
Climate Change Impacts on Grassland Vigour in Northern Portugal

 ,
,  , ,
, ,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
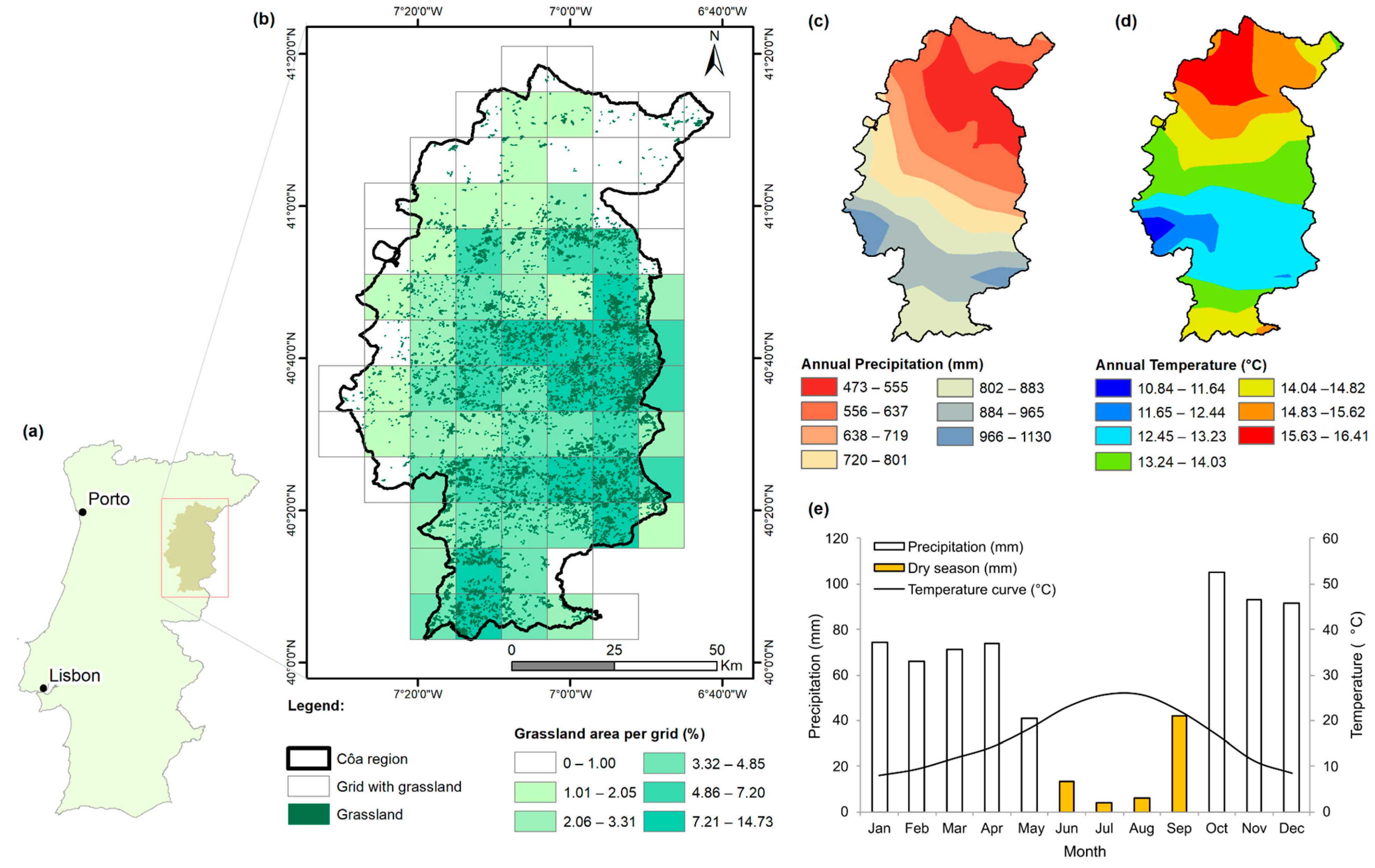
2.1. Study Area
2.2. Future Vigour Model
2.2.1. Grassland Data
2.2.2. NDVI Data
2.2.3. Climatic Data
2.2.4. Data Integration
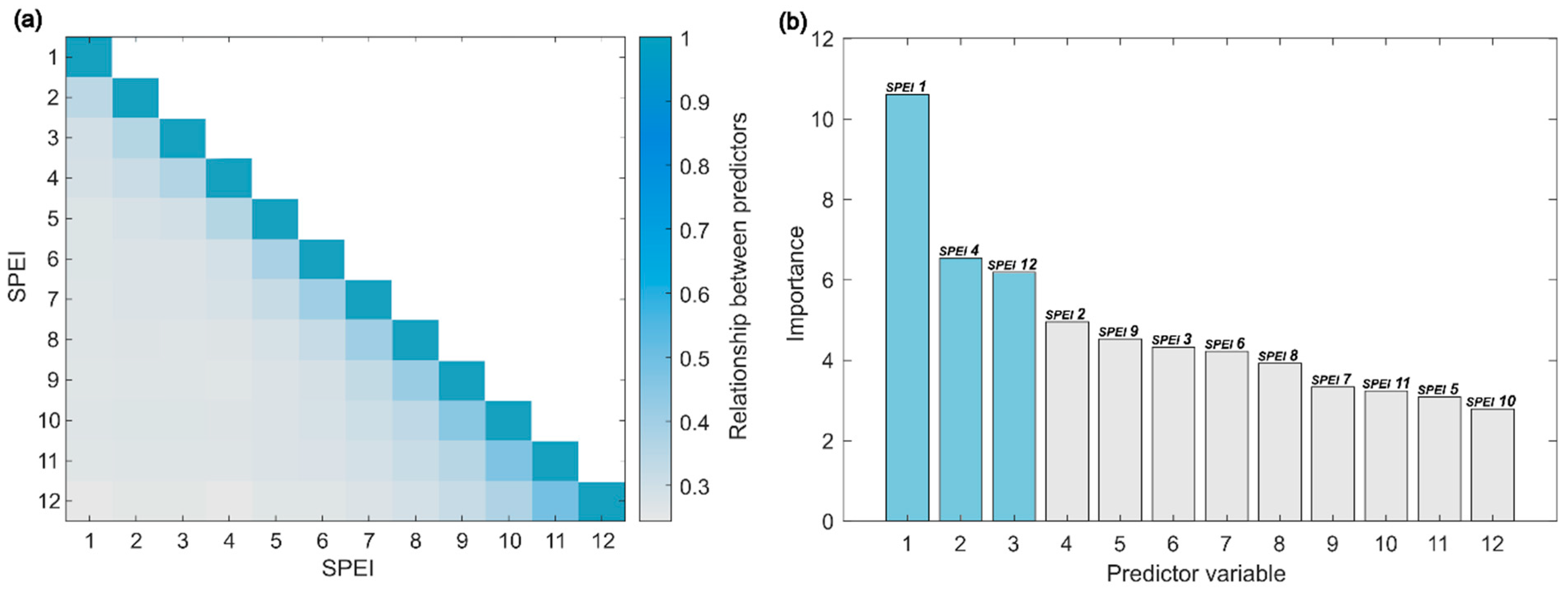
2.3. Data Analysis and Modelling
2.4. Future Projections
3. Results
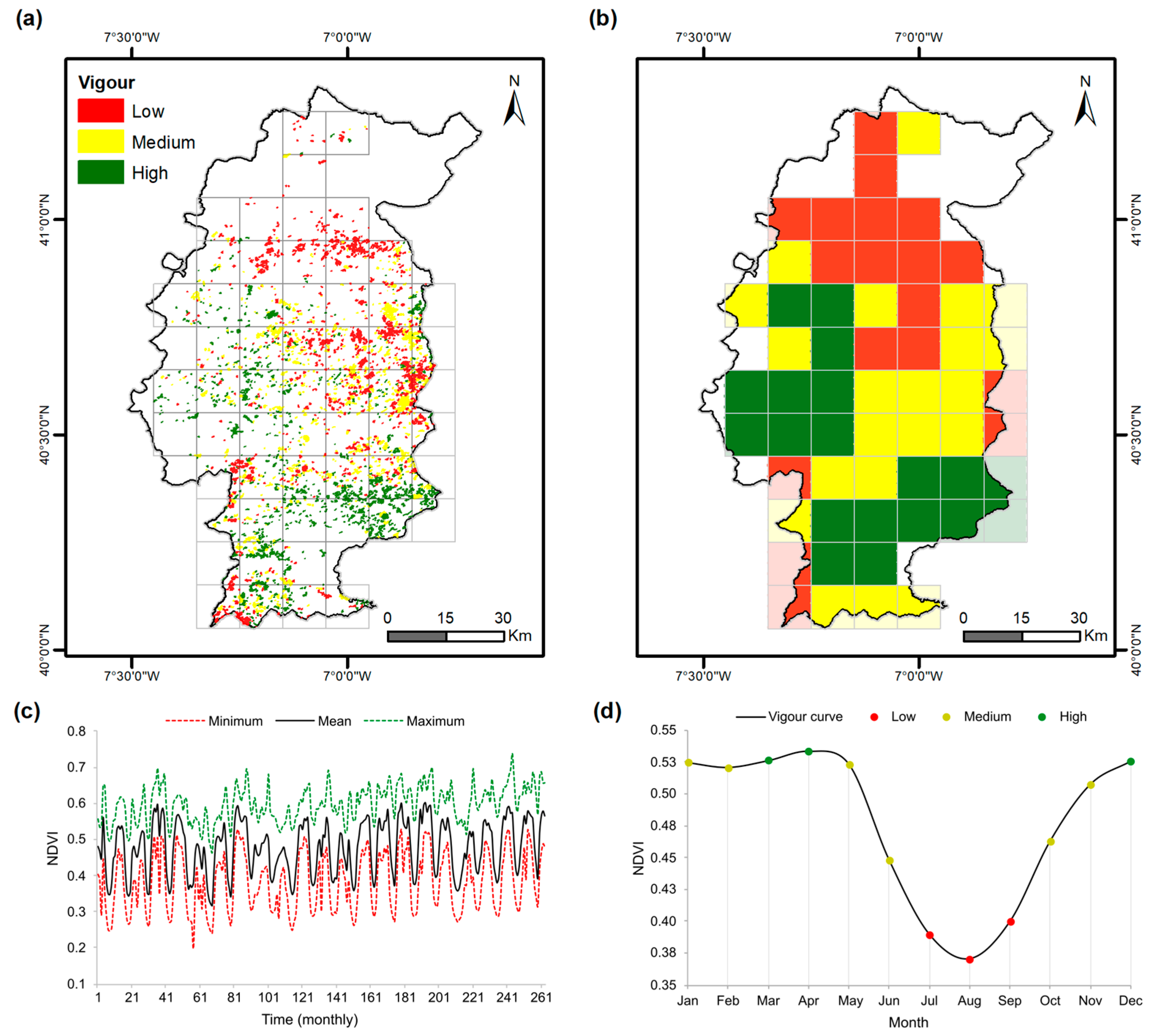
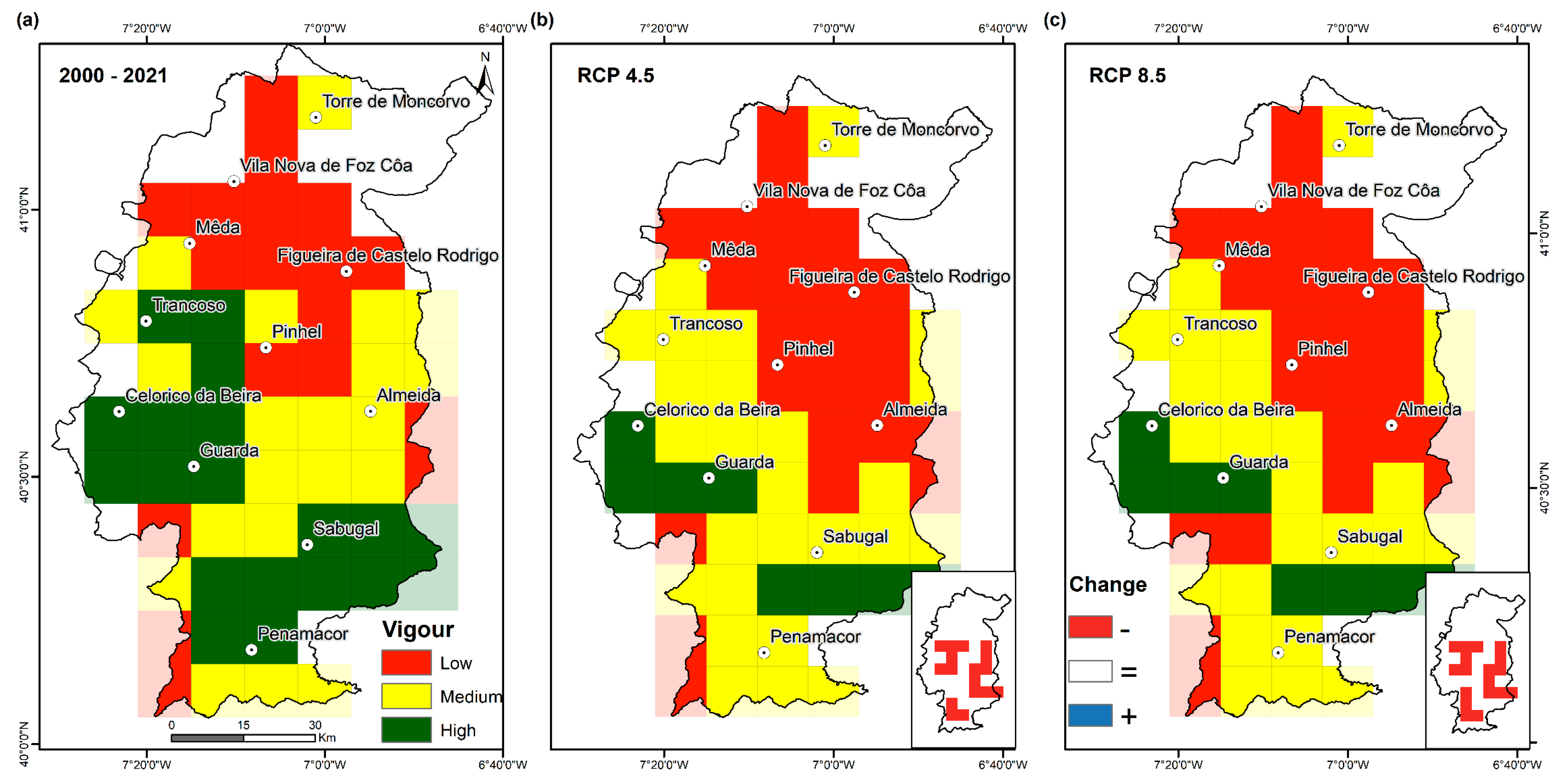
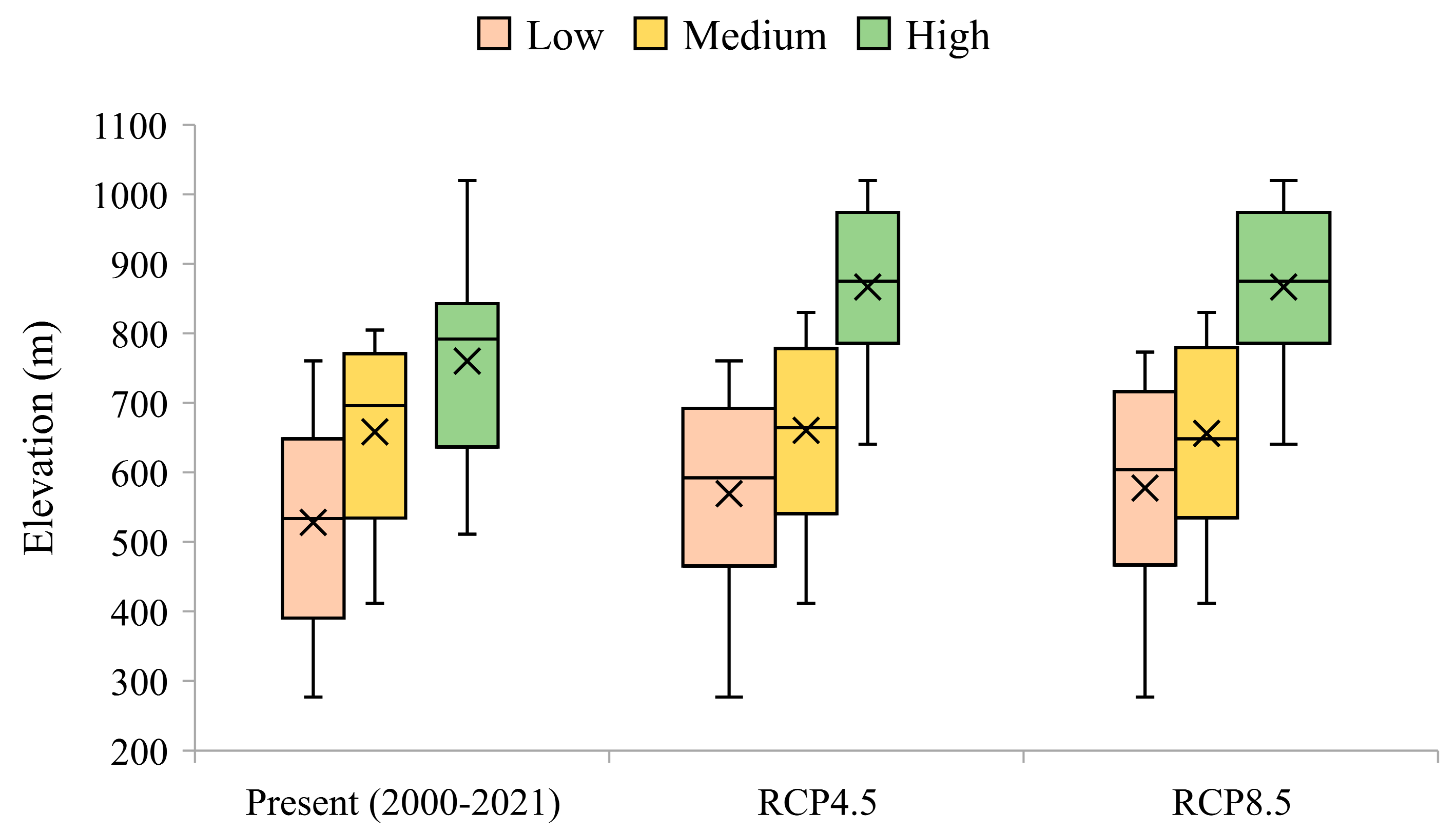
3.1. Recent-Past Grassland Vigour
3.2. Future Changes in Grassland Vigour
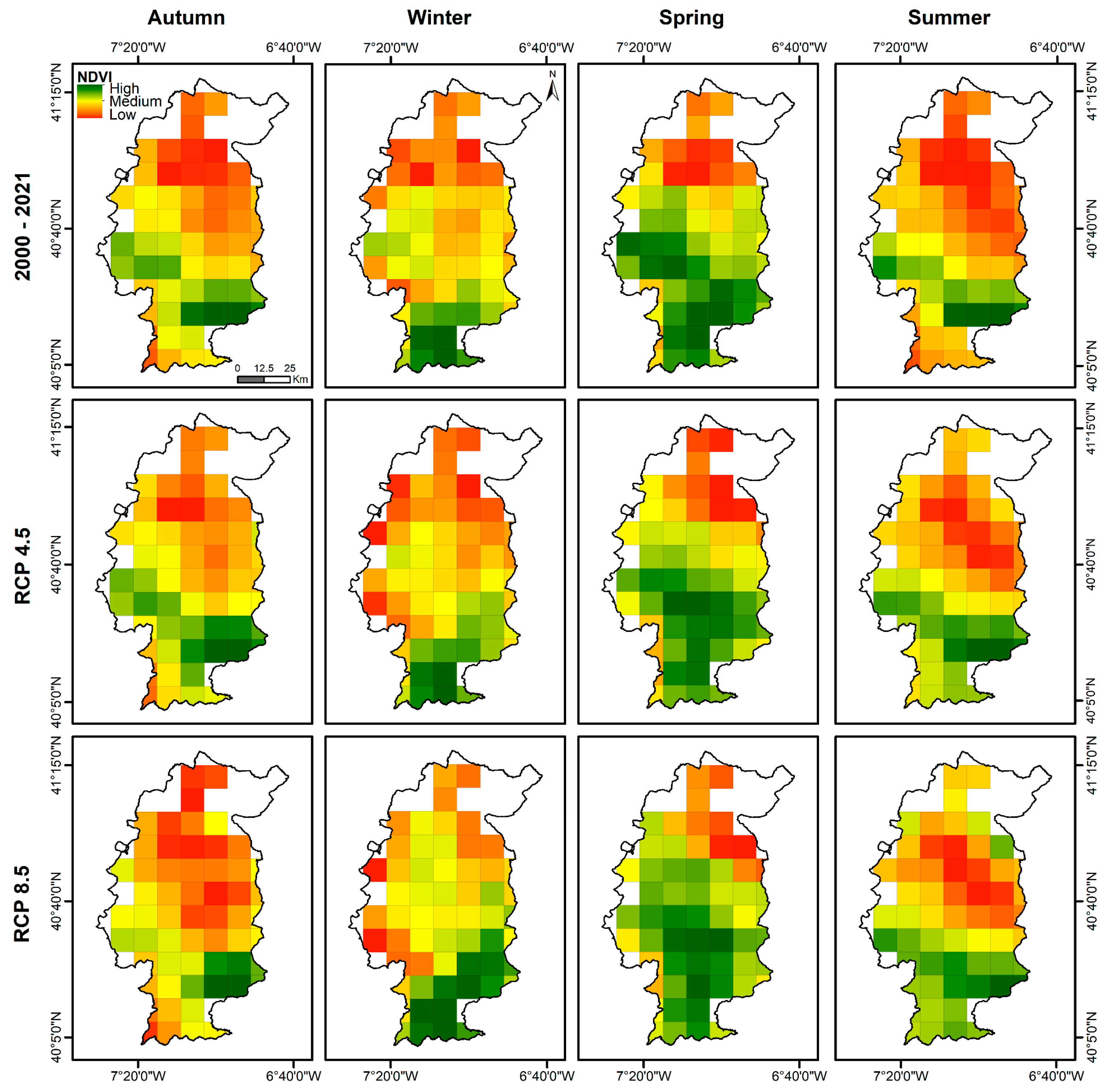
3.3. Seasonal Changes in Grassland Vigour
4. Discussion
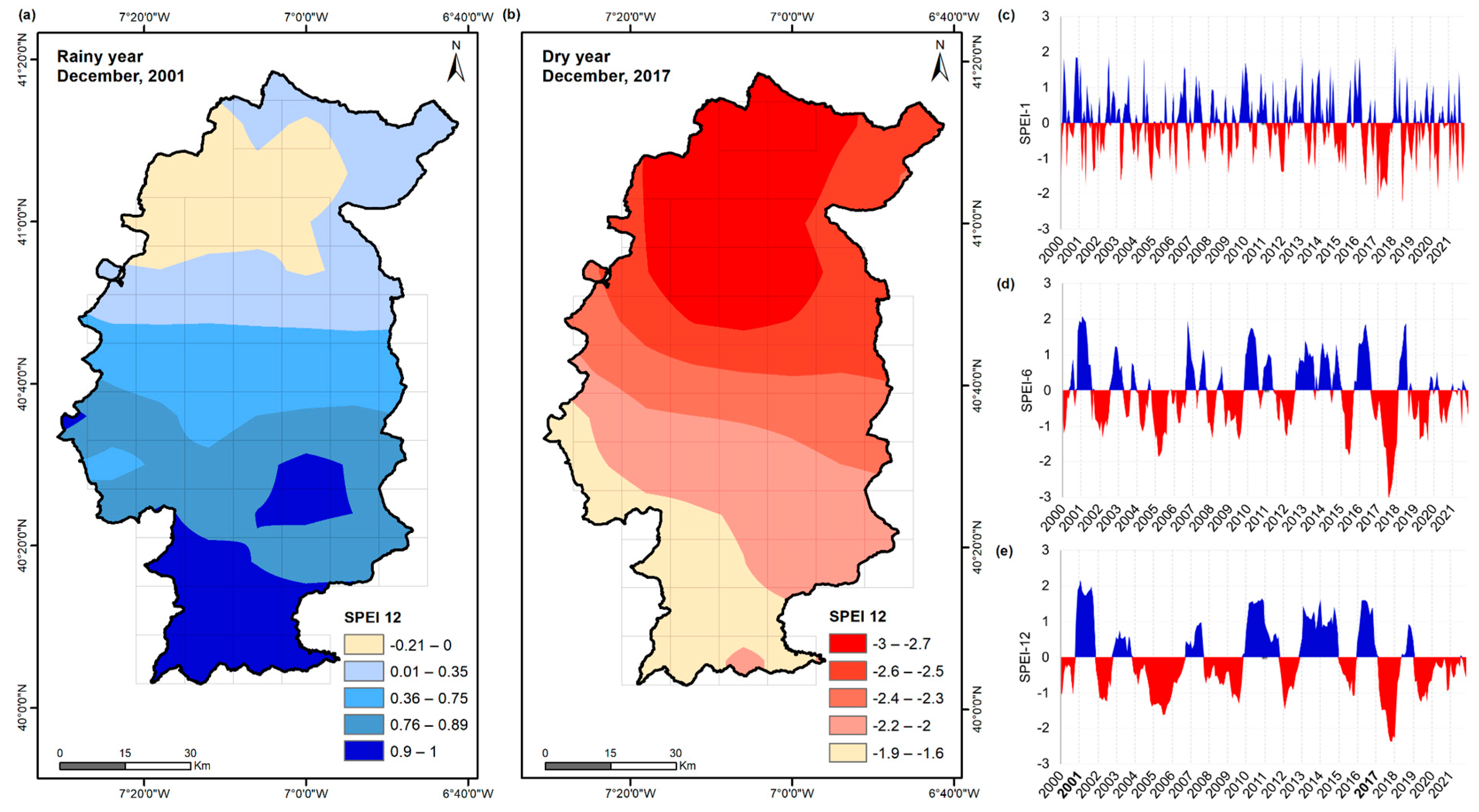
4.1. NDVI vs. SPEI
4.2. Impact of Climate Change on Grassland
4.3. Potential Adaptation Measures for Grasslands
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. World Food and Agriculture—Statistical Yearbook 2022; Food and Agriculture Organization of the United Nations: Rome, Italy, 2022; ISBN 978-92-5-136930-2. [Google Scholar]
- Allen, V.G.; Batello, C.; Berretta, E.J.; Hodgson, J.; Kothmann, M.; Li, X.; McIvor, J.; Milne, J.; Morris, C.; Peeters, A.; et al. An International Terminology for Grazing Lands and Grazing Animals. Grass Forage Sci. 2011, 66, 2–28. [Google Scholar] [CrossRef]
- Peeters, A.; Beaufoy, G.; Canals, R.; Vliegher, A.; Huyghe, C.; Isselstein, J.; Jones, G.; Kessler, W.; Kirilov, A.; Mosquera-Losada, M.R.; et al. Grassland Term Definitions and Classifications Adapted to the Diversity of European Grassland-Based Systems. Grassl. Sci. Eur. 2014, 19, 743–752. [Google Scholar]
- Ali, I.; Cawkwell, F.; Dwyer, E.; Barrett, B.; Green, S. Satellite Remote Sensing of Grasslands: From Observation to Management. J. Plant Ecol. 2016, 9, 649–671. [Google Scholar] [CrossRef]
- Porqueddu, C.; Ates, S.; Louhaichi, M.; Kyriazopoulos, A.P.; Moreno, G.; del Pozo, A.; Ovalle, C.; Ewing, M.A.; Nichols, P.G.H. Grasslands in ‘Old World’ and ‘New World’ Mediterranean-Climate Zones: Past Trends, Current Status and Future Research Priorities. Grass Forage Sci. 2016, 71, 1–35. [Google Scholar] [CrossRef]
- Serrano, J.; Shahidian, S.; Paixão, L.; Marques da Silva, J.; Morais, T.; Teixeira, R.; Domingos, T. Spatiotemporal Patterns of Pasture Quality Based on NDVI Time-Series in Mediterranean Montado Ecosystem. Remote Sens. 2021, 13, 3820. [Google Scholar] [CrossRef]
- Eurostat Share of Main Land Types in Utilised Agricultural Area (UAA) by NUTS 2 Regions. Available online: https://ec.europa.eu/eurostat/databrowser/view/tai05/default/table?lang=en (accessed on 11 January 2023).
- Mittermeier, R.; Gil, P.; Hoffmann, M.; Pilgrim, J.; Brooks, T.; Mittermeier, C.; Lamoreux, J.; Fonseca, G. Hotspots Revisited. Earth’s Biologically Richest and Most Endangered Terrestrial Ecoregions; Cemex: San Pedro Garza García, Mexico, 2004; Volume 392, ISBN 978-968-6397-77-2. [Google Scholar]
- INE. Recenseamento Agrícola—Análise Dos Principais Resultados—2019, 1st ed.; Instituto Nacional de Estatística (INE): Lisboa, Portugal, 2021; ISBN 978-989-25-0562-6. [Google Scholar]
- Bugalho, M.; Abreu, J. The Multifunctional Role of Grasslands. In Sustainable Mediterranean Grasslands and Their Multi-Functions; Porqueddu, C., Tavares de Sousa, M.M., Eds.; CIHEAM/FAO/ENMP/SPPF: Zaragoza, Spain, 2008; pp. 25–30. [Google Scholar]
- O’Mara, F.P. The Role of Grasslands in Food Security and Climate Change. Ann. Bot. 2012, 110, 1263–1270. [Google Scholar] [CrossRef]
- Taube, F.; Gierus, M.; Hermann, A.; Loges, R.; Schönbach, P. Grassland and Globalization—Challenges for North-West European Grass and Forage Research. Grass Forage Sci. 2014, 69, 2–16. [Google Scholar] [CrossRef]
- Dass, P.; Houlton, B.Z.; Wang, Y.; Warlind, D. Grasslands May Be More Reliable Carbon Sinks than Forests in California. Environ. Res. Lett. 2018, 13, 074027. [Google Scholar] [CrossRef]
- Bengtsson, J.; Bullock, J.M.; Egoh, B.; Everson, C.; Everson, T.; O’Connor, T.; O’Farrell, P.J.; Smith, H.G.; Lindborg, R. Grasslands—More Important for Ecosystem Services than You Might Think. Ecosphere 2019, 10, e02582. [Google Scholar] [CrossRef]
- Isselstein, J.; Kayser, M. Functions of Grassland and Their Potential in Delivering Ecosystem Services. Grassl. Sci. Eur. 2014, 19, 199–214. [Google Scholar]
- Soussana, J.-F.; Klumpp, K.; Ehrhardt, F. The Role of Grassland in Mitigating Climate Change; Institute of Biological, Environmental and Rural Sciences (IBERS): Aberystwyth, UK, 2014; Volume 19, 919p. [Google Scholar]
- Collins, M.; Knutti, R.; Arblaster, J.; Dufresne, J.-L.; Fichefet, T.; Friedlingstein, P.; Gao, X.; Gutowski, W.J.; Johns, T.; Krinner, G.; et al. Long-Term Climate Change: Projections, Commitments and Irreversibility. In Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Qin, D., Plattner, G.K., Tignor, M., Allen, S.K., Boschung, J., Naules, A., Xia, Y., Bex, V., Midgley, P.M., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2013; pp. 1029–1136. [Google Scholar]
- Dumont, B.; Andueza, D.; Niderkorn, V.; Lüscher, A.; Porqueddu, C.; Picon-Cochard, C. A Meta-Analysis of Climate Change Effects on Forage Quality in Grasslands: Specificities of Mountain and Mediterranean Areas. Grass Forage Sci. 2015, 70, 239–254. [Google Scholar] [CrossRef]
- Fraga, H.; Molitor, D.; Leolini, L.; Santos, J.A. What Is the Impact of Heatwaves on European Viticulture? A Modelling Assessment. Appl. Sci. 2020, 10, 3030. [Google Scholar] [CrossRef]
- Reichstein, M.; Ciais, P.; Papale, D.; Valentini, R.; Running, S.; Viovy, N.; Cramer, W.; Granier, A.; Ogée, J.; Allard, V.; et al. Reduction of Ecosystem Productivity and Respiration during the European Summer 2003 Climate Anomaly: A Joint Flux Tower, Remote Sensing and Modelling Analysis. Glob. Change Biol. 2007, 13, 634–651. [Google Scholar] [CrossRef]
- Hott, M.C.; Carvalho, L.M.T.; Antunes, M.A.H.; Resende, J.C.; Rocha, W.S.D. Analysis of Grassland Degradation in Zona Da Mata, MG, Brazil, Based on NDVI Time Series Data with the Integration of Phenological Metrics. Remote Sens. 2019, 11, 2956. [Google Scholar] [CrossRef]
- Liu, S.; Zhang, Y.; Cheng, F.; Hou, X.; Zhao, S. Response of Grassland Degradation to Drought at Different Time-Scales in Qinghai Province: Spatio-Temporal Characteristics, Correlation, and Implications. Remote Sens. 2017, 9, 1329. [Google Scholar] [CrossRef]
- Xu, B.; Yang, X.C.; Tao, W.G.; Miao, J.M.; Yang, Z.; Liu, H.Q.; Jin, Y.X.; Zhu, X.H.; Qin, Z.H.; Lv, H.Y.; et al. MODIS-Based Remote-Sensing Monitoring of the Spatiotemporal Patterns of China’s Grassland Vegetation Growth. Int. J. Remote Sens. 2013, 34, 3867–3878. [Google Scholar] [CrossRef]
- Wang, R.; Gamon, J.A.; Montgomery, R.A.; Townsend, P.A.; Zygielbaum, A.I.; Bitan, K.; Tilman, D.; Cavender-Bares, J. Seasonal Variation in the NDVI–Species Richness Relationship in a Prairie Grassland Experiment (Cedar Creek). Remote Sens. 2016, 8, 128. [Google Scholar] [CrossRef]
- Phillips, L.B.; Hansen, A.J.; Flather, C.H. Evaluating the Species Energy Relationship with the Newest Measures of Ecosystem Energy: NDVI versus MODIS Primary Production. Remote Sens. Environ. 2008, 112, 4381–4392. [Google Scholar] [CrossRef]
- Fan, L.; Gao, Y.; Brück, H.; Bernhofer, C. Investigating the Relationship between NDVI and LAI in Semi-Arid Grassland in Inner Mongolia Using in-Situ Measurements. Theor. Appl. Climatol. 2009, 95, 151–156. [Google Scholar] [CrossRef]
- Anderson, M.C.; Hain, C.; Wardlow, B.; Pimstein, A.; Mecikalski, J.R.; Kustas, W.P. Evaluation of Drought Indices Based on Thermal Remote Sensing of Evapotranspiration over the Continental United States. J. Clim. 2011, 24, 2025–2044. [Google Scholar] [CrossRef]
- Starks, P.J.; Steiner, J.L.; Neel, J.P.S.; Turner, K.E.; Northup, B.K.; Gowda, P.H.; Brown, M.A. Assessment of the Standardized Precipitation and Evaporation Index (SPEI) as a Potential Management Tool for Grasslands. Agronomy 2019, 9, 235. [Google Scholar] [CrossRef]
- Ausseil, A.-G.; Weerden, T.; Beare, M.; Teixeira, E.; Baisden, T.; Lieffering, M.; Guo, J.; Keller, E.; Law, R.; Noble, A. Climate Change Impacts on Land Use Suitability; Manaaki Whenua Landcare Research: Lincoln, New Zealand, 2019. [Google Scholar]
- Luo, N.; Mao, D.; Wen, B.; Liu, X. Climate Change Affected Vegetation Dynamics in the Northern Xinjiang of China: Evaluation by SPEI and NDVI. Land 2020, 9, 90. [Google Scholar] [CrossRef]
- Cang, F.A.; Wilson, A.A.; Wiens, J.J. Climate Change Is Projected to Outpace Rates of Niche Change in Grasses. Biol. Lett. 2016, 12, 20160368. [Google Scholar] [CrossRef]
- Morris, C.D.; Kirkman, K.P.; Zacharias, P.J.K. Will the Grass Be Greener on the Other Side of Climate Change? S. Afr. J. Sci. 2022, 118, 1–4. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, B. The Responses of Natural Vegetation Dynamics to Drought during the Growing Season across China. J. Hydrol. 2019, 574, 706–714. [Google Scholar] [CrossRef]
- USGS EarthExplorer. Available online: https://earthexplorer.usgs.gov/ (accessed on 17 January 2023).
- Breiman, L. Random Forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- SNIG. Carta de Uso e Ocupação do Solo. Available online: https://snig.dgterritorio.gov.pt/rndg/srv/por/catalog.search#/home (accessed on 11 January 2023).
- Coelho, P. Pastagens de Altitude—Caracterização e Melhoramento—Pastagens Semeadas Na Região Da Guarda; ISA: Lisboa, Portugal, 2016; 95p. [Google Scholar]
- Rouse, W.; Haas, R.H.; Schell, J.A.; Deering, D.W. Monitoring vegetation systems in the great plains with erts. NASA Spec. Publ. 1974, 351, 309–317. [Google Scholar]
- Vicente-Serrano, S.M.; Beguería, S.; López-Moreno, J.I. A Multiscalar Drought Index Sensitive to Global Warming: The Standardized Precipitation Evapotranspiration Index. J. Clim. 2010, 23, 1696–1718. [Google Scholar] [CrossRef]
- Gutiérrez, J.M.; Herrera, S.; Cardoso, R.M.; Matos Soares, P.; Espírito-Santo, F.; Viterbo, P. Iberia01: Daily Gridded (0.1° Resolution) Dataset of Precipitation and Temperatures over the Iberian Peninsula 2019. Available online: https://essd.copernicus.org/articles/11/1947/2019/ (accessed on 12 December 2022).
- Haylock, M.R.; Hofstra, N.; Klein Tank, A.M.G.; Klok, E.J.; Jones, P.D.; New, M. A European Daily High-Resolution Gridded Data Set of Surface Temperature and Precipitation for 1950–2006. J. Geophys. Res. Atmos. 2008, 113, D20119. [Google Scholar] [CrossRef]
- Cornes, R.C.; van der Schrier, G.; van den Besselaar, E.J.M.; Jones, P.D. An Ensemble Version of the E-OBS Temperature and Precipitation Data Sets. J. Geophys. Res. Atmos. 2018, 123, 9391–9409. [Google Scholar] [CrossRef]
- Herrera, S.; Cardoso, R.M.; Soares, P.M.; Espírito-Santo, F.; Viterbo, P.; Gutiérrez, J.M. Iberia01: A New Gridded Dataset of Daily Precipitation and Temperatures over Iberia. Earth Syst. Sci. Data 2019, 11, 1947–1956. [Google Scholar] [CrossRef]
- Ramirez Villejas, J.; Jarvis, A. Downscaling Global Circulation Model Outputs: The Delta Method Decision and Policy Analysis Working Paper No. 1; International Center for Tropical Agriculture (CIAT): Cali, Colombia, 2010; 18p. [Google Scholar]
- Hargreaves, G.; Samani, Z. Reference Crop Evapotranspiration From Temperature. Appl. Eng. Agric. 1985, 1, 96–99. [Google Scholar] [CrossRef]
- Allan, R.; Pereira, L.; Smith, M. Crop Evapotranspiration-Guidelines for Computing Crop Water Requirements; FAO Irrigation and Drainage Paper 56; FAO: Rome, Italy, 1998; Volume 56. [Google Scholar]
- Faye, C.; Manuela, G.; Wood, S. Use of the Standardized Precipitation and Evapotranspiration Index (SPEI) from 1950 to 2018 to Determine Drought Trends in the Senegalese Territory Citation Climate Change. Clim. Change 2019, 5, 327–341. [Google Scholar]
- Jacob, D.; Petersen, J.; Eggert, B.; Alias, A.; Christensen, O.B.; Bouwer, L.M.; Braun, A.; Colette, A.; Déqué, M.; Georgievski, G.; et al. EURO-CORDEX: New High-Resolution Climate Change Projections for European Impact Research. Reg. Environ. Change 2014, 14, 563–578. [Google Scholar] [CrossRef]
- Navarro-Racines, C.; Tarapues, J.; Thornton, P.; Jarvis, A.; Ramirez-Villegas, J. High-Resolution and Bias-Corrected CMIP5 Projections for Climate Change Impact Assessments. Sci. Data 2020, 7, 7. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, E.; Osborne, T.M.; Ho, C.K.; Challinor, A.J. Calibration and Bias Correction of Climate Projections for Crop Modelling: An Idealised Case Study over Europe. Agric. For. Meteorol. 2013, 170, 19–31. [Google Scholar] [CrossRef]
- Potop, V.; Boroneanţ, C.; Možný, M.; Štěpánek, P.; Skalák, P. Observed Spatiotemporal Characteristics of Drought on Various Time Scales over the Czech Republic. Theor. Appl. Climatol. 2014, 115, 563–581. [Google Scholar] [CrossRef]
- Yang, C.; Menz, C.; De Abreu Jaffe, M.S.; Costafreda-Aumedes, S.; Moriondo, M.; Leolini, L.; Torres-Matallana, A.; Molitor, D.; Junk, J.; Fraga, H.; et al. Projections of Climate Change Impacts on Flowering-Veraison Water Deficits for Riesling and Müller-Thurgau in Germany. Remote Sens. 2022, 14, 1519. [Google Scholar] [CrossRef]
- Guyon, I.; Gunn, S.; Nikravesh, M.; Zadeh, L.A. Feature Extraction: Foundations and Applications; Studies in Fuzziness and Soft Computing; Springer: Berlin/Heidelberg, Germany, 2008; ISBN 978-3-540-35488-8. [Google Scholar]
- Gobiet, A.; Kotlarski, S.; Beniston, M.; Heinrich, G.; Rajczak, J.; Stoffel, M. 21st Century Climate Change in the European Alps—A Review. Sci. Total Environ. 2014, 493, 1138–1151. [Google Scholar] [CrossRef]
- Carlson, B.Z.; Corona, M.C.; Dentant, C.; Bonet, R.; Thuiller, W.; Choler, P. Observed Long-Term Greening of Alpine Vegetation—A Case Study in the French Alps. Environ. Res. Lett. 2017, 12, 114006. [Google Scholar] [CrossRef]
- Jones, M.B.; Donnelly, A. Carbon Sequestration in Temperate Grassland Ecosystems and the Influence of Management, Climate and Elevated CO2. New Phytol. 2004, 164, 423–439. [Google Scholar] [CrossRef]
- Obermeier, W.A.; Lehnert, L.W.; Kammann, C.I.; Müller, C.; Grünhage, L.; Luterbacher, J.; Erbs, M.; Moser, G.; Seibert, R.; Yuan, N.; et al. Reduced CO2 Fertilization Effect in Temperate C3 Grasslands under More Extreme Weather Conditions. Nat. Clim. Change 2017, 7, 137–141. [Google Scholar] [CrossRef]
- Dibari, C.; Costafreda-Aumedes, S.; Argenti, G.; Bindi, M.; Carotenuto, F.; Moriondo, M.; Padovan, G.; Pardini, A.; Staglianò, N.; Vagnoli, C.; et al. Expected Changes to Alpine Pastures in Extent and Composition under Future Climate Conditions. Agronomy 2020, 10, 926. [Google Scholar] [CrossRef]
- Bloor, J.M.G.; Pichon, P.; Falcimagne, R.; Leadley, P.; Soussana, J.-F. Effects of Warming, Summer Drought, and CO2 Enrichment on Aboveground Biomass Production, Flowering Phenology, and Community Structure in an Upland Grassland Ecosystem. Ecosystems 2010, 13, 888–900. [Google Scholar] [CrossRef]
- Kwon, H.-Y.; Nkonya, E.; Johnson, T.; Graw, V.; Kato, E.; Kihiu, E. Global Estimates of the Impacts of Grassland Degradation on Livestock Productivity from 2001 to 2011. In Economics of Land Degradation and Improvement—A Global Assessment for Sustainable Development; Nkonya, E., Mirzabaev, A., von Braun, J., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 197–214. ISBN 978-3-319-19168-3. [Google Scholar]
- Liu, Q.; Fu, Y.H.; Zhu, Z.; Liu, Y.; Liu, Z.; Huang, M.; Janssens, I.A.; Piao, S. Delayed Autumn Phenology in the Northern Hemisphere Is Related to Change in Both Climate and Spring Phenology. Glob. Change Biol. 2016, 22, 3702–3711. [Google Scholar] [CrossRef]
- Casale, F.; Bocchiola, D. Climate Change Effects upon Pasture in the Alps: The Case of Valtellina Valley, Italy. Climate 2022, 10, 173. [Google Scholar] [CrossRef]
- Craine, J.M.; Ocheltree, T.W.; Nippert, J.B.; Towne, E.G.; Skibbe, A.M.; Kembel, S.W.; Fargione, J.E. Global Diversity of Drought Tolerance and Grassland Climate-Change Resilience. Nat. Clim. Change 2013, 3, 63–67. [Google Scholar] [CrossRef]
- Ji, L.; Peters, A.J. A Spatial Regression Procedure for Evaluating the Relationship between AVHRR-NDVI and Climate in the Northern Great Plains. Int. J. Remote Sens. 2004, 25, 297–311. [Google Scholar] [CrossRef]
- Peterson, P.R.; Sheaffer, C.C.; Hall, M.H. Drought Effects on Perennial Forage Legume Yield and Quality. Agron. J. 1992, 84, 774–779. [Google Scholar] [CrossRef]
- Sternberg, M.; Gutman, M.; Perevolotsky, A.; Kigel, J. Effects of Grazing on Soil Seed Bank Dynamics: An Approach with Functional Groups. J. Veg. Sci. 2003, 14, 375–386. [Google Scholar] [CrossRef]
- Kreyling, J.; Thiel, D.; Simmnacher, K.; Willner, E.; Jentsch, A.; Beierkuhnlein, C. Geographic Origin and Past Climatic Experience Influence the Response to Late Spring Frost in Four Common Grass Species in Central Europe. Ecography 2012, 35, 268–275. [Google Scholar] [CrossRef]
- Volaire, F.; Norton, M.R.; Lelièvre, F. Summer Drought Survival Strategies and Sustainability of Perennial Temperate Forage Grasses in Mediterranean Areas. Crop Sci. 2009, 49, 2386–2392. [Google Scholar] [CrossRef]
- Long, R.L.; Gorecki, M.J.; Renton, M.; Scott, J.K.; Colville, L.; Goggin, D.E.; Commander, L.E.; Westcott, D.A.; Cherry, H.; Finch-Savage, W.E. The Ecophysiology of Seed Persistence: A Mechanistic View of the Journey to Germination or Demise. Biol. Rev. 2015, 90, 31–59. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.; Shang, Z.; Gao, J.; Boone, R.B. Enhancing Sustainability of Grassland Ecosystems through Ecological Restoration and Grazing Management in an Era of Climate Change on Qinghai-Tibetan Plateau. Agric. Ecosyst. Environ. 2020, 287, 106684. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Season | RCP4.5delta | RCP8.5delta |
|---|---|---|
| Precipitation (%) | ||
| Autumn | −10 | −20 |
| Winter | 10 | 10 |
| Spring | −10 | −20 |
| Summer | −20 | −30 |
| Temperature (°C) | ||
| Autumn | 2.75 | 4.75 |
| Winter | 1.25 | 3.25 |
| Spring | 1.75 | 2.75 |
| Summer | 2.75 | 5.25 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stolarski, O.; Santos, J.A.; Fonseca, A.; Yang, C.; Trindade, H.; Fraga, H. Climate Change Impacts on Grassland Vigour in Northern Portugal. Land 2023, 12, 1914. https://doi.org/10.3390/land12101914
Stolarski O, Santos JA, Fonseca A, Yang C, Trindade H, Fraga H. Climate Change Impacts on Grassland Vigour in Northern Portugal. Land. 2023; 12(10):1914. https://doi.org/10.3390/land12101914
Chicago/Turabian StyleStolarski, Oiliam, João A. Santos, André Fonseca, Chenyao Yang, Henrique Trindade, and Helder Fraga. 2023. "Climate Change Impacts on Grassland Vigour in Northern Portugal" Land 12, no. 10: 1914. https://doi.org/10.3390/land12101914
APA StyleStolarski, O., Santos, J. A., Fonseca, A., Yang, C., Trindade, H., & Fraga, H. (2023). Climate Change Impacts on Grassland Vigour in Northern Portugal. Land, 12(10), 1914. https://doi.org/10.3390/land12101914