
Selecting Graph Metrics with Ecological Significance for Deepening Landscape Characterization: Review and Applications

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
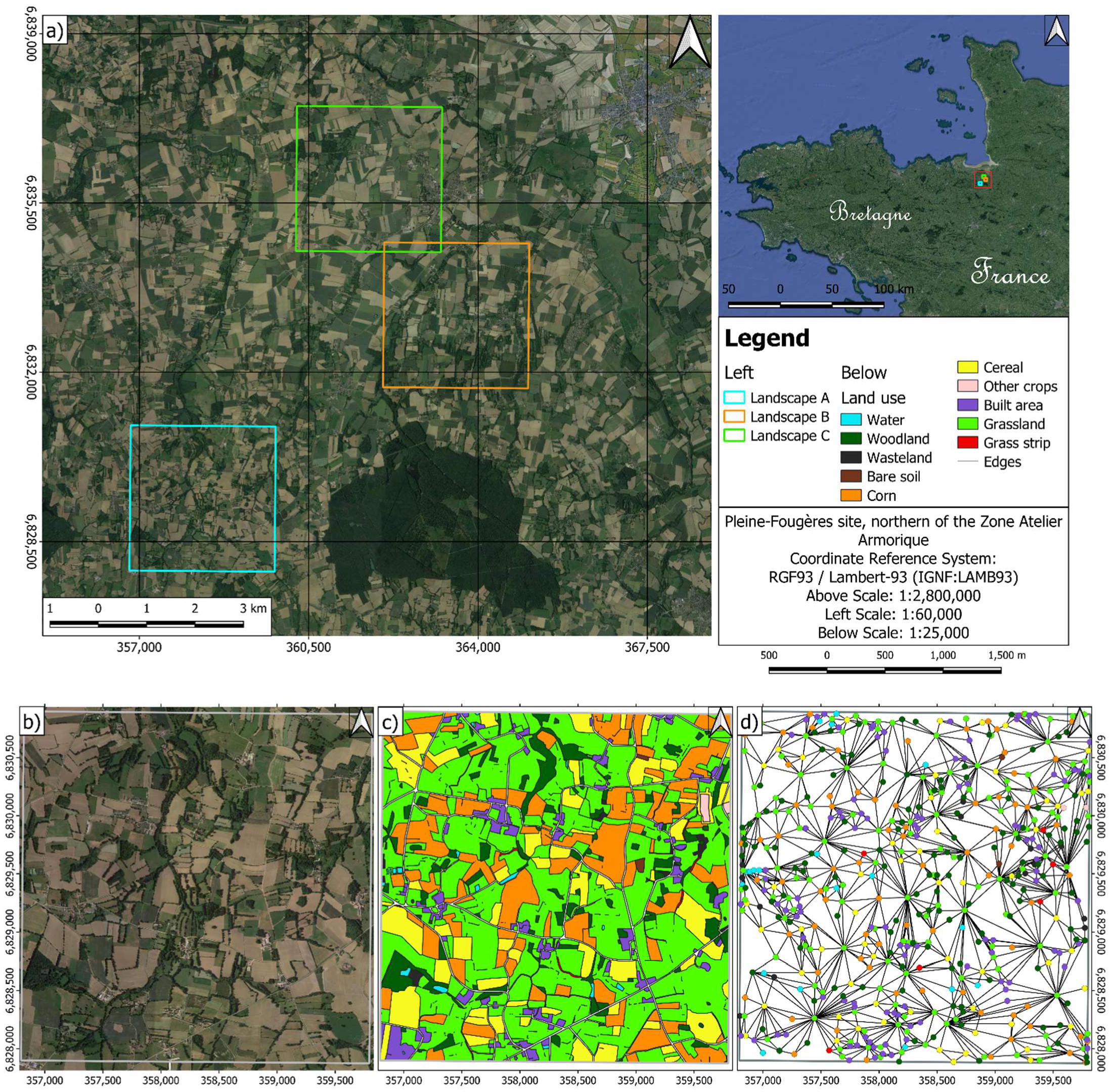
2.1. Landscape Graph Definition
2.2. Bibliographic Review and Graph Selection
2.3. Graph Metrics Application in an Agricultural Forest Mosaic
3. Results
3.1. Review of Graph Metrics
3.1.1. Graph Metrics at the Landscape Scale
Heterogeneity
Connectivity
Clustering
Assortativity
3.1.2. Graph Metrics at the Local Scale
Node Importance/Centrality
Compositional Heterogeneity
Configurational Heterogeneity
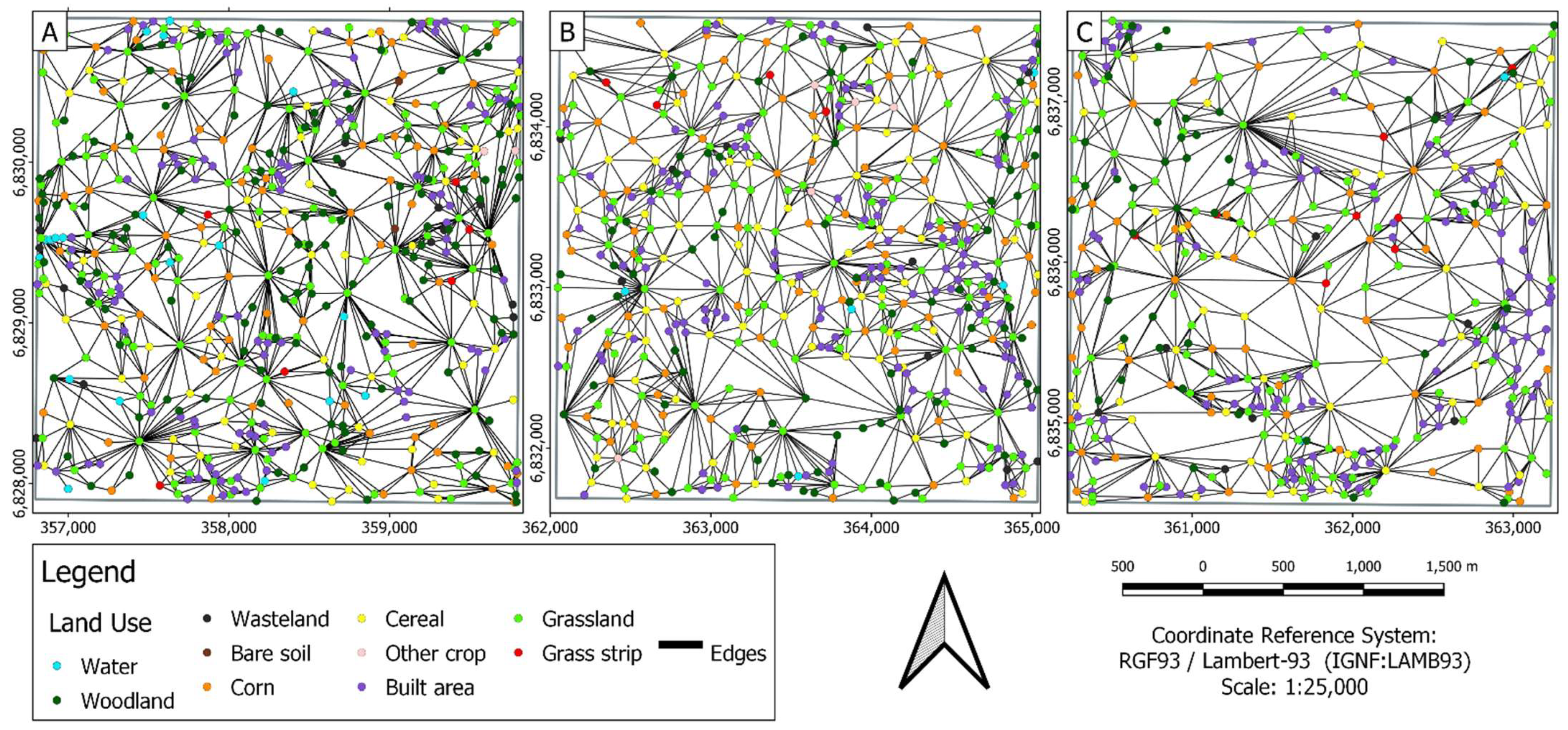
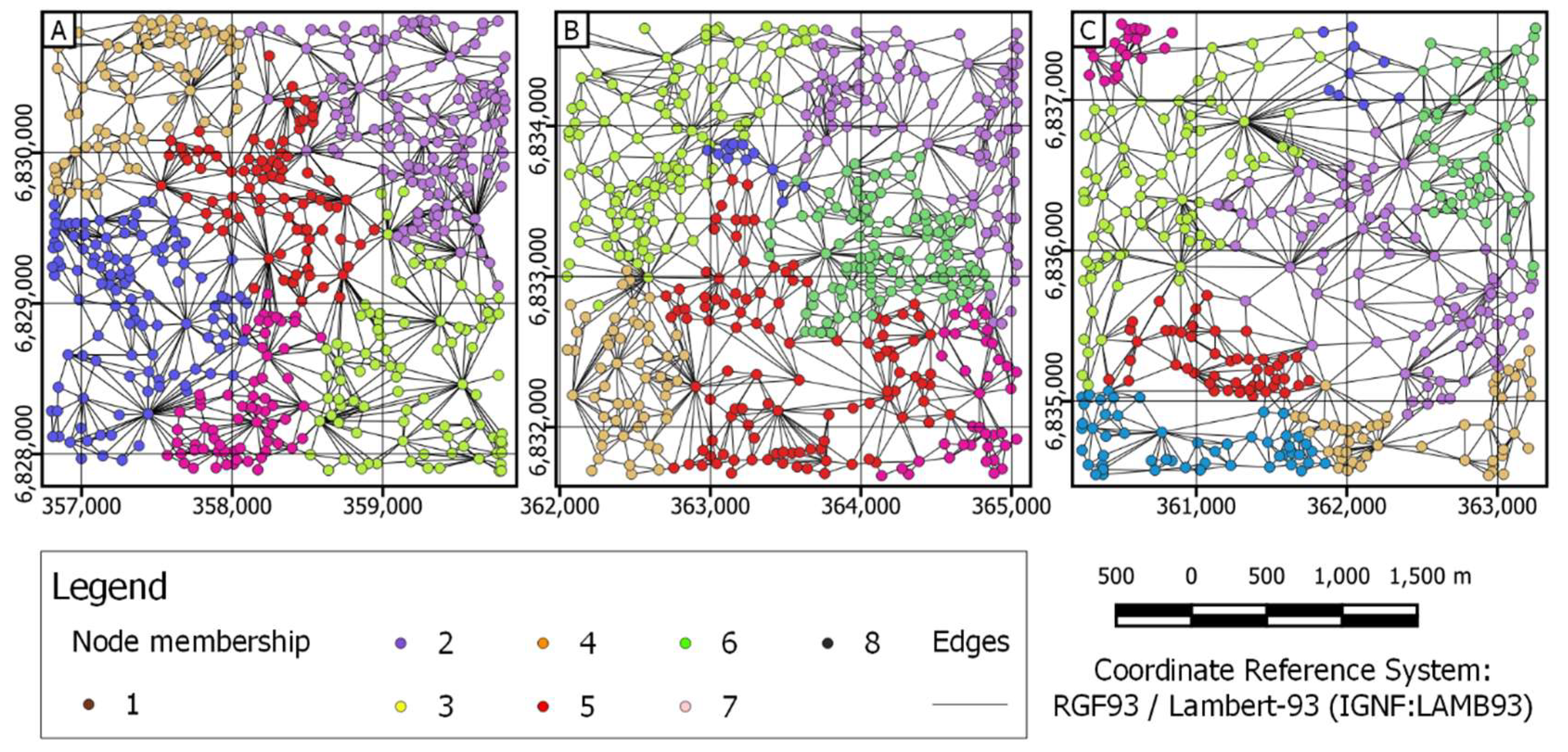
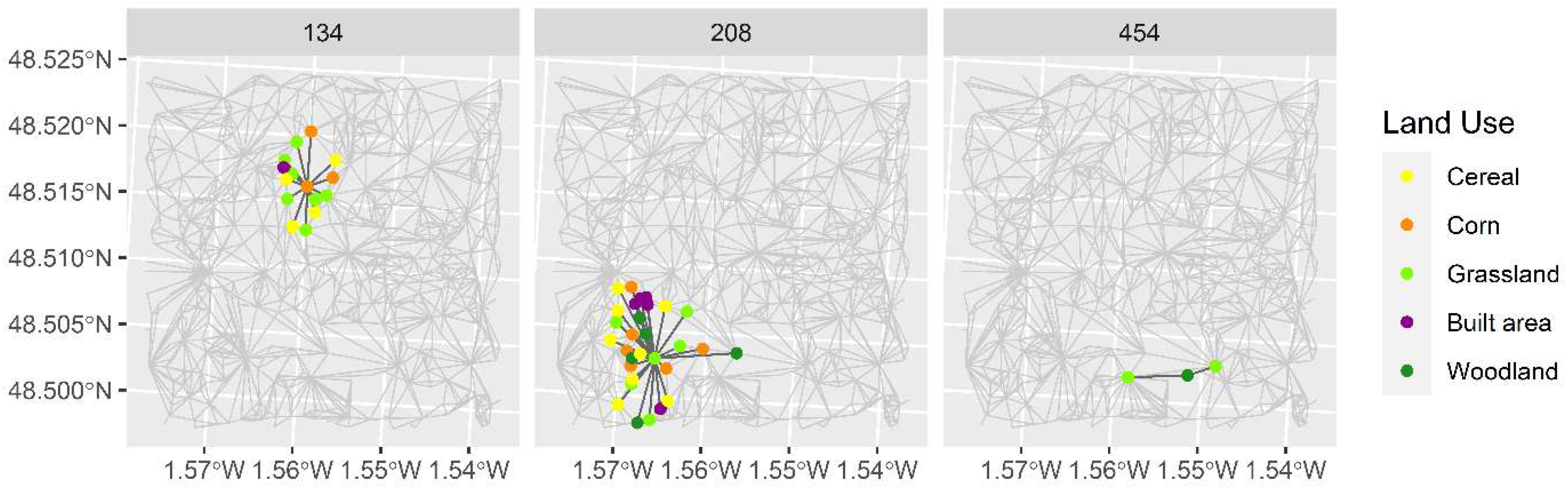
3.2. Graph Metric Application
3.2.1. Landscape Scale
3.2.2. Local Scale
4. Discussion
4.1. Graph Metric Analyses
4.2. Implementation of Graph Metrics
4.3. Case Study and Final Remarks
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Turner, M.G.; Gardner, R.H. Landscape Ecology in Theory and Practice: Pattern and Process, 2nd ed.; Springer: New York, NY, USA, 2015; ISBN 9781493927944. [Google Scholar]
- MacArthur, R.H.; Wilson, E.O. The Theory of Island Biogeography; Princeton University Press: Princeton, NJ, USA, 1967. [Google Scholar]
- Wiens, J.A. Landscape mosaics and ecological theory. In Mosaic Landscapes and Ecological Processes; Springer: Dordrecht, The Netherlands, 1995; pp. 1–26. [Google Scholar]
- Fahrig, L.; Baudry, J.; Brotons, L.; Burel, F.G.; Crist, T.O.; Fuller, R.J.; Sirami, C.; Siriwardena, G.M.; Martin, J.L. Functional landscape heterogeneity and animal biodiversity in agricultural landscapes. Ecol. Lett. 2011, 14, 101–112. [Google Scholar] [CrossRef] [PubMed]
- Pascual-Hortal, L.; Saura, S. Comparison and development of new graph-based landscape connectivity indices: Towards the priorization of habitat patches and corridors for conservation. Landsc. Ecol. 2006, 21, 959–967. [Google Scholar] [CrossRef]
- Saura, S.; Pascual-Hortal, L. A new habitat availability index to integrate connectivity in landscape conservation planning: Comparison with existing indices and application to a case study. Landsc. Urban. Plan. 2007, 83, 91–103. [Google Scholar] [CrossRef]
- Kupfer, J.A. Landscape ecology and biogeography: Rethinking landscape metrics in a post-FRAGSTATS landscape. Prog. Phys. Geogr. 2012, 36, 400–420. [Google Scholar] [CrossRef]
- Forman, R.T.T. Land Mosaics: The Ecology of Landscapes and Regions; University Press: Cambridge, UK, 1995; ISBN 9780521479806. [Google Scholar]
- Farina, A. Ecology, Cognition and Landscape. Linking Natural and Social Systems; Décamps, H., Ed.; Springer: Urbino, Italy, 2010; ISBN 978-90-481-3137-2. [Google Scholar]
- Estrada, E.; Fox, M.; Higham, D.J.; Oppo, G.-L. Network Science; Springer: London, UK, 2010; ISBN 978-1-84996-396-1. [Google Scholar]
- Green, D.G.; Klomp, N.; Rimmington, G.; Sadedin, S. Complexity in Landscape Ecology; Springer: Dordrecht, The Netherlands, 2006; ISBN 978-1-4020-4287-4. [Google Scholar]
- Ager, A.A.; Day, M.A.; Finney, M.A.; Vance-Borland, K.; Vaillant, N.M. Analyzing the transmission of wildfire exposure on a fire-prone landscape in Oregon, USA. For. Ecol. Manag. 2014, 334, 377–390. [Google Scholar] [CrossRef]
- Aquilué, N.; Filotas, É.; Craven, D.; Fortin, M.; Brotons, L.; Messier, C. Evaluating forest resilience to global threats using functional response traits and network properties. Ecol. Appl. 2020, 30. [Google Scholar] [CrossRef]
- Green, D.G. Connectivity and complexity in ecological systems. Pacific Conserv. Biol. 1994, 1, 194–200. [Google Scholar] [CrossRef]
- Hanski, I.; Alho, J.; Moilanen, A. Estimating the parameters of survival and migration of individuals in metapopulations. Ecology 2000, 81, 239–251. [Google Scholar] [CrossRef]
- Uuemaa, E.; Antrop, M.; Roosaare, J.; Marja, R. Landscape Metrics and Indices: An Overview of Their Use in Landscape Research Living Reviews in Landscape Research. Living Rev. Landsc. Res. 2009, 3, 1–28. [Google Scholar] [CrossRef]
- Cantwel, M.; Forman, R. Landscape graphs: Ecological modeling with graph theory to detect configurations common to diverse landscapes. Landsc. Ecol. 1993, 8, 239–255. [Google Scholar] [CrossRef]
- Foltête, J.C.; Savary, P.; Clauzel, C.; Bourgeois, M.; Girardet, X.; Saharoui, Y.; Vuidel, G.; Garnier, S. Coupling landscape graph modeling and biological data: A review. Landsc. Ecol. 2020, 35, 1035–1052. [Google Scholar] [CrossRef]
- Godet, C.; Clauzel, C. Comparison of landscape graph modelling methods for analysing pond network connectivity. Landsc. Ecol. 2020, 9, 735–748. [Google Scholar] [CrossRef]
- Rayfield, B.; Fortin, M.-J.; Fall, A. Connectivity for conservation: A framework to classify network measures. Ecology 2011, 92, 847–858. [Google Scholar] [CrossRef] [PubMed]
- De Franciscis, S.; Johnson, S.; Torres, J.J. Enhancing neural-network performance via assortativity. Phys. Rev. E Stat. Nonlinear Soft Matter Phys. 2011, 83, 36114. [Google Scholar] [CrossRef] [Green Version]
- Newman, M.E.J. Mixing patterns in networks. Phys. Rev. E Stat. Phys. Plasmas Fluids Relat. Interdiscip. Top. 2003, 67, 13. [Google Scholar] [CrossRef] [Green Version]
- Khosla, N.; Marsteller, J.A.; Hsu, Y.J.; Elliott, D.L. Analysing collaboration among HIV agencies through combining network theory and relational coordination. Soc. Sci. Med. 2016, 150, 85–94. [Google Scholar] [CrossRef]
- Wong, F.M.F.; Tan, C.W.; Sen, S.; Chiang, M. Quantifying political leaning from tweets, retweets, and retweeters. IEEE Trans. Knowl. Data Eng. 2016, 28, 2158–2172. [Google Scholar] [CrossRef] [Green Version]
- Han, C.; Kono, K.; Tanaka, S.; Kawakita, M.; Takeuchi, J. Botnet detection using graphical lasso with graph density. In Lecture Notes in Computer Science; Springer: Berlin/Heidelberg, Germany, 2016; Volume 9947, pp. 537–545. [Google Scholar]
- Urban, D.; Keitt, T. Landscape connectivity: A graph-theoretic perspective. Ecology 2001, 82, 1205–1218. [Google Scholar] [CrossRef]
- Dale, M.; Fortin, M.-J. Spatial Analysis a Guide for Ecologists, 2nd ed.; Cambridge University Press: Cambridge, UK, 2014; ISBN 978-0-52114350-9. [Google Scholar]
- Laita, A.; Kotiaho, J.S.; Mönkkönen, M. Graph-theoretic connectivity measures: What do they tell us about connectivity? Landsc. Ecol. 2011, 26, 951–967. [Google Scholar] [CrossRef]
- Mony, C.; Abadie, J.; Gil-Tena, A.; Burel, F.; Ernoult, A. Effects of connectivity on animal-dispersed forest plant communities in agriculture-dominated landscapes. J. Veg. Sci. 2018, 29, 167–178. [Google Scholar] [CrossRef]
- Upadhyay, S.; Roy, A.; Ramprakash, M.; Idiculla, J.; Kumar, A.S.; Bhattacharya, S. A network theoretic study of ecological connectivity in Western Himalayas. Ecol. Model. 2017, 359, 246–257. [Google Scholar] [CrossRef] [Green Version]
- Dunning, J.B.; Danielson, B.J.; Pulliam, H.R. Ecological Processes That Affect Populations in Complex Landscapes. Oikos 1992, 65, 169–175. [Google Scholar] [CrossRef] [Green Version]
- Tscharntke, T.; Tylianakis, J.M.; Rand, T.A.; Didham, R.K.; Fahrig, L.; Batáry, P.; Bengtsson, J.; Clough, Y.; Crist, T.O.; Dormann, C.F.; et al. Landscape moderation of biodiversity patterns and processes - eight hypotheses. Biol. Rev. 2012, 87, 661–685. [Google Scholar] [CrossRef]
- Correa Ayram, C.A.; Mendoza, M.E.; Etter, A.; Salicrup, D.R.P. Habitat connectivity in biodiversity conservation: A review of recent studies and applications. Prog. Phys. Geogr. 2016, 40, 7–37. [Google Scholar] [CrossRef]
- Uroy, L.; Ernoult, A.; Mony, C. Effect of landscape connectivity on plant communities: A review of response patterns. Landsc. Ecol. 2019, 34, 203–225. [Google Scholar] [CrossRef]
- Kwok Lun Cheung, A. Spatial and Temporal Topological Analysis of Landscape Structure Using Graph Theory; University of Auckland: Auckland, New Zealand, 2015. [Google Scholar]
- Fall, A.; Fortin, M.J.; Manseau, M.; O’Brien, D. Spatial graphs: Principles and applications for habitat connectivity. Ecosystems 2007, 10, 448–461. [Google Scholar] [CrossRef]
- Rahman, S. Basic Graph. Theory; Springer: Cham, Switzerland; Dhaka, Bangladesh, 2017; Volume 9, ISBN 978-3-319-49475-3. [Google Scholar]
- Le Coeur, D.; Baudry, J.; Burel, F. Field margins plant assemblages: Variation partitioning between local and landscape factors. Landsc. Urban. Plan. 1997, 37, 57–71. [Google Scholar] [CrossRef]
- Alignier, A. Two decades of change in a field margin vegetation metacommunity as a result of field margin structure and management practice changes. Agric. Ecosyst. Environ. 2018, 251, 1–10. [Google Scholar] [CrossRef]
- ESRI ArcGIS Desktop. 2016. Available online: https://www.safe.com/training/recorded/esri-arcgis-2016/ (accessed on 4 November 2019).
- Saura, S.; Pascual-Hortal, L. Conefor SENSINODE 2.2 User’s Manual. 2007. Available online: http://www.conefor.org/files/usuarios/CS22manual.pdf (accessed on 23 October 2020).
- Julve, P. Baseflor. Index Botanique, Écologique et Chorologique de la Flore de France. Available online: http://philippe.julve.pagesperso-orange.fr/catminat.htm (accessed on 4 November 2019).
- Labadessa, R.; Alignier, A.; Cassano, S.; Forte, L.; Mairota, P. Quantifying edge influence on plant community structure and composition in semi-natural dry grasslands. Appl. Veg. Sci. 2017, 20, 572–581. [Google Scholar] [CrossRef] [Green Version]
- Csardi, G.; Nepusz, T. The igraph software package for complex network research. InterJ. Complex Syst. 2006, 1695, 1–9. [Google Scholar]
- Qgis Development Team QGIS Geographic Information System (3.4.5); Open Source Geospatial Foundation: Madeira, Portugal, 2019.
- R Core Team. R: A Language and Environment for Statistical Computint (3.6.1); R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 23 October 2020).
- Amici, V.; Landi, S.; Frascaroli, F.; Rocchini, D.; Santi, E.; Chiarucci, A. Anthropogenic drivers of plant diversity: Perspective on land use change in a dynamic cultural landscape. Biodivers. Conserv. 2015, 24, 3185–3199. [Google Scholar] [CrossRef]
- Hammill, E.; Hawkins, C.P.; Greig, H.S.; Kratina, P.; Shurin, J.B.; Atwood, T.B. Landscape heterogeneity strengthens the relationship between β-diversity and ecosystem function. Ecology 2018, 99, 2467–2475. [Google Scholar] [CrossRef] [PubMed]
- Poggio, S.L.; Chaneton, E.J.; Ghersa, C.M. Landscape complexity differentially affects alpha, beta, and gamma diversities of plants occurring in fencerows and crop fields. Biol. Conserv. 2010, 143, 2477–2486. [Google Scholar] [CrossRef]
- Mandelik, Y.; Winfree, R.; Neeson, T.; Kremen, C. Complementary habitat use by wild bees in agro-natural landscapes. Ecol. Appl. 2012, 22, 1535–1546. [Google Scholar] [CrossRef] [PubMed]
- Schneider, G.; Krauss, J.; Boetzl, F.A.; Fritze, M.A.; Steffan-Dewenter, I. Spillover from adjacent crop and forest habitats shapes carabid beetle assemblages in fragmented semi-natural grasslands. Oecologia 2016, 182, 1141–1150. [Google Scholar] [CrossRef]
- Wasserman, S. Social Network Analysis: Methods and Applications; Structural Analysis in the Social Sciences; 8; Cambridge University Press: Cambridge, UK, 1994; ISBN 0521387078. [Google Scholar]
- West, D.B. Introduction to Graph Theory, 2nd ed.; Prentice Hall: Hoboken, NJ, USA, 2001; ISBN 0-13-014400-2. [Google Scholar]
- Concepción, E.D.; Fernandez-González, F.; Díaz, M. Plant diversity partitioning in Mediterranean croplands: Effects of farming intensity, field edge, and landscape context. Ecol. Appl. 2012, 22, 972–981. [Google Scholar] [CrossRef]
- Herrera-Arroyo, M.L.; Sork, V.L.; González-Rodríguez, A.; Rocha-Ramírez, V.; Vega, E.; Oyama, K. Seed-mediated connectivity among fragmented populations of Quercus castanea (Fagaceae) in a Mexican landscape. Am. J. Bot. 2013, 100, 1663–1671. [Google Scholar] [CrossRef] [Green Version]
- Ferrari, J.R.; Lookingbill, T.R.; Neel, M.C. Two measures of landscape-graph connectivity: Assessment across gradients in area and configuration. Landsc. Ecol. 2007, 22, 1315–1323. [Google Scholar] [CrossRef]
- Verheijen, B.H.F.; Varner, D.M.; Haukos, D.A. Effects of large-scale wetland loss on network connectivity of the Rainwater Basin, Nebraska. Landsc. Ecol. 2018, 33, 1939–1951. [Google Scholar] [CrossRef] [Green Version]
- Raghavan, U.N.; Albert, R.; Kumara, S. Near linear time algorithm to detect community structures in large-scale networks. Phys. Rev. E Stat. Nonlinear Soft Matter Phys. 2007, 76, 036106. [Google Scholar] [CrossRef] [Green Version]
- Clauset, A.; Newman, M.E.J.; Moore, C. Finding community structure in very large networks. Phys. Rev. E Stat. Phys. Plasmas Fluids Relat. Interdiscip. Top. 2004, 70, 066111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radicchi, F.; Castellano, C.; Cecconi, F.; Loreto, V.; Paris, D. Defining and identifying communities in networks. Proc. Natl. Acad. Sci. USA 2004, 101, 2658–2663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newman, M.E.J.; Girvan, M. Finding and evaluating community structure in networks. Phys. Rev. E Stat. Nonlinear Soft Matter Phys. 2004, 69, 026113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newman, M.E.J. Finding community structure in networks using the eigenvectors of matrices. Phys. Rev. E Stat. Nonlinear Soft Matter Phys. 2006, 74, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noldus, R.; Mieghem, P. Van Assortativity in complex networks. J. Complex. Netw. 2015, 3, 507–542. [Google Scholar] [CrossRef] [Green Version]
- Newman, M.E.J. Assortative Mixing in Networks. Phys. Rev. Lett. 2002, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weibull, A.C.; Östman, Ö.; Agranqvist, A. Species richness in agroecosystems: The effect of landscape, habitat and farm management. Biodivers. Conserv. 2003, 12, 1335–1355. [Google Scholar] [CrossRef]
- Otterstatter, M.C.; Thomson, J.D. Contact networks and transmission of an intestinal pathogen in bumble bee (Bombus impatiens) colonies. Oecologia 2007, 154, 411–421. [Google Scholar] [CrossRef]
- Seidman, S.B. Network structure and minimum degree. Soc. Netw. 1983, 5, 269–287. [Google Scholar] [CrossRef]
- Van Wijk, B.C.M.; Stam, C.J.; Daffertshofer, A. Comparing Brain Networks of Different Size and Connectivity Density Using Graph Theory. PLoS ONE 2010, 5, 13701. [Google Scholar] [CrossRef]
- Danon, L.; Díaz-Guilera, A.; Duch, J.; Arenas, A. Comparing community structure identification. J. Stat. Mech. Theory Exp. 2005, P09008. [Google Scholar] [CrossRef]
- Newman, M.E.J. Communities, modules and large-scale structure in networks. Nat. Phys. 2012, 8, 25–31. [Google Scholar] [CrossRef]
- Han, M.; Yan, M.; Cai, Z.; Li, Y.; Cai, X.; Yu, J. Influence maximization by probing partial communities in dynamic online social networks. Trans. Emerg. Telecommun. Technol. 2017, 28. [Google Scholar] [CrossRef]
- Rojas, A.; Patarroyo, P.; Mao, L.; Bengtson, P.; Kowalewski, M. Global biogeography of Albian ammonoids: A network-based approach. Geology 2017, 45, 659–662. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Cao, G.; Sun, X.; Lee, J.; Rubin, D.L.; Napel, S.; Kurian, A.W.; Daniel, B.L.; Li, R. Intratumoral spatial heterogeneity at perfusion MR imaging predicts recurrence-free survival in locally advanced breast cancer treated with neoadjuvant chemotherapy. Radiology 2018, 288, 26–35. [Google Scholar] [CrossRef]
- Buglass, S.L.; Binder, J.F.; Betts, L.R.; Underwood, J.D.M. When “friends” collide: Social heterogeneity and user vulnerability on social network sites. Comput. Hum. Behav. 2016, 54, 62–72. [Google Scholar] [CrossRef] [Green Version]
- Preti, M.G.; Bolton, T.A.; Van De Ville, D. The dynamic functional connectome: State-of-the-art and perspectives. Neuroimage 2017, 160, 41–54. [Google Scholar] [CrossRef]
- Eguíluz, V.M.; Chialvo, D.R.; Cecchi, G.A.; Baliki, M.; Apkarian, A.V. Scale-free brain functional networks. Phys. Rev. Lett. 2005, 94, 018102. [Google Scholar] [CrossRef] [Green Version]
- Long, X.; Fonseca, P.; Aarts, R.M.; Haakma, R.; Foussier, J. Modeling cardiorespiratory interaction during human sleep with complex networks. Appl. Phys. Lett. 2014, 105, 203701. [Google Scholar] [CrossRef] [Green Version]
- Bae, J.; Kim, S. Identifying and ranking influential spreaders in complex networks by neighborhood coreness. Physica A 2014, 395, 549–559. [Google Scholar] [CrossRef]
- Batagelj, V.; Zaversnik, M. An O(m) Algorithm for Cores Decomposition of Networks. Adv. Data Anal. Classif. 2011, 5, 129–145. [Google Scholar] [CrossRef]
- Bonacich, P. Power and Centrality: A Family of Measures. Am. J. Sociol. 2002, 92, 1170–1182. [Google Scholar] [CrossRef]
- Borgatti, S.P. Centrality and network flow. Soc. Netw. 2005, 27, 55–71. [Google Scholar] [CrossRef]
- Freeman, L. Centrality in Social Networks I: Conceptual Clarification. Soc. Netw. 1979, 1, 215–239. [Google Scholar] [CrossRef] [Green Version]
- Newman, M.E.J. Networks an Introduction; Oxford University Press: Oxford, UK, 2010; ISBN 0-19-159417-2. [Google Scholar]
- Van Dijk, W.F.A.; Van Ruijven, J.; Berendse, F.; de Snoo, G.R. The effectiveness of ditch banks as dispersal corridor for plants in agricultural landscapes depends on species’ dispersal traits. Biol. Conserv. 2014, 171, 91–98. [Google Scholar] [CrossRef]
- Allouche, O.; Kalyuzhny, M.; Moreno-Rueda, G.; Pizarro, M.; Kadmon, R. Area-heterogeneity tradeoff and the diversity of ecological communities. Proc. Natl. Acad. Sci. USA 2012, 109, 17495–17500. [Google Scholar] [CrossRef] [Green Version]
- Gámez-Virués, S.; Perović, D.J.; Perović, P.; Gossner, M.M.; Börschig, C.; Blüthgen, N.; De Jong, H.; Simons, N.K.; Klein, A.-M.; Krauss, J.; et al. Landscape simplification filters species traits and drives biotic homogenization. Nat. Commun. 2015, 6. [Google Scholar] [CrossRef]
- Sirami, C.; Gross, N.; Baillod, A.B.; Bertrand, C.; Carrié, R.; Hass, A.; Henckel, L.; Miguet, P.; Vuillot, C.; Alignier, A.; et al. Increasing crop heterogeneity enhances multitrophic diversity across agricultural regions. Proc. Natl. Acad. Sci. USA 2019, 116, 16442–16447. [Google Scholar] [CrossRef] [Green Version]
- Fahrig, L.; Girard, J.; Duro, D.; Pasher, J.; Smith, A.; Javorek, S.; King, D.; Lindsay, K.F.; Mitchell, S.; Tischendorf, L. Farmlands with smaller crop fields have higher within-field biodiversity. Agric. Ecosyst. Environ. 2015, 200, 219–234. [Google Scholar] [CrossRef]
- Hass, A.L.; Kormann, U.G.; Tscharntke, T.; Clough, Y.; Baillod, A.B.; Sirami, C.; Fahrig, L.; Martin, J.L.; Baudry, J.; Bertrand, C.; et al. Landscape configurational heterogeneity by small-scale agriculture, not crop diversity, maintains pollinators and plant reproduction in western Europe. Proc. R. Soc. B Biol. Sci. 2018, 285. [Google Scholar] [CrossRef]
- Hodgson, J.G.; Grime, J.P.; Hunt, R.; Thompson, K. The Electronic Comparative Plant. Ecology; Springer Science+Business Media: Dordrecht, The Netherlands, 1995; ISBN 978-94-011-7652-1. [Google Scholar]
- Minor, E.S.; Urban, D.L. A Graph-Theory Framework for Evaluating Landscape Connectivity and Conservation Planning. Conserv. Biol. 2008, 22, 297–307. [Google Scholar] [CrossRef] [PubMed]
- Hackett, T.D.; Sauve, A.M.C.; Davies, N.; Montoya, D.; Tylianakis, J.M.; Memmott, J. Reshaping our understanding of species’ roles in landscape-scale networks. Ecol. Lett. 2019. [Google Scholar] [CrossRef] [PubMed]
- Mbaru, E.K.; Barnes, M.L. Key players in conservation diffusion: Using social network analysis to identify critical injection points. Biol. Conserv. 2017, 210, 222–232. [Google Scholar] [CrossRef]
- Rubinov, M.; Sporns, O. Complex network measures of brain connectivity: Uses and interpretations. Neuroimage 2010, 52, 1059–1069. [Google Scholar] [CrossRef] [PubMed]
- Vilchis, F.L. Análisis estructural de la red económica de exportaciones e importaciones de la Alianza del Pacífico y el Mercado Común del Sur. Estud. Gerenc. 2016, 32, 319–325. [Google Scholar] [CrossRef]
- Beauchamp, M.A. an Improved Index of Centrality. Behav. Sci. 1965, 10, 161–163. [Google Scholar] [CrossRef]
- Henriksen, L.F.; Seabrooke, L. Transnational organizing: Issue professionals in environmental sustainability networks. Organization 2016, 23, 722–741. [Google Scholar] [CrossRef] [Green Version]
- Bonacich, P. Factoring and weighting approaches to status scores and clique identification. J. Math. Sociol. 1972, 2, 113–120. [Google Scholar] [CrossRef]
- Pereira, M.; Segurado, P.; Neves, N. Using spatial network structure in landscape management and planning: A case study with pond turtles. Landsc. Urban. Plan. 2011, 100, 67–76. [Google Scholar] [CrossRef]
- Ahmed, H.; Howton, T.C.; Sun, Y.; Weinberger, N.; Belkhadir, Y.; Mukhtar, M.S. Network biology discovers pathogen contact points in host protein-protein interactomes. Nat. Commun. 2018, 9, 1–14. [Google Scholar] [CrossRef]
- Sun, H.-L.; Ch’ng, E.; See, S. Influential spreaders in the political Twitter sphere of the 2013 Malaysian general election. Ind. Manag. Data Syst. 2019, 119, 54–68. [Google Scholar] [CrossRef]
- Andreasen, C.; Streibig, J.C.; Haas, H. Soil properties affecting the distribution of 37 weed species in Danish fields. Weed Res. 1991, 31, 181–187. [Google Scholar] [CrossRef]
- McGarigal, K.; Cushman, S.A.; Ene, E. FRAGSTATS v4: Spatial Pattern Analysis Program for Categorical and Continuous Maps. Computer Software Program Produced by the Authors at the University of Massachusetts (No. 4). 2012. Available online: http://www.umass.edu/landeco/research/fragstats/fragstats.html (accessed on 16 November 2020).
- Gustafson, E.J. How has the state-of-the-art for quantification of landscape pattern advanced in the twenty-first century? Landsc. Ecol. 2019, 34, 2065–2072. [Google Scholar] [CrossRef]
- Frazier, A.E.; Kedron, P. Landscape Metrics: Past Progress and Future Directions. Curr. Landsc. Ecol. Reports 2017, 2, 63–72. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Wu, J. Use and misuse of landscape Índices. Landsc. Ecol. 2004, 19, 389–399. [Google Scholar] [CrossRef] [Green Version]
- Calabrese, J.M.; Fagan, W.F. A comparison-shopper’s guide to connectivity metrics. Front. Ecol. Environ. 2004, 2, 529–536. [Google Scholar] [CrossRef]
- Hesselbarth, M.H.K.; Sciaini, M.; With, K.A.; Wiegand, K.; Nowosad, J. Landscapemetrics: An open-source R tool to calculate landscape metrics. Ecography 2019, 42, 1648–1657. [Google Scholar] [CrossRef] [Green Version]
- Hijmans, R.J. Raster: Geographic Data Analysis and Modeling. R Package. 2020. Available online: https://cran.r-project.org/package=raster (accessed on 23 October 2020).
- Pebesma, E. Simple features for R: Standardized support for spatial vector data. R J. 2018, 10, 439–446. [Google Scholar] [CrossRef] [Green Version]
- Godínez-Gómez, O.; Correa-Ayram, C. Makurhini: Analyzing Landscape Connectivity. 2020. Available online: https://connectscape.github.io/Makurhini/index.html (accessed on 25 June 2021). [CrossRef]
- Savary, P.; Foltête, J.; Moal, H.; Vuidel, G.; Garnier, S. graph4lg: A package for constructing and analysing graphs for landscape genetics in R. Methods Ecol. Evol. 2020. [Google Scholar] [CrossRef]
- Dale, M.; Fortin, M.J. From Graphs to Spatial Graphs. Annu. Rev. Ecol. Evol. Syst. 2010, 41, 21–38. [Google Scholar] [CrossRef]
- Savary, P.; Foltête, J.C.; Moal, H.; Vuidel, G.; Garnier, S. Analysing landscape effects on dispersal networks and gene flow with genetic graphs. Mol. Ecol. Resour. 2021, 21, 1167–1185. [Google Scholar] [CrossRef] [PubMed]
- Burel, F.; Baudry, J.; Butet, A.; Clergeau, P.; Delettre, Y.; Le Coeur, D.; Dubs, F.; Morvan, N.; Paillat, G.; Petit, S.; et al. Comparative biodiversity along a gradient of agricultural landscapes. Acta Oecologica 1998, 19, 47–60. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Metric (Abbreviation) | Calculation | Interpretation | Ecological Meaning | References |
|---|---|---|---|---|---|
| Landscape/graph heterogeneity | Number of nodes (N nodes) | How many patches does the landscape have | The higher the value, the higher the heterogeneity of the landscape | [47,48,49] | |
| Total perimeter (Total per) | Sum of the perimeters of all patches | ||||
| Landscape/graph connectivity | Graph density (Density) | Ratio of the number of edges to the number of possible edges | The higher the value, the greater the connectivity/interactions between patches | [50,51,52] | |
| Number of edges (N edges) | How many connections does this landscape have | [4,37,50,51,53,54,55] | |||
| Maximum degree (Max degree) | Number of links of the most connected patch | ||||
| Mean node degree (Mean degree) | Average number of connections per patch | ||||
| Median node degree (Median degree) | The “middle” value of the degree of nodes | ||||
| Graph diameter (Diameter) | Longest geodesic distance (length of the shortest path) between nodes | The lower the value, the greater the connectivity/interactions between patches | [53,56,57] | ||
| Mean node distance (Mean distance) | Average geodesic distance between patches | ||||
| Cluster metrics | Number of communities based on propagating labels (N CLP) | See text (details in [58]). | Number of identified groups based on different methods Measures how clustered the graph is | The higher the value, the higher the heterogeneity of the landscape | [30,58,59,60,61,62] |
| Number of communities based on greedy optimization of modularity (N CGO) | Max | ||||
| Number of communities based on the leading eigenvector of the community matrix (N CLEV) | Max [Q] See text (details in [62]) | ||||
| Assortativity coefficient | Degree of assortativity (Ass deg) | The assortativity coefficient is positive if similar vertices (based on some external property) tend to connect between them. | The lower the value, the lower the robustness of landscape flow (of organism) | [22,30,63,64] |
| Group | Metrics (abb.) | Calculation | Interpretation | Ecological Meaning | References |
|---|---|---|---|---|---|
| Node importance | Node degree (Degree) | Measures the mean of node importance in relation to landscape connectivity | The greater the importance or centrality, the higher the flow (or fluxes) | [12,30,42,78,79,80,81,82,83,84] | |
| Node closeness (Closeness) | |||||
| Node eigenvector (Eigenvector) | → | ||||
| Node Bonacich power (B power) | |||||
| Node coreness (Coreness) | Mean of coreness (see text and details in [79]. | ||||
| Compositional heterogeneity | Community based on propagating labels (N CLP) | Number of community members surrounding each node | Identify group membership on the basis of main land use | The higher the diversity of communities or patches, the higher the local/patch heterogeneity | [4,29,50,51,65,85,86,87] |
| Configurational heterogeneity | Community based on greedy optimization of modularity (N CGO) | Number of community members surrounding each node after Max [Q] | Identify group membership on the basis of local topological structure | The greater the number of communities surrounding the node, the higher the local/patch heterogeneity | [4,29,30,54,87,88,89] |
| Community based on the leading eigenvector of the community matrix (NCLEV) | Number of community members surrounding each node after Max [Q] | ||||
| Patch area (Area) | Area of the patches/nodes | With more area, the patch is bigger | The larger the area, the lower the local/patch heterogeneity | ||
| Patch perimeter (Perimeter) | Perimeter of patches/nodes | A patch with a larger perimeter is bigger or more complex | The larger the perimeter, the higher the local/patch heterogeneity |
| Typology | Parallel Duplication | Serial Duplication | Transfer |
|---|---|---|---|
| Geodesics | dPCflux + dPCconnector | Freeman closeness (Mitotic reproduction) | Freeman closeness Freeman betweenness (Package delivery) |
| Paths | Freeman closeness Degree centrality (Internet name server) | (Viral infection) | (Mooch) |
| Trails | Freeman closeness Degree centrality (E-mail broadcast) | (Gossip) | (Used goods) |
| Walks | Freeman closeness Degree centrality Bonacich eigenvector (Attitude influencing) | (Emotional support) | (Money exchange) |
| Group | Metric | Landscape A | Landscape B | Landscape C |
|---|---|---|---|---|
| Landscape/graph heterogeneity | Number of nodes | 553 | 506 | 395 |
| Total perimeter | 314,451.41 | 295,448.28 | 252,918.45 | |
| Landscape/graph connectivity | Graph density | 9.96 × 10−3 | 11.30 × 10−3 | 14.08 × 10−3 |
| Number of edges | 1520 | 1444 | 1096 | |
| Maximum degree | 31 | 29 | 33 | |
| Mean node degree | 5.49 | 5.71 | 5.55 | |
| Median node degree | 4 | 5 | 5 | |
| Graph diameter | 15 | 16 | 16 | |
| Mean node distance | 6.32 | 6.72 | 6.15 | |
| Cluster metrics | Number of communities based on propagating labels | 3 | 6 | 5 |
| Number of communities based on greedy optimization of modularity | 6 | 7 | 8 | |
| Number of communities based on the leading eigenvector of the community matrix | 12 | 12 | 11 | |
| Assortativity coefficient | Degree of assortativity | −0.20 | −0.18 | −0.13 |
| Group | Metric | Node 134 | Node 208 | Node 454 |
|---|---|---|---|---|
| Node importance | Node degree (degree) | 14 | 29 | 2 |
| Node closeness (closeness) | 0.20 | 0.18 | 0.16 | |
| Node eigenvector (eigenvector) | 0.05 | 1 | 0.06 | |
| Node Bonacich power (B Power) | 4.96 × 10−3 | 12.13 × 10−3 | 1.51 × 10−3 | |
| Node coreness (Coreness) | 4 | 4 | 2 | |
| Compositional heterogeneity | Community based on propagating labels (N CLP) | 2 | 3 | 1 |
| Configurational heterogeneity | Community based on greedy optimization of modularity (N CGO) | 5 | 2 | 1 |
| Community based on the leading eigenvector of the community matrix (NCLEV) | 3 | 2 | 1 | |
| Patch area (m2) | 82,877.01 | 197,840.49 | 903.87 | |
| Patch perimeter (m) | 1624.94 | 4226.61 | 132.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de la Barra, F.; Alignier, A.; Reyes-Paecke, S.; Duane, A.; Miranda, M.D. Selecting Graph Metrics with Ecological Significance for Deepening Landscape Characterization: Review and Applications. Land 2022, 11, 338. https://doi.org/10.3390/land11030338
de la Barra F, Alignier A, Reyes-Paecke S, Duane A, Miranda MD. Selecting Graph Metrics with Ecological Significance for Deepening Landscape Characterization: Review and Applications. Land. 2022; 11(3):338. https://doi.org/10.3390/land11030338
Chicago/Turabian Stylede la Barra, Felipe, Audrey Alignier, Sonia Reyes-Paecke, Andrea Duane, and Marcelo D. Miranda. 2022. "Selecting Graph Metrics with Ecological Significance for Deepening Landscape Characterization: Review and Applications" Land 11, no. 3: 338. https://doi.org/10.3390/land11030338
APA Stylede la Barra, F., Alignier, A., Reyes-Paecke, S., Duane, A., & Miranda, M. D. (2022). Selecting Graph Metrics with Ecological Significance for Deepening Landscape Characterization: Review and Applications. Land, 11(3), 338. https://doi.org/10.3390/land11030338