A Novel Panel of Rabbit Monoclonal Antibodies and Their Diverse Applications Including Inhibition of Clostridium perfringens Epsilon Toxin Oligomerization

Abstract
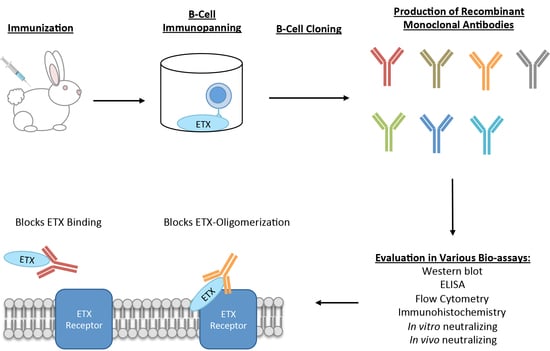
:
1. Introduction
2. Materials and Methods
2.1. Epsilon Protoxin
2.2. Activation of ETX
2.3. Rabbit Immunization
2.4. Affinity Purification of Rabbit Polyclonal Antibodies against pETX
2.5. Rabbit PBMC Isolation
2.6. B-Cell Panning
2.7. B-Cell Cloning and Monoclonal Antibody Production
2.8. rMAL-CHO Cell Culture
2.9. rMAL-CHO Cell Lysates from Western Blot Analysis
2.10. Western Blot (WB) Analysis and Densitometry Measurements
2.11. Indirect ELISA
2.12. Sandwich ELISA
2.13. Immunocytochemistry (ICC)
2.14. Flow Cytometry
2.15. Cell Death Assays with rMAL-CHO Cells
2.16. In Vivo Mouse Experiments
3. Results
3.1. Production of Monocolonal Antibodies Against ETX
3.2. Evaluation of Anti-ETX Antibodies for Detection of pETX, ETX, and ETX-Oligomer Complex via WB
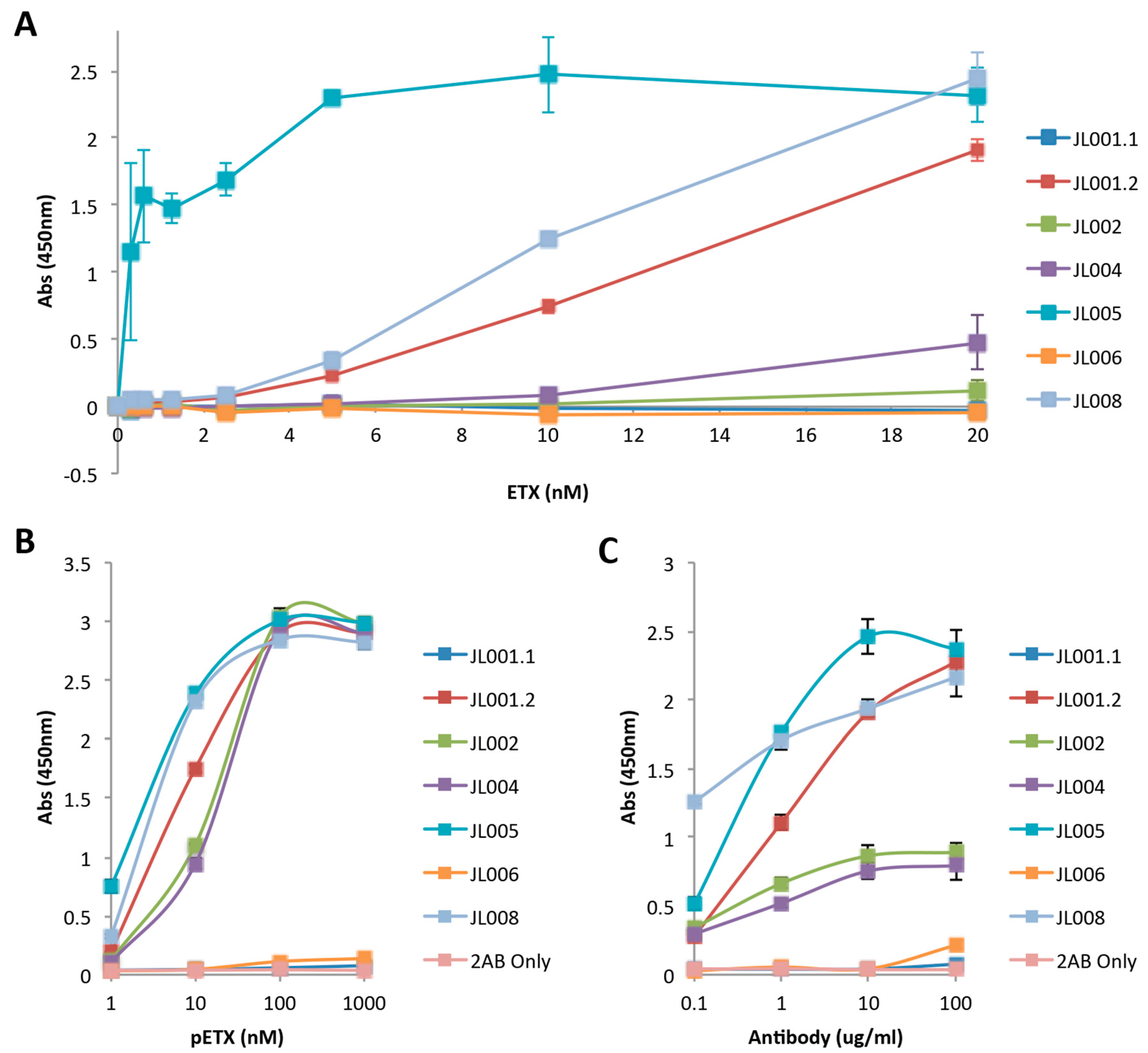
3.3. Evaluation of Anti-ETX Antibodies for Detection of ETX in Indirect ELISA
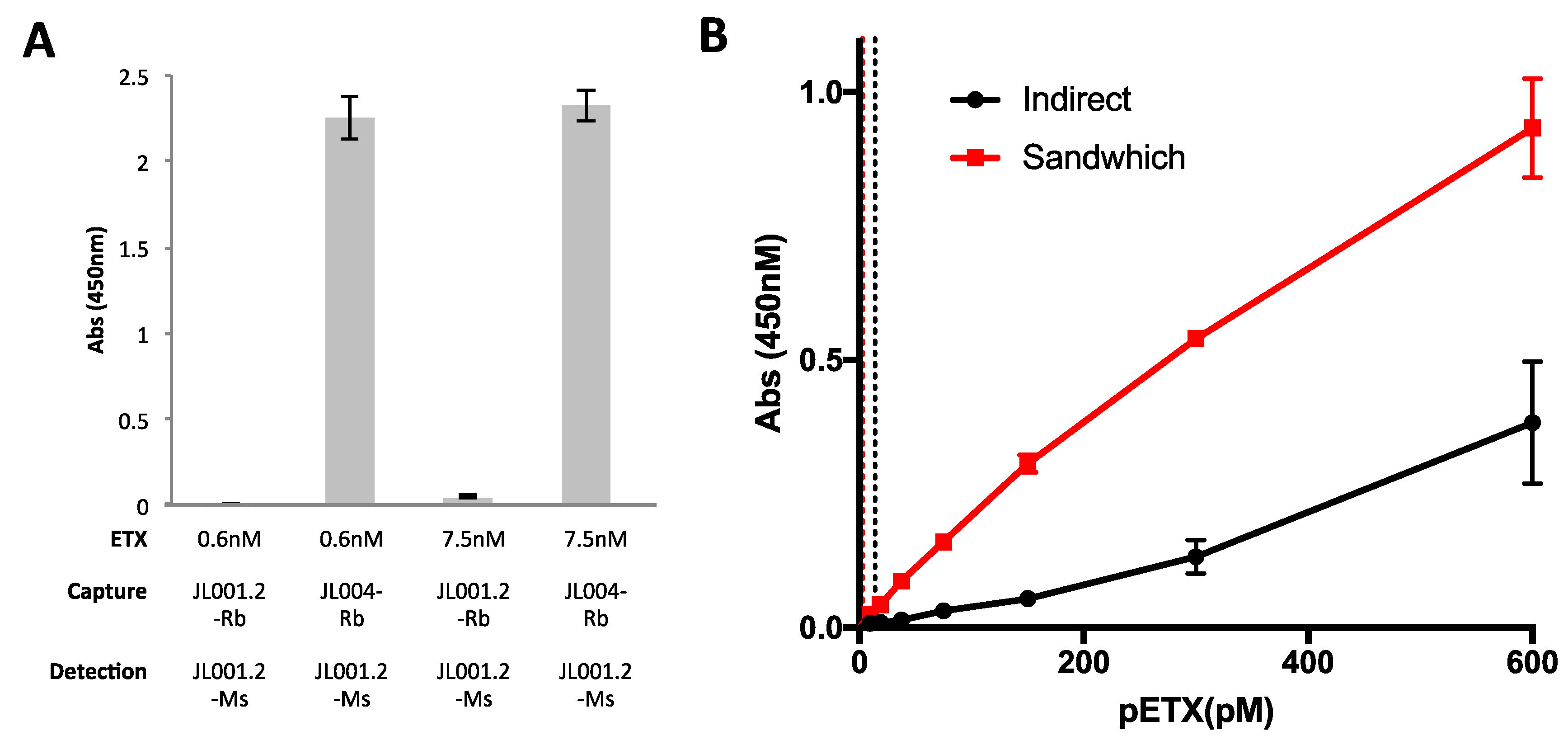
3.4. Evaluation of Anti-ETX Antibodies for Detection of pETX in Sandwich ELISA
3.5. Evaluation of Anti-ETX Antibodies for Detection of ETX via Immunocytochemistry (ICC)
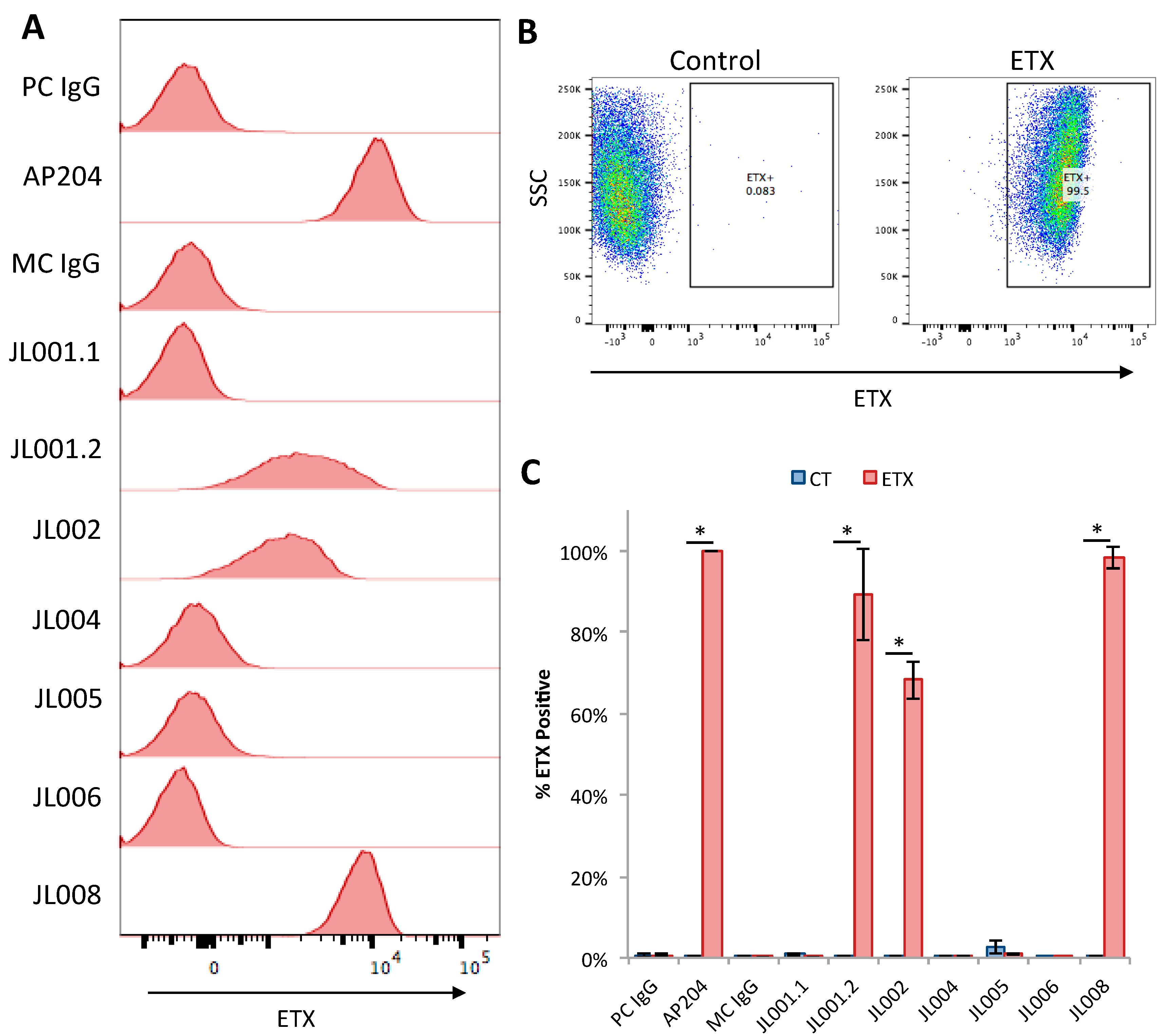
3.6. Evaluation of Anti-ETX Antibodies for Detection of ETX via Flow Cytometry
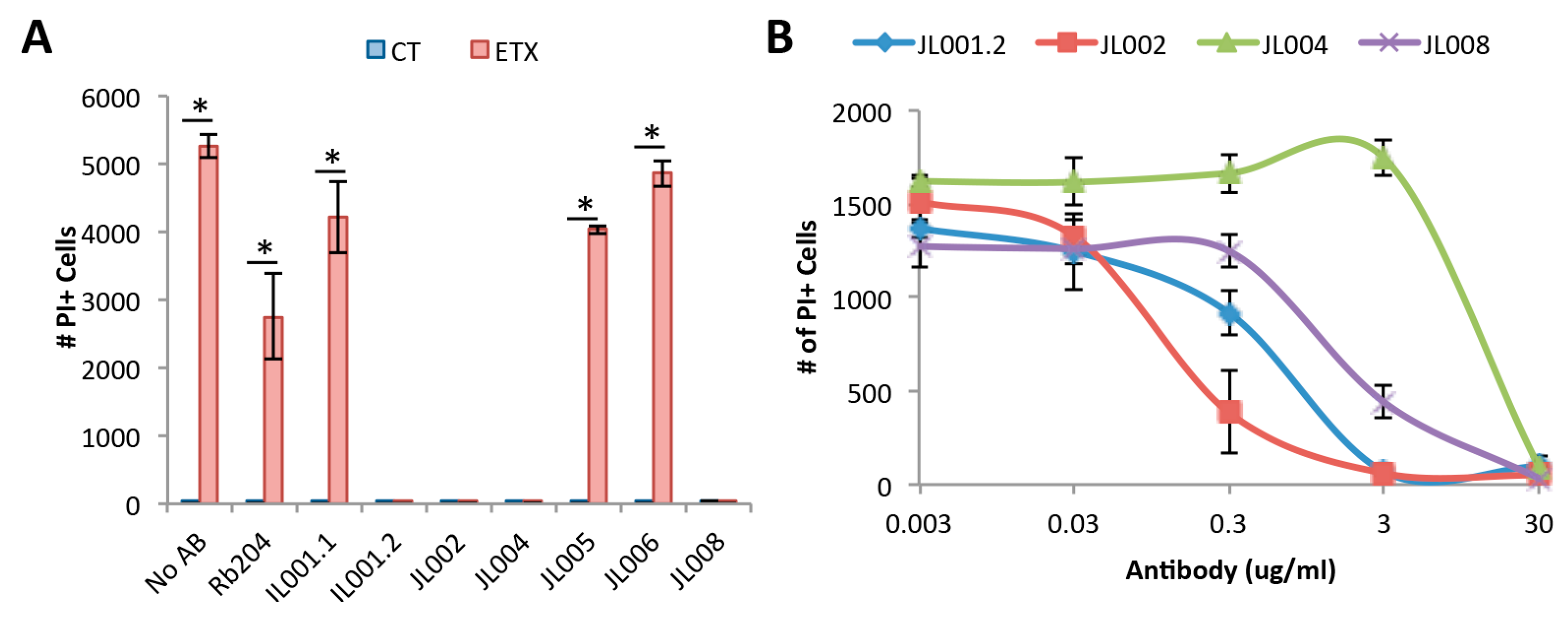
3.7. Neutralizing Ability of Anti-ETX Antibodies
3.8. Anti-ETX Antibodies Neutralize ETX by Blocking ETX Binding and Oligomerization
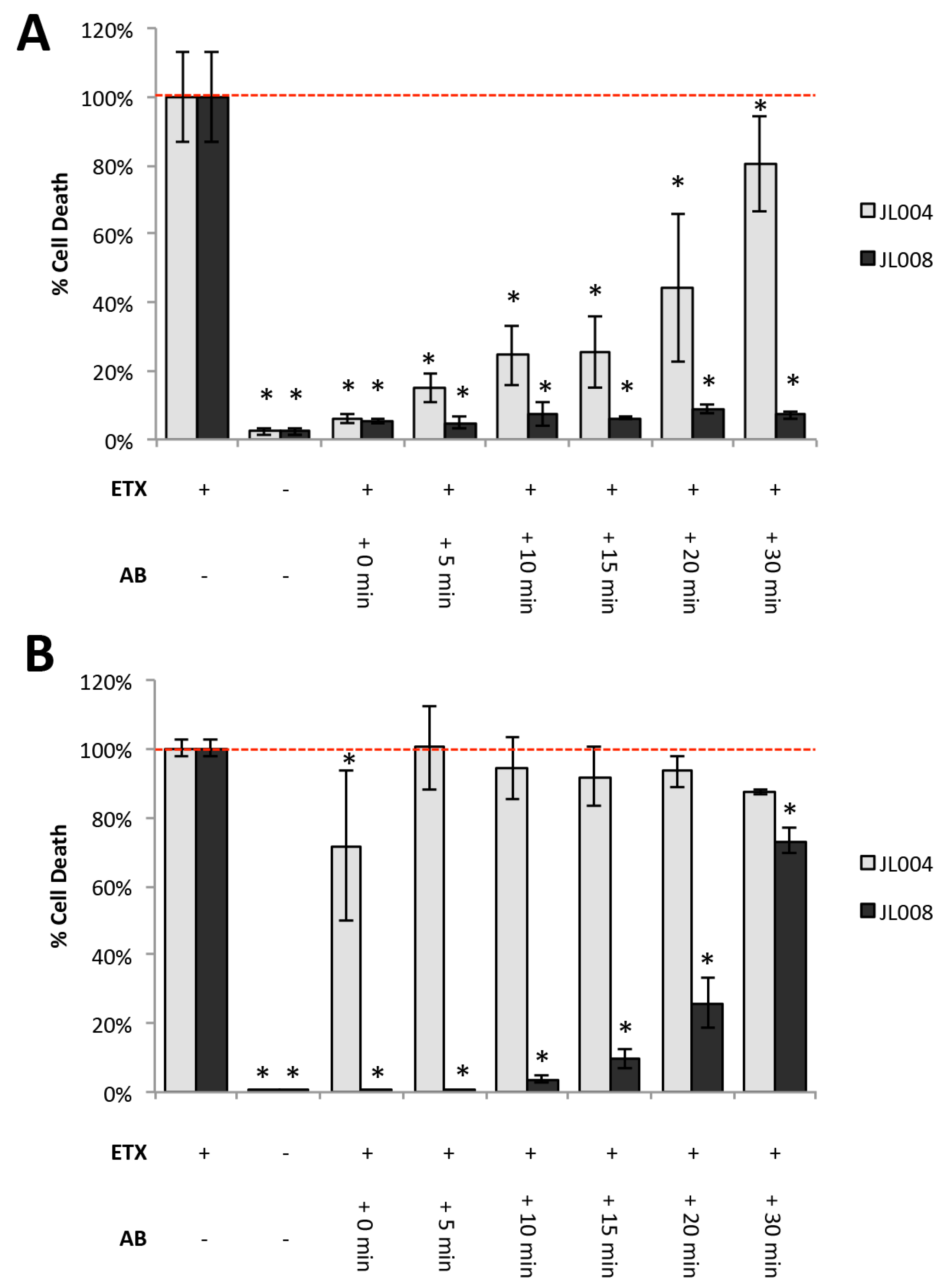
3.9. Post-Exposure Treatment With Anti-ETX Antibodies Protects Against Cytotoxicity in Vitro and in Vivo
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Garcia, J.P.; Adams, V.; Beingesser, J.; Hughes, M.L.; Poon, R.; Lyras, D.; Hill, A.; McClane, B.A.; Rood, J.I.; Uzal, F.A. Epsilon toxin is essential for the virulence of Clostridium perfringens type d infection in sheep, goats, and mice. Infect. Immun. 2013, 81, 2405–2414. [Google Scholar] [CrossRef] [PubMed]
- Uzal, F.A.; Pasini, M.I.; Olaechea, F.V.; Robles, C.A.; Elizondo, A. An outbreak of enterotoxaemia caused by Clostridium perfringens type d in goats in patagonia. Vet. Rec. 1994, 135, 279–280. [Google Scholar] [CrossRef] [PubMed]
- Alves, G.G.; Machado de Avila, R.A.; Chavez-Olortegui, C.D.; Lobato, F.C. Clostridium perfringens epsilon toxin: The third most potent bacterial toxin known. Anaerobe 2014, 30, 102–107. [Google Scholar] [CrossRef] [PubMed]
- Fernandez Miyakawa, M.E.; Zabal, O.; Silberstein, C. Clostridium perfringens epsilon toxin is cytotoxic for human renal tubular epithelial cells. Hum. Exp. Toxicol. 2011, 30, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Shortt, S.J.; Titball, R.W.; Lindsay, C.D. An assessment of the in vitro toxicology of Clostridium perfringens type d epsilon-toxin in human and animal cells. Hum. Exp. Toxicol. 2000, 19, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Ivie, S.E.; Fennessey, C.M.; Sheng, J.; Rubin, D.H.; McClain, M.S. Gene-trap mutagenesis identifies mammalian genes contributing to intoxication by Clostridium perfringens epsilon-toxin. PLoS ONE 2011, 6, e17787. [Google Scholar] [CrossRef] [PubMed]
- Blanch, M.; Dorca-Arevalo, J.; Not, A.; Cases, M.; Gomez de Aranda, I.; Martinez Yelamos, A.; Martinez Yelamos, S.; Solsona, C.; Blasi, J. The cytotoxicity of epsilon toxin from Clostridium perfringens on lymphocytes is mediated by mal protein expression. Mol. Cell. Biol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Murrell, T.G.; O’Donoghue, P.J.; Ellis, T. A review of the sheep-multiple sclerosis connection. Med. Hypotheses 1986, 19, 27–39. [Google Scholar] [CrossRef]
- Rumah, K.R.; Linden, J.; Fischetti, V.A.; Vartanian, T. Isolation of Clostridium perfringens type b in an individual at first clinical presentation of multiple sclerosis provides clues for environmental triggers of the disease. PLoS ONE 2013, 8, e76359. [Google Scholar] [CrossRef] [PubMed]
- Wagley, S.; Bokori-Brown, M.; Morcrette, H.; Malaspina, A.; D’Arcy, C.; Gnanapavan, S.; Lewis, N.; Popoff, M.R.; Raciborska, D.; Nicholas, R.; et al. Evidence of Clostridium perfringens epsilon toxin associated with multiple sclerosis. Mult. Scler. 2018. [Google Scholar] [CrossRef] [PubMed]
- Miyata, S.; Matsushita, O.; Minami, J.; Katayama, S.; Shimamoto, S.; Okabe, A. Cleavage of a c-terminal peptide is essential for heptamerization of Clostridium perfringens epsilon-toxin in the synaptosomal membrane. J. Biol. Chem. 2001, 276, 13778–13783. [Google Scholar] [CrossRef] [PubMed]
- Worthington, R.W.; Mulders, M.S. Physical changes in the epsilon prototoxin molecule of Clostridium perfringens during enzymatic activation. Infect. Immun. 1977, 18, 549–551. [Google Scholar] [PubMed]
- Minami, J.; Katayama, S.; Matsushita, O.; Matsushita, C.; Okabe, A. Lambda-toxin of Clostridium perfringens activates the precursor of epsilon-toxin by releasing its n- and c-terminal peptides. Microbiol. Immunol. 1997, 41, 527–535. [Google Scholar] [CrossRef] [PubMed]
- Robertson, S.L.; Li, J.; Uzal, F.A.; McClane, B.A. Evidence for a prepore stage in the action of Clostridium perfringens epsilon toxin. PLoS ONE 2011, 6, e22053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rumah, K.R.; Ma, Y.; Linden, J.R.; Oo, M.L.; Anrather, J.; Schaeren-Wiemers, N.; Alonso, M.A.; Fischetti, V.A.; McClain, M.S.; Vartanian, T. The myelin and lymphocyte protein mal is required for binding and activity of Clostridium perfringens epsilon-toxin. PLoS Pathog. 2015, 11, e1004896. [Google Scholar] [CrossRef] [PubMed]
- Fennessey, C.M.; Sheng, J.; Rubin, D.H.; McClain, M.S. Oligomerization of Clostridium perfringens epsilon toxin is dependent upon caveolins 1 and 2. PLoS ONE 2012, 7, e46866. [Google Scholar] [CrossRef] [PubMed]
- Ferrarezi, M.C.; Curci, V.C.; Cardoso, T.C. Cellular vacuolation and mitochondrial-associated factors induced by Clostridium perfringens epsilon toxin detected using acoustic flow cytometry. Anaerobe 2013, 24, 55–59. [Google Scholar] [CrossRef] [PubMed]
- Chassin, C.; Bens, M.; de Barry, J.; Courjaret, R.; Bossu, J.L.; Cluzeaud, F.; Ben Mkaddem, S.; Gibert, M.; Poulain, B.; Popoff, M.R.; et al. Pore-forming epsilon toxin causes membrane permeabilization and rapid atp depletion-mediated cell death in renal collecting duct cells. Am. J. Physiol. Renal Physiol. 2007, 293, F927–F937. [Google Scholar] [CrossRef] [PubMed]
- Petit, L.; Maier, E.; Gibert, M.; Popoff, M.R.; Benz, R. Clostridium perfringens epsilon toxin induces a rapid change of cell membrane permeability to ions and forms channels in artificial lipid bilayers. J. Biol. Chem. 2001, 276, 15736–15740. [Google Scholar] [CrossRef] [PubMed]
- Nagahama, M.; Ochi, S.; Sakurai, J. Assembly of Clostridium perfringens epsilon-toxin on mdck cell membrane. J. Nat. Toxins 1998, 7, 291–302. [Google Scholar] [PubMed]
- Petit, L.; Gibert, M.; Gillet, D.; Laurent-Winter, C.; Boquet, P.; Popoff, M.R. Clostridium perfringens epsilon-toxin acts on mdck cells by forming a large membrane complex. J. Bacteriol. 1997, 179, 6480–6487. [Google Scholar] [CrossRef] [PubMed]
- Nestorovich, E.M.; Karginov, V.A.; Bezrukov, S.M. Polymer partitioning and ion selectivity suggest asymmetrical shape for the membrane pore formed by epsilon toxin. Biophys. J. 2010, 99, 782–789. [Google Scholar] [CrossRef] [PubMed]
- Cole, A.R.; Gibert, M.; Popoff, M.; Moss, D.S.; Titball, R.W.; Basak, A.K. Clostridium perfringens epsilon-toxin shows structural similarity to the pore-forming toxin aerolysin. Nat. Struct. Mol. Biol. 2004, 11, 797–798. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Xin, W.; Gao, S.; Kang, L.; Wang, J. A low-toxic site-directed mutant of Clostridium perfringens epsilon-toxin as a potential candidate vaccine against enterotoxemia. Hum. Vaccines Immunother. 2013, 9, 2386–2392. [Google Scholar] [CrossRef]
- Kang, J.; Gao, J.; Yao, W.; Kang, L.; Gao, S.; Yang, H.; Ji, B.; Li, P.; Liu, J.; Yao, J.; et al. F199e substitution reduced toxicity of Clostridium perfringens epsilon toxin by depriving the receptor binding capability. Hum. Vaccines Immunother. 2017, 13, 1598–1608. [Google Scholar] [CrossRef] [PubMed]
- Yao, W.; Kang, J.; Kang, L.; Gao, S.; Yang, H.; Ji, B.; Li, P.; Liu, J.; Xin, W.; Wang, J. Immunization with a novel Clostridium perfringens epsilon toxin mutant retx(y196e)-c confers strong protection in mice. Sci. Rep. 2016, 6, 24162. [Google Scholar] [CrossRef] [PubMed]
- Yao, W.; Kang, L.; Gao, S.; Zhuang, X.; Zhang, T.; Yang, H.; Ji, B.; Xin, W.; Wang, J. Amino acid residue y196e substitution and c-terminal peptide synergistically alleviate the toxicity of Clostridium perfringens epsilon toxin. Toxicon 2015, 100, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Chang, J.; Wang, F.; Yu, L. Identification of tyrosine 71 as a critical residue for the cytotoxic activity of Clostridium perfringens epsilon toxin towards mdck cells. J. Microbiol. 2015, 53, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Pelish, T.M.; McClain, M.S. Dominant-negative inhibitors of the Clostridium perfringens epsilon-toxin. J. Biol. Chem. 2009, 284, 29446–29453. [Google Scholar] [CrossRef] [PubMed]
- Bokori-Brown, M.; Kokkinidou, M.C.; Savva, C.G.; Fernandes da Costa, S.; Naylor, C.E.; Cole, A.R.; Moss, D.S.; Basak, A.K.; Titball, R.W. Clostridium perfringens epsilon toxin h149a mutant as a platform for receptor binding studies. Protein Sci. 2013, 22, 650–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bokori-Brown, M.; Savva, C.G.; Fernandes da Costa, S.P.; Naylor, C.E.; Basak, A.K.; Titball, R.W. Molecular basis of toxicity of Clostridium perfringens epsilon toxin. FEBS J. 2011, 278, 4589–4601. [Google Scholar] [CrossRef] [PubMed]
- Knapp, O.; Maier, E.; Benz, R.; Geny, B.; Popoff, M.R. Identification of the channel-forming domain of Clostridium perfringens epsilon-toxin (etx). Biochim. Biophys. Acta 2009, 1788, 2584–2593. [Google Scholar] [CrossRef] [PubMed]
- Ivie, S.E.; McClain, M.S. Identification of amino acids important for binding of Clostridium perfringens epsilon toxin to host cells and to havcr1. Biochemistry 2012, 51, 7588–7595. [Google Scholar] [CrossRef] [PubMed]
- Alves, G.G.; Machado-de-Avila, R.A.; Chavez-Olortegui, C.D.; Silva, R.O.S.; Lobato, F.C.F. Mapping of the continuous epitopes displayed on the Clostridium perfringens type d epsilon-toxin. Braz. J. Microbiol. 2017, 48, 570–575. [Google Scholar] [CrossRef] [PubMed]
- McClain, M.S.; Cover, T.L. Functional analysis of neutralizing antibodies against Clostridium perfringens epsilon-toxin. Infect. Immun. 2007, 75, 1785–1793. [Google Scholar] [CrossRef] [PubMed]
- Feraudet-Tarisse, C.; Mazuet, C.; Pauillac, S.; Kruger, M.; Lacroux, C.; Popoff, M.R.; Dorner, B.G.; Andreoletti, O.; Plaisance, M.; Volland, H.; et al. Highly sensitive sandwich immunoassay and immunochromatographic test for the detection of clostridial epsilon toxin in complex matrices. PLoS ONE 2017, 12, e0181013. [Google Scholar] [CrossRef] [PubMed]
- Hauer, P.J.; Clough, N.E. Development of monoclonal antibodies suitable for use in antigen quantification potency tests for clostridial veterinary vaccines. Dev. Biol. Stand. 1999, 101, 85–94. [Google Scholar] [PubMed]
- Garcia, J.P.; Beingesser, J.; Bohorov, O.; Bohorova, N.; Goodman, C.; Kim, D.; Pauly, M.; Velasco, J.; Whaley, K.; Zeitlin, L.; et al. Prevention and treatment of Clostridium perfringens epsilon toxin intoxication in mice with a neutralizing monoclonal antibody (c4d7) produced in nicotiana benthamiana. Toxicon 2014, 88, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Sully, E.K.; Whaley, K.; Bohorova, N.; Bohorov, O.; Goodman, C.; Kim, D.; Pauly, M.; Velasco, J.; Holtsberg, F.W.; Stavale, E.; et al. A tripartite cocktail of chimeric monoclonal antibodies passively protects mice against ricin, staphylococcal enterotoxin b and Clostridium perfringens epsilon toxin. Toxicon 2014, 92, 36–41. [Google Scholar] [CrossRef] [PubMed]
- el Idrissi, A.H.; Ward, G.E. Development of double sandwich elisa for Clostridium perfringens beta and epsilon toxins. Vet. Microbiol. 1992, 31, 89–99. [Google Scholar] [CrossRef]
- Nagahama, M.; Kobayashi, K.; Ochi, S.; Sakurai, J. Enzyme-linked immunosorbent assay for rapid detection of toxins from Clostridium perfringens. FEMS Microbiol. Lett. 1991, 68, 41–44. [Google Scholar] [CrossRef] [PubMed]
- Uzal, F.A.; Kelly, W.R.; Thomas, R.; Hornitzky, M.; Galea, F. Comparison of four techniques for the detection of Clostridium perfringens type d epsilon toxin in intestinal contents and other body fluids of sheep and goats. J. Vet. Diagn. Investig. 2003, 15, 94–99. [Google Scholar] [CrossRef] [PubMed]
- Rossi, S.; Laurino, L.; Furlanetto, A.; Chinellato, S.; Orvieto, E.; Canal, F.; Facchetti, F.; Dei Tos, A.P. Rabbit monoclonal antibodies: A comparative study between a novel category of immunoreagents and the corresponding mouse monoclonal antibodies. Am. J. Clin. Pathol. 2005, 124, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Vilches-Moure, J.G.; Ramos-Vara, J.A. Comparison of rabbit monoclonal and mouse monoclonal antibodies in immunohistochemistry in canine tissues. J. Vet. Diagn. Investig. 2005, 17, 346–350. [Google Scholar] [CrossRef] [PubMed]
- Liguori, M.J.; Hoff-Velk, J.A.; Ostrow, D.H. Recombinant human interleukin-6 enhances the immunoglobulin secretion of a rabbit-rabbit hybridoma. Hybridoma 2001, 20, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Popkov, M.; Mage, R.G.; Alexander, C.B.; Thundivalappil, S.; Barbas, C.F., 3rd; Rader, C. Rabbit immune repertoires as sources for therapeutic monoclonal antibodies: The impact of kappa allotype-correlated variation in cysteine content on antibody libraries selected by phage display. J. Mol. Biol. 2003, 325, 325–335. [Google Scholar] [CrossRef]
- Babcook, J.S.; Leslie, K.B.; Olsen, O.A.; Salmon, R.A.; Schrader, J.W. A novel strategy for generating monoclonal antibodies from single, isolated lymphocytes producing antibodies of defined specificities. Proc. Natl. Acad. Sci. USA 1996, 93, 7843–7848. [Google Scholar] [CrossRef] [PubMed]
- Spieker-Polet, H.; Sethupathi, P.; Yam, P.C.; Knight, K.L. Rabbit monoclonal antibodies: Generating a fusion partner to produce rabbit-rabbit hybridomas. Proc. Natl. Acad. Sci. USA 1995, 92, 9348–9352. [Google Scholar] [CrossRef] [PubMed]
- Lipman, N.S.; Jackson, L.R.; Trudel, L.J.; Weis-Garcia, F. Monoclonal versus polyclonal antibodies: Distinguishing characteristics, applications, and information resources. ILAR J. 2005, 46, 258–268. [Google Scholar] [CrossRef] [PubMed]
- Lightwood, D.J.; Carrington, B.; Henry, A.J.; McKnight, A.J.; Crook, K.; Cromie, K.; Lawson, A.D. Antibody generation through b cell panning on antigen followed by in situ culture and direct rt-pcr on cells harvested en masse from antigen-positive wells. J. Immunol. Methods 2006, 316, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Nagahama, M.; Itohayashi, Y.; Hara, H.; Higashihara, M.; Fukatani, Y.; Takagishi, T.; Oda, M.; Kobayashi, K.; Nakagawa, I.; Sakurai, J. Cellular vacuolation induced by Clostridium perfringens epsilon-toxin. FEBS J. 2011, 278, 3395–3407. [Google Scholar] [CrossRef] [PubMed]
- Worthington, R.W.; Mulders, M.S. Effect of Clostridium perfringens epsilon toxin on the blood brain barrier of mice. Onderstepoort J. Vet. Res. 1975, 42, 25–27. [Google Scholar] [PubMed]
- Finnie, J.W. Histopathological changes in the brain of mice given Clostridium perfringens type d epsilon toxin. J. Comp. Pathol. 1984, 94, 363–370. [Google Scholar] [CrossRef]
- Finnie, J.W.; Hajduk, P. An immunohistochemical study of plasma albumin extravasation in the brain of mice after the administration of Clostridium perfringens type d epsilon toxin. Aust. Vet. J. 1992, 69, 261–262. [Google Scholar] [CrossRef] [PubMed]
- Finnie, J.W.; Blumbergs, P.C.; Manavis, J. Neuronal damage produced in rat brains by Clostridium perfringens type d epsilon toxin. J. Comp. Pathol. 1999, 120, 415–420. [Google Scholar] [CrossRef] [PubMed]
- Finnie, J.W.; Manavis, J.; Blumbergs, P.C. Aquaporin-4 in acute cerebral edema produced by Clostridium perfringens type d epsilon toxin. Vet. Pathol. 2008, 45, 307–309. [Google Scholar] [CrossRef] [PubMed]
- Finnie, J.W.; Manavis, J.; Chidlow, G. Loss of endothelial barrier antigen immunoreactivity as a marker of Clostridium perfringens type d epsilon toxin-induced microvascular damage in rat brain. J. Comp. Pathol. 2014, 151, 153–156. [Google Scholar] [CrossRef] [PubMed]
- Idone, V.; Tam, C.; Goss, J.W.; Toomre, D.; Pypaert, M.; Andrews, N.W. Repair of injured plasma membrane by rapid Ca2+-dependent endocytosis. J. Cell Biol. 2008, 180, 905–914. [Google Scholar] [CrossRef] [PubMed]
- Tam, C.; Idone, V.; Devlin, C.; Fernandes, M.C.; Flannery, A.; He, X.; Schuchman, E.; Tabas, I.; Andrews, N.W. Exocytosis of acid sphingomyelinase by wounded cells promotes endocytosis and plasma membrane repair. J. Cell Biol. 2010, 189, 1027–1038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thiery, J.; Keefe, D.; Saffarian, S.; Martinvalet, D.; Walch, M.; Boucrot, E.; Kirchhausen, T.; Lieberman, J. Perforin activates clathrin- and dynamin-dependent endocytosis, which is required for plasma membrane repair and delivery of granzyme b for granzyme-mediated apoptosis. Blood 2010, 115, 1582–1593. [Google Scholar] [CrossRef] [PubMed]
- Husmann, M.; Beckmann, E.; Boller, K.; Kloft, N.; Tenzer, S.; Bobkiewicz, W.; Neukirch, C.; Bayley, H.; Bhakdi, S. Elimination of a bacterial pore-forming toxin by sequential endocytosis and exocytosis. FEBS Lett. 2009, 583, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, M.G.; Saka, H.A.; Chinen, I.; Zoppino, F.C.; Yoshimori, T.; Bocco, J.L.; Colombo, M.I. Protective role of autophagy against vibrio cholerae cytolysin, a pore-forming toxin from v. Cholerae. Proc. Natl. Acad. Sci. USA 2007, 104, 1829–1834. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Western Blot | ELISA | Flow | ICC | Neutralizing | Neutralizing MOA | |||
|---|---|---|---|---|---|---|---|---|---|
| pETX | ETX | Bound ETX | Pore | ETX | ETX | ETX | |||
| JL001.1 | + | + | − | − | − | − | − | − | n/a |
| JL001.2 | + | + | + | − | + | + | + | + | Oligomerization |
| JL002 | + | + | - | − | − | + | + | + | Oligomerization |
| JL004 | + | + | + | + | + | − | − | + | Binding |
| JL005 | + | + | + | + | + | − | − | − | n/a |
| JL006 | + | + | - | − | − | − | − | − | n/a |
| JL008 | + | + | + | − | + | + | + | + | Oligomerization |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Linden, J.R.; Telesford, K.; Shetty, S.; Winokour, P.; Haigh, S.; Cahir-McFarland, E.; Antognetti, G.; Datta, A.; Wang, T.; Meier, W.; et al. A Novel Panel of Rabbit Monoclonal Antibodies and Their Diverse Applications Including Inhibition of Clostridium perfringens Epsilon Toxin Oligomerization. Antibodies 2018, 7, 37. https://doi.org/10.3390/antib7040037
Linden JR, Telesford K, Shetty S, Winokour P, Haigh S, Cahir-McFarland E, Antognetti G, Datta A, Wang T, Meier W, et al. A Novel Panel of Rabbit Monoclonal Antibodies and Their Diverse Applications Including Inhibition of Clostridium perfringens Epsilon Toxin Oligomerization. Antibodies. 2018; 7(4):37. https://doi.org/10.3390/antib7040037
Chicago/Turabian StyleLinden, Jennifer R., Kiel Telesford, Samantha Shetty, Paige Winokour, Sylvia Haigh, Ellen Cahir-McFarland, Giovanna Antognetti, Abhishek Datta, Tao Wang, Werner Meier, and et al. 2018. "A Novel Panel of Rabbit Monoclonal Antibodies and Their Diverse Applications Including Inhibition of Clostridium perfringens Epsilon Toxin Oligomerization" Antibodies 7, no. 4: 37. https://doi.org/10.3390/antib7040037
APA StyleLinden, J. R., Telesford, K., Shetty, S., Winokour, P., Haigh, S., Cahir-McFarland, E., Antognetti, G., Datta, A., Wang, T., Meier, W., & Vartanian, T. (2018). A Novel Panel of Rabbit Monoclonal Antibodies and Their Diverse Applications Including Inhibition of Clostridium perfringens Epsilon Toxin Oligomerization. Antibodies, 7(4), 37. https://doi.org/10.3390/antib7040037