Connectivity of Protected Areas: Effect of Human Pressure and Subnational Contributions in the Ecoregions of Tropical Andean Countries

 ,
,  ,
,  , ,
, , 
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Geographical Scope
2.2. Database of Protected Areas
2.3. Ecological Units of Analysis
2.4. Network Connectivity and Landscape Heterogeneity
2.5. Data Analyses
3. Results
3.1. Database of Protected Areas
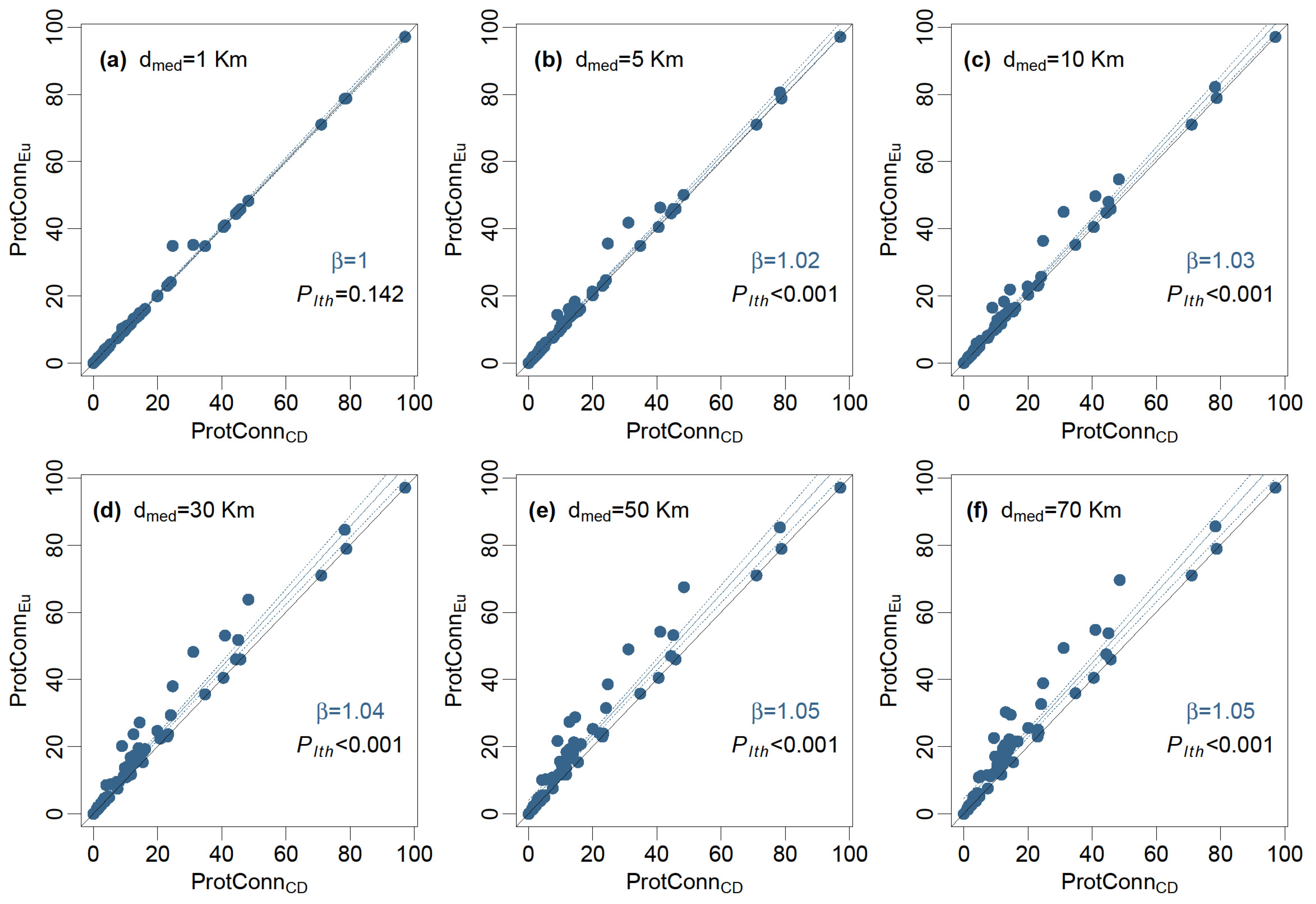
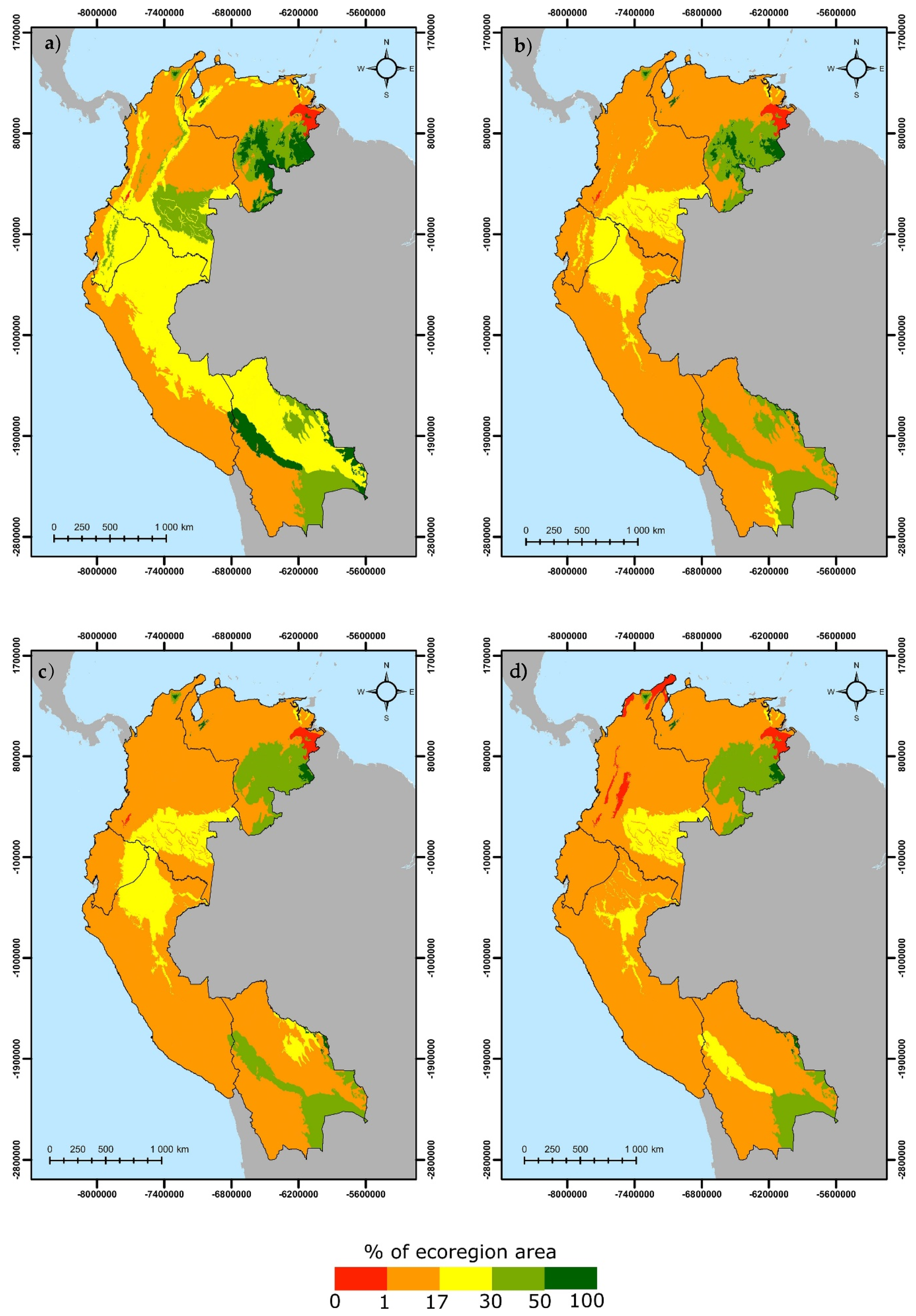
3.2. Landscape Heterogeneity and PA Connectivity
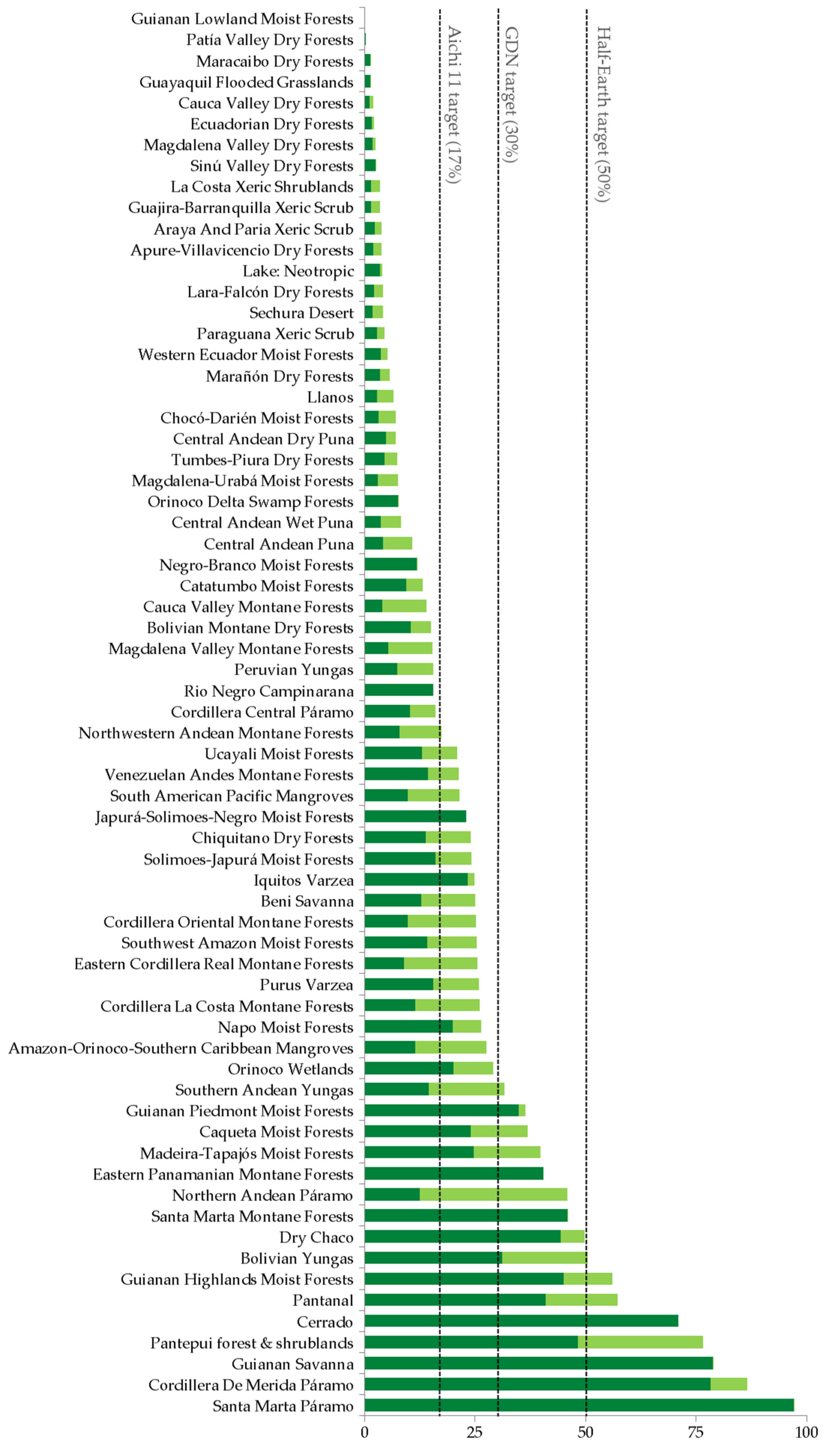
3.3. Protected and Connected Ecoregions and Global Targets
4. Discussion
4.1. Protected Land and Governmental Reports
4.2. Landscape Heterogeneity and PA Connectivity
4.3. Protected and Connected Ecoregions
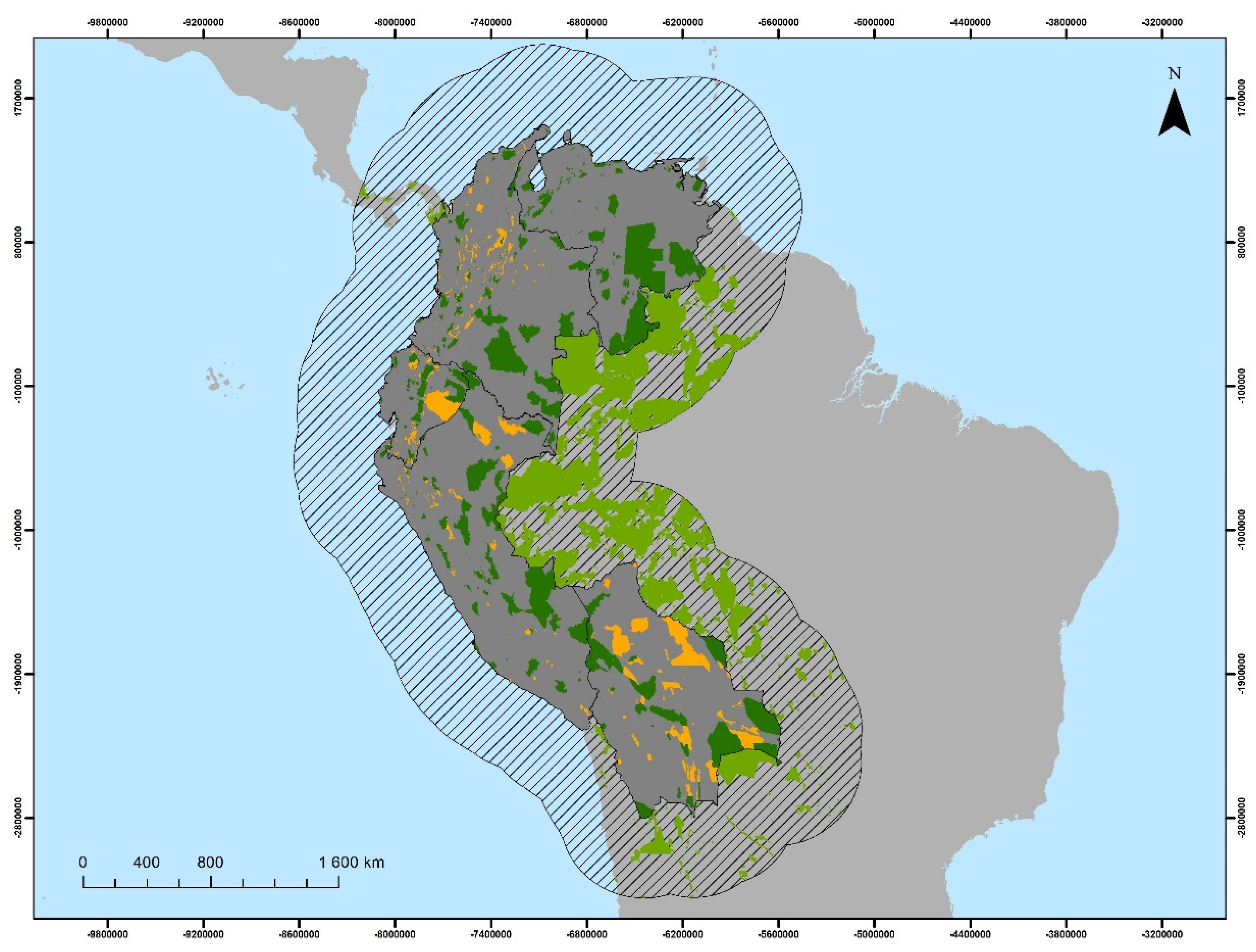
4.4. Subnational Contributions
4.5. Limitations and Further Research
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

References
- Locke, H.; Ellis, E.C.; Venter, O.; Schuster, R.; Ma, K.; Shen, X.; Woodley, S.; Kingston, N.; Bhola, N.; Strassburg, B.B.N.; et al. Three global conditions for biodiversity conservation and sustainable use: An implementation framework. Natl. Sci. Rev. 2019, 6, 1080–1082. [Google Scholar] [CrossRef] [Green Version]
- Haddad, N.M.; Brudvig, L.A.; Clobert, J.; Davies, K.F.; Gonzalez, A.; Holt, R.D.; Lovejoy, T.E.; Sexton, J.O.; Austin, M.P.; Collins, C.D.; et al. Habitat fragmentation and its lasting impact on Earth’s ecosystems. Sci. Adv. 2015, 1, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zemanova, M.A.; Perotto-Baldivieso, H.L.; Dickins, E.L.; Gill, A.B.; Leonard, J.P.; Wester, D.B. Impact of deforestation on habitat connectivity thresholds for large carnivores in tropical forests. Ecol. Process. 2017, 6, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Fahrig, L. Habitat fragmentation: A long and tangled tale. Glob. Ecol. Biogeogr. 2019, 28, 33–41. [Google Scholar] [CrossRef]
- Fletcher, R.J.; Didham, R.K.; Banks-Leite, C.; Barlow, J.; Ewers, R.M.; Rosindell, J.; Holt, R.D.; Gonzalez, A.; Pardini, R.; Damschen, E.I.; et al. Is habitat fragmentation good for biodiversity? Biol. Conserv. 2018, 226, 9–15. [Google Scholar] [CrossRef] [Green Version]
- Crooks, K.R.; Burdett, C.L.; Theobald, D.M.; Rondinini, C.; Boitani, L. Global patterns of fragmentation and connectivity of mammalian carnivore habitat. Philos. Trans. R. Soc. B Biol. Sci. 2011, 366, 2642–2651. [Google Scholar] [CrossRef]
- DeFries, R.; Hansen, A.; Newton, A.C.; Hansen, M.C. Increasing isolation of protected areas in tropical forests over the past twenty years. Ecol. Appl. 2005, 15, 19–26. [Google Scholar] [CrossRef] [Green Version]
- Laurance, W.F.; Carolina Useche, D.; Rendeiro, J.; Kalka, M.; Bradshaw, C.J.A.; Sloan, S.P.; Laurance, S.G.; Campbell, M.; Abernethy, K.; Alvarez, P.; et al. Averting biodiversity collapse in tropical forest protected areas. Nature 2012, 489, 290–293. [Google Scholar] [CrossRef] [Green Version]
- UNEP-WCMC; IUCN; NGS. Protected Planet Report 2018; Gland: Cambridge, UK; Washington, DC, USA, 2018. [Google Scholar]
- CBD. Strategic Plan for Biodiversity 2011–2020; CBD: Montreal, QC, Canada, 2011. [Google Scholar]
- Watson, J.E.M.; Dudley, N.; Segan, D.B.; Hockings, M. The performance and potential of protected areas. Nature 2014, 515, 67–73. [Google Scholar] [CrossRef]
- Coetzee, B.W.T.; Gaston, K.J.; Chown, S.L. Local scale comparisons of biodiversity as a test for global protected area ecological performance: A meta-analysis. PLoS ONE 2014, 9, e0105824. [Google Scholar] [CrossRef] [PubMed]
- Gray, C.L.; Hill, S.L.L.; Newbold, T.; Hudson, L.N.; Börger, L.; Contu, S.; Hoskins, A.J.; Ferrier, S.; Purvis, A.; Scharlemann, J.P.W. Local biodiversity is higher inside than outside terrestrial protected areas worldwide. Nat. Commun. 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Dinerstein, E.; Vynne, C.; Sala, E.; Joshi, A.R.; Fernando, S.; Lovejoy, T.E.; Mayorga, J.; Olson, D.; Asner, G.P.; Baillie, J.E.M.; et al. A Global Deal for Nature: Guiding principles, milestones, and targets. Sci. Adv. 2019, 5, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Wilson, E.O. Half-Earth: Our Planet’s Fight for Life; Liveright: New York, NY, USA, 2016; ISBN 9781631490828. [Google Scholar]
- Locke, H. Nature Needs (at least) Half: A Necessary New Agenda for Protected Areas. In Protecting the Wild; Wuerthner, G., Crist, E.B.T., Eds.; Island Press: Washington, DC, USA, 2015; pp. 3–15. ISBN 978-1-59726-111-1. [Google Scholar]
- Dinerstein, E.; Olson, D.; Joshi, A.; Vynne, C.; Burgess, N.D.; Wikramanayake, E.; Hahn, N.; Palminteri, S.; Hedao, P.; Noss, R.; et al. An Ecoregion-Based Approach to Protecting Half the Terrestrial Realm. Bioscience 2017, 67, 534–545. [Google Scholar] [CrossRef] [PubMed]
- Barnes, M.D.; Glew, L.; Wyborn, C.; Craigie, I.D. Prevent perverse outcomes from global protected area policy. Nat. Ecol. Evol. 2018, 2, 759–762. [Google Scholar] [CrossRef] [PubMed]
- Visconti, P.; Butchart, S.H.M.; Brooks, T.M.; Langhammer, P.F.; Marnewick, D.; Vergara, S.; Yanosky, A.; Watson, J.E.M. Protected area targets post-2020. Science 2019, 364, 239–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geldmann, J.; Barnes, M.; Coad, L.; Craigie, I.D.; Hockings, M.; Burgess, N.D. Effectiveness of terrestrial protected areas in reducing habitat loss and population declines. Biol. Conserv. 2013, 161, 230–238. [Google Scholar] [CrossRef]
- Geldmann, J.; Coad, L.; Barnes, M.D.; Craigie, I.D.; Woodley, S.; Balmford, A.; Brooks, T.M.; Hockings, M.; Knights, K.; Mascia, M.B.; et al. A global analysis of management capacity and ecological outcomes in terrestrial protected areas. Conserv. Lett. 2018, 11, 1–10. [Google Scholar] [CrossRef]
- Santini, L.; Saura, S.; Rondinini, C. Connectivity of the global network of protected areas. Divers. Distrib. 2015, 199–211. [Google Scholar] [CrossRef] [Green Version]
- Saura, S.; Estreguil, C.; Mouton, C.; Rodríguez-Freire, M. Network analysis to assess landscape connectivity trends: Application to European forests (1990–2000). Ecol. Indic. 2011, 11, 407–416. [Google Scholar] [CrossRef]
- Saura, S.; Bastin, L.; Battistella, L.; Mandrici, A.; Dubois, G. Protected areas in the world’s ecoregions: How well connected are they? Ecol. Indic. 2017, 76, 144–158. [Google Scholar] [CrossRef]
- Saura, S.; Bertzky, B.; Bastin, L.; Battistella, L.; Mandrici, A.; Dubois, G. Protected area connectivity: Shortfalls in global targets and country-level priorities. Biol. Conserv. 2018, 219, 53–67. [Google Scholar] [CrossRef] [PubMed]
- Saura, S.; Bertzky, B.; Bastin, L.; Battistella, L.; Mandrici, A.; Dubois, G. Global trends in protected area connectivity from 2010 to 2018. Biol. Conserv. 2019, 238, 108183. [Google Scholar] [CrossRef] [PubMed]
- Sayre, R.; Karagulle, D.; Frye, C.; Boucher, T.; Wolff, N.H.; Breyer, S.; Wright, D.; Martin, M.; Butler, K.; Van Graafeiland, K.; et al. An assessment of the representation of ecosystems in global protected areas using new maps of World Climate Regions and World Ecosystems. Glob. Ecol. Conserv. 2020, 21, e00860. [Google Scholar] [CrossRef]
- Naidoo, R.; Brennan, A. Connectivity of protected areas must consider landscape heterogeneity: A response to Saura et al. Biol. Conserv. 2019, 239, 108316. [Google Scholar] [CrossRef]
- Visconti, P.; Di Marco, M.; Álvarez-Romero, J.G.; Januchowski-Hartley, S.R.; Pressey, R.L.; Weeks, R.; Rondinini, C. Effects of errors and gaps in spatial data sets on assessment of conservation progress. Conserv. Biol. 2013, 27, 1000–1010. [Google Scholar] [CrossRef]
- Stolton, S.; Redford, K.H.; Dudley, N. The Futures of Privately Protected Areas; UICN: Gland, Switzerland, 2014; ISBN 978-2-8317-1675. [Google Scholar]
- You, Z.; Hu, J.; Wei, Q.; Li, C.; Deng, X.; Jiang, Z. Pitfall of big databases. Proc. Natl. Acad. Sci. USA 2018, 115, E9026–E9028. [Google Scholar] [CrossRef] [Green Version]
- Bargelt, L.; Fortin, M.J.; Murray, D.L. Assessing connectivity and the contribution of private lands to protected area networks in the United States. PLoS ONE 2020, 15, e0228946. [Google Scholar] [CrossRef] [Green Version]
- Baldwin, R.F.; Fouch, N.T. Understanding the biodiversity contributions of small protected areas presents many challenges. Land 2018, 7, 123. [Google Scholar] [CrossRef] [Green Version]
- FAO. GeoNetwork: Global Administrative Unit Layers. Available online: http://www.fao.org/geonetwork/srv/en/metadata.show?id=12691 (accessed on 20 November 2019).
- Butler, R.A. The Top 10 Most Biodiverse Countries—What Are the World’s Most Biodiverse Countries? Available online: https://news.mongabay.com/2016/05/top-10-biodiverse-countries/ (accessed on 29 April 2020).
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Fonseca, G.A.B.; da Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef]
- Dudley, N. Directrices Para la Aplicación de las Categorías de Gestión de Áreas Protegidas; UICN: Gland, Switzerland, 2008; ISBN 978-2-8317-1088-4. [Google Scholar]
- Olson, D.M.; Dinerstein, E.; Wikramanayake, E.D.; Burgess, N.D.; Powell, G.V.N.; Underwood, E.C.; D’amico, J.A.; Itoua, I.; Strand, H.E.; Morrison, J.C.; et al. Terrestrial Ecoregions of the World: A New Map of Life on Earth. Bioscience 2001, 51, 933. [Google Scholar] [CrossRef]
- Sierra, R. A transnational perspective on national protected areas and ecoregions in the Tropical Andean Countries. In Globalization and New Geographies of Conservation; Zimmerer, K.S., Ed.; The University of Chicago Press: Chicago, IL, USA; London, UK, 2006. [Google Scholar]
- Shanee, S.; Shanee, N.; Monteferri, B.; Allgas, N.; Alarcon Pardo, A.; Horwich, R.H. Protected area coverage of threatened vertebrates and ecoregions in Peru: Comparison of communal, private and state reserves. J. Environ. Manag. 2017, 202, 12–20. [Google Scholar] [CrossRef] [Green Version]
- Butchart, S.H.M.; Clarke, M.; Smith, R.J.; Sykes, R.E.; Scharlemann, J.P.W.; Harfoot, M.; Buchanan, G.M.; Angulo, A.; Balmford, A.; Bertzky, B.; et al. Shortfalls and Solutions for Meeting National and Global Conservation Area Targets. Conserv. Lett. 2015, 8, 329–337. [Google Scholar] [CrossRef]
- Saura, S.; Pascual-Hortal, L. A new habitat availability index to integrate connectivity in landscape conservation planning: Comparison with existing indices and application to a case study. Landsc. Urban Plan. 2007, 83, 91–103. [Google Scholar] [CrossRef]
- Golden Kroner, R.E.; Qin, S.; Cook, C.N.; Krithivasan, R.; Pack, S.M.; Bonilla, O.D.; Cort-kansinally, K.A.; Coutinho, B.; Feng, M.; Garcia Martínez, M.I.; et al. The uncertain future of protected lands and waters. Science 2019, 364, 881–886. [Google Scholar] [CrossRef] [PubMed]
- Hilty, J.A.; Keeley, A.T.H.; Lidicker, W.Z., Jr.; Merenlender, A.M. Corridor Ecology: Linking Landscapes for Biodiversity Conservation and Climate Adaptation; Island Press: Washington, DC, USA, 2019. [Google Scholar]
- Sutherland, G.D.; Harestad, A.S.; Price, K.; Lertzman, K.P. Scaling of natal dispersal distances in terrestrial birds and mammals. Ecol. Soc. 2000, 4. [Google Scholar] [CrossRef]
- Gonzalez-Borrajo, N.; López-Bao, J.V.; Palomares, F. Spatial ecology of jaguars, pumas, and ocelots: A review of the state of knowledge. Mamm. Rev. 2017, 47, 62–75. [Google Scholar] [CrossRef] [Green Version]
- Maehr, D.S.; Land, E.D.; Shindle, D.B.; Bass, O.L.; Hoctor, T.S. Florida panther dispersal and conservation. Biol. Conserv. 2002, 106, 187–197. [Google Scholar] [CrossRef]
- Venter, O.; Sanderson, E.W.; Magrach, A.; Allan, J.R.; Beher, J.; Jones, K.R.; Possingham, H.P.; Laurance, W.F.; Wood, P.; Fekete, B.M.; et al. Sixteen years of change in the global terrestrial human footprint and implications for biodiversity conservation. Nat. Commun. 2016, 7, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Baldwin, R.F.; Perkl, R.M.; Trombulak, S.C.; Burwell, W.B. Modeling Ecoregional Connectivity. In Landscape-Scale Conservation Planning; Trombulak, S.C., Baldwin, R.F., Eds.; Springer: Berlin, Germany, 2010; pp. 349–367. ISBN 9789048195749. [Google Scholar]
- Alagador, D.; Triviño, M.; Cerdeira, J.O.; Brás, R.; Cabeza, M.; Araújo, M.B. Linking like with like: Optimising connectivity between environmentally-similar habitats. Landsc. Ecol. 2012, 27, 291–301. [Google Scholar] [CrossRef] [Green Version]
- Correa-Ayram, C.A.; Mendoza, M.E.; Etter, A.; Pérez Salicrup, D.R. Anthropogenic impact on habitat connectivity: A multidimensional human footprint index evaluated in a highly biodiverse landscape of Mexico. Ecol. Indic. 2017, 72, 895–909. [Google Scholar] [CrossRef]
- Belote, R.T.; Dietz, M.S.; McRae, B.H.; Theobald, D.M.; McClure, M.L.; Irwin, G.H.; McKinley, P.S.; Gage, J.A.; Aplet, G.H. Identifying corridors among large protected areas in the United States. PLoS ONE 2016, 11, e0154223. [Google Scholar] [CrossRef] [Green Version]
- Chapin III, F.S.; Matson, P.A.; Vitousek, P.M. Landscape Heterogeneity and Ecosystem Dynamics. In Principles of Terrestrial Ecosystem Ecology; Springer: New York, NY, USA, 2011; pp. 369–397. ISBN 978-1-4419-9504-9. [Google Scholar]
- Godínez-Gómez, O.; Correa-Ayram, C. Makurhini: Analyzing landscape connectivity. Zenodo 2020. [Google Scholar] [CrossRef]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2019. [Google Scholar]
- Dudley, N.; Jonas, H.; Nelson, F.; Parrish, J.; Pyhälä, A.; Stolton, S.; Watson, J.E.M. The essential role of other effective area-based conservation measures in achieving big bold conservation targets. Glob. Ecol. Conserv. 2018, 15, 1–7. [Google Scholar] [CrossRef]
- UNEP-WCMC. IUCN Protected Planet: Argentina, Bolivia, Brasil, Chile, Colombia, Ecuador, Guyana, Panamá, Paraguay, Perú, Venezuela; The World Database on Protected Areas (WDPA) [on-line], [Nov/2019]. Cambridge, UK. Available online: https://www.protectedplanet.net/ (accessed on 26 June 2020).
- Redparques Pronatura México. Progreso del cumplimiento de la Meta 11 de Aichi en los Países de la Redparques: Resultados y Perspectivas al 2020; Convention on Biological Diversity (CBD): Bogotá, Colombia, 2018. [Google Scholar]
- UNEP-WCMC. Manual de Usuario para la Base de Datos Mundial Sobre Áreas Protegidas y Base de Datos Mundial Sobre Otras Medidas Eficaces de Conservación Basadas en Áreas: 1.6; UNEP-WCMC: Cambridge, UK, 2020. [Google Scholar]
- Clements, H.S.; Selinske, M.J.; Archibald, C.L.; Cooke, B.; Fitzsimons, J.A.; Groce, J.E.; Torabi, N.; Hardy, M.J. Fairness and transparency are required for the inclusion of privately protected areas in publicly accessible conservation databases. Land 2018, 7, 96. [Google Scholar] [CrossRef] [Green Version]
- Pressey, R.L.; Cabeza, M.; Watts, M.E.; Cowling, R.M.; Wilson, K.A. Conservation planning in a changing world. Trends Ecol. Evol. 2007, 22, 583–592. [Google Scholar] [CrossRef] [PubMed]
- Franklin, J.F. Preserving Biodiversity: Species, Ecosystems, or Landscapes? Ecol. Appl. 1993, 3, 202–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, J.; Fazey, I.; Briese, R.; Lindenmayer, D.B. Making the matrix matter: Challenges in Australian grazing landscapes. Biodivers. Conserv. 2005, 14, 561–578. [Google Scholar] [CrossRef]
- Matallana Tobón, C.L.; Santamaría, M.; Areiza, A.; Solano, C.; Galán, S. Rethinking nature conservation in Colombia: A case study of other effective area—Based conservation measures. Parks 2018, 24, 89–98. [Google Scholar] [CrossRef]
- Margules, C.R.; Pressey, R.L. Systematic conservation planning. Nature 2000, 405, 243–253. [Google Scholar] [CrossRef]
- Venter, O.; Magrach, A.; Outram, N.; Klein, C.J.; Possingham, H.P.; Di Marco, M.; Watson, J.E.M. Bias in protected-area location and its effects on long-term aspirations of biodiversity conventions. Conserv. Biol. 2018, 32, 127–134. [Google Scholar] [CrossRef]
- Jenkins, C.N.; Van Houtan, K.S.; Pimm, S.L.; Sexton, J.O. US protected lands mismatch biodiversity priorities. Proc. Natl. Acad. Sci. USA 2015, 112, 5081–5086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miles, L.; Newton, A.C.; DeFries, R.S.; Ravilious, C.; May, I.; Blyth, S.; Kapos, V.; Gordon, J.E. A global overview of the conservation status of tropical dry forests. J. Biogeogr. 2006, 33, 491–505. [Google Scholar] [CrossRef]
- Portillo-Quintero, C.A.; Sánchez-Azofeifa, G.A. Extent and conservation of tropical dry forests in the Americas. Biol. Conserv. 2010, 143, 144–155. [Google Scholar] [CrossRef]
- Areiza, A.; Corzo, G.; Castillo, L.S.; Matallana, C.; Correa-Ayram, C.A. Áreas protegidas regionales y reservas privadas: Las protagonistas de las últimas décadas. In Biodiversidad 2018. Estado y Tendencias de la Biodiversidad Continental de Colombia; Moreno, L.A., Andrade, G.I., Goméz, M.F., Eds.; Instituto de Investigación de Recursos Biológicos Alexander von Humboldt: Bogotá, Colombia, 2019; ISBN 978-958-5418-63-9. [Google Scholar]
- SERNANP GEO ANP—Visor De Las Áreas Naturales Protegidas del Perú. Available online: http://geo.sernanp.gob.pe/visorsernanp/# (accessed on 30 May 2020).
- Armsworth, P.R.; Cantú-Salazar, L.; Parnell, M.; Davies, Z.G.; Stoneman, R. Management costs for small protected areas and economies of scale in habitat conservation. Biol. Conserv. 2011, 144, 423–429. [Google Scholar] [CrossRef] [Green Version]
- Larson, L.R.; Jennings, V.; Cloutier, S.A. Public parks and wellbeing in urban areas of the United States. PLoS ONE 2016, 11, e0153211. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TAC Country | National Extent 1 (Mha) | No. PA 2 | No. Subnal. PA 2 | PA Extent 1 (Mha) | Subnal. PA Extent 1 (Mha) | Subnal. Contrib. to PA Extent (%) | Protected Land (Prot) (%) | ||
|---|---|---|---|---|---|---|---|---|---|
| Our Study 1 | WDPA 3 | Redpar-Ques 4 | |||||||
| Bolivia | 108.9 | 132 | 110 | 29.5 | 12.4 | 41.9 | 27.1 | 30.9 | 30.9 |
| Colombia | 114.4 | 1154 | 1037 | 18.4 | 3.1 | 16.7 | 16.1 | 14.8 | 15.9 |
| Ecuador | 24.7 | 170 | 122 | 7.7 | 3.6 | 47.0 | 31.1 | 21.8 | 20.2 |
| Perú | 129.8 | 231 | 151 | 22.0 | 3.5 | 16.0 | 17.0 | 21.5 | 16.9 |
| Venezuela | 90.4 | 88 | 0 | 20.9 | 0.0 | 0.0 | 23.1 | 54.1 | 54.1 |
| TAC | 468.3 | 1775 | 1420 | 98.5 | 22.6 | 22.9 | 21.0 | – | – |
| Category (% of Ecoregion Area) | Prot | ProtConnEu | ProtConnCD | ProtConnCD without Subnal. | ||||
|---|---|---|---|---|---|---|---|---|
| No. Ecoreg. | Ext. (%) | No. Ecoreg. | Ext. (%) | No. Ecoreg. | Ext. (%) | No. Ecoreg. | Ext. (%) | |
| Omission (<1) | 2 | 0.7 | 2 | 0.7 | 2 | 0.7 | 5 | 2.0 |
| Insufficient for all targets (1–6.9) | 32 | 45.1 | 45 | 73.2 | 47 | 74.5 | 46 | 79.9 |
| Sufficient for Aichi 11 (17–29.9) | 17 | 36.2 | 7 | 12.9 | 6 | 13.0 | 5 | 8.3 |
| Sufficient for GDN (30–49.9) | 8 | 12.0 | 8 | 11.7 | 8 | 11.3 | 7 | 9.4 |
| Sufficient for all targets (≥50) | 8 | 5.9 | 5 | 1.4 | 4 | 0.5 | 4 | 0.5 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castillo, L.S.; Correa Ayram, C.A.; Matallana Tobón, C.L.; Corzo, G.; Areiza, A.; González-M., R.; Serrano, F.; Chalán Briceño, L.; Sánchez Puertas, F.; More, A.; et al. Connectivity of Protected Areas: Effect of Human Pressure and Subnational Contributions in the Ecoregions of Tropical Andean Countries. Land 2020, 9, 239. https://doi.org/10.3390/land9080239
Castillo LS, Correa Ayram CA, Matallana Tobón CL, Corzo G, Areiza A, González-M. R, Serrano F, Chalán Briceño L, Sánchez Puertas F, More A, et al. Connectivity of Protected Areas: Effect of Human Pressure and Subnational Contributions in the Ecoregions of Tropical Andean Countries. Land. 2020; 9(8):239. https://doi.org/10.3390/land9080239
Chicago/Turabian StyleCastillo, Luis Santiago, Camilo Andrés Correa Ayram, Clara L. Matallana Tobón, Germán Corzo, Alexandra Areiza, Roy González-M., Felipe Serrano, Luis Chalán Briceño, Felipe Sánchez Puertas, Alexander More, and et al. 2020. "Connectivity of Protected Areas: Effect of Human Pressure and Subnational Contributions in the Ecoregions of Tropical Andean Countries" Land 9, no. 8: 239. https://doi.org/10.3390/land9080239
APA StyleCastillo, L. S., Correa Ayram, C. A., Matallana Tobón, C. L., Corzo, G., Areiza, A., González-M., R., Serrano, F., Chalán Briceño, L., Sánchez Puertas, F., More, A., Franco, O., Bloomfield, H., Aguilera Orrury, V. L., Rivadeneira Canedo, C., Morón-Zambrano, V., Yerena, E., Papadakis, J., Cárdenas, J. J., Golden Kroner, R. E., & Godínez-Gómez, O. (2020). Connectivity of Protected Areas: Effect of Human Pressure and Subnational Contributions in the Ecoregions of Tropical Andean Countries. Land, 9(8), 239. https://doi.org/10.3390/land9080239