
IgG Avidity Test as a Tool for Discrimination between Recent and Distant Toxoplasma gondii Infection—Current Status of Studies


Abstract
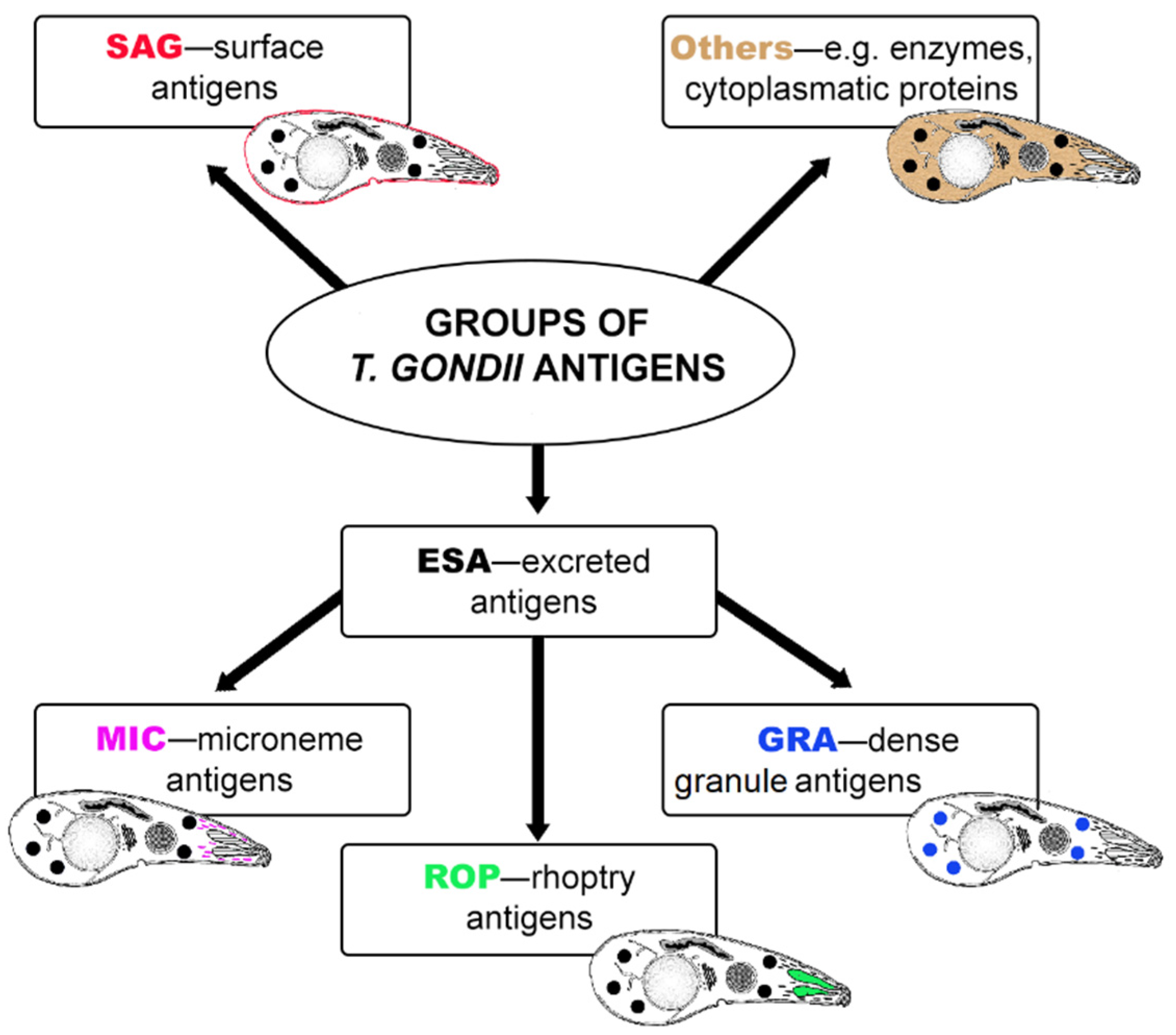
:1. Introduction
2. Antibody Affinity and Avidity
3. IgG Avidity Assays in Serodiagnosis of T. gondii Infection
4. Recombinant Antigens in IgG Avidity Tests
5. Summary and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dubey, J.P. The History of Toxoplasma gondii—The First 100 Years. J. Eukaryot. Microbiol. 2008, 55, 467–475. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Ocampo, F. Genomics of Apicomplexa. In Farm Animals Diseases, Recent Omic Trends and New Strategies of Treatment; IntechOpen: London, UK, 2018; p. 72633. [Google Scholar]
- Chakraborty, S.; Roy, S.; Mistry, H.U.; Murthy, S.; George, N.; Bhandari, V.; Sharma, P. Potential Sabotage of Host Cell Physiology by Apicomplexan Parasites for Their Survival Benefits. Front. Immunol. 2015, 8, 01261. [Google Scholar] [CrossRef]
- Gubbels, M.J.; Duraisingh, M.T. Evolution of apicomplexan secretory organelles. Int. J. Parasitol. 2012, 42, 1071–1081. [Google Scholar] [CrossRef] [PubMed]
- Dubey, J.P. The History and Life Cycle of Toxoplasma gondii. In Toxoplasma gondii the Model Apicomplexan: Perspectives and Methods; Weiss, L.M., Kim, K., Eds.; Academic Press: London, UK, 2007; pp. 1–17. [Google Scholar]
- Dubey, J.P.; Lindsay, D.S.; Speer, C.A. Structure of Toxoplasma gondii tachyzoites, bradyzoites and sporozoites, and biology and development of tissue cysts. Clin. Microbiol. Rev. 1998, 11, 267–299. [Google Scholar] [CrossRef]
- Hunter, C.A.; Sibley, L.D. Modulation of innate immunity by Toxoplasma gondii virulence effectors. Nat. Rev. Microbiol. 2012, 10, 766–778. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.L.; Dubey, J.P. Foodborne Toxoplasmosis. Clin. Infect. Dis. 2012, 55, 845–851. [Google Scholar] [CrossRef]
- Tenter, A.M.; Heckeroth, A.R.; Weiss, L.M. Toxoplasma gondii: From animals to humans. Int. J. Parasitol. 2000, 30, 1217–1258. [Google Scholar] [CrossRef]
- Weiss, L.M.; Kim, K. The International Congress on Toxoplasmosis. Int. J. Parasitol. 2004, 34, 249–252. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.-D.; Liu, H.-H.; Ma, Z.-X.; Ma, H.-Y.; Li, Z.-Y.; Yang, Z.-B.; Zhu, X.-Q.; Xu, B.; Wei, F.; Liu, Q. Toxoplasma gondii Infection in Immunocompromised Patients: A Systematic Review and Meta-Analysis. Front. Microbiol. 2017, 8, 389. [Google Scholar] [CrossRef]
- Montoya, J.G.; Remington, J.S. Management of Toxoplasma gondii infection during pregnancy. Clin. Infect. Dis. 2008, 47, 554–566. [Google Scholar] [CrossRef]
- Ortiz-Alegria, L.B.; Caballero-Ortega, H.; Cañedo-Solares, I.; Rico-Torres, C.P.; Sahagún-Ruiz, A.; Medina-Escuti, M.E.; Correa, D. Congenital toxoplasmosis: Candidate host immune genes relevant for vertical transmission and pathogenesis. Genes Immun. 2010, 11, 363–373. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Lin, G.; Han, Y.; Li, J. Serological diagnosis of toxoplasmosis and standardization. Clin. Chim. Acta 2016, 461, 83–89. [Google Scholar] [CrossRef]
- Dard, C.; Fricker-Hidalgo, H.; Brenier-Pinchart, M.P.; Pelloux, H. Relevance of and New Developments in Serology for Toxoplasmosis. Trends Parasitol. 2016, 32, 492–506. [Google Scholar] [CrossRef] [PubMed]
- Murat, J.B.; Hidalgo, H.F.; Brenier-Pinchart, M.P.; Pelloux, H. Human toxoplasmosis: Which biological diagnostic tests are best suited to which clinical situations? Expert Rev. Anti Infect. Ther. 2013, 11, 943–956. [Google Scholar] [CrossRef]
- Del Bono, V.; Canessa, A.; Bruzzi, P.; Fiorelli, M.A.; Terragna, A. Significance of specific immunoglobulin M in the chronological diagnosis of 38 cases of toxoplasmic lymphadenopathy. J. Clin. Microbiol. 1989, 27, 2133–2135. [Google Scholar] [CrossRef] [PubMed]
- Bobic, B.; Sibalic, D.; Djurkovic-Djakovic, O. High levels of IgM antibodies specific for Toxoplasma gondii in pregnancy 12 years after primary toxoplasma infection. Gynecol. Obstet. Investig. 1991, 31, 182–184. [Google Scholar]
- Liesenfeld, O.; Montoya, J.G.; Tathineni, N.J.; Davis, M.; Brown, B.W., Jr.; Cobb, K.L.; Parsonnet, J.; Remington, J.S. Confirmatory serologic testing for acute toxoplasmosis and rate of induced abortions among women reported to have positive Toxoplasma immunoglobulin M antibody titers. Am. J. Obstet. Gynecol. 2001, 184, 140–145. [Google Scholar] [CrossRef]
- Liesenfeld, O.; Press, C.; Montoya, J.G.; Gill, R.; Isaac-Renton, J.L.; Hedman, K.; Remington, J.S. False-positive results in immunoglobulin M (IgM) toxoplasma antibody tests and importance of confirmatory testing: The Platelia Toxo IgM test. J. Clin. Microbiol. 1997, 35, 174–178. [Google Scholar] [CrossRef] [PubMed]
- Stepick-Biek, P.; Thulliez, P.; Araujo, F.G.; Remington, J.S. IgA antibodies for diagnosis of acute congenital and acquired toxoplasmosis. J. Infect. Dis. 1990, 162, 270–273. [Google Scholar] [CrossRef]
- Wong, S.Y.; Hajdu, M.P.; Ramirez, R.; Thulliez, P.; McLeod, R.; Remington, J.S. Role of specific immunoglobulin E in diagnosis of acute toxoplasma infection and toxoplasmosis. J. Clin. Microbiol. 1993, 31, 2952–2959. [Google Scholar] [CrossRef]
- Paul, M.; Goullier-Fleuret, A.; Pelloux, H.; Ambroise-Thomas, P. The importance of the detection of anti-P30 IgA antibodies in acquired toxoplasmosis. Acta Parasitol. 1996, 41, 4. [Google Scholar]
- Gorgievski-Hrisoho, M.; Germann, D.; Matter, L. Diagnostic implications of kinetics of immunoglobulin M and A antibody responses to Toxoplasma gondii. J. Clin. Microbiol. 1996, 34, 1506–1511. [Google Scholar] [CrossRef]
- Robert-Gangneux, F.; Dardé, M.L. Epidemiology of and diagnostic strategies for toxoplasmosis. Clin. Microbiol. Rev. 2012, 25, 264–296. [Google Scholar] [CrossRef]
- Reiter-Owona, I. Laboratory diagnosis of toxoplasmosis—Possibilities and limitations. J. Lab. Med. 2005, 29, 439–445. [Google Scholar]
- Petersen, E.; Borobio, M.V.; Guy, E.; Liesenfeld, O.; Meroni, V.; Naessens, A.; Spranzi, E.; Thulliez, P. European multicenter study of the LIAISON automated diagnostic system for determination of Toxoplasma gondii-specific immunoglobulin G (IgG) and IgM and the IgG avidity index. J. Clin. Microbiol. 2005, 43, 1570–1574. [Google Scholar] [CrossRef] [PubMed]
- Jenum, P.A.; Stray-Pedersen, B.; Gundersen, A.G. Improved diagnosis of primary Toxoplasma gondii infection in early pregnancy by determination of antitoxoplasma immunoglobulin G avidity. J. Clin. Microbiol. 1997, 35, 1972–1977. [Google Scholar] [CrossRef] [PubMed]
- Cozon, G.J.; Ferrandiz, J.; Nebhi, H.; Wallon, M.; Peyron, F. Estimation of the avidity of immunoglobulin G for routine diagnosis of chronic Toxoplasma gondii infection in pregnant women. Eur. J. Clin. Microbiol. Infect. Dis. 1998, 17, 32–36. [Google Scholar] [CrossRef]
- Villard, O.; Breit, L.; Cimon, B.; Franck, J.; Fricker-Hidalgo, H.; Godineau, N.; Houze, S.; Paris, L.; Pelloux, H.; Villena, I.; et al. Comparison of four commercially available avidity tests for Toxoplasma gondii-specific IgG antibodies. Clin. Vaccine Immunol. 2013, 20, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Eisen, H.N.; Siskind, G.W. Variations in affinities of antibodies during the immune response. Biochemistry 1964, 3, 996–1008. [Google Scholar] [CrossRef]
- Werblin, T.P.; Kim, Y.T.; Quagliata, F.; Siskind, G.W. Studies on the control of antibody synthesis. III. Changes in heterogeneity of antibody affinity during the course of the immune response. Immunology 1973, 24, 477–492. [Google Scholar]
- Hedman, K.; Lappalainen, M.; Söderlund, M.; Hedman, L. Avidity of IgG in serodiagnosis of infectious diseases. Rev. Med. Microbiol. 1993, 4, 123–129. [Google Scholar] [CrossRef]
- Hedman, K.; Lappalainen, M.; Seppala, I.; Makela, O. Recent primary toxoplasma infection indicated by a low avidity specific IgG. J. Infect. Dis. 1989, 159, 736–739. [Google Scholar] [CrossRef] [PubMed]
- Hedman, K.; Rousseau, S.A. Measurement of avidity of specific IgG for verification of recent primary rubella. J. Med. Virol. 1989, 27, 288–292. [Google Scholar] [CrossRef]
- Hedman, K.; Seppala, I. Recent rubella virus infection indicated by a low avidity of specific IgG. J. Clin. Immunol. 1988, 8, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Grangeot-Keros, L.; Mayaux, M.J.; Lebon, P.; Freymuth, F.; Eugene, G.; Stricker, R.; Dussaix, E. Value of cytomegalovirus (CMV) IgG avidity index for the diagnosis of primary CMV infection in pregnant women. J. Infect. Dis. 1997, 175, 944–946. [Google Scholar] [CrossRef] [PubMed]
- Prince, H.E.; Lapé-Nixon, M. Role of cytomegalovirus (CMV) IgG avidity testing in diagnosing primary CMV infection during pregnancy. Clin. Vaccine Immunol. 2014, 21, 1377–1384. [Google Scholar] [CrossRef]
- Gaudy-Graffin, C.; Lesage, G.; Kousignian, I.; Laperche, S.; Girault, A.; Dubois, F.; Goudeau, A.; Barin, F. Use of an anti-hepatitis C virus (HCV) IgG avidity assay to identify recent HCV infection. J. Clin. Microbiol. 2010, 48, 3281–3287. [Google Scholar] [CrossRef]
- Junker, A.K.; Tilley, P. Varicella-zoster virus antibody avidity and IgG-subclass patterns in children with recurrent chickenpox. J. Med. Virol. 1994, 43, 119–124. [Google Scholar] [CrossRef]
- Absolom, D.R.; van Oss, C.J. The nature of the antigen-antibody bond and the factors affecting its association and dissociation. CRC Crit. Rev. Immunol. 1986, 6, 1–46. [Google Scholar]
- Liesenfeld, O.; Montoya, J.G.; Kinney, S.; Press, C.; Remington, J.S. Effect of testing for IgG avidity in the diagnosis of Toxoplasma gondii infection in pregnant women: Experience in a US reference laboratory. J. Infect. Dis. 2001, 183, 1248–1253. [Google Scholar] [CrossRef]
- Paul, M. Immunoglobulin G, avidity in diagnosis of toxolasmic lymphadenopathy and ocular toxoplasmosis. Clin. Diagn. Lab. Immunol. 1999, 6, 514–518. [Google Scholar] [CrossRef] [PubMed]
- Pietkiewicz, H.; Hiszczyńska-Sawicka, E.; Kur, J.; Petersen, E.; Nielsen, H.V.; Paul, M.; Stankiewicz, M.; Myjak, P. Usefulness of Toxoplasma gondii recombinant antigens (GRA1, GRA7 and SAG1) in an immunoglobulin G avidity test for the serodiagnosis of toxoplasmosis. Parasitol. Res. 2007, 100, 333–337. [Google Scholar] [CrossRef] [PubMed]
- Mechain, B.; Garin, Y.J.; Robert-Gangneux, F.; Dupouy-Camet, J.; Derouin, F. Lack of utility of specific immunoglobulin G antibody avidity for serodiagnosis of reactivated toxoplasmosis in immunocompromised patients. Clin. Diagn. Lab. Immunol. 2000, 7, 703–705. [Google Scholar] [CrossRef] [PubMed]
- Lappalainen, M.; Hedman, K. Serodiagnosis of toxoplasmosis. The impact of measurement of IgG avidity. Ann. Ist. Super. Sanita. 2004, 40, 81–88. [Google Scholar]
- Holec-Gasior, L. Toxoplasma gondii recombinant antigens as tools for serodiagnosis of human toxoplasmosis: Current status of studies. Clin. Vaccine Immunol. 2013, 20, 1343–1351. [Google Scholar] [CrossRef]
- Kotresha, D.; Noordin, R. Recombinant proteins in the diagnosis of toxoplasmosis. APMIS 2010, 18, 529–542. [Google Scholar] [CrossRef]
- Ferra, B.; Holec-Gąsior, L.; Grąźlewska, W. Toxoplasma gondii Recombinant Antigens in the Serodiagnosis of Toxoplasmosis in Domestic and Farm Animals. Animals 2020, 10, 1245. [Google Scholar] [CrossRef]
- Dubremetz, J.F.; Achbarou, A.; Bermudes, D.; Joiner, K.A. Kinetics and pattern of organelle exocytosis during Toxoplasma gondii/host-cell interaction. Parasitol. Res. 1993, 79, 402–408. [Google Scholar] [CrossRef]
- Carruthers, V.B.; Sibley, L.D. Sequential protein secretion from three distinct organelles of Toxoplasma gondii accompanies invasion of human fibroblasts. Eur. J. Cell Biol. 1997, 73, 114–123. [Google Scholar]
- Carruthers, V.B.; Giddings, O.K.; Sibley, L.D. Secretion of micronemal proteins is associated with Toxoplasma invasion of host cells. Cell. Microbiol. 1999, 1, 225–235. [Google Scholar] [CrossRef]
- Joiner, K.A.; Roos, D.S. Secretory traffic in the eukaryotic parasite Toxoplasma gondii: Less is more. J. Cell Biol. 2002, 157, 557–563. [Google Scholar] [CrossRef] [PubMed]
- Ngo, H.M.; Hoppe, H.C.; Joiner, K.A. Differential sorting and post-secretory targeting of proteins in parasitic invasion. Trends Cell Biol. 2000, 10, 67–72. [Google Scholar] [CrossRef]
- Sam-Yellowe, T.Y. Rhoptry organelles of the Apicomplexa: Their role in host cell invasion and intracellular survival. Parasitol. Today 1996, 12, 308–316. [Google Scholar] [CrossRef]
- Mercier, C.; Dubremetz, J.F.; Rauscher, B.; Lecordier, L.; Sibley, L.D.; Cesbron-Delauw, M.F. Biogenesis of nanotubular network in Toxoplasma parasitophorous vacuole induced by parasite proteins. Mol. Biol. Cell 2002, 13, 2397–2409. [Google Scholar] [CrossRef]
- Mercier, C.; Adjogble, K.D.; Däubener, W.; Delauw, M.F. Dense granules: Are they key organelles to help understand the parasitophorous vacuole of all apicomplexa parasites? Int. J. Parasitol. 2005, 35, 829–849. [Google Scholar] [CrossRef]
- Hughes, H.P.; Van, K.F. Characterization of a secretory antigen from Toxoplasma gondii and its role in circulating antigen production. Int. J. Parasitol. 1982, 12, 433–437. [Google Scholar] [CrossRef]
- Lekutis, C.; Ferguson, D.J.P.; Grigg, M.E.; Camps, M.; Boothroyd, J.C. Surface antigens of Toxoplasma gondii: Variations of a theme. Int. J. Parasitol. 2001, 31, 1285–1292. [Google Scholar] [CrossRef]
- Beghetto, E.; Buffolano, W.; Spadoni, A.; Del Pezzo, M.; Di Cristina, M.; Minenkova, O.; Petersen, E.; Felici, F.; Gargano, N. Use of an immunoglobulin G avidity assay based on recombinant antigens for diagnosis of primary Toxoplasma gondii infection during pregnancy. J. Clin. Microbiol. 2003, 41, 5414–5418. [Google Scholar] [CrossRef]
- Pfrepper, K.I.; Enders, G.; Gohl, M.; Krczal, D.; Hlobil, H.; Wassenberg, D.; Soutschek, E. Seroreactivity to and avidity for recombinant antigens in toxoplasmosis. Clin. Diagn. Lab. Immunol. 2005, 12, 977–982. [Google Scholar] [CrossRef]
- Béla, S.R.; Oliveira Silva, D.A.; Cunha-Júnior, J.P.; Pirovani, C.P.; Chaves-Borges, F.A.; Reis de Carvalho, F.; Carrijo de Oliveira, T.; Mineo, J.R. Use of SAG2A recombinant Toxoplasma gondii surface antigen as a diagnostic marker for human acute toxoplasmosis: Analysis of titers and avidity of IgG and IgG1 antibodies. Diagn. Microbiol. Infect. Dis. 2008, 62, 245–254. [Google Scholar] [CrossRef]
- Holec-Gąsior, L.; Drapała, D.; Lautenbach, D.; Kur, J. Toxoplasma gondii: Usefulness of ROP1 recombinant antigen in an immunoglobulin G avidity assay for diagnosis of acute toxoplasmosis in humans. Pol. J. Microbiol. 2010, 59, 307–310. [Google Scholar] [CrossRef] [PubMed]
- Elyasi, H.; Babaie, J.; Fricker-Hidalgo, H.; Brenier-Pinchart, M.P.; Zare, M.; Sadeghiani, G.; Assmar, M.; Pelloux, H.; Golkar, M. Use of dense granule antigen GRA6 in an immunoglobulin g avidity test to exclude acute Toxoplasma gondii infection during pregnancy. Clin. Vaccine Immunol. 2010, 17, 1349–1355. [Google Scholar] [CrossRef] [PubMed]
- Deshpande, P.S.; Kotresha, D.; Noordin, R.; Yunus, M.H.; Saadatnia, G.; Golkar, M.; Osman, S.; Karim, I.Z.A.; Ghaffarifar, F. IgG avidity Western blot using Toxoplasma gondii rGRA-7 cloned from nucleotides 39-711 for serodiagnosis of acute toxoplasmosis. Rev. Inst. Med. Trop. Sao Paulo 2013, 55, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Teh, A.Y.; Amerizadeh, A.; Osman, S.; Yunus, M.H.; Noordin, R. Identification, production and assessment of two Toxoplasma gondii recombinant proteins for use in a Toxoplasma IgG avidity assay. Pathog. Glob. Health 2016, 110, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Drapała, D.; Holec-Gąsior, L.; Kur, J.; Ferra, B.; Hiszczyńska-Sawicka, E.; Lautenbach, D. A new human IgG avidity test, using mixtures of recombinant antigens (rROP1, rSAG2, rGRA6), for the diagnosis of difficult-to-identify phases of toxoplasmosis. Diagn. Microbiol. Infect. Dis. 2014, 79, 342–346. [Google Scholar] [CrossRef]
- Teimouri, A.; Afshar, M.J.A.; Mohtasebi, S.; Azami, S.J.; Rasoul Alimi, R.; Keshavarz, H. Assessment of an In-House Enzyme-Linked Immunosorbent Assay and IgG Avidity Test Based on SAG1 and GRA7 Proteins for Discriminating between Acute and Chronic Toxoplasmosis in Humans. J. Clin. Microbiol. 2021, 59, e0041621. [Google Scholar] [CrossRef]
- Ferra, B.F.; Holec-Gąsior, L.; Gatkowska, J.; Dziadek, B.; Dzitko, K.; Grąźlewska, W.; Lautenbach, D. The first study on the usefulness of recombinant tetravalent chimeric proteins containing fragments of SAG2, GRA1, ROP1 and AMA1 antigens in the detection of specific anti-Toxoplasma gondii antibodies in mouse and human sera. PLoS ONE 2019, 14, e0217866. [Google Scholar] [CrossRef]
- Sickinger, E.; Gay-Andrieu, F.; Jonas, G.; Schultess, J.; Stieler, M.; Smith, D.; Hausmann, M.; Stricker, R.; Stricker, R.; Dhein, J.; et al. Performance characteristics of the new ARCHITECT Toxo IgG and Toxo IgG Avidity assays. Diagn. Microbiol. Infect. Dis. 2008, 62, 235–244. [Google Scholar] [CrossRef]
- Marcolino, P.T.; Silva, D.A.; Leser, P.G.; Camargo, M.E.; Mineo, J.R. Molecular markers in acute and chronic phases of human toxoplasmosis: Determination of immunoglobulin G avidity by Western blotting. Clin. Diagn. Lab. Immunol. 2000, 7, 384–389. [Google Scholar] [CrossRef]
- Soldati, D.; Lassen, A.; Dubremetz, J.F.; Boothroyd, J.C. Processing of Toxoplasma ROP1 protein in nascent rhoptries. Mol. Biochem. Parasitol. 1998, 96, 37–48. [Google Scholar] [CrossRef]
- Bradley, P.J.; Hsieh, C.L.; Boothroyd, J.C. Unprocessed Toxoplasma ROP1 is effectively targeted and secreted into the nascent parasitophorous vacuole. Mol. Biochem. Parasitol. 2002, 125, 189–193. [Google Scholar] [CrossRef]
- Pietkiewicz, H.; Hiszczyńska-Sawicka, E.; Kur, J.; Petersen, E.; Nielsen, H.V.; Stankiewicz, M.; Andrzejewska, I.; Myjak, P. Usefulness of Toxoplasma gondii-specific recombinant antigens in serodiagnosis of human toxoplasmosis. J. Clin. Microbiol. 2004, 42, 1779–1781. [Google Scholar] [CrossRef] [PubMed]
- Holec-Gąsior, L.; Kur, J.; Hiszczyńska-Sawicka, E. GRA2 and ROP1 recombinant antigens as potential markers for detection of Toxoplasma gondii-specific immunoglobulin G in humans with acute toxoplasmosis. Clin. Vaccine Immunol. 2009, 6, 510–514. [Google Scholar] [CrossRef] [PubMed]
- Malan Borel, I.; Gentile, T.; Angelucci, J.; Pividori, J.; Guala, M.D.C.; Binaghi, R.A.; Margni, R.A. IgG asymmetric molecules with antipaternal activity isolated from sera and placenta of pregnant human. J. Reprod. Immunol. 1991, 20, 129–140. [Google Scholar] [CrossRef]
- Pelloux, H.; Brun, E.; Vernet, G.; Marcillat, S.; Jolivet, M.; Guergour, D.; Fricker-Hidalgo, H.; Goullier-Fleuret, A.; Ambroise-Thomas, P. Determination of anti-Toxoplasma gondii immunoglobulin G avidity: Adaptation to the Vidas system (bioMérieux). Diagn. Microbiol. Infect. Dis. 1998, 32, 69–73. [Google Scholar] [CrossRef]
- Barberi, A.; Gistri, A.; Cappelletti, F.; Giordano, I. Diagnostic value of IgG avidity in Toxoplasma infection: Comparison of 3 commercial kits. J. Infect. Dis. 2001, 184, 944–946. [Google Scholar] [CrossRef]


{kind=link}
| Antigen | Type of Test | Denaturing Agent | Results | Literature, Year |
|---|---|---|---|---|
| GRA3, GRA7, MIC3 and SAG1 | ELISA | 6 M urea | MIC3 antigen—a marker of the early phase of toxoplasmosis. Low avidity index for people infected in the last 2 months. | [60], 2003 |
| ROP1, GRA8, MAG1, SAG1, GRA7 | nitrocellulose strips | 6 M urea | The research became the basis for the development of the recomLine Toxoplasma IgG Avidity diagnostic kit, in which 3 antigens (MAG1, GRA7, SAG1) were used. | [61], 2005 |
| GRA1, GRA7, SAG1 | ELISA | 6 M urea | The mixture of 3 antigens allows to obtain results similar to those in which Toxoplasma lysate antigen (TLA) was used. | [44], 2007 |
| P30 (SAG1), P35 (GRA8) | ARCHITECT Analyzer and chemiluminescent detection | not specified | IgG avidity test based on SAG1 antigen detected 100% of acute phase specimens (4 months after infection) as low avidity. | [70], 2008 |
| SAG2A | ELISA | 6 M urea | SAG2A antigen—suitable for use with the ELISA-IgG1 avidity assay. | [62], 2008 |
| ROP1 | ELISA | 6 M urea | ROP1 antigen—a potential marker of the early phase of toxoplasmosis. | [63], 2010 |
| GRA6 | ELISA | 8 M urea | IgG avidity ELISA based on GRA6 antigen was characterized as better performance than commercially available Euroimmun avidity for exclusion of early infection occurring less than 4 months previously. | [64], 2010 |
| rGRA7 cloned from nucleotides 39-711 | Western blot | 8 M urea | GRA7 recombinant antigen cloned from the gene segment comprising nucleotides 39-711 showed the potential of discrimination between acute and chronic toxoplasmosis | [65], 2013 |
| rROP1, rSAG2 and rGRA6 | ELISA | 6 M urea | A different trend in avidity maturation of IgG antibodies against two mixture of recombinant antigens (rROP1 + rSAG2 and rROP1 + rGRA6) compression with native antigens was observed. IgG avidity test using these mixtures of antigens may be useful for the diagnosis of difficult-to-identify phases of toxoplasmosis. | [67], 2014 |
| rAG12b and rAG18 | Western blot ELISA | 8 M urea | rAG12b and rAG18 antigens were able to differentiate low avidity and high avidity serum samples | [66], 2016 |
| Trivalent and tetravalent chimeric proteins: SAG2-GRA1-ROP1, SAG2-GRA1-ROP1-AMA1N, AMA1N-SAG2-GRA1-ROP1, AMA1C-SAG2-GRA1-ROP1, AMA1-SAG2-GRA1-ROP1 (AMA—apical membrane antigen) | ELISA | 6 M urea | AMA1-SAG2-GRA1-ROP1 chimeric protein – potential marker for distinguish specific antibodies from sera of individuals with the early and chronic phases T. gondii infections | [69], 2019 |
| SAG1 and GRA7 | ELISA | 8 M urea | SAG1 + GRA7 mixture showed the greatest potential for assessing avidity of antibodies | [68], 2021 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Holec-Gąsior, L.; Sołowińska, K. IgG Avidity Test as a Tool for Discrimination between Recent and Distant Toxoplasma gondii Infection—Current Status of Studies. Antibodies 2022, 11, 52. https://doi.org/10.3390/antib11030052
Holec-Gąsior L, Sołowińska K. IgG Avidity Test as a Tool for Discrimination between Recent and Distant Toxoplasma gondii Infection—Current Status of Studies. Antibodies. 2022; 11(3):52. https://doi.org/10.3390/antib11030052
Chicago/Turabian StyleHolec-Gąsior, Lucyna, and Karolina Sołowińska. 2022. "IgG Avidity Test as a Tool for Discrimination between Recent and Distant Toxoplasma gondii Infection—Current Status of Studies" Antibodies 11, no. 3: 52. https://doi.org/10.3390/antib11030052
APA StyleHolec-Gąsior, L., & Sołowińska, K. (2022). IgG Avidity Test as a Tool for Discrimination between Recent and Distant Toxoplasma gondii Infection—Current Status of Studies. Antibodies, 11(3), 52. https://doi.org/10.3390/antib11030052