; Yang, L.; Yang, Y. Microbial Biomass Is More Important than Runoff Export in Predicting Soil Inorganic Nitrogen Concentrations Following Forest Conversion in Subtropical China. Land 2022, 11, 295. https://doi.org/10.3390/land11020295
Xu C, Lin T-C, Huang J-C, Yang Z, Liu X, Xiong D, Chen S, Wang M, Yang L, Yang Y. Microbial Biomass Is More Important than Runoff Export in Predicting Soil Inorganic Nitrogen Concentrations Following Forest Conversion in Subtropical China. Land. 2022; 11(2):295. https://doi.org/10.3390/land11020295
Chicago/Turabian StyleXu, Chao, Teng-Chiu Lin, Jr-Chuan Huang, Zhijie Yang, Xiaofei Liu, Decheng Xiong, Shidong Chen, Minhuang Wang, Liuming Yang, and Yusheng Yang. 2022. "Microbial Biomass Is More Important than Runoff Export in Predicting Soil Inorganic Nitrogen Concentrations Following Forest Conversion in Subtropical China" Land 11, no. 2: 295. https://doi.org/10.3390/land11020295
APA StyleXu, C., Lin, T.-C., Huang, J.-C., Yang, Z., Liu, X., Xiong, D., Chen, S., Wang, M., Yang, L., & Yang, Y. (2022). Microbial Biomass Is More Important than Runoff Export in Predicting Soil Inorganic Nitrogen Concentrations Following Forest Conversion in Subtropical China. Land, 11(2), 295. https://doi.org/10.3390/land11020295