
Temperature Mainly Determined the Seasonal Variations in Soil Faunal Communities in Semiarid Areas

Abstract
:
1. Introduction
2. Materials and Methods
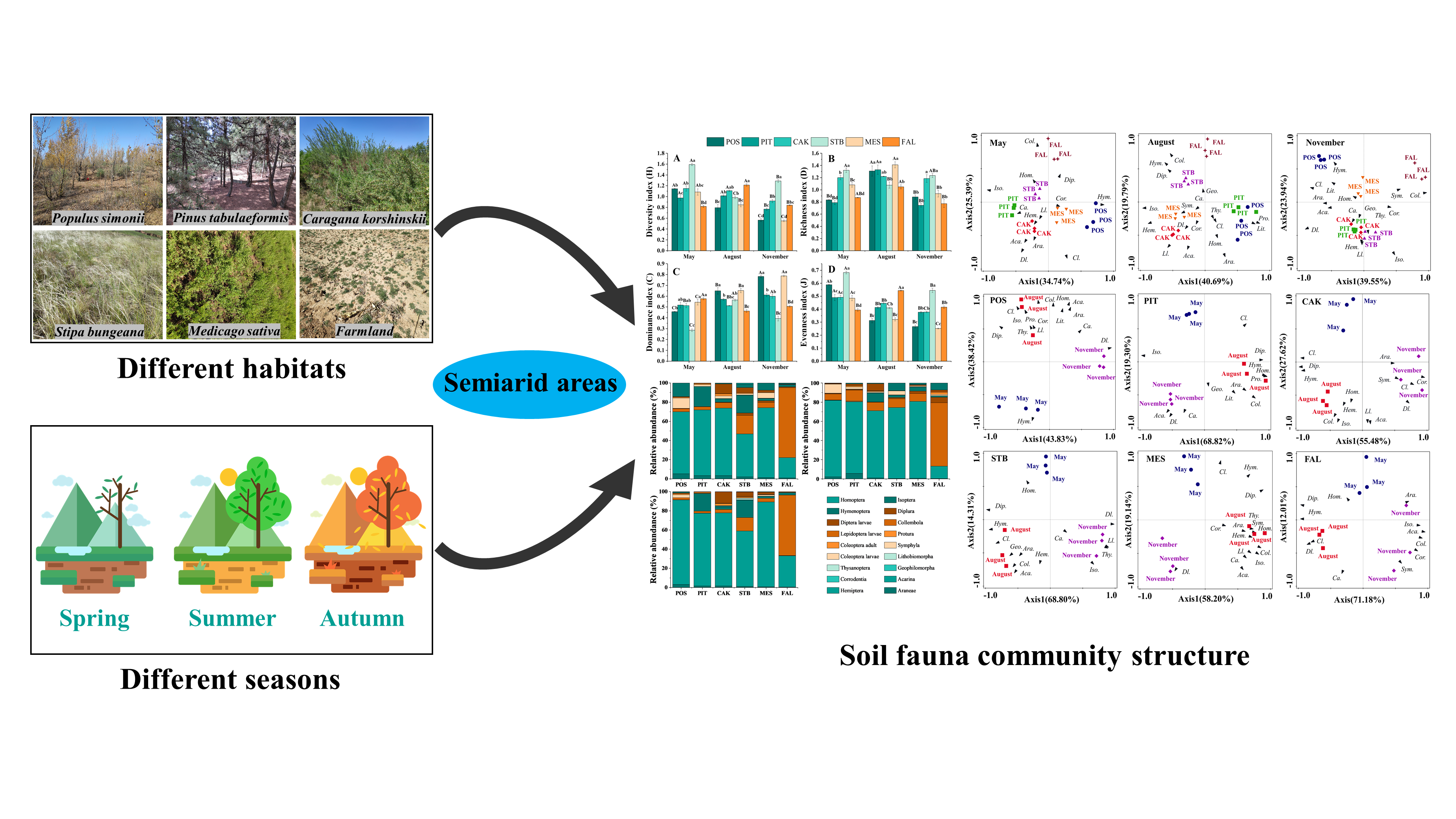
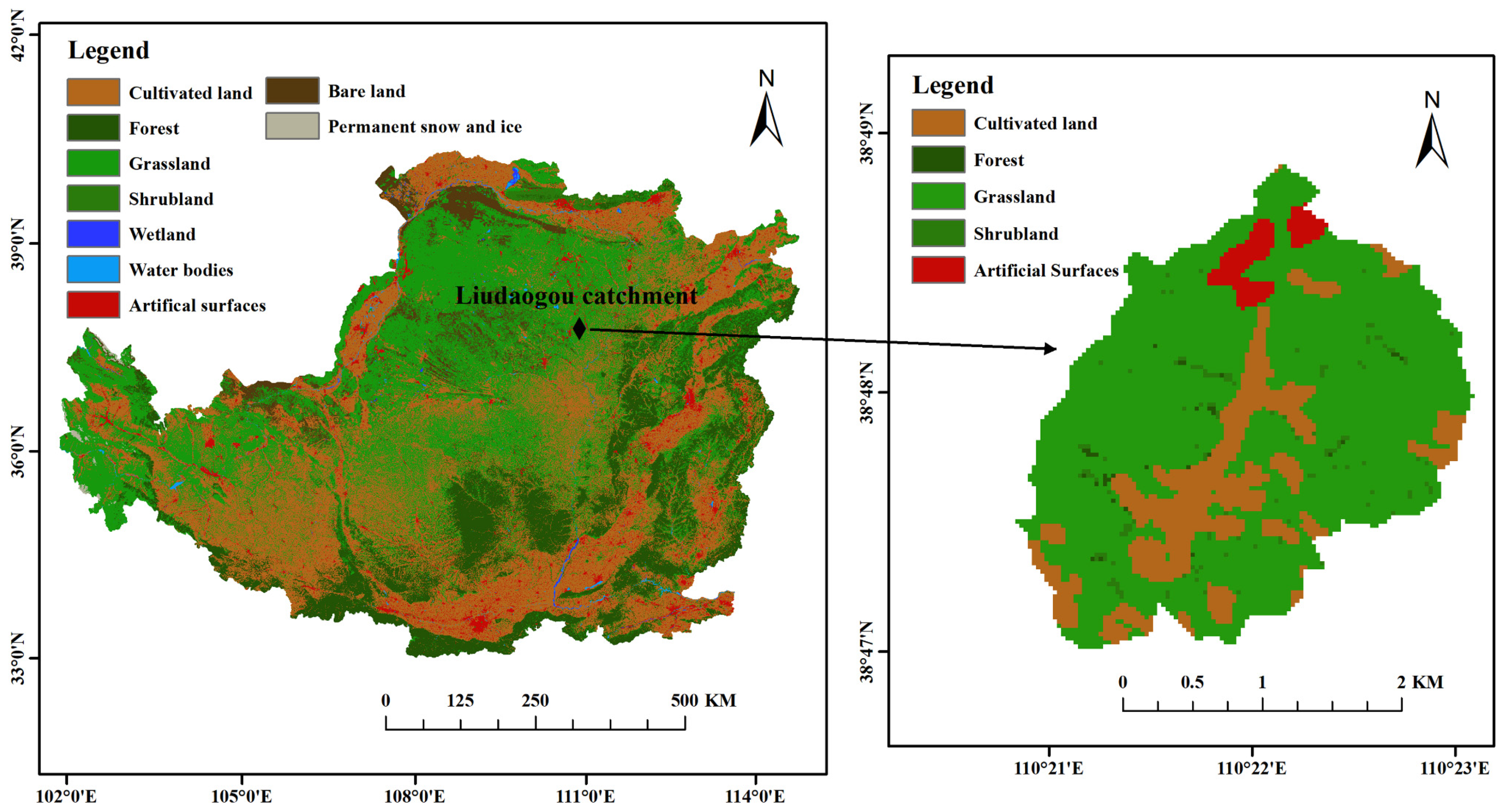
2.1. Study Sites and Selection of Habitats
2.2. Collection and Processing of Samples
2.3. Statistical Analysis
3. Results
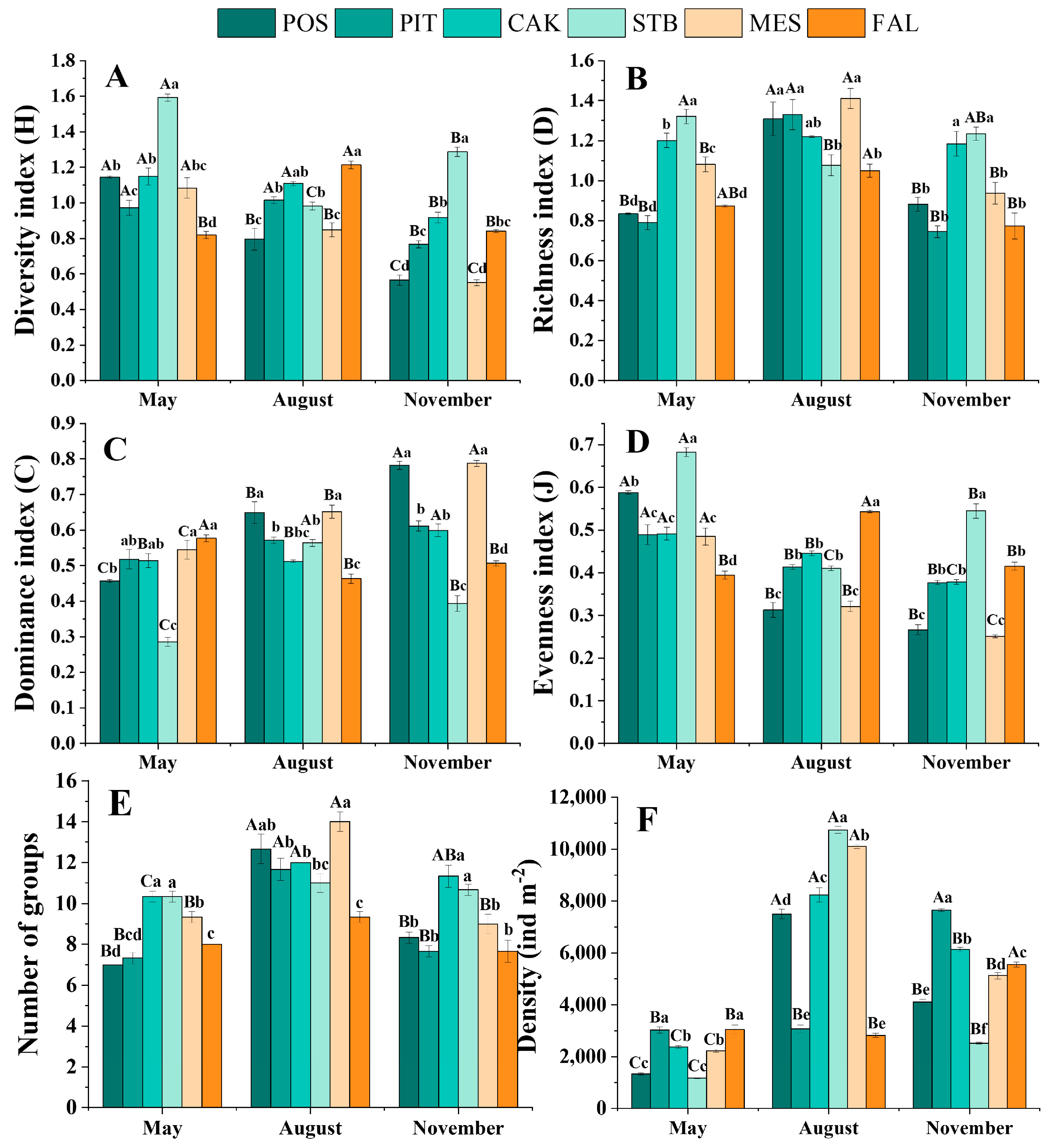
3.1. Seasonal Variation in Soil Faunal Density and Diversity
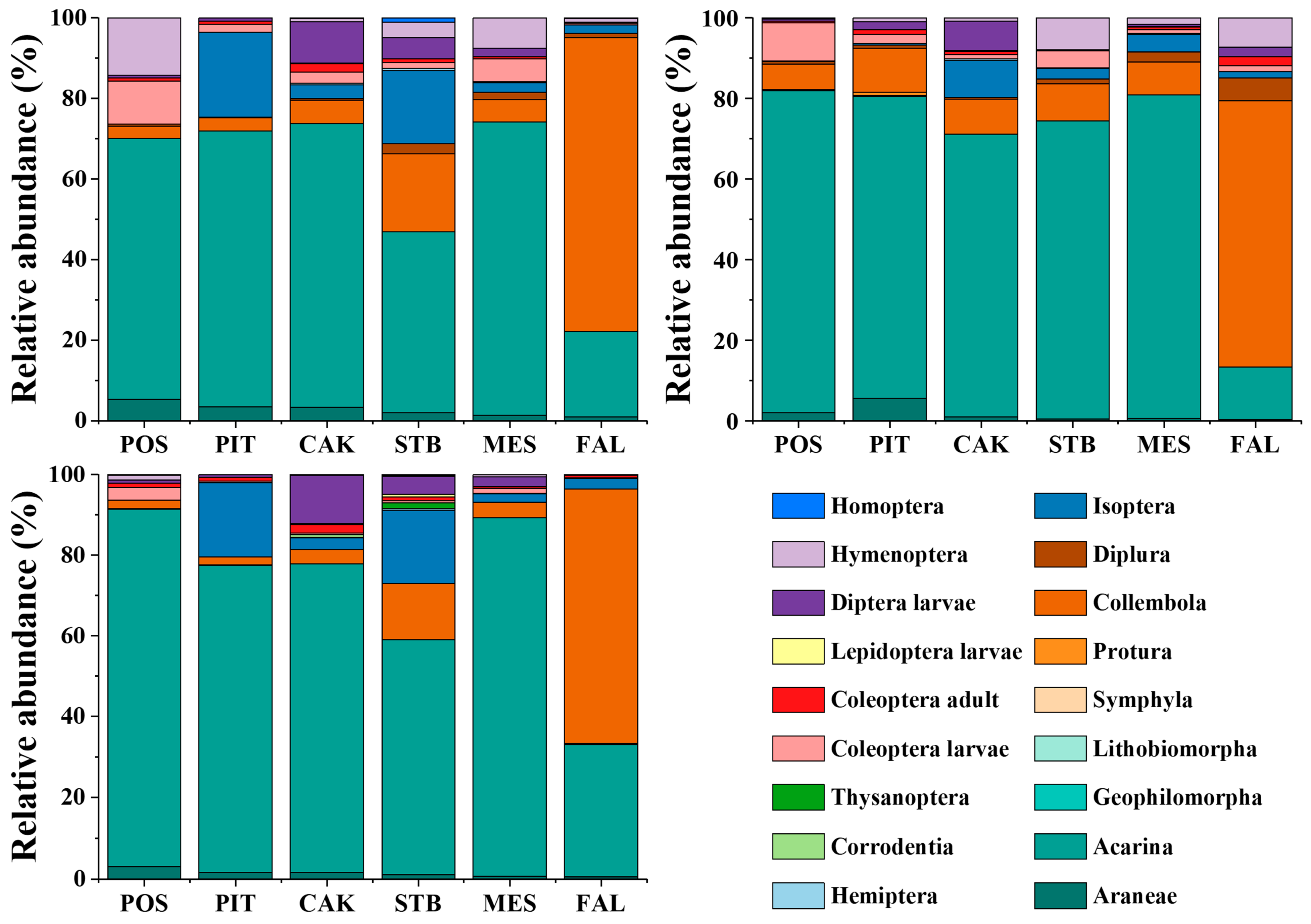
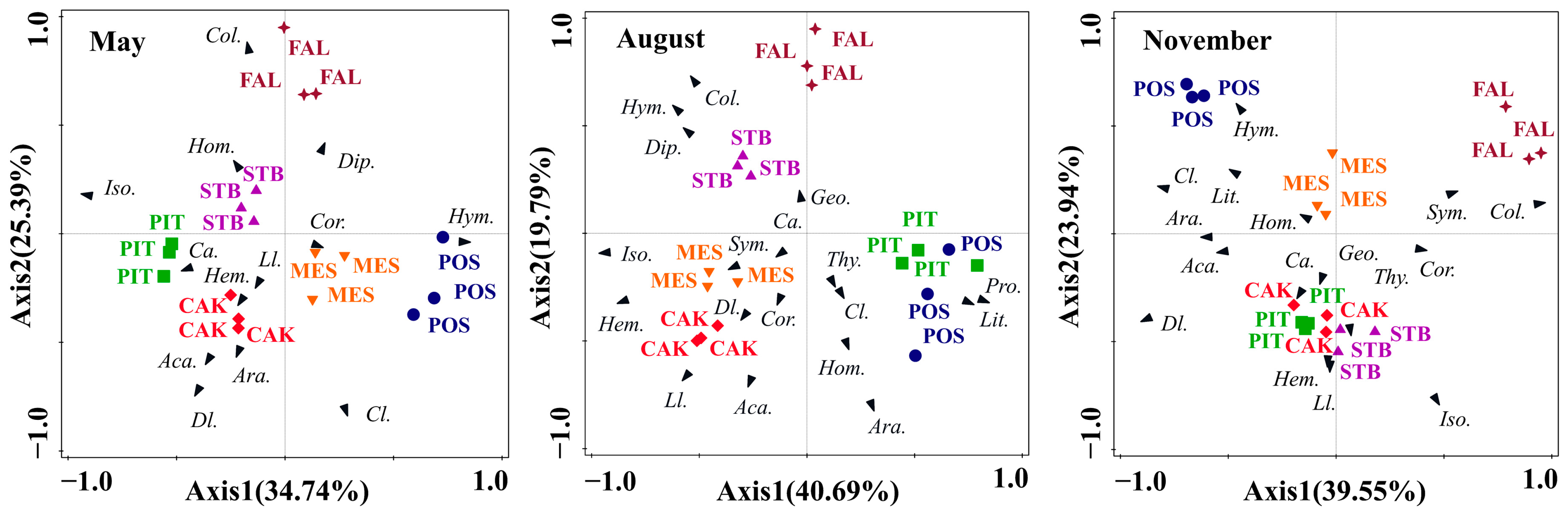
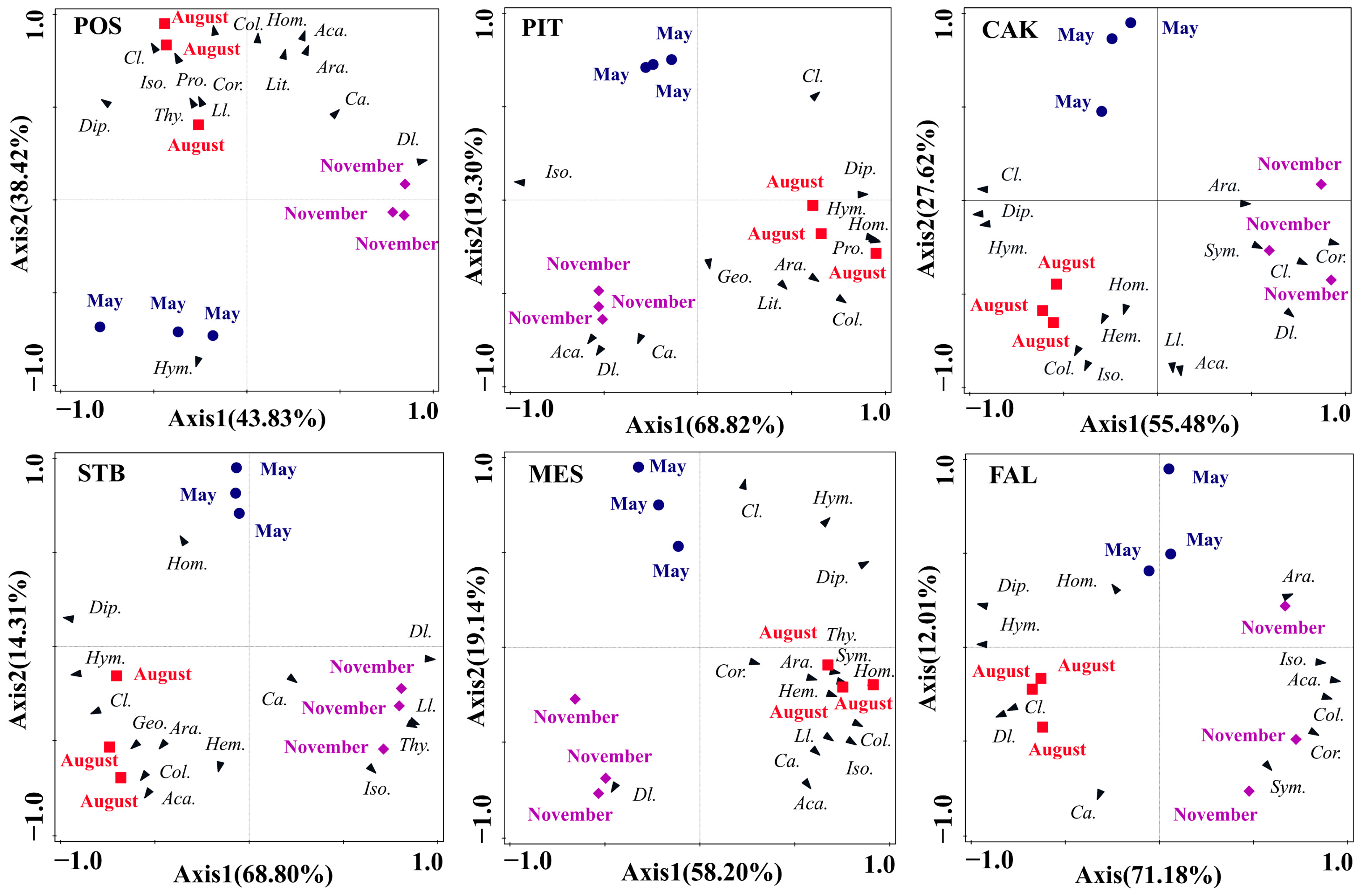
3.2. Seasonal Variation in Soil Faunal Community Composition
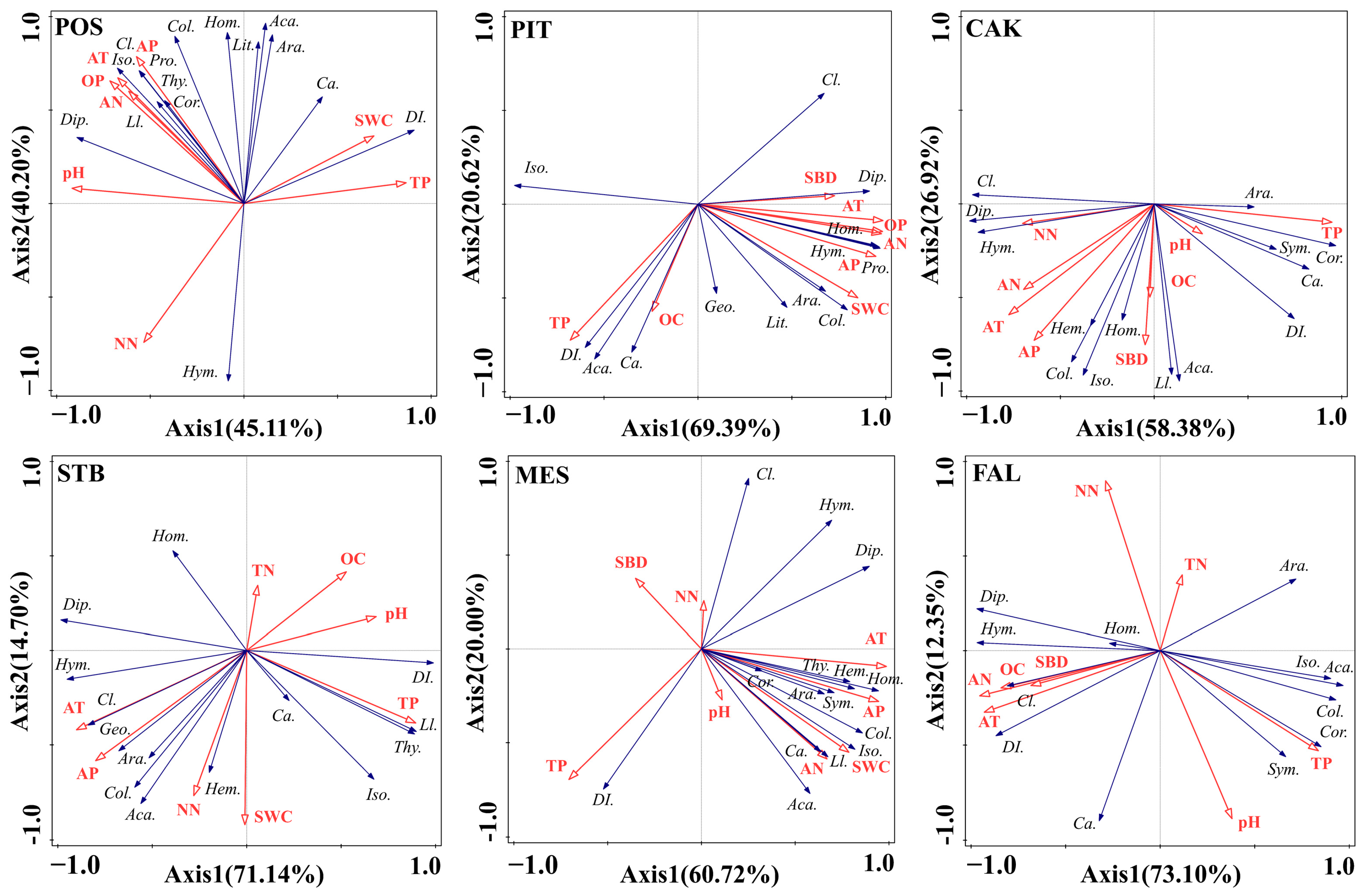
3.3. Driving Factors of Seasonal Variation of the Soil Faunal Community
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bardgett, R.D.; van der Putten, W.H. Belowground biodiversity and ecosystem functioning. Nature 2014, 515, 505–511. [Google Scholar] [CrossRef] [PubMed]
- Wagg, C.; Bender, S.F.; Widmer, F.; van der Heijden, M.G.A. Soil biodiversity and soil community composition determine ecosystem multifunctionality. Proc. Natl. Acad. Sci. USA 2014, 111, 5266–5270. [Google Scholar] [CrossRef] [PubMed]
- David, J.F. The role of litter-feeding macroarthropods in decomposition processes: A reappraisal of common views. Soil Biol. Biochem. 2014, 76, 109–118. [Google Scholar] [CrossRef]
- Frouz, J. Effects of soil macro- and mesofauna on litter decomposition and soil organic matter stabilization. Geoderma 2018, 332, 161–172. [Google Scholar] [CrossRef]
- Li, T.; Jia, Y.; Shao, M.a.; Shen, N. Camponotus japonicus burrowing activities exacerbate soil erosion on bare slopes. Geoderma 2019, 348, 158–167. [Google Scholar] [CrossRef]
- Li, T.; Shao, M.a.; Jia, Y.; Jia, X.; Huang, L. Small-scale observation on the effects of the burrowing activities of mole crickets on soil erosion and hydrologic processes. Agric. Ecosyst. Environ. 2018, 261, 136–143. [Google Scholar] [CrossRef]
- Bradford, M.A.; Jones, T.H.; Bardgett, R.D.; Black, H.I.J.; Boag, B.; Bonkowski, M.; Cook, R.; Eggers, T.; Gange, A.C.; Grayston, S.J.; et al. Impacts of Soil Faunal Community Composition on Model Grassland Ecosystems. Science 2002, 298, 615–618. [Google Scholar] [CrossRef] [PubMed]
- Eisenhauer, N.; Sabais, A.C.W.; Scheu, S. Collembola species composition and diversity effects on ecosystem functioning vary with plant functional group identity. Soil Biol. Biochem. 2011, 43, 1697–1704. [Google Scholar] [CrossRef]
- Lensing, J.R.; Wise, D.H. Predicted climate change alters the indirect effect of predators on an ecosystem process. Proc. Natl. Acad. Sci. USA 2006, 103, 15502–15505. [Google Scholar] [CrossRef]
- Erktan, A.; Or, D.; Scheu, S. The physical structure of soil: Determinant and consequence of trophic interactions. Soil Biol. Biochem. 2020, 148, 107876. [Google Scholar] [CrossRef]
- Gongalsky, K.B. Soil macrofauna: Study problems and perspectives. Soil Biol. Biochem. 2021, 159, 108281. [Google Scholar] [CrossRef]
- Eisenhauer, N.; Cesarz, S.; Koller, R.; Worm, K.; Reich, P.B. Global change belowground: Impacts of elevated CO2, nitrogen, and summer drought on soil food webs and biodiversity. Glob. Chang. Biol. 2012, 18, 435–447. [Google Scholar] [CrossRef]
- Kardol, P.; Reynolds, W.N.; Norby, R.J.; Classen, A.T. Climate change effects on soil microarthropod abundance and community structure. Appl. Soil Ecol. 2011, 47, 37–44. [Google Scholar] [CrossRef]
- Berg, M.P.; Bengtsson, J. Temporal and spatial variability in soil food web structure. Oikos 2007, 116, 1789–1804. [Google Scholar] [CrossRef]
- Handa, I.T.; Aerts, R.; Berendse, F.; Berg, M.P.; Bruder, A.; Butenschoen, O.; Chauvet, E.; Gessner, M.O.; Jabiol, J.; Makkonen, M.; et al. Consequences of biodiversity loss for litter decomposition across biomes. Nature 2014, 509, 218–221. [Google Scholar] [CrossRef]
- Wu, P.; Liu, S.; Liu, X. Composition and spatio-temporal changes of soil macroinvertebrates in the biodiversity hotspot of northern Hengduanshan Mountains, China. Plant Soil 2012, 357, 321–338. [Google Scholar] [CrossRef]
- Han, F.; Yu, C.; Fu, G. Non-growing/growing season non-uniform-warming increases precipitation use efficiency but reduces its temporal stability in an alpine meadow. Front. Plant Sci. 2023, 14, 1090204. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.; Wang, C. Differences in spatiotemporal dynamics between soil macrofauna and mesofauna communities in forest ecosystems: The significance for soil fauna diversity monitoring. Geoderma 2019, 337, 266–272. [Google Scholar] [CrossRef]
- Gao, H.; Pang, G.; Li, Z.; Chang, S. Evaluating the potential of vegetation restoration in the Loess Plateau. Acta Geogr. Sin. 2017, 72, 863–874. [Google Scholar]
- Wang, Y.; Shao, M.a.; Liu, Z. Large-scale spatial variability of dried soil layers and related factors across the entire Loess Plateau of China. Geoderma 2010, 159, 99–108. [Google Scholar] [CrossRef]
- Zhu, Y.; Wang, Y.; Chen, L.; Li, Z. Does non-native black locust afforestation affect soil biodiversity at the regional scale? Case study of soil macroinvertebrates across the Chinese Loess Plateau. Catena 2021, 200, 105171. [Google Scholar] [CrossRef]
- Yang, X.; Shao, M.a.; Li, T.; Gan, M.; Chen, M. Community characteristics and distribution patterns of soil fauna after vegetation restoration in the northern Loess Plateau. Ecol. Indic. 2021, 122, 107236. [Google Scholar] [CrossRef]
- The Food and Agriculture Organization of the United Nations. World Reference Base for Soil Resources; World Soil Resources Reports 103; The Food and Agriculture Organization of the United Nations: Rome, Italy, 2006. [Google Scholar]
- Anderson, J.; Ingram, J. Tropical Soil Biology and Fertility: A Handbook of Methods. Soil Sci. 1994, 157, 265. [Google Scholar] [CrossRef]
- Yang, X. Geographic Distribution of Grassland Soil Fauna and Their Ecological Functions on the Loess Plateau. Ph.D. Thesis, Northwest A&F University, Yangling, China, 2023. [Google Scholar]
- Zhu, X.; Zhu, B. Diversity and abundance of soil fauna as influenced by long-term fertilization in cropland of purple soil, China. Soil Tillage Res. 2015, 146, 39–46. [Google Scholar] [CrossRef]
- Wenying, Y. Pictorial Keys to Soil Animals of China; Science Press: Beijing, China, 2000. [Google Scholar]
- Page, A.L. Methods of Soil Analysis. Part 2. Chemical and Microbiological Properties; Amer Society of Agronomy: Madison, WI, USA, 1982. [Google Scholar]
- Kachurina, O.; Zhang, H.; Raun, W.; Krenzer, E. Simultaneous determination of soil aluminum, ammonium- and nitrate-nitrogen using 1 M potassium chloride extraction. Commun. Soil Sci. Plant Anal. 2000, 31, 893–903. [Google Scholar] [CrossRef]
- Dick, W.A.; Cheng, L.; Wang, P. Soil acid and alkaline phosphatase activity as pH adjustment indicators. Soil Biol. Biochem. 2000, 32, 1915–1919. [Google Scholar] [CrossRef]
- Cao, W. Community Structure of Soil Fauna and Its Seasonal Dynaic of Wetland in Nanniwan; Yan’An University: Yan’an City, China, 2017. [Google Scholar]
- Wu, P.; Liu, X.; Liu, S.; Wang, J.; Wang, Y. Composition and spatio-temporal variation of soil microarthropods in the biodiversity hotspot of northern Hengduan Mountains, China. Eur. J. Soil Biol. 2014, 62, 30–38. [Google Scholar] [CrossRef]
- Decaëns, T.; Jiménez, J.J.; Barros, E.; Chauvel, A.; Blanchart, E.; Fragoso, C.; Lavelle, P. Soil macrofaunal communities in permanent pastures derived from tropical forest or savanna. Agric. Ecosyst. Environ. 2004, 103, 301–312. [Google Scholar] [CrossRef]
- Rossi, J.-P.; Blanchart, E. Seasonal and land-use induced variations of soil macrofauna composition in the Western Ghats, southern India. Soil Biol. Biochem. 2005, 37, 1093–1104. [Google Scholar] [CrossRef]
- Liu, Y.; Chang, L.; Zhang, S.; Gao, Q.; Xin, S. Effects of Farmland Management Measures on the Characteristics of Soil Animal Community. Chin. J. Soil Sci. 2023, 54, 989. [Google Scholar]
- Sauvadet, M.; Chauvat, M.; Brunet, N.; Bertrand, I. Can changes in litter quality drive soil fauna structure and functions? Soil Biol. Biochem. 2017, 107, 94–103. [Google Scholar] [CrossRef]
- Cui, Y.; Fang, L.; Guo, X.; Wang, X.; Zhang, Y.; Li, P.; Zhang, X. Ecoenzymatic stoichiometry and microbial nutrient limitation in rhizosphere soil in the arid area of the northern Loess Plateau, China. Soil Biol. Biochem. 2018, 116, 11–21. [Google Scholar] [CrossRef]
- Chang, H. Spatial-Temporal Distribution Characteristics of Soil Faunal Community Structure after Planting Caragana Korshinskii in Reaumuria Soongorica Shrubbery Land of Urad Desert Steppe; Ningxia University: Yinchuan, China, 2020. [Google Scholar]
- Aupic-Samain, A.; Baldy, V.; Delcourt, N.; Krogh, P.H.; Gauquelin, T.; Fernandez, C.; Santonja, M. Water availability rather than temperature control soil fauna community structure and prey–predator interactions. Funct. Ecol. 2021, 35, 1550–1559. [Google Scholar] [CrossRef]
- Schlesinger, W.; Bernhardt, E. Biogeochemistry: An Analysis of Global Change, 3rd ed.; Academic Press: Cambridge, MA, USA, 2013; 672p. [Google Scholar]
- Bokhorst, S.; Huiskes, A.; Convey, P.; van Bodegom, P.M.; Aerts, R. Climate change effects on soil arthropod communities from the Falkland Islands and the Maritime Antarctic. Soil Biol. Biochem. 2008, 40, 1547–1556. [Google Scholar] [CrossRef]
- Melillo, J.M.; McGuire, A.D.; Kicklighter, D.W.; Moore, B.; Vorosmarty, C.J.; Schloss, A.L. Global climate change and terrestrial net primary production. Nature 1993, 363, 234–240. [Google Scholar] [CrossRef]
- Blankinship, J.C.; Niklaus, P.A.; Hungate, B.A. A meta-analysis of responses of soil biota to global change. Oecologia 2011, 165, 553–565. [Google Scholar] [CrossRef]
- Coulson, S.J.; Leinaas, H.P.; Ims, R.A.; Søvik, G. Experimental manipulation of the winter surface ice layer: The effects on a High Arctic soil microarthropod community. Ecography 2000, 23, 299–306. [Google Scholar] [CrossRef]
- Wiwatwitaya, D.; Takeda, H. Seasonal changes in soil arthropod abundance in the dry evergreen forest of north-east Thailand, with special reference to collembolan communities. Ecol. Res. 2005, 20, 59–70. [Google Scholar] [CrossRef]
- Wang, S.; Ruan, H.; Wang, J.; Xu, Z.; Wu, Y. Composition structure of soil fauna community under the typical vegetations in the Wuyi Mountains, China. Acta Ecol. Sin. 2010, 30, 5174–5184. [Google Scholar]
- Zhu, Y.; Bian, H.; Ju, C.; Xu, C.; Zhou, Y.; Zhang, H.; Xu, X. Fertilization alters the abundance but not the diversity of soil fauna: A meta-analysis. Glob. Ecol. Biogeogr. 2023, 32, 482–494. [Google Scholar] [CrossRef]
- Berlinches de Gea, A.; Hautier, Y.; Geisen, S. Interactive effects of global change drivers as determinants of the link between soil biodiversity and ecosystem functioning. Glob. Chang. Biol. 2023, 29, 296–307. [Google Scholar] [CrossRef]
- Yin, R.; Qin, W.; Wang, X.; Xie, D.; Wang, H.; Zhao, H.; Zhang, Z.; He, J.-S.; Schädler, M.; Kardol, P.; et al. Experimental warming causes mismatches in alpine plant-microbe-fauna phenology. Nat. Commun. 2023, 14, 2159. [Google Scholar] [CrossRef]
- Zhang, B.; Ma, W.; Song, L.; Liang, X.; Xi, X.; Wang, Z. Nitrogen addition and experimental drought simplified arthropod network in temperate grassland. Funct. Ecol. 2023, 37, 1815–1826. [Google Scholar] [CrossRef]
- Peng, Y.; Peñuelas, J.; Vesterdal, L.; Yue, K.; Peguero, G.; Fornara, D.A.; Heděnec, P.; Steffens, C.; Wu, F. Responses of soil fauna communities to the individual and combined effects of multiple global change factors. Ecol. Lett. 2022, 25, 1961–1973. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Habitats | Latitude and Longitude | Altitude (m) | Soil Texture | Density (ind m−2) | Vegetation Coverage (%) | Litter Thickness (cm) |
|---|---|---|---|---|---|---|
| Populus simonii | 110°21′40.39″ E 38°47′5.57″ N | 1221 | loamy sand | 0.83 | 63.67 | 3.40 |
| Pinus tabuliformis | 110°21′50.49″ E 38°47′8.00″ N | 1232 | loam | 0.83 | 92.36 | 5.20 |
| Caragana korshinskii | 110°21′54.45″ E 38°47′25.29″ N | 1216 | loam | 0.41 | 80.38 | 2.20 |
| Stipa bungeana | 110°21′50.95″ E 38°47′26.59″ N | 1224 | loam | 64 | 85.56 | 0.30 |
| Medicago Sativa | 110°21′52.62″ E 38°47′35.85″ N | 1187 | loam | 14.23 | 89.04 | 0.83 |
| Farmland (Zea mays) | 110°21′50.19″ E 38°47′42.97″ N | 1155 | loam | \ | \ | \ |
| Habitats | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| POS | PIT | CAK | STB | MES | FAL | |||||||||||||
| Taxa | May | August | November | May | August | November | May | August | November | May | August | November | May | August | November | May | August | November |
| Araneae | 70 ± 8 | 150 ± 22 | 127 ± 10 | 103 ± 10 | 170 ± 17 | 123 ± 11 | 80 ± 8 | 77 ± 12 | 97 ± 7 | 23 ± 3 | 40 ± 5 | 27 ± 3 | 30 ± 5 | 63 ± 7 | 37 ± 10 | 30 ± 8 | 7 ± 3 | 30 ± 5 |
| Acarina | 867 ± 27 | 5980 ± 295 | 3627 ± 126 | 2080 ± 165 | 2293 ± 116 | 5810 ± 94 | 1677 ± 64 | 5773 ± 213 | 4680 ± 78 | 527 ± 44 | 7947 ± 178 | 1463 ± 95 | 1627 ± 69 | 8087 ± 92 | 4540 ± 96 | 647 ± 22 | 367 ± 24 | 1807 ± 52 |
| Geophilomorpha | / | / | / | / | 3 ± 3 | 3 ± 3 | / | / | / | / | 7 ± 3 | / | / | / | / | / | / | / |
| Lithobiomorpha | / | 10 ± 0 | 7 ± 3 | / | 7 ± 3 | 3 ± 3 | / | / | / | / | / | / | / | / | / | / | / | / |
| Symphyla | / | / | / | / | / | / | / | / | 3 ± 3 | / | / | / | / | 7 ± 3 | / | / | / | 10 ± 5 |
| Protura | / | 10 ± 5 | / | / | 23 ± 3 | / | / | / | / | / | / | / | / | / | / | / | / | / |
| Collembola | 40 ± 5 | 477 ± 28 | 87 ± 10 | 100 ± 12 | 337 ± 21 | 157 ± 7 | 137 ± 18 | 720 ± 37 | 217 ± 20 | 227 ± 44 | 983 ± 32 | 353 ± 31 | 123 ± 12 | 820 ± 66 | 197 ± 22 | 2230 ± 143 | 1867 ± 76 | 3507 ± 110 |
| Diplura | 7 ± 3 | 40 ± 5 | / | 3 ± 3 | 23 ± 3 | / | 10 ± 5 | 37 ± 5 | / | 30 ± 5 | 130 ± 12 | / | 40 ± 5 | 257 ± 28 | / | 30 ± 5 | 160 ± 5 | / |
| Isoptera | / | 7 ± 3 | / | 637 ± 50 | 13 ± 5 | 1410 ± 95 | 80 ± 12 | 757 ± 87 | 180 ± 14 | 213 ± 10 | 277 ± 10 | 460 ± 36 | 53 ± 3 | 443 ± 37 | 110 ± 12 | 67 ± 10 | 47 ± 7 | 140 ± 12 |
| Hemiptera | / | / | / | / | / | / | 10 ± 5 | 33 ± 7 | 10 ± 0 | 7 ± 3 | 17 ± 3 | 10 ± 0 | 3 ± 3 | 13 ± 3 | 3 ± 3 | / | / | / |
| Corrodentia | / | 3 ± 3 | / | / | / | / | / | / | 40 ± 5 | / | / | / | 3 ± 3 | 7 ± 3 | 3 ± 3 | / | / | 10 ± 0 |
| Thysanoptera | / | 7 ± 3 | / | / | / | / | / | / | / | / | / | 37 ± 3 | / | 7 ± 3 | / | / | / | / |
| Coleoptera larvae | 143 ± 12 | 710 ± 92 | 127 ± 7 | 60 ± 9 | 67 ± 11 | 23 ± 3 | 67 ± 5 | 87 ± 3 | 23 ± 3 | 17 ± 3 | 447 ± 32 | 13 ± 7 | 127 ± 11 | 90 ± 0 | 63 ± 5 | 10 ± 5 | 37 ± 5 | 3 ± 3 |
| Coleoptera adult | 10 ± 5 | 23 ± 5 | 43 ± 10 | 27 ± 5 | 37 ± 7 | 73 ± 7 | 50 ± 5 | 60 ± 9 | 127 ± 10 | 10 ± 5 | 20 ± 9 | 23 ± 7 | 10 ± 5 | 63 ± 10 | 20 ± 8 | 10 ± 0 | 63 ± 5 | 37 ± 10 |
| Lepidoptera larvae | / | 3 ± 3 | / | / | / | / | 3 ± 3 | 23 ± 3 | 17 ± 5 | / | / | 17 ± 3 | / | 10 ± 0 | 3 ± 3 | / | / | / |
| Diptera larvae | 10 ± 5 | 33 ± 3 | 37 ± 10 | 23 ± 3 | 60 ± 9 | 53 ± 10 | 243 ± 14 | 600 ± 8 | 737 ± 81 | 63 ± 11 | 13 ± 3 | 113 ± 10 | 50 ± 12 | 53 ± 7 | 123 ± 18 | / | 67 ± 7 | / |
| Hymenoptera | 190 ± 9 | 27 ± 3 | 47 ± 10 | / | 30 ± 5 | / | 20 ± 5 | 63 ± 7 | 3 ± 3 | 43 ± 5 | 853 ± 53 | 7 ± 3 | 167 ± 24 | 167 ± 21 | 27 ± 3 | 30 ± 5 | 207 ± 15 | 3 ± 3 |
| Homoptera | / | 20 ± 5 | 7 ± 3 | / | 13 ± 3 | / | 3 ± 3 | 13 ± 3 | 7 ± 3 | 13 ± 3 | 10 ± 5 | 3 ± 3 | / | 13 ± 3 | / | 3 ± 3 | 7 ± 3 | 3 ± 3 |
| Total density | 1337 ± 45 | 7500 ± 182 | 4107 ± 112 | 3033 ± 120 | 3077 ± 143 | 7657 ± 56 | 2380 ± 50 | 8243 ± 276 | 6140 ± 79 | 1173 ± 19 | 10743 ± 144 | 2527 ± 40 | 2233 ± 50 | 10100 ± 78 | 5127 ± 127 | 3057 ± 165 | 2827 ± 86 | 5550 ± 96 |
| Total number of groups | 7 ± 0 | 13 ± 1 | 8 ± 0 | 7 ± 0 | 12 ± 1 | 8 ± 0 | 10 ± 0 | 12 ± 0 | 11 ± 1 | 10 ± 0 | 11 ± 0 | 11 ± 0 | 9 ± 0 | 14 ± 0 | 9 ± 0 | 8 ± 0 | 9 ± 0 | 8 ± 1 |
| Diversity Index (H) | Richness Index (D) | Dominance Index (C) | Evenness Index (J) | Density | Number of Groups | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| F | p | F | p | F | p | F | p | F | p | F | p | |
| Habitat | 113.25 | <0.001 | 63.83 | <0.001 | 95.92 | <0.001 | 188.42 | <0.001 | 1981.47 | <0.001 | 107.62 | <0.001 |
| Season | 72.16 | <0.001 | 27.90 | <0.001 | 76.66 | <0.001 | 66.24 | <0.001 | 98.59 | <0.001 | 26.97 | <0.001 |
| Habitat × Season | 30.75 | <0.001 | 12.51 | <0.001 | 29.39 | <0.001 | 41.08 | <0.001 | 370.54 | <0.001 | 8.34 | <0.001 |
| Habitats | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| POS | PIT | CAK | STB | MES | FAL | |||||||||||||
| Parameters | May | August | November | May | August | November | May | August | November | May | August | November | May | August | November | May | August | November |
| SBD (g/cm3) | 1.2 ± 0.0 | 1.3 ± 0.0 | 1.3 ± 0.0 | 1.3 ± 0.0 | 1.4 ± 0.0 | 1.2 ± 0.1 | 1.2 ± 0.0 | 1.4 ± 0.1 | 1.4 ± 0.1 | 1.3 ± 0.0 | 1.3 ± 0.1 | 1.2 ± 0.1 | 1.3 ± 0.0 | 1.2 ± 0.1 | 1.2 ± 0.0 | 1.2 ± 0.0 | 1.3 ± 0.0 | 1.2 ± 0.0 |
| SWC (%) | 3.5 ± 0.1 | 4.8 ± 0.3 | 7.3 ± 1.2 | 4.6 ± 0.5 | 17.6 ± 0.2 | 8.5 ± 0.2 | 6.0 ± 0.1 | 12.6 ± 2.3 | 8.4 ± 1.6 | 2.7 ± 0.1 | 9.9 ± 0.8 | 9.3 ± 0.1 | 5.4 ± 0.7 | 12.8 ± 0.7 | 8.1 ± 0.6 | 2.8 ± 0.5 | 20.9 ± 1.3 | 12.8 ± 0.1 |
| pH | 8.6 ± 0.0 | 8.8 ± 0.1 | 7.9 ± 0.1 | 8.7 ± 0.0 | 8.8 ± 0.0 | 8.9 ± 0.2 | 8.4 ± 0.1 | 8.5 ± 0.1 | 8.7 ± 0.2 | 8.9 ± 0.0 | 8.7 ± 0.1 | 8.9 ± 0.1 | 8.5 ± 0.1 | 8.7 ± 0.1 | 8.7 ± 0.1 | 8.3 ± 0.0 | 8.7 ± 0.0 | 9.0 ± 0.0 |
| OC (g/kg) | 6.7 ± 0.4 | 3.6 ± 0.3 | 3.2 ± 0.5 | 5.4 ± 0.2 | 5.6 ± 0.8 | 6.7 ± 0.3 | 5.6 ± 0.9 | 7.4 ± 0.6 | 6.6 ± 1.0 | 8. ± 0.1 | 7.4 ± 1.0 | 9.0 ± 0.6 | 13.8 ± 0.3 | 6.5 ± 0.2 | 11.3 ± 0.2 | 7.4 ± 0.4 | 9.6 ± 0.8 | 6.5 ± 0.1 |
| TN (g/kg) | 0.4 ± 0.0 | 0.3 ± 0.0 | 0.4 ± 0.0 | 0.4 ± 0.0 | 0.6 ± 0.1 | 0.5 ± 0.0 | 0.5 ± 0.1 | 0.7 ± 0.1 | 0.6 ± 0.0 | 0.7 ± 0.0 | 0.7 ± 0.1 | 0.7 ± 0.1 | 0.9 ± 0.1 | 1.1 ± 0.2 | 0.9 ± 0.1 | 0.6 ± 0.0 | 0.5 ± 0.1 | 0.5 ± 0.0 |
| TP (g/kg) | 0.3 ± 0.0 | 0.3 ± 0.0 | 0.5 ± 0.0 | 0.4 ± 0.0 | 0.4 ± 0.0 | 0.8 ± 0.0 | 0.4 ± 0.0 | 0.4 ± 0.0 | 0.7 ± 0.1 | 0.5 ± 0.0 | 0.4 ± 0.0 | 0.9 ± 0.0 | 0.5 ± 0.0 | 0.4 ± 0.0 | 0.8 ± 0.0 | 0.5 ± 0.0 | 0.5 ± 0.0 | 1.0 ± 0.0 |
| NN (mg/kg) | 2.4 ± 0.3 | 1.0 ± 0.0 | 0.4 ± 0.1 | 0.6 ± 0.1 | 1.8 ± 0.1 | 0.5 ± 0.2 | 5.5 ± 0.2 | 6.6 ± 1.3 | 3.5 ± 0.3 | 0.5 ± 0.0 | 3.1 ± 0.7 | 2.1 ± 0.1 | 9.4 ± 2.3 | 9.1 ± 1.5 | 8.6 ± 1.1 | 15.2 ± 0.8 | 6.6 ± 0.5 | 2.6 ± 0.4 |
| AN (mg/kg) | 3.7 ± 0.6 | 6.8 ± 0.4 | 3.3 ± 0.7 | 2.9 ± 0.1 | 9.1 ± 0.5 | 2.2 ± 0.3 | 4.4 ± 1.2 | 6.9 ± 0.6 | 3.0 ± 0.6 | 2.8 ± 0.1 | 8.5 ± 0.5 | 2.9 ± 0.4 | 2.4 ± 0.1 | 9.8 ± 1.9 | 5.8 ± 0.9 | 3.3 ± 0.2 | 6.5 ± 0.3 | 2.3 ± 0.3 |
| OP (mg/kg) | 5.0 ± 0.2 | 24.5 ± 3.2 | 2.3 ± 0.4 | 2.6 ± 0.0 | 17.4 ± 2.1 | 1.2 ± 0.1 | 3.4 ± 0.2 | 34.0 ± 2.5 | 2.3 ± 0.4 | 3.3 ± 0.0 | 3.2 ± 0.4 | 1.7 ± 0.3 | 4.4 ± 0.3 | 3.5 ± 0.4 | 2.1 ± 0.2 | 4.7 ± 0.0 | 59.7 ± 3.9 | 0.7 ± 0.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Z.; Yang, X.; Long, W.; Song, R.; Zhu, X.; Li, T.; Shao, M.; Chen, M.; Gan, M. Temperature Mainly Determined the Seasonal Variations in Soil Faunal Communities in Semiarid Areas. Land 2024, 13, 505. https://doi.org/10.3390/land13040505
Li Z, Yang X, Long W, Song R, Zhu X, Li T, Shao M, Chen M, Gan M. Temperature Mainly Determined the Seasonal Variations in Soil Faunal Communities in Semiarid Areas. Land. 2024; 13(4):505. https://doi.org/10.3390/land13040505
Chicago/Turabian StyleLi, Zhiyong, Xi Yang, Wei Long, Ranran Song, Xuchao Zhu, Tongchuan Li, Ming’an Shao, Mingyu Chen, and Miao Gan. 2024. "Temperature Mainly Determined the Seasonal Variations in Soil Faunal Communities in Semiarid Areas" Land 13, no. 4: 505. https://doi.org/10.3390/land13040505
APA StyleLi, Z., Yang, X., Long, W., Song, R., Zhu, X., Li, T., Shao, M., Chen, M., & Gan, M. (2024). Temperature Mainly Determined the Seasonal Variations in Soil Faunal Communities in Semiarid Areas. Land, 13(4), 505. https://doi.org/10.3390/land13040505