
Epitope Mapping of Monoclonal Antibodies to Calreticulin Reveals That Charged Amino Acids Are Essential for Antibody Binding

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Design of Synthetic Peptides
2.3. Screening of Antibody Reactivity to Wild Type Calreticulin by Enzyme-Linked Immunosorbent Assay
2.4. Screening of Resin-Bound Peptides by Modified Enzyme-Linked Immunosorbent Assay
2.5. PAGE and Western Blotting
3. Results
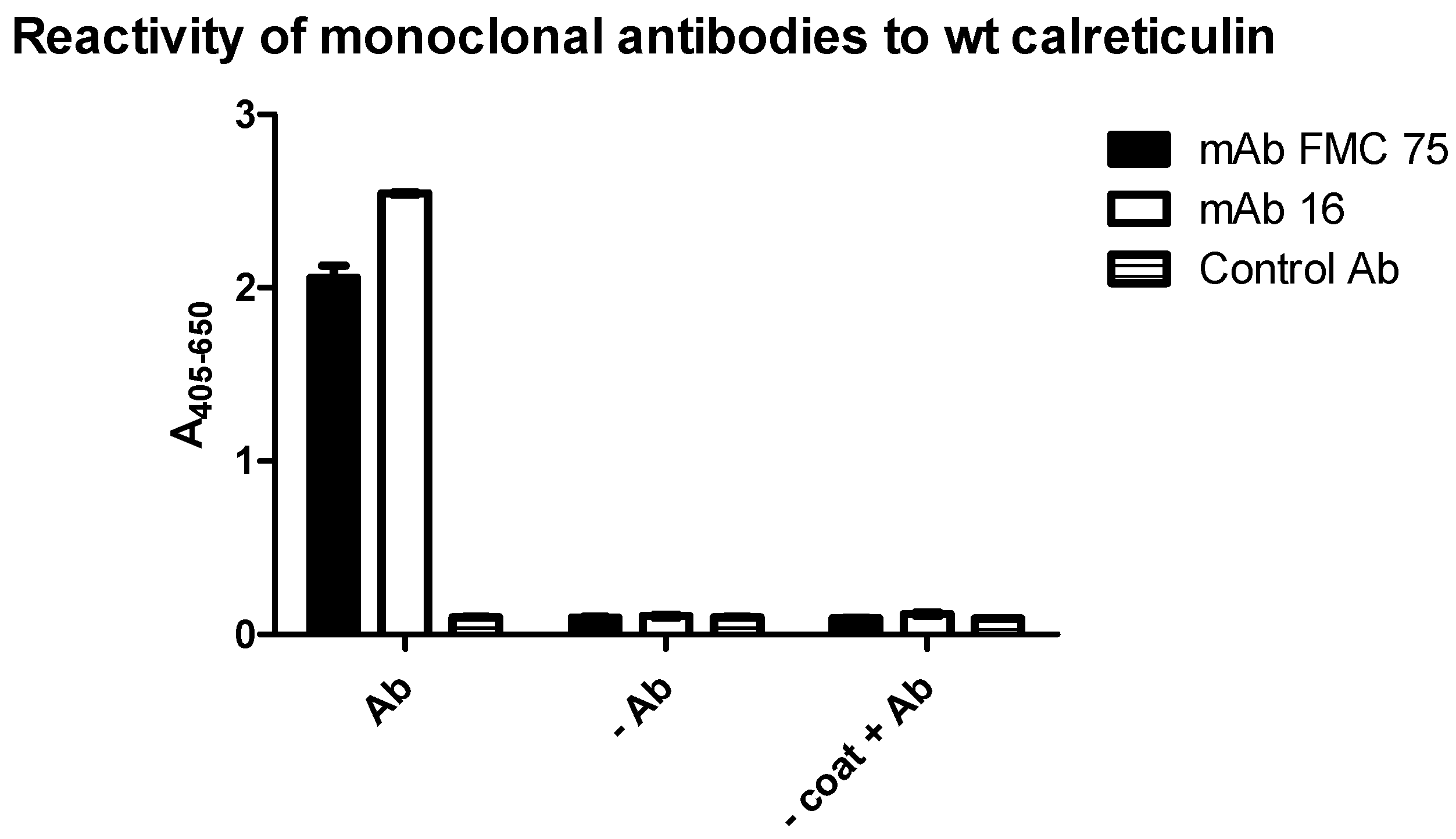
3.1. Reactivity of Monoclonal Antibodies to Wild Type Calreticulin
3.2. Reactivity of Calreticulin Antibodies to Overlapping Synthetic Peptides
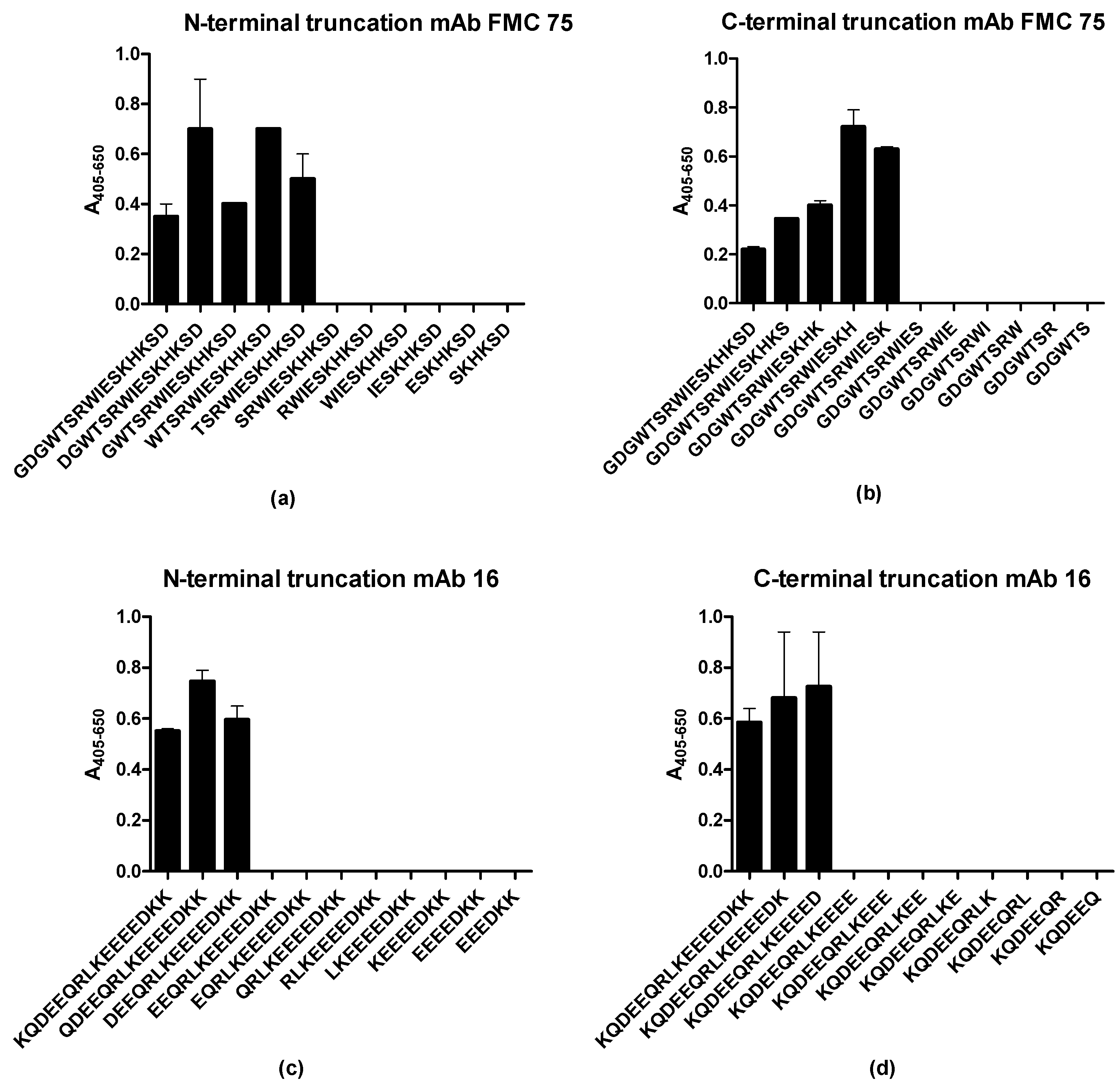
3.3. Identification of Minimal Sequences for Antibody Binding
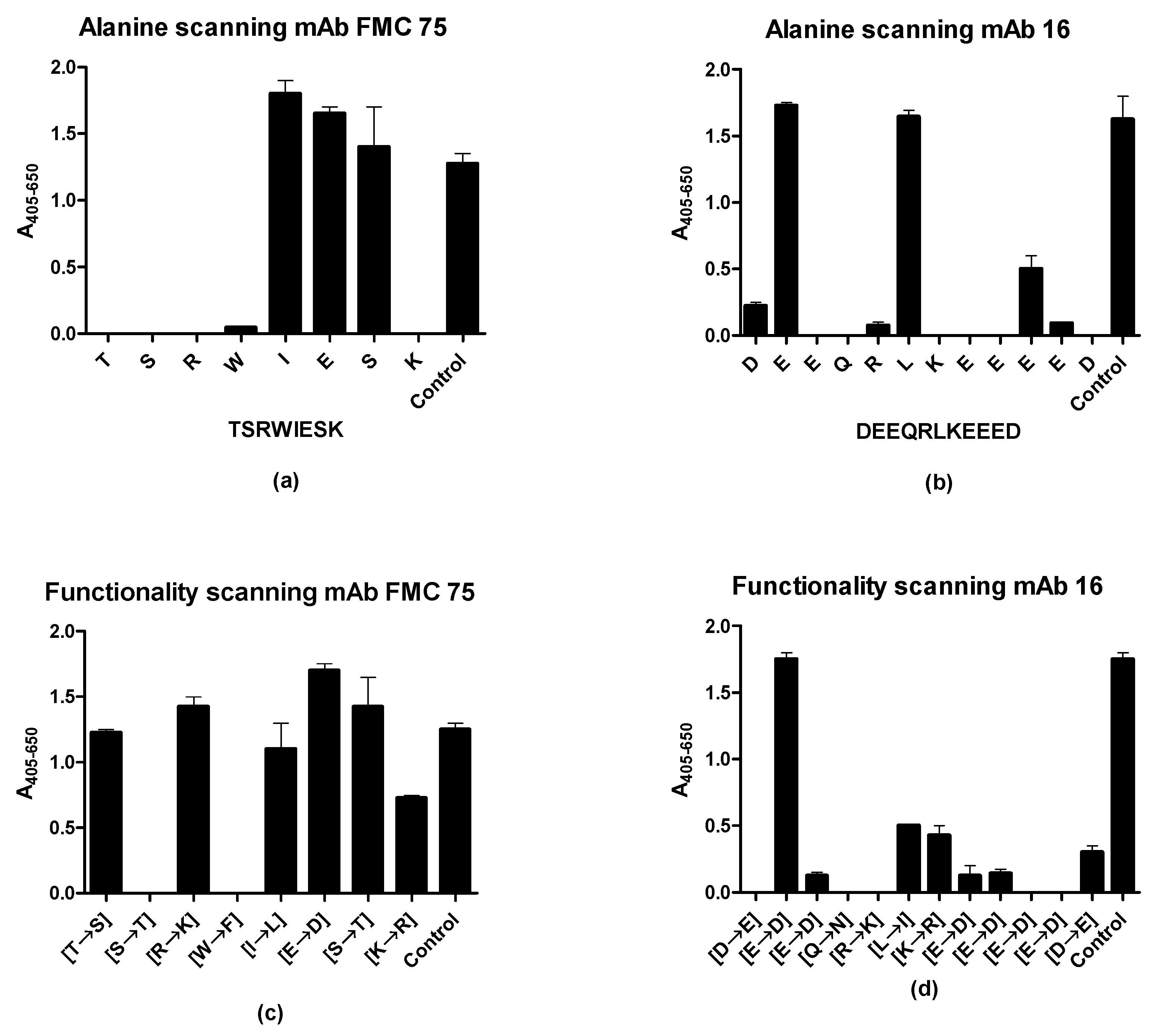
3.4. Determination of Antigenic Amino Acids Essential for Antibody Reactivity
3.5. Epitope Presentation in the Native Calreticulin Structure
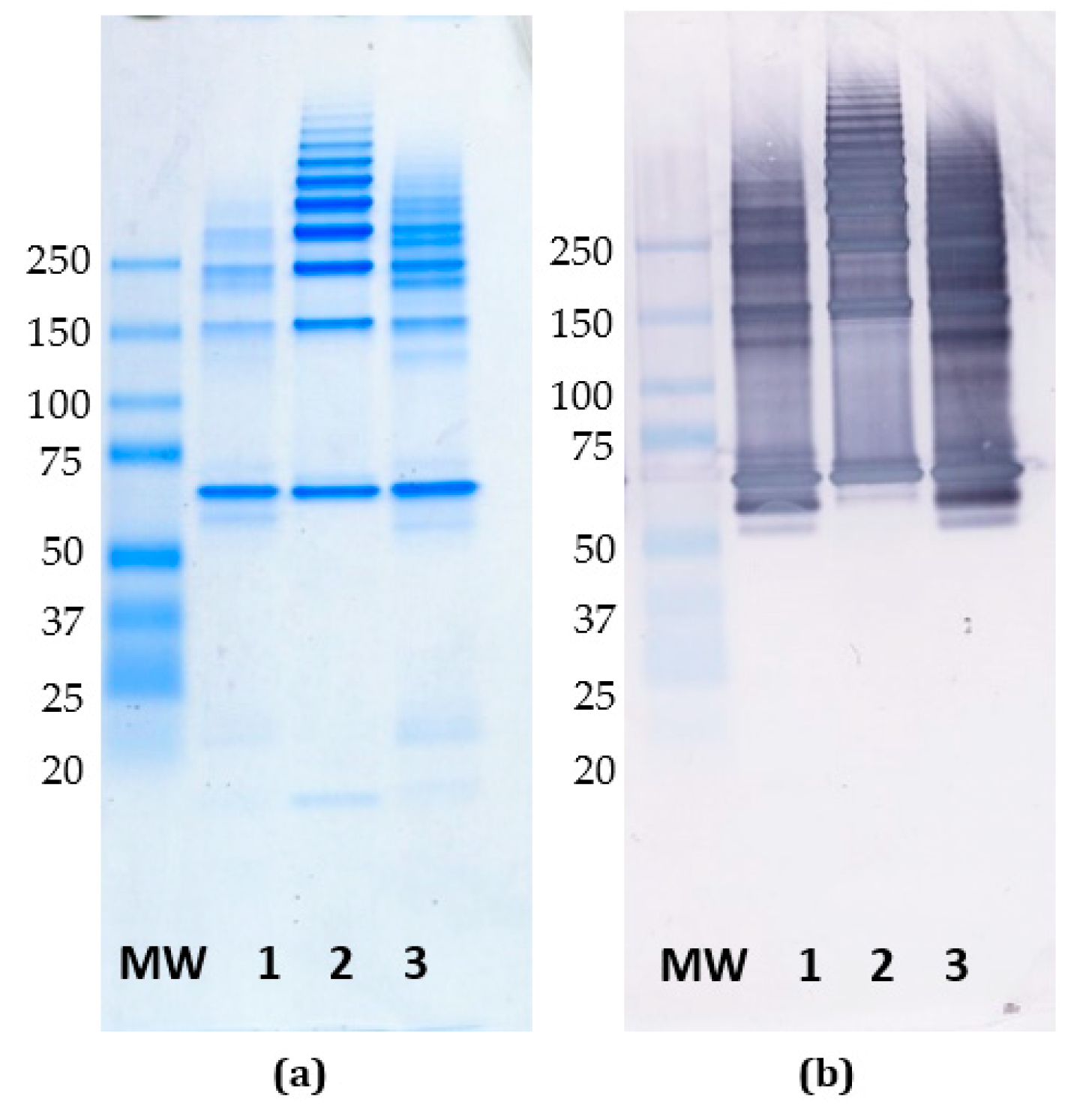
3.6. Reactivity of mAb FMC 75 to Oligomerized Calreticulin
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| # | Amino Acid | Peptide |
|---|---|---|
| 1 | 18–37 | EPAVYFKEQFLDGDGWTSRW |
| 2 | 28–47 | LDGDGWTSRWIESKHKSDFG |
| 3 | 38–57 | IESKHKSDFGKFVLSSGKFY |
| 4 | 48–67 | KFVLSSGKFYGDEEKDKGLQ |
| 5 | 58–77 | GDEEKDKGLQTSQDARFYAL |
| 6 | 68–87 | TSQDARFYALSASFEPFSNK |
| 7 | 78–97 | SASFEPFSNKGQTLVVQFTV |
| 8 | 88–107 | GQTLVVQFTVKHEQNIDCGG |
| 9 | 98–117 | KHEQNIDCGGGYVKLFPNSL |
| 10 | 108–127 | GYVKLFPNSLDQTDMHGDSE |
| 11 | 118–137 | DQTDMHGDSEYNIMFGPDIC |
| 12 | 128–147 | YNIMFGPDICGPGTKKVHVI |
| 13 | 138–157 | GPGTKKVHVIFNYKGKNVLI |
| 14 | 148–167 | FNYKGKNVLINKDIRCKDDE |
| 15 | 158–177 | NKDIRCKDDEFTHLYTLIVR |
| 16 | 168–187 | FTHLYTLIVRPDNTYEVKID |
| 17 | 178–197 | PDNTYEVKIDNSQVESGSLE |
| 18 | 188–207 | NSQVESGSLEDDWDFLPPKK |
| 19 | 198–217 | DDWDFLPPKKIKDPDASKPE |
| 20 | 208–227 | IKDPDASKPEDWDERAKIDD |
| 21 | 218–237 | DWDERAKIDDPTDSKPEDWD |
| 22 | 228–247 | PTDSKPEDWDKPEHIPDPDA |
| 23 | 238–257 | KPEHIPDPDAKKPEDWDEEM |
| 24 | 248–267 | KKPEDWDEEMDGEWEPPVIQ |
| 25 | 258–277 | DGEWEPPVIQNPEYKGEWKP |
| 26 | 268–287 | NPEYKGEWKPRQIDNPDYKG |
| 27 | 278–297 | RQIDNPDYKGTWIHPEIDNP |
| 28 | 288–307 | TWIHPEIDNPEYSPDPSIYA |
| 29 | 298–317 | EYSPDPSIYAYDNFGVLGLD |
| 30 | 308–327 | YDNFGVLGLDLWQVKSGTIF |
| 31 | 318–337 | LWQVKSGTIFDNFLITNDEA |
| 32 | 328–347 | DNFLITNDEAYAEEFGNETW |
| 33 | 338–357 | YAEEFGNETWGVTKAAEKQM |
| 34 | 348–367 | GVTKAAEKQMKDKQDEEQRL |
| 35 | 358–377 | KDKQDEEQRLKEEEEDKKRK |
| 36 | 368–387 | KEEEEDKKRKEEEEAEDKED |
| 37 | 378–397 | EEEEAEDKEDDEDKDEDEED |
| 38 | 388–407 | DEDKDEDEEDEEDKEEDEEE |
| 39 | 398–417 | EEDKEEDEEEDVPGQAKDEL |
References
- Wang, W.-A.; Groenendyk, J.; Michalak, M. Calreticulin signaling in health and disease. Int. J. Biochem. Cell Biol. 2012, 44, 842–846. [Google Scholar] [CrossRef]
- Michalak, M.; Groenendyk, J.; Szabo, E.; Gold, L.I.; Opas, M. Calreticulin, a multi-process calcium-buffering chaperone of the endoplasmic reticulum. Biochem. J. 2009, 417, 651–666. [Google Scholar] [CrossRef]
- Martins, I.; Kepp, O.; Galluzzi, L.; Senovilla, L.; Schlemmer, F.; Adjemian, S.; Menger, L.; Michaud, M.; Zitvogel, L.; Kroemer, G. Surface-exposed calreticulin in the interaction between dying cells and phagocytes. Ann. N. Y. Acad. Sci. 2010, 1209, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Holoshitz, J.; De Almeida, D.E.; Ling, S. A role for calreticulin in the pathogenesis of rheumatoid arthritis. Ann. N. Y. Acad. Sci. 2010, 1209, 91–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gold, L.I.; Eggleton, P.; Sweetwyne, M.T.; Van Duyn, L.B.; Greives, M.R.; Naylor, S.; Michalak, M.; Murphy-Ullrich, J.E. Calreticulin: Non-endoplasmic reticulum functions in physiology and disease. FASEB J. 2010, 24, 665–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumari, R.; Bansal, S.; Gupta, G.; Arora, S.; Kumar, A.; Goel, S.; Singh, P.; Ponnan, P.; Priya, N.; Tyagi, T.K.; et al. Calreticulin transacylase: Genesis, mechanism of action and biological applications. Biochimie 2010, 92, 1173–1179. [Google Scholar] [CrossRef]
- Villagomez, M.; Szabo, E.; Podcheko, A.; Feng, T.; Papp, S.; Opas, M. Calreticulin and focal-contact-dependent adhesion. Biochem. Cell Biol. 2009, 87, 545–556. [Google Scholar] [CrossRef]
- Chaput, N.; De Botton, S.; Obeid, M.; Apetoh, L.; Ghiringhelli, F.; Panaretakis, T.; Flament, C.; Zitvogel, L.; Kroemer, G. Molecular determinants of immunogenic cell death: Surface exposure of calreticulin makes the difference. J. Mol. Med. 2007, 85, 1069–1076. [Google Scholar] [CrossRef]
- Imai, M.; Araki, M.; Komatsu, N. Somatic mutations of calreticulin in myeloproliferative neoplasms. Int. J. Hematol. 2017, 105, 743–747. [Google Scholar] [CrossRef]
- Clinton, A.; McMullin, M.F. The Calreticulin gene and myeloproliferative neoplasms. J. Clin. Pathol. 2016, 69, 841–845. [Google Scholar] [CrossRef] [Green Version]
- Chi, J.; Manoloukos, M.; Pierides, C.; Nicolaidou, V.; Nicolaou, K.; Kleopa, M.; Vassiliou, G.; Costeas, P. Calreticulin mutations in myeloproliferative neoplasms and new methodology for their detection and monitoring. Ann. Hematol. 2015, 94, 399–408. [Google Scholar] [CrossRef]
- Klampfl, T.; Gisslinger, H.; Harutyunyan, A.; Nivarthi, H.; Rumi, E.; Milosevic, J.D.; Them, N.C.; Berg, T.; Gisslinger, B.; Pietra, D.; et al. Somatic Mutations of Calreticulin in Myeloproliferative Neoplasms. N. Engl. J. Med. 2013, 369, 2379–2390. [Google Scholar] [CrossRef] [Green Version]
- Nangalia, J.; Massie, C.E.; Baxter, E.J.; Nice, F.L.; Gundem, G.; Wedge, D.C.; Avezov, E.; Li, J.; Kollmann, K.; Kent, D.G.; et al. Somatic CALR mutations in myeloproliferative neoplasms with nonmutated JAK2. N. Engl. J. Med. 2013, 369, 2391–2405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozlov, G.; Gehring, K. Calnexin cycle—Structural features of the ER chaperone system. FEBS J. 2020, 287, 4322–4340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozlov, G.; Pocanschi, C.L.; Rosenauer, A.; Bastos-Aristizabal, S.; Gorelik, A.; Williams, D.B.; Gehring, K. Structural Basis of Carbohydrate Recognition by Calreticulin. J. Biol. Chem. 2010, 285, 38612–38620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chouquet, A.; Païdassi, H.; Ling, W.L.; Frachet, P.; Houen, G.; Arlaud, G.J.; Gaboriaud, C. X-ray structure of the human calreticulin globular domain reveals a peptide-binding area and suggests a multi-molecular mechanism. PLoS ONE 2011, 6, e17886. [Google Scholar] [CrossRef] [PubMed]
- Boelt, S.G.; Norn, C.; Rasmussen, M.I.; André, I.; Čiplys, E.; Slibinskas, R.; Houen, G.; Højrup, P. Mapping the Ca(2+) induced structural change in calreticulin. J. Proteomics 2016, 142, 138–148. [Google Scholar] [CrossRef] [PubMed]
- Houen, G.; Højrup, P.; Ciplys, E.; Gaboriaud, C.; Slibinskas, R. Structural Analysis of calreticulin, an endoplasmic reticulum-resident molecular chaperone. In Cellular Biology of the Endoplasmic Reticulum; Springer: Basel, Switzerland, 2021; pp. 13–25. [Google Scholar]
- Jørgensen, C.S.; Ryder, L.R.; Steinø, A.; Højrup, P.; Hansen, J.; Beyer, N.H.; Heegaard, N.H.H.; Houen, G. Dimerization and oligomerization of the chaperone calreticulin. Eur. J. Biochem. 2003, 270, 4140–4148. [Google Scholar] [CrossRef] [Green Version]
- Rizvi, S.M.; Mancino, L.; Thammavongsa, V.; Cantley, R.L.; Raghavan, M. A Polypeptide Binding Conformation of Calreticulin Is Induced by Heat Shock, Calcium Depletion, or by Deletion of the C-Terminal Acidic Region. Mol. Cell 2004, 15, 913–923. [Google Scholar] [CrossRef]
- Huang, S.-H.; Zhao, L.-X.; Hong, C.; Duo, C.-C.; Guo, B.-N.; Zhang, L.-J.; Gong, Z.; Xiong, S.-D.; Gong, F.-Y.; Gao, X.-M. Self-Oligomerization Is Essential for Enhanced Immunological Activities of Soluble Recombinant Calreticulin. PLoS ONE 2013, 8, e64951. [Google Scholar] [CrossRef] [Green Version]
- Araki, M.; Yang, Y.; Imai, M.; Mizukami, Y.; Kihara, Y.; Sunami, Y.; Masubuchi, N.; Edahiro, Y.; Hironaka, Y.; Osaga, S.; et al. Homomultimerization of mutant calreticulin is a prerequisite for MPL binding and activation. Leukemia 2019, 33, 122–131. [Google Scholar] [CrossRef] [PubMed]
- Rivera, J.F.; Baral, A.J.; Nadat, F.; Boyd, G.; Smyth, R.; Patel, H.; Burman, E.L.; Alameer, G.; Boxall, S.A.; Jackson, B.R.; et al. Zinc-dependent multimerization of mutant calreticulin is required for MPL binding and MPN pathogenesis. Blood Adv. 2021, 5, 1922–1932. [Google Scholar] [CrossRef] [PubMed]
- Venkatesan, A.; Geng, J.; Kandarpa, M.; Wijeyesakere, S.J.; Bhide, A.; Talpaz, M.; Pogozheva, I.D.; Raghavan, M. Mechanism of mutant calreticulin-mediated activation of the thrombopoietin receptor in cancers. J. Cell Biol. 2021, 220, e202009179. [Google Scholar] [CrossRef]
- Houen, G.; Trier, N. Molecular Recognition and Advances in Antibody Design and Antigenic Peptide Targeting. Int. J. Mol. Sci. 2020, 21, 1405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trier, N.H.; Houen, G. Antibodies as Diagnostic Targets and as Reagents for Diagnostics. Antibodies 2020, 9, 15. [Google Scholar] [CrossRef] [PubMed]
- Trier, N.; Hansen, P.; Houen, G. Peptides, antibodies, peptide antibodies and more. Int. J. Mol. Sci. 2019, 20, 6289. [Google Scholar] [CrossRef] [Green Version]
- Trier, N.H.; Houen, G. Peptide antibodies in clinical laboratory diagnostics. Adv. Clin. Chem. 2017, 81, 43–96. [Google Scholar]
- Wang, G.; Yang, J.; Liu, C. A mouse monoclonal antibody specific for calreticulin. Hybridoma 2012, 31, 382–385. [Google Scholar] [CrossRef]
- Corsiero, E.; Jagemann, L.; Perretti, M.; Pitzalis, C.; Bombardieri, M. Characterization of a synovial B cell-derived recombinant monoclonal antibody targeting stromal calreticulin in the rheumatoid joints. J. Immunol. 2018, 201, 1373–1381. [Google Scholar] [CrossRef]
- Stein, H.; Bob, R.; Dürkop, H.; Erck, C.; Kämpfe, D.; Kvasnicka, H.M.; Martens, H.; Roth, A.; Streubel, A. A new monoclonal antibody (CAL2) detects calreticulin mutations in formalin-fixed and paraffin-embedded bone marrow biopsies. Leukemia 2016, 30, 131–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandya, U.M.; Manzanares, M.A.; Tellechea, A.; Egbuta, C.; Daubriac, J.; Jimenez-Jaramillo, C.; Samra, F.; Fredston-Hermann, A.; Saadipour, K. Calreticulin exploits TGF-β for extracellular matrix induction engineering a tissue regenerative process. FASEB J. 2020, 34, 15849–15874. [Google Scholar] [CrossRef]
- Kasikova, L.; Truxova, I.; Cremer, I.; Sautes-Fridman, C.; Kepp, O.; Kroemer, G.; Spisek, R.; Fucikova, J. Side-by-side comparison of flow cytometry and immunohistochemistry for detection of calreticulin exposure in the course of immunogenic cell death. Methods Enzym. 2020, 632, 15–25. [Google Scholar]
- Pandya, U.M.; Egbuta, C.; Norman, T.F.A.; Chiang, C.; Wiersma, V.R.; Panchal, R.G.; Bremer, E.; Eggleton, P.; Gold, L.I. The biophysical interaction of the danger-associated molecular pattern (DAMP) calreticulin with the pattern-associated molecular pattern (PAMP) lipopolysaccharide. Int. J. Mol. Sci. 2019, 20, 408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tu, L.; Wang, J.; Barathi, V.A.; Prea, S.M.; He, Z.; Lee, J.H.; Bender, J.; King, A.E.; Logan, G.J.; Alexander, I.E.; et al. AAV-mediated gene delivery of the calreticulin anti-angiogenic domain inhibits ocular neovascularization. Angiogenesis 2018, 21, 95–109. [Google Scholar] [CrossRef] [PubMed]
- Arshad, N.; Cresswell, P. Tumor-associated calreticulin variants functionally compromise the peptide loading complex and impair its recruitment of MHC-I. J. Biol. Chem. 2018, 293, 9555–9569. [Google Scholar] [CrossRef] [Green Version]
- Ito, H.; Seyama, Y.; Kubota, S. Calreticulin is directly involved in anti-alpha3 integrin antibody-mediated secretion and activation of matrix metalloprotease-2. Biochem. Biophys. Res. Commun. 2001, 283, 297–302. [Google Scholar] [CrossRef]
- Nair, S.; Wearsch, P.A.; Mitchell, D.A.; Wassenberg, J.J.; Gilboa, E.; Nicchitta, C.V. Calreticulin displays in vivo peptide-binding activity and can elicit CTL responses against bound peptides. J. Immunol. 1999, 162, 6426–6432. [Google Scholar]
- Čiplys, E.; Žitkus, E.; Gold, L.I.; Daubriac, J.; Pavlides, S.C.; Højrup, P.; Houen, G.; Wang, W.A.; Michalak, M.; Slibinskas, R. High-level secretion of native recombinant human calreticulin in yeast. Microb. Cell Fact. 2015, 14, 165. [Google Scholar] [CrossRef] [Green Version]
- Amrutkar, S.D.; Trier, N.H.; Hansen, P.R.; Houen, G. Fine mapping of a monoclonal antibody to the N-Methyl D-aspartate receptor reveals a short linear epitope. Biopolymers 2012, 98, 567–575. [Google Scholar] [CrossRef]
- Petersen, N.H.; Hansen, P.R.; Houen, G. Fast and efficient characterization of an anti-gliadin monoclonal antibody epitope related to celiac disease using resin-bound peptides. J. Immunol. Methods 2011, 365, 174–182. [Google Scholar] [CrossRef]
- Valdarnini, N.; Holm, B.; Hansen, P.; Rovero, P.; Houen, G.; Trier, N. Fine mapping of glutamate decarboxylase 65 epitopes reveals dependency on hydrophobic amino acids for specific interactions. Int. J. Mol. Sci. 2019, 20, 2909. [Google Scholar] [CrossRef] [Green Version]
- Trier, N.H.; Hansen, P.R.; Vedeler, C.A.; Somnier, F.E.; Houen, G. Identification of continuous epitopes of HuD antibodies related to paraneoplastic diseases/small cell lung cancer. J. Neuroimmunol. 2012, 243, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Welner, S.; Trier, N.H.; Houen, G.; Hansen, P.R. Identification and mapping of a linear epitope of centromere protein F using monoclonal antibodies. J. Pept. Sci. 2013, 19, 95–101. [Google Scholar] [CrossRef]
- Agca, S.; Houen, G.; Trier, N.H. Characterization of continuous B-cell epitopes in the N-terminus of glutamate decarboxylase67 using monoclonal antibodies. J. Pept. Sci. 2014, 20, 928–934. [Google Scholar] [CrossRef] [PubMed]
- Fisette, O.; Schröder, G.F.; Schäfer, L.V. Atomistic structure and dynamics of the human MHC-I peptide loading complex. Proc. Natl. Acad. Sci. USA 2020, 117, 20597–20606. [Google Scholar] [CrossRef] [PubMed]
- Trier, N.H.; Holm, B.E.; Slot, O.; Locht, H.; Lindegaard, H.; Svendsen, A.; Houen, G. Physical characteristics of a citrullinated pro-filaggrin epitope recognized by anti-citrullinated protein antibodies in rheumatoid arthritis sera. PLoS ONE 2016, 11, e0168542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blees, A.; Januliene, D.; Hofmann, T.; Koller, N.; Schmidt, C.; Trowitzsch, S.; Moeller, A.; Tampé, R. Structure of the human MHC-I peptide-loading complex. Nature 2017, 551, 525–528. [Google Scholar] [CrossRef]
- Trier, N.H.; Dam, C.E.; Olsen, D.T.; Hansen, P.R.; Houen, G. Contribution of peptide backbone to citrulline-dependent antibody reactivity. PLoS ONE 2015, 10, e0144707. [Google Scholar] [CrossRef]






| N-Terminal Truncated Peptide | C-Terminal Truncated Peptides | Functionality-Substituted Peptides | Ala-Substituted Peptides | |
|---|---|---|---|---|
| mAb FMC 75 | ||||
| Template | GDGWTSRWIESKHKSD | TSRWIESK | ||
| DGWTSRWIESKHKSD | GDGWTSRWIESKHKS | SSRWIESK | ASRWIESK | |
| GWTSRWIESKHKSD | GDGWTSRWIESKHK | TTRWIESK | TARWIESK | |
| WTSRWIESKHKSD | GDGWTSRWIESKH | TSKWIESK | TSAWIESK | |
| TSRWIESKHKSD | GDGWTSRWIESK | TSRFIESK | TSRAIESK | |
| SRWIESKHKSD | GDGWTSRWIES | TSRWLESK | TSRWAESK | |
| RWIESKHKSD | GDGWTSRWIE | TSRWIDSK | TSRWIASK | |
| WIESKHKSD | GDGWTSRWI | TSRWIETK | TSRWIEAK | |
| IESKHKSD | GDGWTSRW | TSRWIESR | TSRWIESA | |
| ESKHKSD | GDGWTSR | |||
| SKHKSD | GDGWTS | |||
| KHKSD | GDGWT | |||
| HKSD | GDGW | |||
| mAb 16 | ||||
| Template | KDKQDEEQRLKEEEEDKKRK | DEEQRLKEEEED | ||
| QDEEQRLKEEEEDKK | KQDEEQRLKEEEEDK | EEEQRLKEEEED | AEEQRLKEEEED | |
| DEEQRLKEEEEDKK | KQDEEQRLKEEEED | DDEQRLKEEEED | DAEQRLKEEEED | |
| EEQRLKEEEEDKK | KQDEEQRLKEEEE | DEDQRLKEEEED | DEAQRLKEEEED | |
| EQRLKEEEEDKK | KQDEEQRLKEEE | DEENRLKEEEED | DEEARLKEEEED | |
| QRLKEEEEDKK | KQDEEQRLKEE | DEEQKLKEEEED | DEEQALKEEEED | |
| RLKEEEEDKK | KQDEEQRLKE | DEEQRIKEEEED | DEEQRAKEEEED | |
| LKEEEEDKK | KQDEEQRLK | DEEQRLREEEED | DEEQRLAEEEED | |
| KEEEEDKK | KQDEEQRL | DEEQRLKDEEED | DEEQRLKAEEED | |
| EEEEDKK | KQDEEQR | DEEQRLKEDEED | DEEQRLKEAEED | |
| EEEDKK | KQDEEQ | DEEQRLKEEDED | DEEQRLKEEAED | |
| EEDKK | KQDEE | DEEQRLKEEEDD | DEEQRLKEEEAD | |
| EDKK | KQDE | DEEQRLKEEEEE | DEEQRLKEEEEA | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bergmann, A.C.; Kyllesbech, C.; Slibinskas, R.; Ciplys, E.; Højrup, P.; Trier, N.H.; Houen, G. Epitope Mapping of Monoclonal Antibodies to Calreticulin Reveals That Charged Amino Acids Are Essential for Antibody Binding. Antibodies 2021, 10, 31. https://doi.org/10.3390/antib10030031
Bergmann AC, Kyllesbech C, Slibinskas R, Ciplys E, Højrup P, Trier NH, Houen G. Epitope Mapping of Monoclonal Antibodies to Calreticulin Reveals That Charged Amino Acids Are Essential for Antibody Binding. Antibodies. 2021; 10(3):31. https://doi.org/10.3390/antib10030031
Chicago/Turabian StyleBergmann, Ann Christina, Cecilie Kyllesbech, Rimantas Slibinskas, Evaldas Ciplys, Peter Højrup, Nicole Hartwig Trier, and Gunnar Houen. 2021. "Epitope Mapping of Monoclonal Antibodies to Calreticulin Reveals That Charged Amino Acids Are Essential for Antibody Binding" Antibodies 10, no. 3: 31. https://doi.org/10.3390/antib10030031
APA StyleBergmann, A. C., Kyllesbech, C., Slibinskas, R., Ciplys, E., Højrup, P., Trier, N. H., & Houen, G. (2021). Epitope Mapping of Monoclonal Antibodies to Calreticulin Reveals That Charged Amino Acids Are Essential for Antibody Binding. Antibodies, 10(3), 31. https://doi.org/10.3390/antib10030031