Impact of Water Level on Species Quantity and Composition Grown from the Soil Seed Bank of the Inland Salt Marsh: An Ex-Situ Experiment


 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Factors Influencing the Experimental Design
- to keep the time of germination consistent, it is better to sample the soil seed bank in autumn;
- the sampling time and outdoor experimental conditions ensure that the seeds experience “natural” temperatures needed for germination;
- water in a sufficient amount should be present at all times;
- seeds require and should experience similar light conditions as they would in their habitat outdoors.
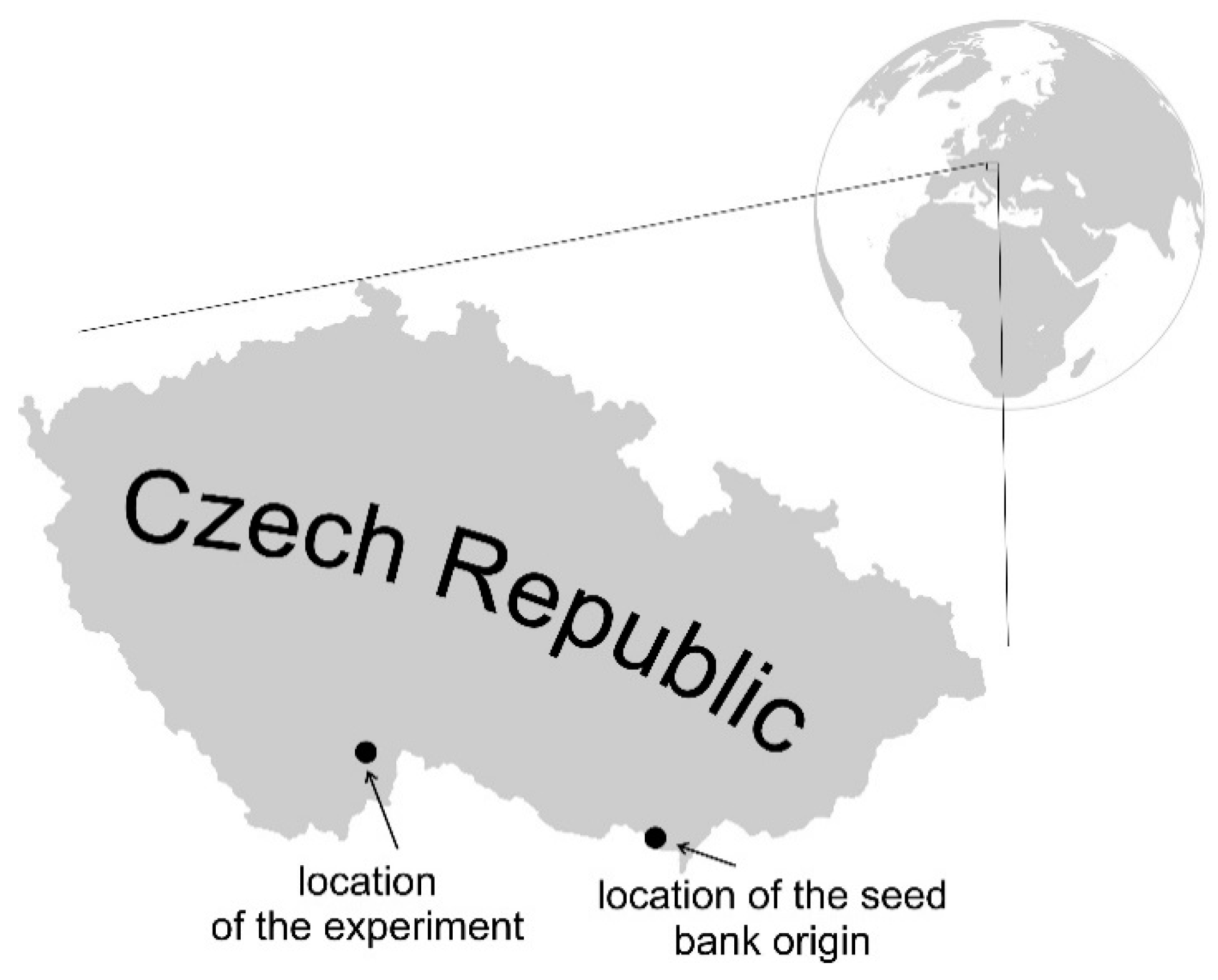
2.2. Seed Bank
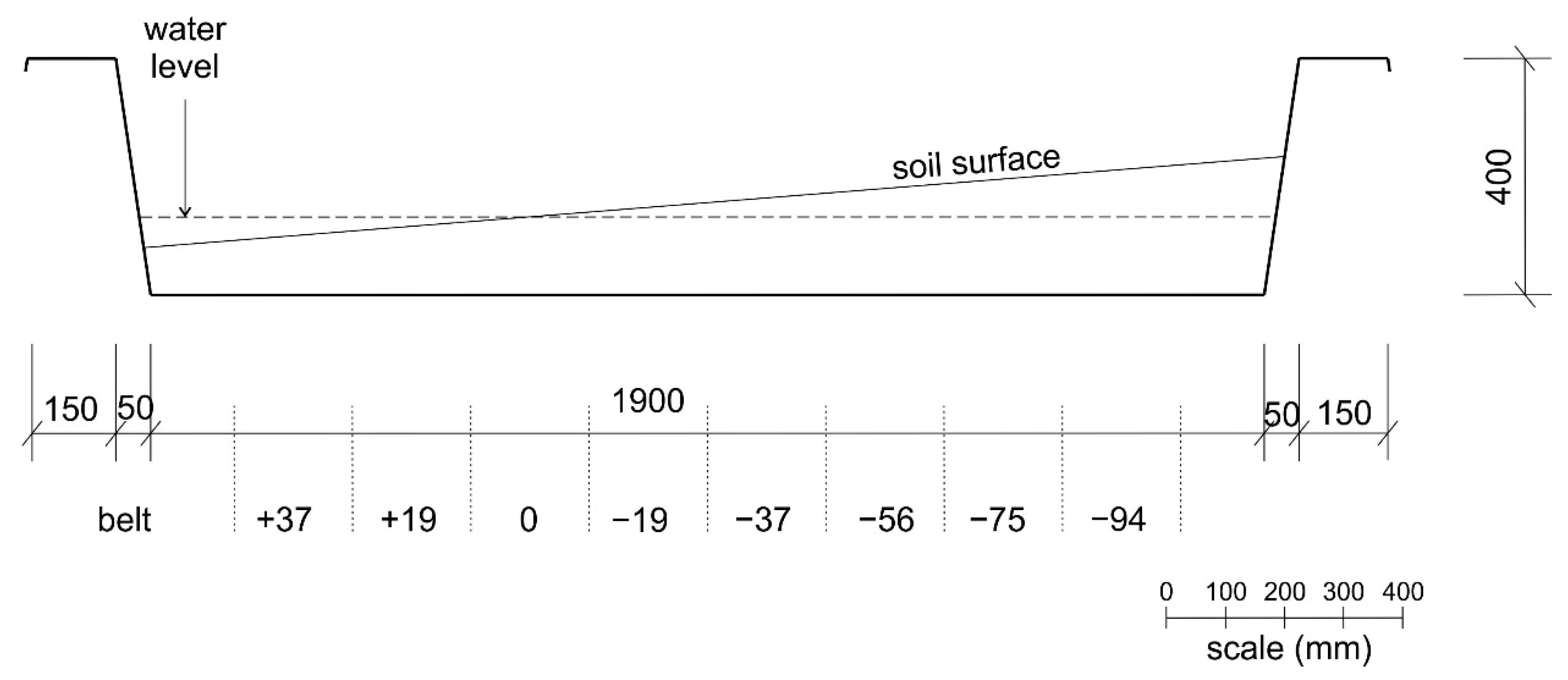
2.3. Experimental Design
2.4. Data Collection
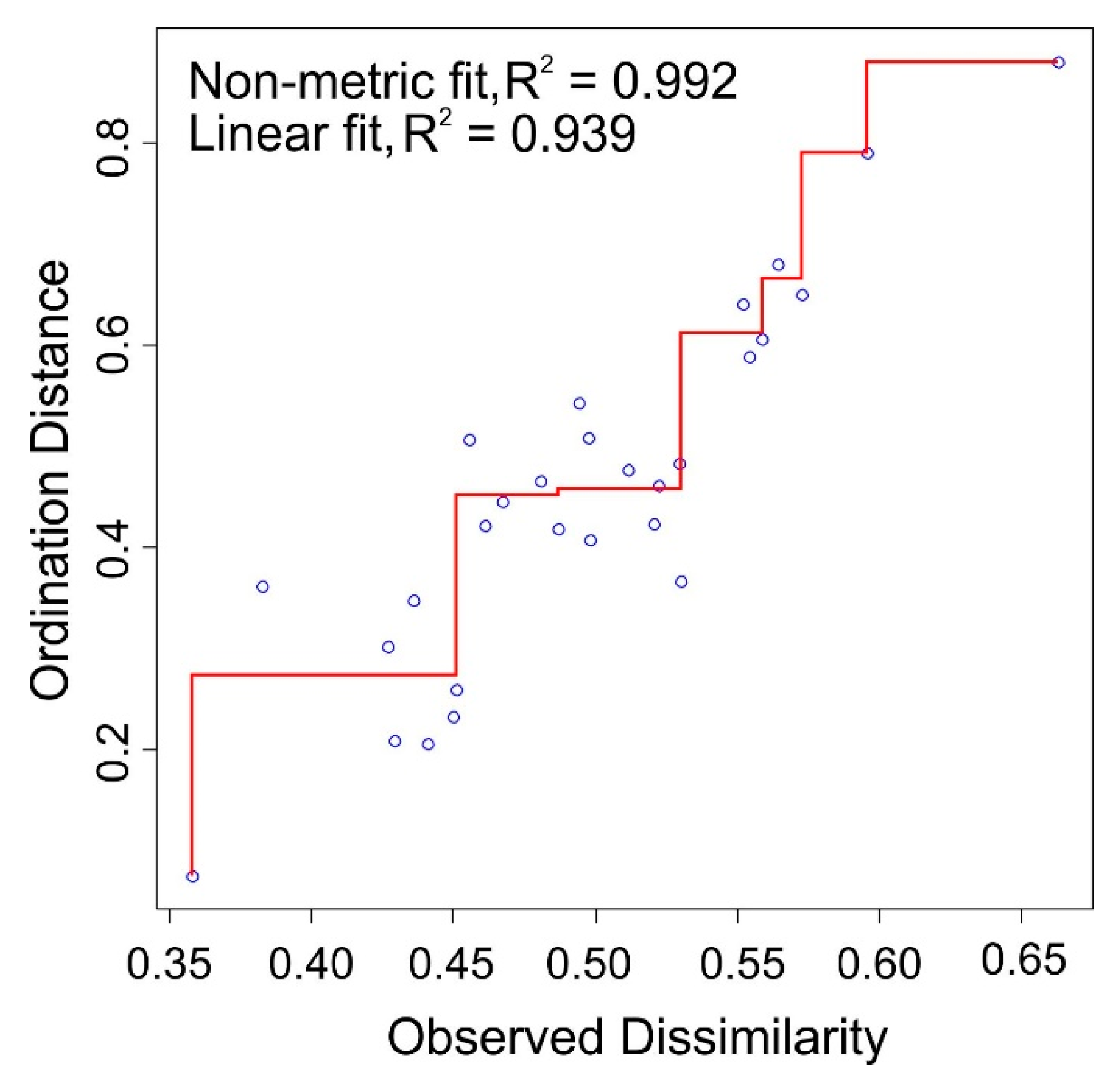
2.5. Data Analyses
3. Results
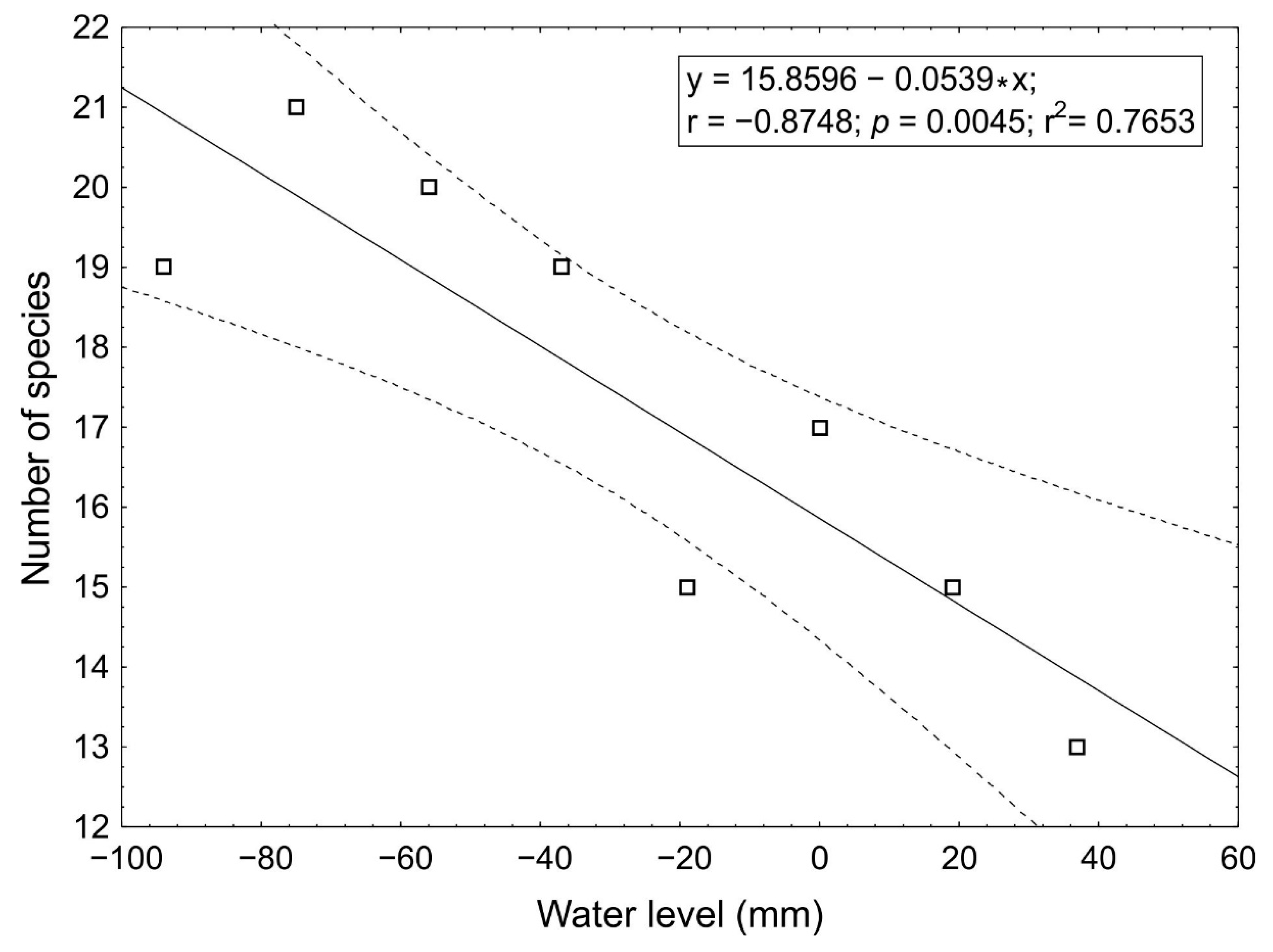
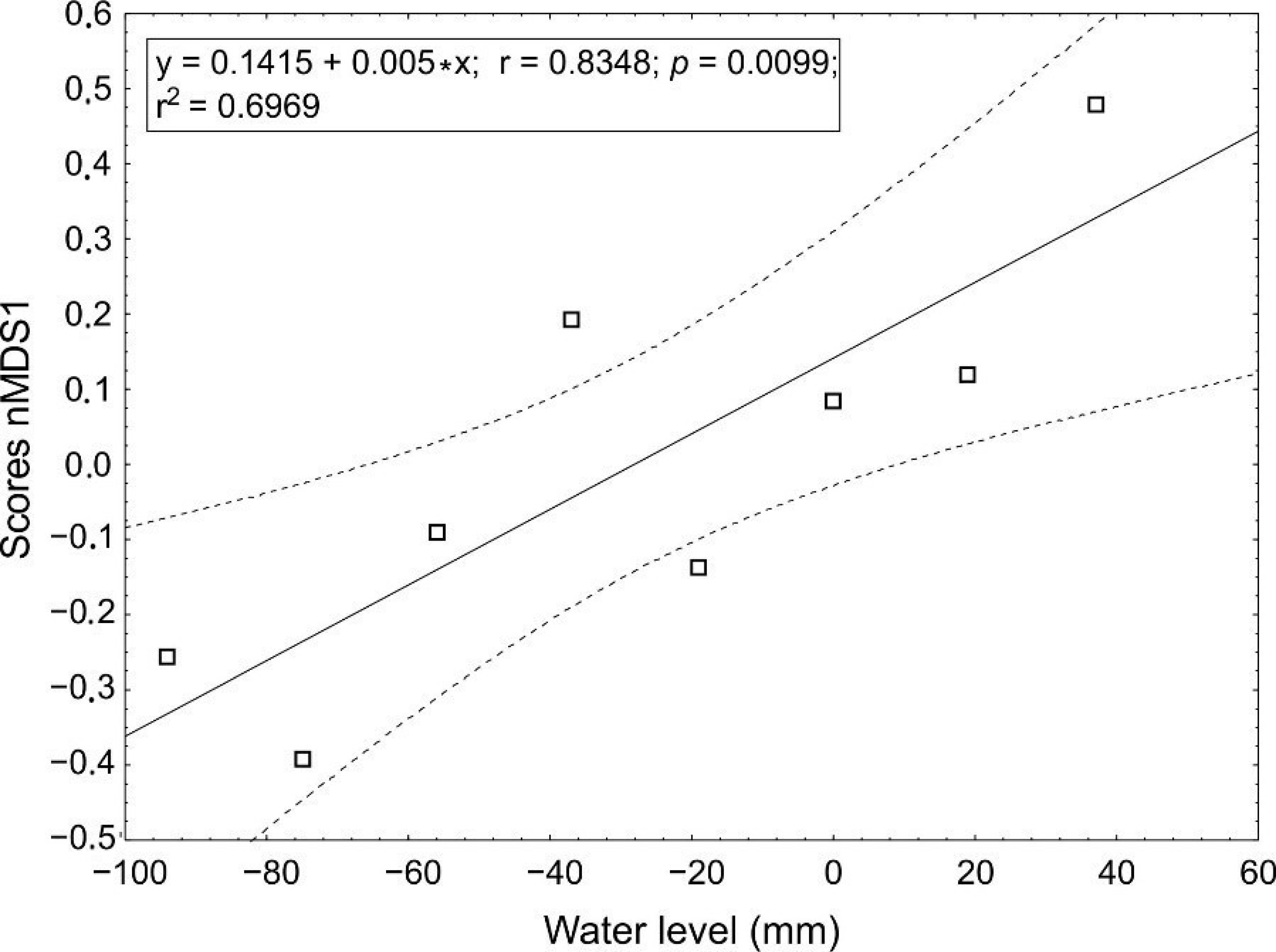
3.1. The Relationship between the Number of Species and Water Level
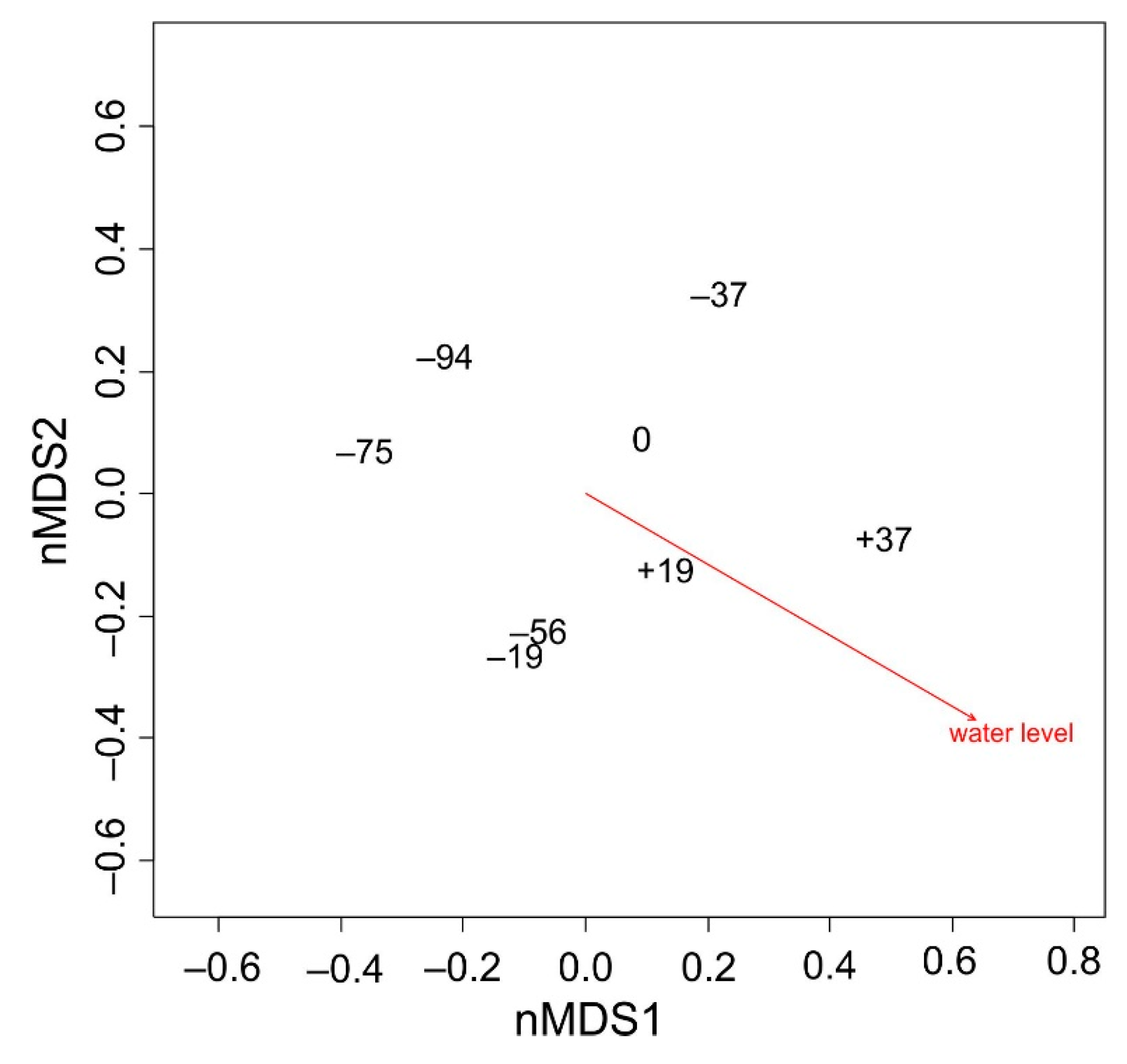
3.2. Species Composition and Water Level
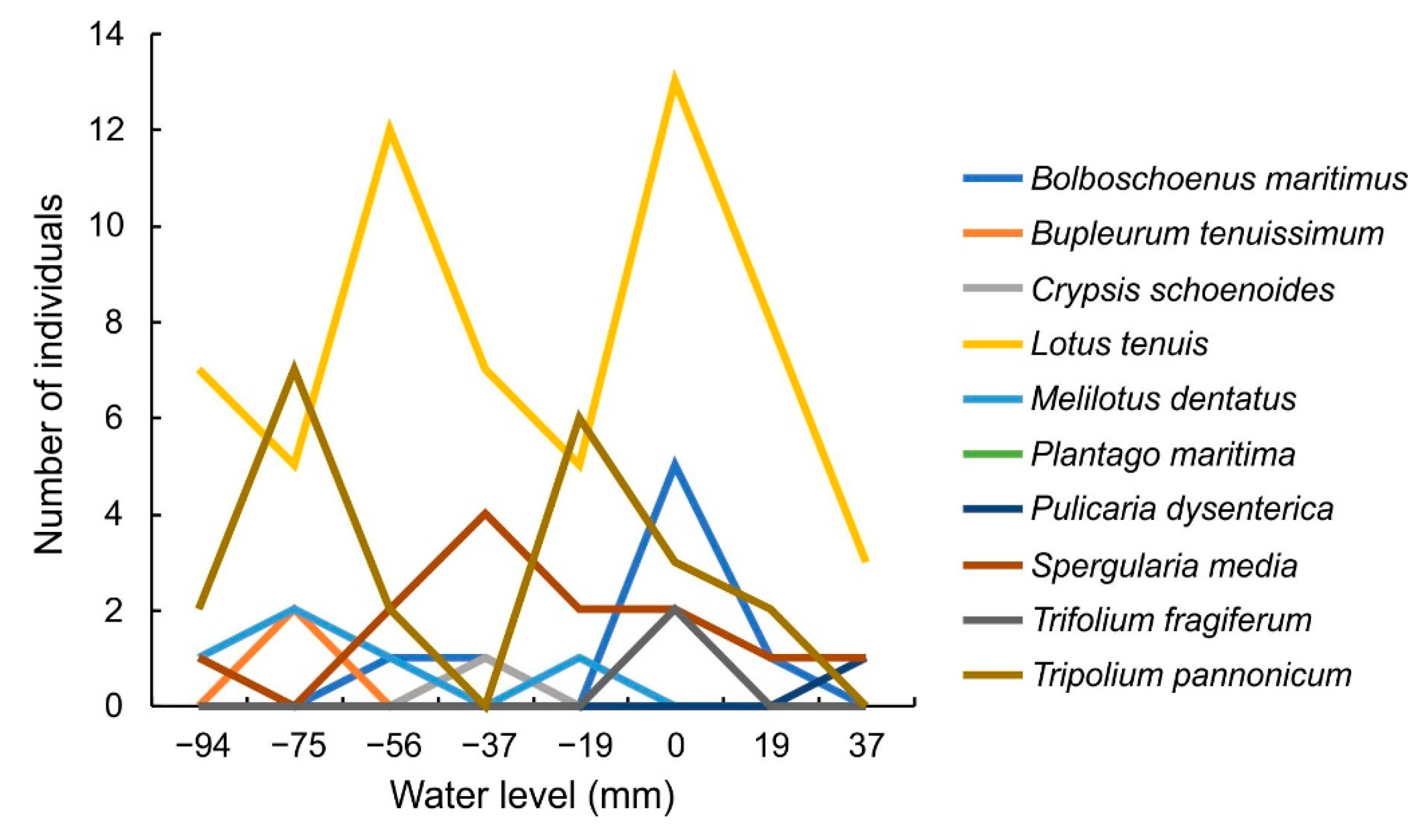
3.3. Growth of Halophytes in Water Level Belts
4. Discussion
4.1. The Impact of Water Level on the Number of Species and Their Distribution
4.2. Halophytes in Experiment
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species/Belt (mm) | −96 | −75 | −56 | −37 | −19 | 0 | +19 | +37 | Total Frequency | EIV Salinity |
|---|---|---|---|---|---|---|---|---|---|---|
| Alopecurus aequalis | 1 | 1 | 2 | 2 | ||||||
| Atriplex prostrata | 7 | 11 | 9 | 4 | 8 | 2 | 6 | 1 | 48 | 3 |
| Bolboschoenus maritimus | 1 | 1 | 5 | 1 | 8 | 5 | ||||
| Bupleurum tenuissimum | 2 | 1 | 3 | 6 | ||||||
| Cirsium vulgare | 1 | 1 | 2 | 1 | ||||||
| Centaurium pulchellum | 4 | 1 | 1 | 2 | 8 | 3 | ||||
| Cerastium brachypetalum | 1 | 1 | 2 | 0 | ||||||
| Cerastium dubium | 2 | 2 | 3 | |||||||
| Conyza canadensis | 1 | 1 | 1 | |||||||
| Epilobium hirsutum | 1 | 1 | 2 | 1 | ||||||
| Epilobium parviflorum | 1 | 1 | 1 | |||||||
| Geranium pusillum | 1 | 1 | 0 | |||||||
| Crypsis schoenoides | 1 | 1 | 9 | |||||||
| Hypericum sp. | 1 | 1 | - | |||||||
| Chenopodium album agg. | 1 | 1 | 1 | |||||||
| Inula britannica | 3 | 8 | 2 | 2 | 2 | 17 | 3 | |||
| Juncus articulatus | 3 | 6 | 9 | 3 | 5 | 8 | 6 | 4 | 44 | 1 |
| Juncus bufonius agg. | 1 | 7 | 1 | 2 | 5 | 1 | 3 | 1 | 21 | 1 |
| Juncus compressus | 41 | 45 | 20 | 35 | 36 | 27 | 38 | 20 | 262 | 3 |
| Juncus inflexus | 1 | 2 | 1 | 4 | 2 | |||||
| Lamium purpureum | 2 | 2 | 0 | |||||||
| Lotus tenuis | 7 | 5 | 12 | 7 | 5 | 13 | 8 | 3 | 60 | 6 |
| Lycopus europaeus | 2 | 2 | 1 | |||||||
| Medicago lupulina | 1 | 1 | 1 | |||||||
| Melilotus altissimus | 1 | 1 | 2 | |||||||
| Melilotus dentatus | 1 | 2 | 1 | 1 | 5 | 5 | ||||
| Mentha aquatica | 3 | 5 | 7 | 2 | 6 | 4 | 3 | 2 | 32 | 1 |
| Plantago lanceolata | 1 | 1 | 1 | |||||||
| Plantago major | 1 | 1 | 1 | |||||||
| Plantago maritima | 1 | 1 | 7 | |||||||
| Plantago uliginosa | 82 | 92 | 108 | 92 | 73 | 62 | 57 | 32 | 598 | 2 |
| Poa annua | 2 | 1 | 1 | 1 | 2 | 7 | 1 | |||
| Poaceae | 8 | 5 | 4 | 6 | 8 | 4 | 2 | 37 | - | |
| Potentilla supina | 2 | 2 | 4 | 1 | ||||||
| Pulicaria dysenterica | 1 | 1 | 4 | |||||||
| Ranunculus sceleratus | 1 | 1 | 1 | |||||||
| Typha sp. | 1 | 1 | 2 | - | ||||||
| Spergularia media | 1 | 2 | 4 | 2 | 2 | 1 | 1 | 13 | 8 | |
| Taraxacum sp. | 1 | 1 | - | |||||||
| Trifolium fragiferum | 2 | 2 | 6 | |||||||
| Tripolium pannonicum subsp. pannonicum | 2 | 7 | 2 | 6 | 3 | 2 | 22 | 8 | ||
| Veronica anagallis-aquatica | 1 | 2 | 2 | 1 | 6 | 2 | ||||
| Veronica anagalloides | 1 | 1 | 3 | |||||||
| Veronica scutellata | 1 | 1 | 0 |
Appendix B
- Pulicaria dysenterica is to be found usually on wet and mildly saline soils [57] and is known for chemicals contained in its oils [58,59]. In the Czech Republic, it is scattered only across the valleys in southern Moravia and can be rarely seen in the White Carpathians, in Haná region, or the Moravian Gate and Ostrava regions [60].
- Melilotus dentatus is species the prefers wet saline grasslands, often ruderalized [61]. In the Czech Republic, it can be found only in dry, warm areas of southern Moravia and central and north-western Bohemia. It is a species with declined occurrence due to the loss of suitable habitats in the Czech Republic [62,63].
- Bolboschoenus maritimus dominates reed vegetation of saline waters both along the seashore and inland saltwater bodies in Central Europe [64] as well as in other parts of the world [65]. In the Czech Republic, it is a rare species with declining occurrence [62] concentrated to the remains of saline wetlands in dry and warm areas of southern Moravia and central and north-western Bohemia [66].
- Lotus tenuis is European species of wet salt-rich soils and quite rare in the Czech Republic [62]. It is naturalized in other parts of the world, where it is known as an important forage crop of otherwise useless saline soils [68]. It is flood tolerant [69] and well established on restored saline wetlands [39].
- Plantago maritima is another obligate halophyte that tolerates being trodden down. However, it is often found on gradually disrupted saline soils (due to restoration) and in localities overgrown with other vegetation. Much of this species preferred habitat has disappeared from the Czech Republic, and thus its occurrence is rare and only in the north-western Bohemia and southern Moravia [71].
- Crypsis schoenoides is also an obligate halophyte, but it grows on heavily saline soils that accumulate an extreme amount of salt during the dry season. At the same time, this species is to be found in full-sun localities with wet soils rich in nutrients. In the Czech Republic, they can be found very rarely [62], only in southern Moravia [76].
References
- Šumberová, K.; Novák, J.; Sádlo, J. Slaniskové trávníky. In Vegetace České Republiky. 2, Travinná a Keříčková Vegetace, 2nd ed.; Chytrý, M., Ed.; Academia: Praha, Czech Republic, 2010; pp. 150–164. [Google Scholar]
- Sádlo, J. T7 Slaniska. In Katalog Biotopů České Republiky, 2nd ed.; Chytrý, M., Kučera, T., Kočí, M., Grulich, V., Lustyk, P., Eds.; Agentura Ochrany Přírody a Krajiny ČR: Praha, Czech Republic, 2010; pp. 240–243. [Google Scholar]
- Piernik, A. Ecological Pattern of Inland Salt Marsh Vegetation in Central Europe; Wydawnictwo Naukowe Uniwersytetu Mikołaja Kopernika: Toruń, Czech Republic, 2012; p. 219. [Google Scholar]
- Šumberová, K.; Chytrý, M.; Sádlo, J. Rákosiny a vegetace vysokých ostřic. In Katalog Biotopů České Republiky, 2nd ed.; Chytrý, M., Kučera, T., Kočí, M., Grulich, V., Lustyk, P., Eds.; Agentura Ochrany Přírody a Krajiny České Republiky: Praha, Czech Republic, 2010; pp. 34–52. [Google Scholar]
- Galvanek, D.; Dite, D.; Elias, P.; Dite, Z. Regeneration of threatened alkali steppe vegetation after a heavy disturbance by disk tillage. Plant Ecol. 2020, 221, 1177–1186. [Google Scholar] [CrossRef]
- Elias, P.; Sopotlieva, D.; Dite, D.; Hajkova, P.; Apostolova, I.; Senko, D.; Meleckova, Z.; Hajek, M. Vegetation diversity of salt-rich grasslands in Southeast Europe. Appl. Veg. Sci. 2013, 16, 521–537. [Google Scholar] [CrossRef]
- Eliáš, P.; Dítě, D.; Dítě, Z. Halophytic Vegetation in the Pannonian Basin: Origin, Syntaxonomy, Threat, and Conservation. In Handbook of Halophytes: From Molecules to Ecosystems towards Biosaline Agriculture; Springer International Publishing: Berlin/Heidelberg, Germany, 2020; pp. 1–38. [Google Scholar]
- Chytry, M. Vegetation of the Czech Republic: Diversity, ecology, history and dynamics. Preslia 2012, 84, 427–504. [Google Scholar]
- Hroudová, Z.; Hrivnák, R.; Šumberová, K. MCB01. Astero Pannonici-Bolboschoenetum Compacti. In Vegetace České Republiky. 3, Vodní a Mokřadní Vegetace, 1st ed.; Chytrý, M., Ed.; Academia: Praha, Czech Republic, 2011; pp. 434–437. [Google Scholar]
- Šumberová, K. Mokřady a pobřežní vegetace. In Červený Seznam Biotopů České Republiky; Agentura Ochrany Přírody a Krajiny ČR: Praha, Czech Republic, 2020; Volume 41, pp. 47–52. [Google Scholar]
- Chytrý, M. Sekundární trávníky a vřesoviště. In Červený Seznam Biotopů České Republiky; Agentura Ochrany Přírody a Krajiny ČR: Praha, Czech Republic, 2020; Volume 41, pp. 63–69. [Google Scholar]
- Navratilova, J.; Havlicek, M.; Navratil, J.; Frazier, R.J. Land cover changes on temperate organic substrates over last 150years: Evidence from the Czech Republic. Biologia 2019, 74, 361–373. [Google Scholar] [CrossRef]
- Hroudová, Z. Svaz, M.C.B. Meliloto Dentati-Bolboschoenion Maritimi. In Vegetace České republiky. 3, Vodní a Mokřadní Vegetace, 1st ed.; Chytrý, M., Ed.; Academia: Praha, Czech Republic, 2011; pp. 428–434. [Google Scholar]
- Ditetova, Z.; Dite, D.; Elias, P.; Galvanek, D. The impact of grazing absence in inland saline vegetation—A case study from Slovakia. Biologia 2016, 71, 980–988. [Google Scholar] [CrossRef]
- Šumberová, K. Vegetace jednoletých sukulentních halofytů. In Vegetace České Republiky. 2, Travinná a Keříčková Vegetace, 2nd ed.; Chytrý, M., Ed.; Academia: Praha, Czech Republic, 2010; pp. 143–149. [Google Scholar]
- Boon, P.J.; Baxter, J.M. Aquatic conservation: Reflections on the first 25 years. Aquat. Conserv. Mar. Freshw. Ecosyst. 2016, 26, 809–816. [Google Scholar] [CrossRef]
- Kaplan, Z.; Sumberova, K.; Formanova, I.; Duchacek, M. Re-establishment of an extinct population of the endangered aquatic plant Potamogeton coloratus. Aquat. Bot. 2014, 119, 91–99. [Google Scholar] [CrossRef]
- Schaich, H.; Barthelmes, B. Management of grasslands in rewetted floodplains: Effects of grazing and cutting on vegetation development. Tuexenia 2012, 32, 207–231. [Google Scholar]
- Klotzli, F.; Grootjans, A.P. Restoration of natural and semi-natural wetland systems in Central Europe: Progress and predictability of developments. Restor. Ecol. 2001, 9, 209–219. [Google Scholar] [CrossRef]
- Williams, P.; Biggs, J.; Stoate, C.; Szczur, J.; Brown, C.; Bonney, S. Nature based measures increase freshwater biodiversity in agricultural catchments. Biol. Conserv. 2020, 244. [Google Scholar] [CrossRef]
- Jellinek, S.; Harrison, P.A.; Tuck, J.; Te, T. Replanting agricultural landscapes: How well do plants survive after habitat restoration? Restor. Ecol. 2020, 28, 1454–1463. [Google Scholar] [CrossRef]
- Van der Horst, D.; Martinat, S.; Navratil, J.; Dvorak, P.; Chmielova, P. What can the location of biogas plants tell us about agricultural change? A Case Study from the Czech Republic. Deturope Cent. Eur. J. Reg. Dev. Tour. 2018, 10, 33–52. [Google Scholar]
- Alonso, N.M.; Muniz, I.O.; Aja, A.H.; Garcia, F.F. Challenges for the revitalisation of peri-urban agriculture in Spain: Territorial analysis of the Madrid and Oviedo metropolitan areas. Morav. Geogr. Rep. 2017, 25, 192–207. [Google Scholar] [CrossRef] [Green Version]
- Chodkowska-Miszczuk, J.; Kulla, M.; Novotny, L. Biogas Energy—A Chance for Agriculture and Rural Development? Insight from the Post-Communist Central Europe. Deturope Cent. Eur. J. Reg. Dev. Tour. 2019, 11, 30–53. [Google Scholar]
- Hildebrandt, U.; Janetta, K.; Ouziad, F.; Renne, B.; Nawrath, K.; Bothe, H. Arbuscular mycorrhizal colonization of halophytes in Central European salt marshes. Mycorrhiza 2001, 10, 175–183. [Google Scholar] [CrossRef]
- Muhsin, T.M.; Booth, T. Fungi Associated with Halophytes of an Inland Salt-Marsh, Manitoba, Canada. Can. J. Bot. 1987, 65, 1137–1151. [Google Scholar] [CrossRef]
- Sychra, J.; Adamek, Z.; Petrivalska, K. Distribution and diversity of littoral macroinvertebrates within extensive reed beds of a lowland pond. Ann. Limnol. Int. J. Limnol. 2010, 46, 281–289. [Google Scholar] [CrossRef]
- Beyen, W.; Meire, P. Ecohydrology of saline grasslands: Consequences for their restoration. Appl. Veg. Sci. 2003, 6, 153–160. [Google Scholar] [CrossRef]
- Miklin, J.; Machacek, P. Birds of Lednicke rybniky Fishponds (Czech Republic). J. Maps 2016, 12, 239–248. [Google Scholar] [CrossRef]
- Mitsch, W.J.; Wu, X.; Nairn, R.W.; Weihe, P.E.; Wang, N.; Deal, R.; Boucher, C.E. Creating and restoring wetlands. BioScience 1998, 48, 1019–1030. [Google Scholar] [CrossRef]
- Zedler, J.B. Progress in wetland restoration ecology. Trends Ecol. Evol. 2000, 15, 402–407. [Google Scholar] [CrossRef]
- Wang, G.M.; Lv, J.Z.; Han, G.X.; Zhu, S.Y.; Liu, X.L.; Wang, A.D.; Guan, B.; Zhao, Y.J. Ecological Restoration of Degraded Supratidal Wetland Based on Microtopography Modification: A Case Study in the Yellow River Delta. Wetlands 2020. [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M. Seeds: Ecology, Biogeography, and Evolution of Dormancy and Germination; Academic Press: Cambridge, MA, USA, 1998. [Google Scholar]
- Wolters, M.; de Vries, S.; Ozinga, W.A.; Bakker, J.P. Restoration of inland brackish vegetation by large-scale transfer of coastal driftline material. Appl. Veg. Sci. 2017, 20, 641–650. [Google Scholar] [CrossRef]
- Gilhaus, K.; Vogt, V.; Holzel, N. Restoration of sand grasslands by topsoil removal and self-greening. Appl. Veg. Sci. 2015, 18, 661–673. [Google Scholar] [CrossRef]
- Meleckova, Z.; Galvanek, D.; Dite, D.; Elias, P. Effect of experimental top soil removal on vegetation of Pannonian salt steppes. Cent. Eur. J. Biol. 2013, 8, 1204–1215. [Google Scholar] [CrossRef]
- Bakker, J.P.; Esselink, P.; Dijkema, K.S.; Van Duin, W.E.; De Jong, D.J. Restoration of salt marshes in the Netherlands. Hydrobiologia 2002, 478, 29–51. [Google Scholar] [CrossRef]
- Šumberová, K.; Chytrý, M. Vegetace jednoletých vlhkomilných bylin. In Katalog Biotopů České Republiky, 2nd ed.; Chytrý, M., Kučera, T., Kočí, M., Grulich, V., Lustyk, P., Eds.; Agentura Ochrany Přírody a Krajiny České Republiky: Praha, Czech Republic, 2010; pp. 54–62. [Google Scholar]
- Meleckova, Z.; Dite, D.; Elias, P.; Pis, V.; Galvanek, D. Succession of saline vegetation in Slovakia after a large-scale disturbance. Ann. Bot. Fenn. 2014, 51, 285–296. [Google Scholar] [CrossRef]
- Bakker, J.P.; Berendse, F. Constraints in the restoration of ecological diversity in grassland and heathland communities. Trends Ecol. Evol. 1999, 14, 63–68. [Google Scholar] [CrossRef]
- Shannon, M.C.; Grieve, C.M.; Francois, L.E. Whole—Plant Response to Salinity. In Plant—Environment Interactions; Wilkinson, R.E., Ed.; Marcel Dekker Inc.: New York, NY, USA, 1994; pp. 199–244. [Google Scholar]
- Al-Hawija, B.N.; Partzsch, M.; Hensen, I. Effects of temperature, salinity and cold stratification on seed germination in halophytes. Nord. J. Bot. 2012, 30, 627–634. [Google Scholar] [CrossRef]
- Grulich, V.; Danihelka, J. NPR Slanisko u Nesytu. In Chráněná Území Přírody ČR, Svazek IX. Brněnsko; Mackovcin, P., Ed.; AOPK ČR, EkoCentrum Brno: Praha, Czech Republic, 2007; pp. 766–767. [Google Scholar]
- Oksanen, J.; Guillaume Blanchet, F.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Package ‘Vegan’. Available online: https://cran.r-project.org/web/packages/vegan/vegan.pdf (accessed on 12 November 2020).
- The-R-Foundation. The R Project for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 12 November 2020).
- TIBCO. TIBCO Statistica™ Quick Reference. Available online: https://docs.tibco.com/pub/stat/13.3.0/doc/pdf/TIB_stat_13.3_quick_ref.pdf (accessed on 12 November 2020).
- Keiffer, C.H.; Ungar, I.A. The effect of extended exposure to hypersaline conditions on the germination of five inland halophyte species. Am. J. Bot. 1997, 84, 104–111. [Google Scholar] [CrossRef]
- Ungar, I.A. Germination ecology of halophytes. In Contributions to the Ecology of Halophytes; Sen, D.N., Rajpurohit, K.S., Eds.; Springer: Dordrecht, The Netherlands, 1982; Volumes 143–154, p. 272. [Google Scholar]
- Elsey-Quirk, T.; Middleton, B.A.; Proffitt, C.E. Seed flotation and germination of salt marsh plants: The effects of stratification, salinity, and/or inundation regime. Aquat. Bot. 2009, 91, 402009. [Google Scholar] [CrossRef]
- Wang, M.; Wang, G.D.; Lu, X.G.; Jiang, M.; Wang, S.Z. Soil seed banks and their implications for wetland restoration along the Nongjiang River, Northeastern China. Ecol. Eng. 2016, 96, 26–33. [Google Scholar] [CrossRef]
- Magee, T.K.; Kentula, M.E. Response of wetland plant species to hydrologic conditions. Wetl. Ecol. Manag. 2005, 13, 163–181. [Google Scholar] [CrossRef]
- Wang, Z.; Xu, H.; Yin, L.; Li, J.; Zhang, Z.; Li, Y. Effects of water treatments on the activation of soil seed banks-A case study on the lower reaches of the Tarim River. Prog. Nat. Sci. 2009, 19, 733–740. [Google Scholar] [CrossRef]
- Chytry, M.; Tichy, L.; Drevojan, P.; Sadlo, J.; Zeleny, D. Ellenberg-type indicator values for the Czech flora. Preslia 2018, 90, 83–103. [Google Scholar] [CrossRef] [Green Version]
- Zeleny, D.; Chytry, M. Ecological Specialization Indices for species of the Czech flora. Preslia 2019, 91, 93–116. [Google Scholar] [CrossRef]
- Deák, B.; Valkó, O.; Török, P.; Tóthmérész, B. Solonetz meadow vegetation (Beckmannion eruciformis) in East-Hungary—An alliance driven by moisture and salinity. Tuexenia 2014, 34, 187–203. [Google Scholar] [CrossRef]
- Can, E.; Arslan, M.; Sener, O.; Daghan, H. Response of strawberry clover (Trifolium fragiferum L.) to salinity stress. Res. Crops 2013, 14, 576–584. [Google Scholar]
- Kaplan, Z.; Danihelka, J.; Sumberova, K.; Chrtek, J.; Rotreklova, O.; Ekrt, L.; Stepankova, J.; Taraska, V.; Travnicek, B.; Prancl, J.; et al. Distributions of vascular plants in the Czech Republic. Part 5. Preslia 2017, 89, 333–439. [Google Scholar] [CrossRef]
- Basta, A.; Tzakou, O.; Couladis, M.; Pavlovic, M. Chemical composition of Pulicaria dysenterica (L.) Bernh. from Greece. J. Essent. Oil Res. 2007, 19, 333–335. [Google Scholar] [CrossRef]
- Cdiz-Gurrea, M.D.; Zengin, G.; Kayacik, O.; Lobine, D.; Mahomoodally, M.F.; Leyva-Jimnez, F.J.; Segura-Carretero, A. Innovative perspectives on Pulicaria dysenterica extracts: Phyto-pharmaceutical properties, chemical characterization and multivariate analysis. J. Sci. Food Agric. 2019, 99, 6001–6010. [Google Scholar] [CrossRef] [PubMed]
- Hrouda, L. Pulicaria Gaertner. In Květena České Republiky 7; Slavík, B., Štěpánková, J., Eds.; Academia: Praha, Czech Republic, 2004. [Google Scholar]
- Tishchenko, M.P.; Korolyuk, A.Y. The syntaxonomy of the meadow vegetation of Kulunda and Kasmala Pine Forest Strips (Altai Territory). Rastit. Ross. 2018, 2018, 101–119. [Google Scholar] [CrossRef]
- Danihelka, J.; Chrtek, J.; Kaplan, Z. Checklist of vascular plants of the Czech Republic. Preslia 2012, 84, 647–811. [Google Scholar]
- Chytry, M.; Hajek, M.; Koci, M.; Pesout, P.; Rolecek, J.; Sadlo, J.; Sumberova, K.; Sychra, J.; Boublik, K.; Douda, J.; et al. Red List of Habitats of the Czech Republic. Ecol. Indic. 2019, 106. [Google Scholar] [CrossRef]
- Ljevnaic-Masic, B.; Dzigurski, D.; Nikolic, L.; Brdar-Jokanovic, M.; Cabilovski, R.; Ciric, V.; Petrovic, A. Assessment of the habitat conditions of a rare and endangered inland saline wetland community with Bolboschoenus maritimus (L.) Palla dominance in Southeastern Europe: The effects of physical-chemical water and soil properties. Wetl. Ecol. Manag. 2020, 28, 421–438. [Google Scholar] [CrossRef]
- Kettenring, K.M. Viability, dormancy, germination, and intraspecific variation of Bolboschoenus maritimus (alkali bulrush) seeds. Aquat. Bot. 2016, 134, 26–30. [Google Scholar] [CrossRef]
- Hroudova, Z.; Zakravsky, P.; Duchacek, M.; Marhold, K. Taxonomy, distribution and ecology of Bolboschoenus in Europe. Ann. Bot. Fenn. 2007, 44, 81–102. [Google Scholar]
- Šourková, M.; Hrouda, L.; Bupleurum, L. Květena České Republiky 5; Slavík, B., Ed.; Academia: Praha, Czech Republic, 1997. [Google Scholar]
- Espasandin, F.D.; Brugnoli, E.A.; Ayala, P.G.; Ayala, L.P.; Ruiz, O.A.; Sansberro, P.A. Long-term preservation of Lotus tenuis adventitious buds. Plant Cell Tissue Organ Cult. 2019, 136, 373–382. [Google Scholar] [CrossRef]
- Striker, G.G.; Colmer, T.D. Flooding tolerance of forage legumes. J. Exp. Bot. 2017, 68, 1851–1872. [Google Scholar] [CrossRef] [Green Version]
- Haerinasab, M.; Ali-Farsangi, F.; Bordbar, F.; Farouji, A.E. Genetic Diversity and Infraspecific Relationships of Trifolium fragiferum L. in Iran. Iran. J. Sci. Technol. Trans. A Sci. 2020, 44, 345–354. [Google Scholar] [CrossRef]
- Chrtek, J.S.; Plantago, L. Květena České Republiky 6; Slavík, B., Ed.; Academia: Praha, Czech Republic, 2000. [Google Scholar]
- Kaplan, Z.; Danihelka, J. Květena České Republiky 2; Hejný, S., Slavík, B., Eds.; Academia: Praha, Czech Republic, 1990. [Google Scholar]
- Kaplan, Z.; Danihelka, J.; Stepankova, J.; Ekrt, L.; Chrtek, J.; Zazvorka, J.; Grulich, V.; Repka, R.; Prancl, J.; Duchacek, M.; et al. Distributions of vascular plants in the Czech Republic. Part 2. Preslia 2016, 88, 229–322. [Google Scholar]
- Eliáš, P.; Dítě, D.; Dítě, Z.; Eliášová, M. Distribution and habitat preferences of Tripolium pannonicum subsp. pannonicum (Asteraceae) in Slovakia. Thaiszia J. Bot. 2018, 28, 111–123. [Google Scholar]
- Kovanda, K.; Kubát, K.; Aster, L. Květena České Republiky 7; Slavík, B., Štěpánková, J., Eds.; Academia: Praha, Czech Republic, 2004. [Google Scholar]
- Kaplan, Z.; Danihelka, J.; Ekrt, L.; Stech, M.; Repka, R.; Chrtek, J.; Grulich, V.; Rotreklova, O.; Drevojan, P.; Sumberova, K.; et al. Distributions of vascular plants in the Czech Republic. Part 9. Preslia 2020, 92, 255–340. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Čížková, E.; Navrátilová, J.; Martinát, S.; Navrátil, J.; Frazier, R.J. Impact of Water Level on Species Quantity and Composition Grown from the Soil Seed Bank of the Inland Salt Marsh: An Ex-Situ Experiment. Land 2020, 9, 533. https://doi.org/10.3390/land9120533
Čížková E, Navrátilová J, Martinát S, Navrátil J, Frazier RJ. Impact of Water Level on Species Quantity and Composition Grown from the Soil Seed Bank of the Inland Salt Marsh: An Ex-Situ Experiment. Land. 2020; 9(12):533. https://doi.org/10.3390/land9120533
Chicago/Turabian StyleČížková, Eva, Jana Navrátilová, Stanislav Martinát, Josef Navrátil, and Ryan J. Frazier. 2020. "Impact of Water Level on Species Quantity and Composition Grown from the Soil Seed Bank of the Inland Salt Marsh: An Ex-Situ Experiment" Land 9, no. 12: 533. https://doi.org/10.3390/land9120533
APA StyleČížková, E., Navrátilová, J., Martinát, S., Navrátil, J., & Frazier, R. J. (2020). Impact of Water Level on Species Quantity and Composition Grown from the Soil Seed Bank of the Inland Salt Marsh: An Ex-Situ Experiment. Land, 9(12), 533. https://doi.org/10.3390/land9120533