
_Sun.png)


Homogenization of Urban Forests across the Subtropical Zones of China

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Data Collection and Processing
2.2.1. Tree Community Data of Urban Forests
2.2.2. Data for the Community Traits of Rural Forests
2.2.3. Socioeconomic Attributes
2.2.4. Data for Climate and Calculations
2.2.5. Data for Distance between Cities
2.3. Calculations
3. Results
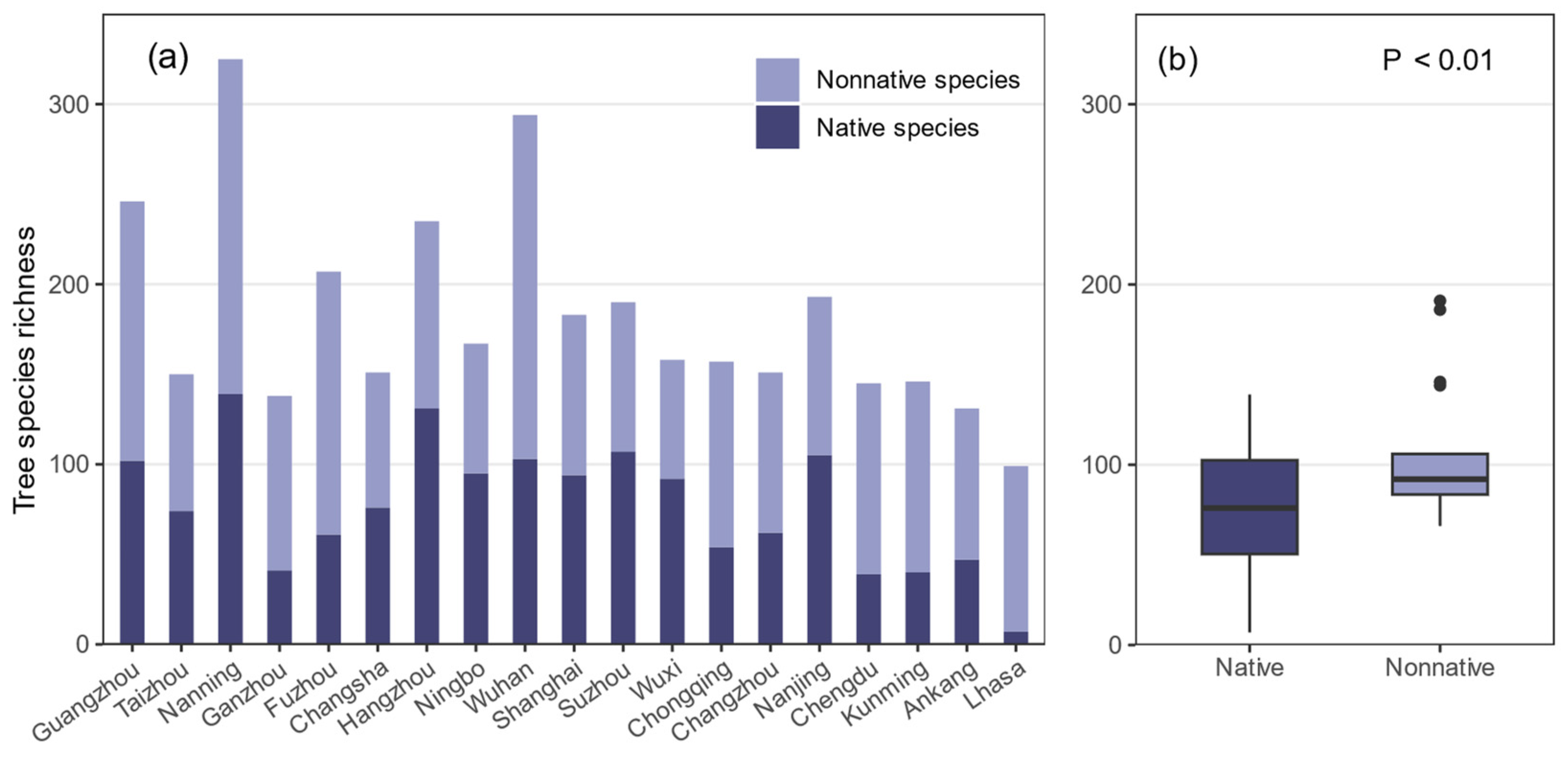
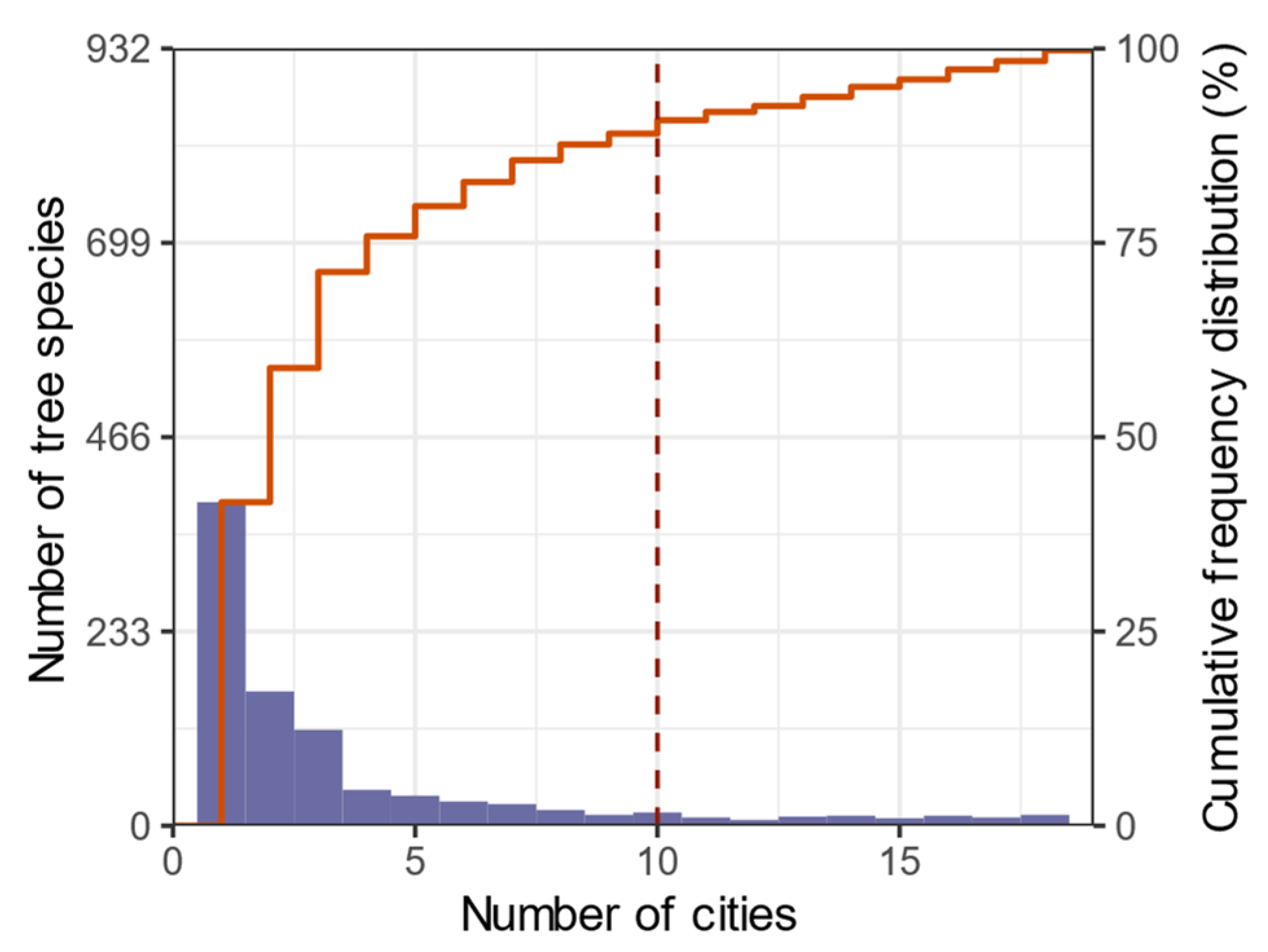
3.1. Tree Species Richness in Urban Forests
3.2. Variations in Tree Diversity in Urban Forests and Rural Forests along the Precipitation Gradient
3.3. Similarities in Tree Species Composition between Urban Forests or Rural Forests along the Precipitation Gradient
3.4. Relationships between Tree Diversity of Urban Forests and Socioeconomic Factors
4. Discussion
4.1. The Tree Species Composition Homogenized along the Precipitation Gradient in Subtropical China
4.2. The Biotic Homogenization of Urban Forests in Humid Areas Due to Removal More than the Introduction
4.3. The Tree Diversity of Urban Forests Tends to Converge along the Precipitation Gradient
4.4. Implications of Biotic Homogenization in Urban Forests to Sustainable Cities
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chang, J.; Qu, Z.; Xu, R.; Pan, K.; Xu, B.; Min, Y.; Ren, Y.; Yang, G.; Ge, Y. Assessing the ecosystem services provided by urban green spaces along urban center-edge gradients. Sci. Rep. 2017, 7, 11226. [Google Scholar] [CrossRef] [Green Version]
- Portner, H.-O.; Scholes, R.J.; Arneth, A.; Barnes, D.K.A.; Burrows, M.T.; Diamond, S.E.; Duarte, C.M.; Kiessling, W.; Leadley, P.; Managi, S.; et al. Overcoming the coupled climate and biodiversity crises and their societal impacts. Science 2023, 380, eabl4881. [Google Scholar] [CrossRef]
- Endreny, T.A. Strategically growing the urban forest will improve our world. Nat. Commun. 2018, 9, 1160. [Google Scholar] [CrossRef] [Green Version]
- Le Provost, G.; Schenk, N.V.; Penone, C.; Thiele, J.; Westphal, C.; Allan, E.; Ayasse, M.; Blüthgen, N.; Boeddinghaus, R.S.; Boesing, A.L.; et al. The supply of multiple ecosystem services requires biodiversity across spatial scales. Nat. Ecol. Evol. 2023, 7, 236–249. [Google Scholar] [CrossRef]
- Simkin, R.D.; Seto, K.C.; McDonald, R.I.; Jetz, W. Biodiversity impacts and conservation implications of urban land expansion projected to 2050. Proc. Natl. Acad. Sci. USA 2022, 119, e2117297119. [Google Scholar] [CrossRef]
- Delgado-Baquerizo, M.; Eldridge, D.J.; Liu, Y.-R.; Sokoya, B.; Wang, J.-T.; Hu, H.-W.; He, J.-Z.; Bastida, F.; Moreno, J.L.; Bamigboye, A.R.; et al. Global homogenization of the structure and function in the soil microbiome of urban greenspaces. Sci. Adv. 2021, 7, eabg5809. [Google Scholar] [CrossRef]
- Gao, Z.; Pan, Y.; Van Bodegom, P.M.; Cieraad, E.; Xing, D.; Yang, Y.; Xia, T.; Luo, X.; Song, K.; Da, L.; et al. Beta diversity of urban spontaneous plants and its drivers in 9 major cities of Yunnan province, China. Landsc. Urban Plan. 2023, 234, 104741. [Google Scholar] [CrossRef]
- Wang, X.; Svenning, J.-C.; Liu, J.; Zhao, Z.; Zhang, Z.; Feng, G.; Si, X.; Zhang, J. Regional effects of plant diversity and biotic homogenization in urban greenspace—The case of university campuses across China. Urban For. Urban Green. 2021, 62, 127170. [Google Scholar] [CrossRef]
- McKinney, M.L. Urbanization as a major cause of biotic homogenization. Biol. Conserv. 2006, 127, 247–260. [Google Scholar] [CrossRef]
- Qian, S.; Qi, M.; Huang, L.; Zhao, L.; Lin, D.; Yang, Y. Biotic homogenization of China’s urban greening: A meta-analysis on woody species. Urban For. Urban Green. 2016, 18, 25–33. [Google Scholar] [CrossRef]
- Ouyang, Y.; Chen, Y.; Yang, G.; Ren, Y.; Yu, M.; Chang, J.; Ge, Y. Homogenization of trees in urban green spaces along the moisture gradient in China. Urban For. Urban Green. 2023, 83, 127892. [Google Scholar] [CrossRef]
- Groffman, P.M.; Avolio, M.; Cavender-Bares, J.; Bettez, N.D.; Grove, J.M.; Hall, S.J.; Hobbie, S.E.; Larson, K.L.; Lerman, S.B.; Locke, D.H.; et al. Ecological homogenization of residential macrosystems. Nat. Ecol. Evol. 2017, 1, 0191. [Google Scholar] [CrossRef] [Green Version]
- Harrison, S.; Spasojevic, M.J.; Li, D. Climate and plant community diversity in space and time. Proc. Natl. Acad. Sci. USA 2020, 117, 4464–4470. [Google Scholar] [CrossRef]
- Chen, Y.; Ge, Y.; Wu, Z.; Ouyang, Y.; Yang, G.; Du, Y.; Pan, K.; Chang, J. Homogenization of tree species diversity in urban green spaces along a temperature gradient in eastern China. Urban For. Urban Green. 2021, 66, 127388. [Google Scholar] [CrossRef]
- Capinha, C.; Essl, F.; Seebens, H.; Moser, D.; Pereira, H.M. The dispersal of alien species redefines biogeography in the Anthropocene. Science 2015, 348, 1248–1251. [Google Scholar] [CrossRef]
- Ossola, A.; Hopton, M.E. Climate differentiates forest structure across a residential macrosystem. Sci. Total Environ. 2018, 639, 1164–1174. [Google Scholar] [CrossRef]
- Qian, H.; Ricklefs, R.E. A latitudinal gradient in large-scale beta diversity for vascular plants in North America. Ecol. Lett. 2007, 10, 737–744. [Google Scholar] [CrossRef]
- Anguelovski, I.; Connolly, J.J.T.; Cole, H.; Garcia-Lamarca, M.; Triguero-Mas, M.; Baró, F.; Martin, N.; Conesa, D.; Shokry, G.; Del Pulgar, C.P.; et al. Green gentrification in European and North American cities. Nat. Commun. 2022, 13, 3816. [Google Scholar] [CrossRef]
- Wheeler, M.M.; Neill, C.; Groffman, P.M.; Avolio, M.; Bettez, N.; Cavender-Bares, J.; Roy Chowdhury, R.; Darling, L.; Grove, J.M.; Hall, S.J.; et al. Continental-scale homogenization of residential lawn plant communities. Landsc. Urban Plan. 2017, 165, 54–63. [Google Scholar] [CrossRef]
- Hutyra, L.R.; Yoon, B.; Alberti, M. Terrestrial carbon stocks across a gradient of urbanization: A study of the Seattle, WA region. Glob. Chang. Biol. 2011, 17, 783–797. [Google Scholar] [CrossRef]
- Yang, G.; Xu, R.; Chen, Y.; Wu, Z.; Du, Y.; Liu, S.; Qu, Z.; Guo, K.; Peng, C.; Chang, J.; et al. Identifying the greenhouses by Google Earth Engine to promote the reuse of fragmented land in urban fringe. Sustain. Cities Soc. 2021, 67, 102743. [Google Scholar] [CrossRef]
- Nowak, D.J.; Walton, J.T.; Stevens, J.C.; Crane, D.E.; Hoehn, R.E. Effect of Plot and Sample Size on Timing and Precision of Urban Forest Assessments. Arboric. Urban For. 2008, 34, 386–390. [Google Scholar] [CrossRef]
- Lososová, Z.; Chytrý, M.; Tichý, L.; Danihelka, J.; Fajmon, K.; Hájek, O.; Kintrová, K.; Láníková, D.; Otýpková, Z.; Řehořek, V. Biotic homogenization of Central European urban floras depends on residence time of alien species and habitat types. Biol. Conserv. 2012, 145, 179–184. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2023; Available online: https://www.R-project.org (accessed on 29 April 2023).
- Groffman, P.M.; Cavender-Bares, J.; Bettez, N.D.; Grove, J.M.; Hall, S.J.; Heffernan, J.B.; Hobbie, S.E.; Larson, K.L.; Morse, J.L.; Neill, C.; et al. Ecological homogenization of urban USA. Front. Ecol. Environ. 2014, 12, 74–81. [Google Scholar] [CrossRef] [Green Version]
- Qian, S.; Qin, D.; Wu, X.; Hu, S.; Hu, L.; Lin, D.; Zhao, L.; Shang, K.; Song, K.; Yang, Y. Urban growth and topographical factors shape patterns of spontaneous plant community diversity in a mountainous city in southwest China. Urban For. Urban Green. 2020, 55, 126814. [Google Scholar] [CrossRef]
- Seto, K.C.; Güneralp, B.; Hutyra, L.R. Global forecasts of urban expansion to 2030 and direct impacts on biodiversity and carbon pools. Proc. Natl. Acad. Sci. USA 2012, 109, 16083–16088. [Google Scholar] [CrossRef]
- Hope, D.; Gries, C.; Zhu, W.; Fagan, W.F.; Redman, C.L.; Grimm, N.B.; Nelson, A.L.; Martin, C.; Kinzig, A. Socioeconomics drive urban plant diversity. Proc. Natl. Acad. Sci. USA 2003, 100, 8788–8792. [Google Scholar] [CrossRef]
- Stuhlmacher, M.; Georgescu, M.; Turner, B.L.; Hu, Y.N.; Goldblatt, R.; Gupta, S.; Frazier, A.E.; Kim, Y.; Balling, R.C.; Clinton, N. Are global cities homogenizing? An assessment of urban form and heat island implications. Cities 2022, 126, 103705. [Google Scholar] [CrossRef]
- Dornelas, M.; Gotelli, N.J.; McGill, B.; Shimadzu, H.; Moyes, F.; Sievers, C.; Magurran, A.E. Assemblage Time Series Reveal Biodiversity Change but Not Systematic Loss. Science 2014, 344, 296–299. [Google Scholar] [CrossRef] [Green Version]
- Dehling, D.M.; Dehling, J.M. Elevated alpha diversity in disturbed sites obscures regional decline and homogenization of amphibian taxonomic, functional and phylogenetic diversity. Sci. Rep. 2023, 13, 1710. [Google Scholar] [CrossRef]
- Dembicz, I.; Dengler, J.; Steinbauer, M.J.; Matthews, T.J.; Bartha, S.; Burrascano, S.; Chiarucci, A.; Filibeck, G.; Gillet, F.; Janišová, M.; et al. Fine-grain beta diversity of Palaearctic grassland vegetation. J. Veg. Sci. 2021, 32, e13045. [Google Scholar] [CrossRef]
- Fan, K.; Chu, H.; Eldridge, D.J.; Gaitan, J.J.; Liu, Y.-R.; Sokoya, B.; Wang, J.-T.; Hu, H.-W.; He, J.-Z.; Sun, W.; et al. Soil biodiversity supports the delivery of multiple ecosystem functions in urban greenspaces. Nat. Ecol. Evol. 2023, 7, 113–126. [Google Scholar] [CrossRef]
- Hosokawa, S.; Momota, K.; Chariton, A.A.; Naito, R.; Nakamura, Y. The use of diversity indices for local assessment of marine sediment quality. Sci. Rep. 2021, 11, 14991. [Google Scholar] [CrossRef] [PubMed]
- Strong, W.L. Biased richness and evenness relationships within Shannon–Wiener index values. Ecol. Indic. 2016, 67, 703–713. [Google Scholar] [CrossRef]
- Lambert, M.R.; Donihue, C.M. Urban biodiversity management using evolutionary tools. Nat. Ecol. Evol. 2020, 4, 903–910. [Google Scholar] [CrossRef] [PubMed]
- Leong, M.; Dunn, R.R.; Trautwein, M.D. Biodiversity and socioeconomics in the city: A review of the luxury effect. Biol. Lett. 2018, 14, 20180082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaninotto, V.; Thebault, E.; Dajoz, I. Native and exotic plants play different roles in urban pollination networks across seasons. Oecologia 2023, 201, 525–536. [Google Scholar] [CrossRef]
- Remme, R.P.; Frumkin, H.; Guerry, A.D.; King, A.C.; Mandle, L.; Sarabu, C.; Bratman, G.N.; Giles-Corti, B.; Hamel, P.; Han, B.; et al. An ecosystem service perspective on urban nature, physical activity, and health. Proc. Natl. Acad. Sci. USA 2021, 118, e2018472118. [Google Scholar] [CrossRef]
- Grêt-Regamey, A.; Galleguillos-Torres, M. Global urban homogenization and the loss of emotions. Sci. Rep. 2022, 12, 22515. [Google Scholar] [CrossRef]
- Bureau of Forestry and Landscape of Changchun, Landscape Plants in Changchun. 2016. Available online: http://ccylj.changchun.gov.cn/ylkp/ylkpej/201612/t20161222_793304.html (accessed on 13 September 2021). (In Chinese)
- Chen, D.; Fan, R.; Bai, S.L.; Ci, Z.L. Application Situation and Analysis of Garden Afforestation in the City of Huhhot. Tianjin Agric. Sci. 2009, 15, 61–64, (In Chinese, with English Summary). [Google Scholar]
- Fan, Q. A Comparative Study on the Change of Landscape Pattern and Biodiversity in Moderate-temperate Steppe Cities and Warm-temperate Steppe Cities in Inner Mongolia. Master’s Thesis, Inner Mongolia University, Hohhot, China, 2017. (In Chinese with English Summary). [Google Scholar]
- Guo, L.; Zhou, Z.L.; He, Y. Changes of landscape plants in Beijing in the past 60 years. In Beijing landscape architecture society, Beijing Bureau of Landscape Architecture; Beijing Park Management Center, Ed.; Beijing Ecological Garden City Construction; China Forestry Publishing House: Beijing, China, 2010; pp. 201–208. (In Chinese) [Google Scholar]
- Guo, Y. Design and Application of Landscape Plants; Heilongjiang Fine Art Press: Harbin, China, 2011. (In Chinese) [Google Scholar]
- Li, J. Study on Necessity and Feasibility of Urban Forestry Construction in Xi’an. Master’s Thesis, Northwest A&F University, Xianyang, China, 2009. (In Chinese, with English Summary). [Google Scholar]
- Li, N. The Present Evolution of Plant Diversity in Urumqi. Master’s Thesis, Xinjiang Agricultural University, Urumqi, China, 2009. (In Chinese, with English Summary). [Google Scholar]
- Li, P.B. Investigation, Evaluation and Planning of Landscape Woody Plants of Taian City. Master’s Thesis, Shandong Agricultural University, Taian, China, 2002. (In Chinese, with English Summary). [Google Scholar]
- Qingdao Bureau of Landscape Architecture and Forestry, Two Books, “The Monographs of Trees in Qingdao” and “Atlas of Qingdao Main Greening Trees”, Were Published. 2016. Available online: http://ly.qingdao.gov.cn/n32205788/n32205856/160122091229024186.html (accessed on 10 September 2021). (In Chinese)
- Shijiazhuang Bureau of Landscape Architecture, Procurement of Landscape Plants. 2020. Available online: http://ylj.sjz.gov.cn/col/1526374018015/index.html (accessed on 10 September 2021). (In Chinese)
- Tao, L. Investigation and construction of greening in Taiyuan. Inn. Mong. For. Sci. Technol. 2010, 36, 86–88, (In Chinese, with English Summary). [Google Scholar]
- Tianjin Municipal Commission of Urban Management, List of Trees and Other Plants Suitable for Planting in Tianjin. 2014. Available online: http://csgl.tj.gov.cn/zwgk_57/xzcwj/scsglw/CGWWJ/202011/t20201111_4052654.html (accessed on 13 September 2021). (In Chinese)
- Wang, H.J. Study on Community Structure, Ecological Benefits and Optimized Models of Waterfront Green Space in Jilin City. Ph.D. Thesis, Northeast Forestry University, Harbin, China, 2014. (In Chinese, with English Summary). [Google Scholar]
- Wei, Z.H. Study on the Geographical Composition of the Flora of Woody Garden Plants in Anyang City. Master’s Thesis, Henan Agricultural University, Zhengzhou, China, 2006. (In Chinese, with English Summary). [Google Scholar]
- Wu, Z.Y. China Vegetation; Science Press: Beijing, China, 1980. (In Chinese) [Google Scholar]
- Xin, Q.R. Atlas of Woody Plants in Xilingol League, Inner Mongolia Autonomous Region; China Machine Press: Beijing, China, 2016. (In Chinese) [Google Scholar]
- Yu, J.Q.; Su, X.C.; L, L. Planning of greening tree species in Jinan City. Sci. Technol. Landsc. Archit. 2007, 1, 32–37. (In Chinese) [Google Scholar]
- Zhang, N.N. Zhengzhou City Park Green Plant Diversity and Landscape Research. Master’s Thesis, Northwest A&F University, Xianyang, China, 2013. (In Chinese, with English Summary). [Google Scholar]
- Zhang, Y.; Zhao, F.; Wu, Y.H.; Huang, R. Selection and quantitative evaluation of main suitable ornamental tree species for gardens in Lanzhou City. J. Northwest For. Univ. 2019, 34, 255–267, (In Chinese, with English Summary). [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Richness | Simpson Index | Shannon–Wiener Index | Pielou Index | |||||
|---|---|---|---|---|---|---|---|---|
| UFs | RFs | UFs | RFs | UFs | RFs | UFs | RFs | |
| MIN | 99 | 22 | 0.33 | 0.12 | 0.40 | 0.19 | 0.43 | 0.13 |
| MAX | 325 | 675 | 0.68 | 0.74 | 1.29 | 2.23 | 0.82 | 0.78 |
| MEAN | 182 | 484 | 0.53 | 0.67 | 0.84 | 1.84 | 0.66 | 0.68 |
| SD | 57.33 | 129.25 | 0.10 | 0.14 | 0.18 | 0.42 | 0.11 | 0.14 |
| CV (%) | 26.70 | 31.43 | 18.87 | 20.23 | 21.67 | 23.12 | 16.60 | 20.75 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, G.; Ouyang, Y.; Hou, X.; Zhou, T.; Ge, Y.; Lu, Y.; Wang, Y.; Chang, J. Homogenization of Urban Forests across the Subtropical Zones of China. Land 2023, 12, 1559. https://doi.org/10.3390/land12081559
Yang G, Ouyang Y, Hou X, Zhou T, Ge Y, Lu Y, Wang Y, Chang J. Homogenization of Urban Forests across the Subtropical Zones of China. Land. 2023; 12(8):1559. https://doi.org/10.3390/land12081559
Chicago/Turabian StyleYang, Guofu, Yan Ouyang, Xiaoyu Hou, Tianyi Zhou, Ying Ge, Yijun Lu, Yue Wang, and Jie Chang. 2023. "Homogenization of Urban Forests across the Subtropical Zones of China" Land 12, no. 8: 1559. https://doi.org/10.3390/land12081559
APA StyleYang, G., Ouyang, Y., Hou, X., Zhou, T., Ge, Y., Lu, Y., Wang, Y., & Chang, J. (2023). Homogenization of Urban Forests across the Subtropical Zones of China. Land, 12(8), 1559. https://doi.org/10.3390/land12081559