Isolation and Characterization of Nanobodies against a Zinc-Transporting P-Type ATPase

 , ,
, ,
Abstract
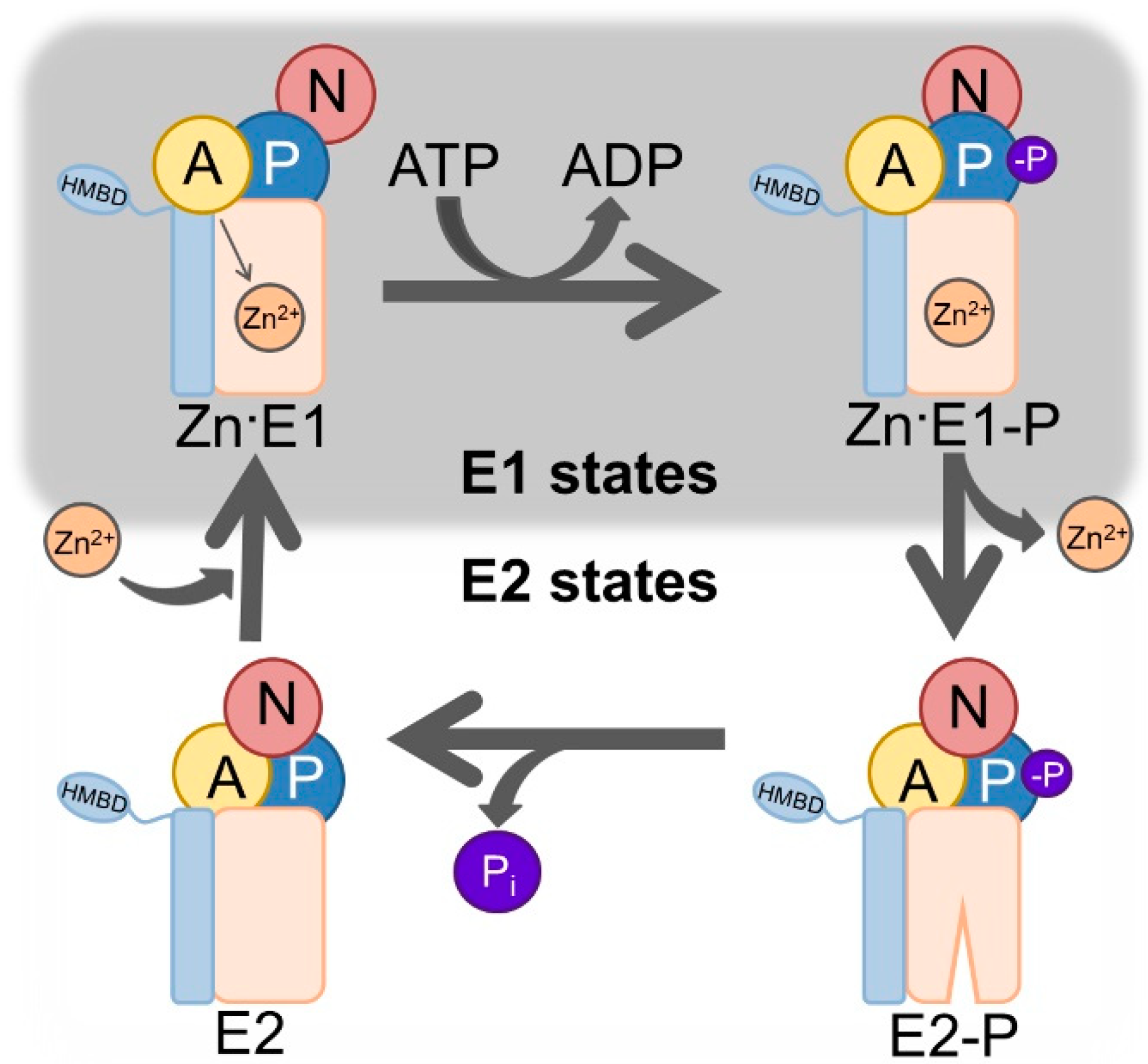
:1. Introduction
2. Materials and Methods
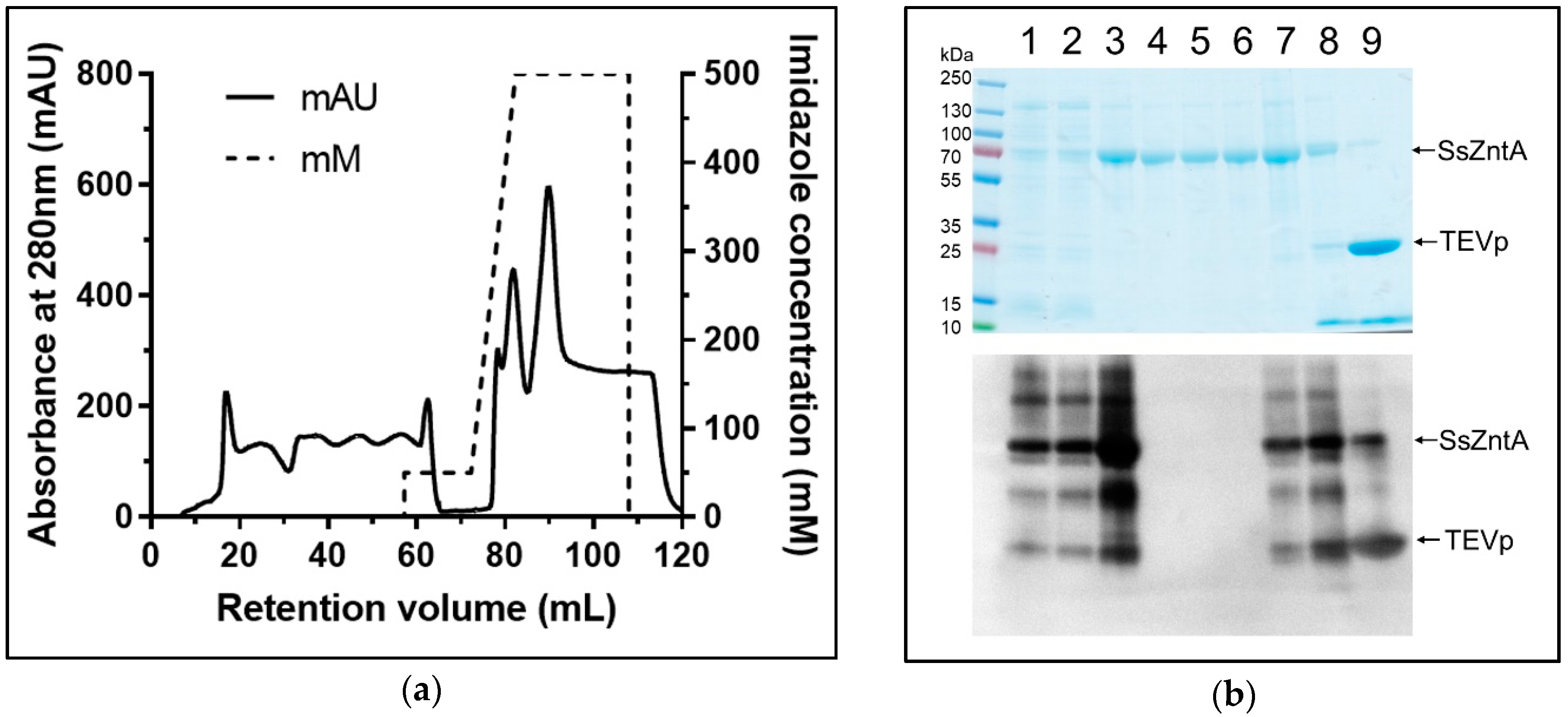
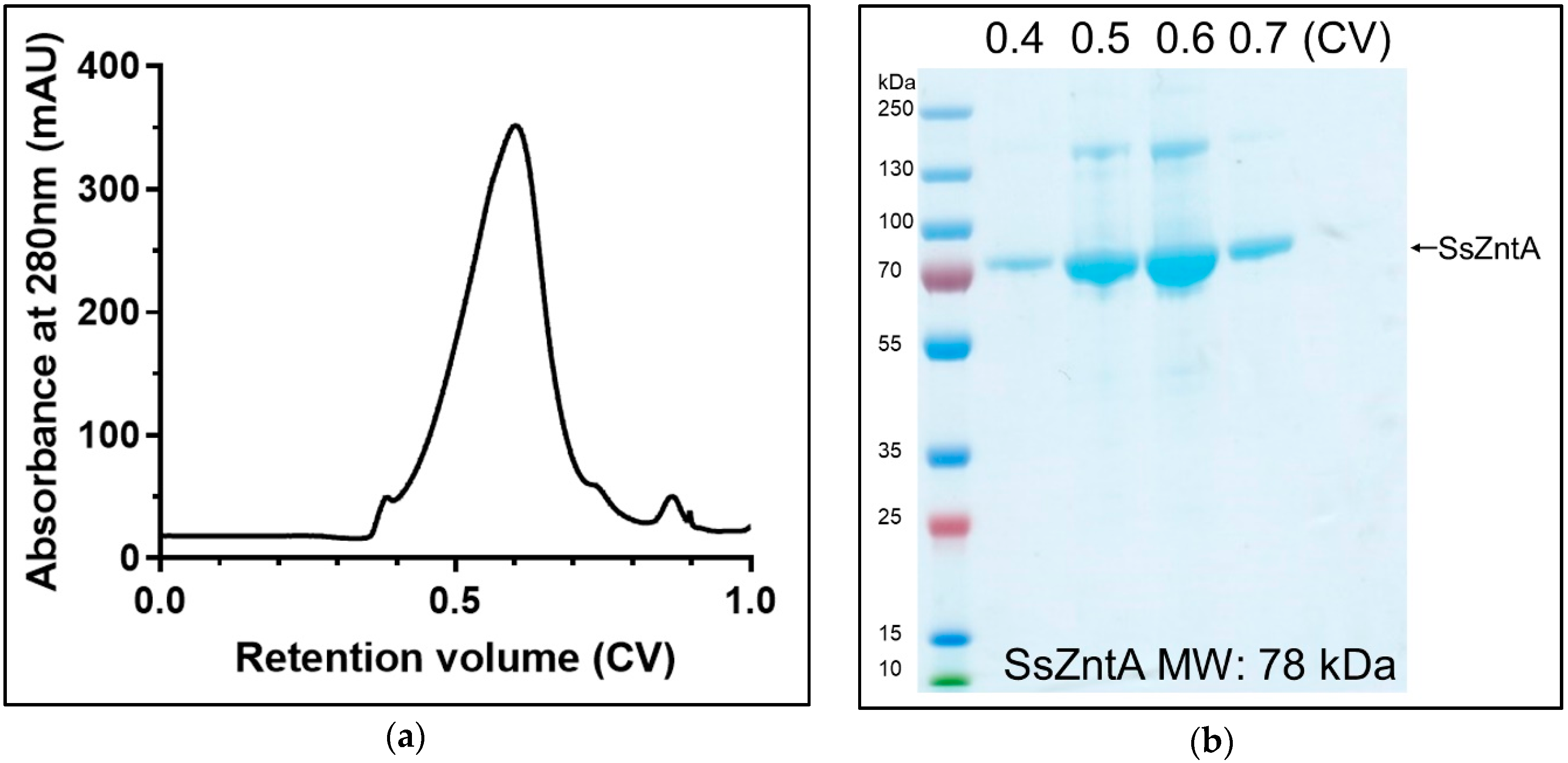
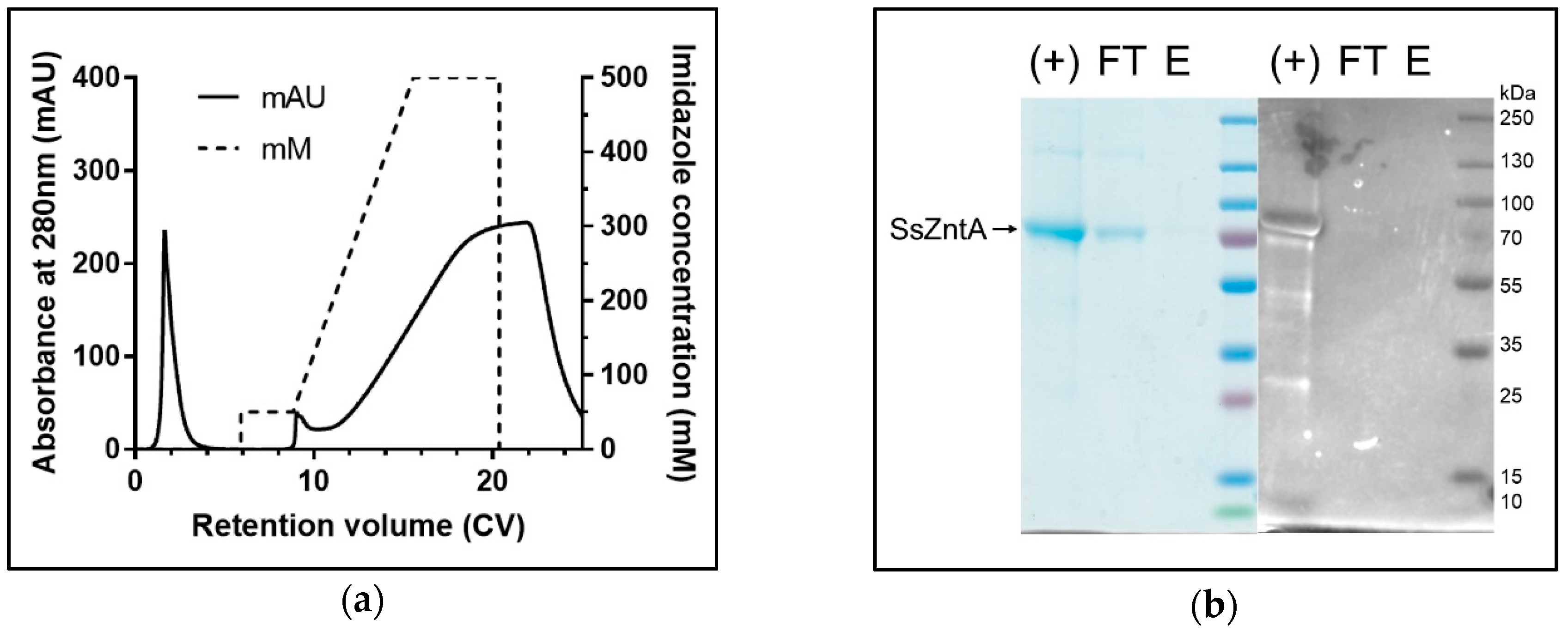
2.1. SsZntA Production
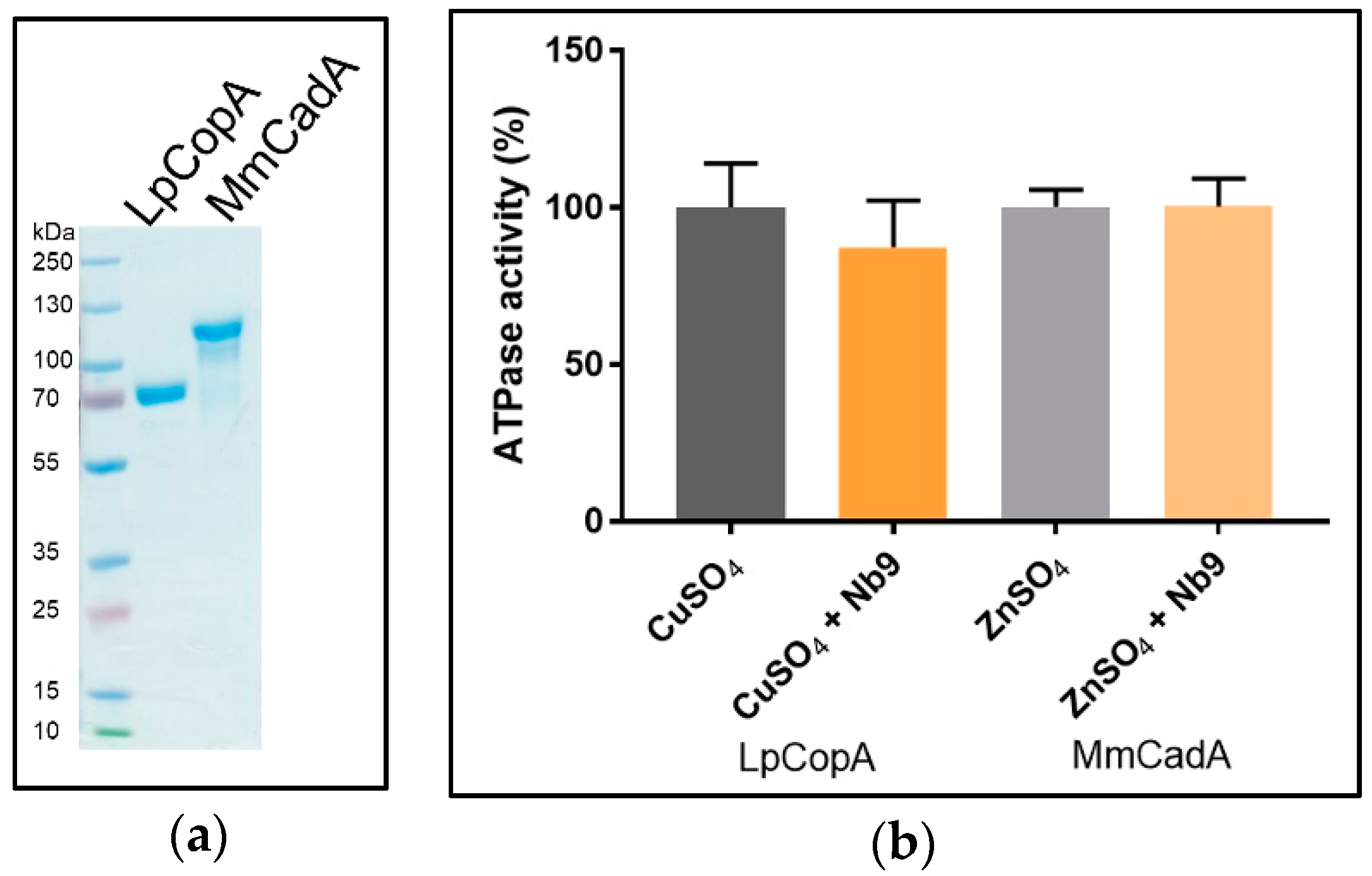
2.2. LpCopA and MmCadA Production
2.3. Llama Immunization and Nanobodies Identification
2.4. Nanobodies Sequence Analysis
2.5. Nanobodies Production
2.6. Functional Assay
2.6.1. SEC Co-Elution
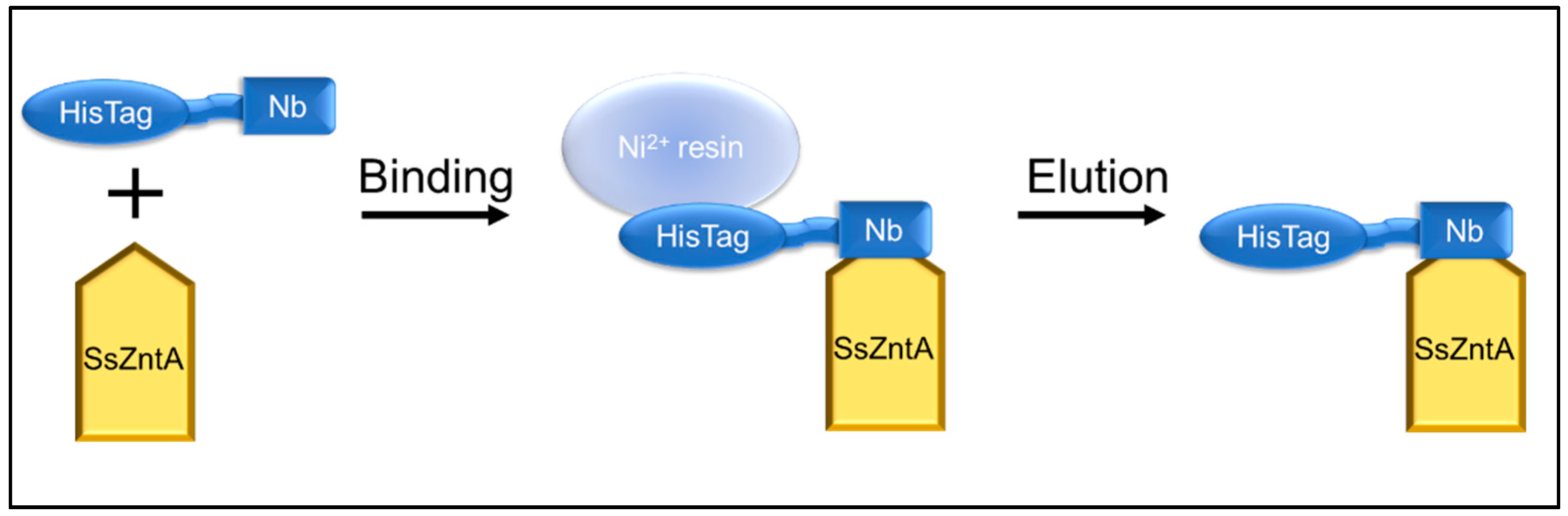
2.6.2. Ni-NTA Co-Elution
2.6.3. ATPase Activity Assay
3. Results
3.1. Isolation of Native SsZntA
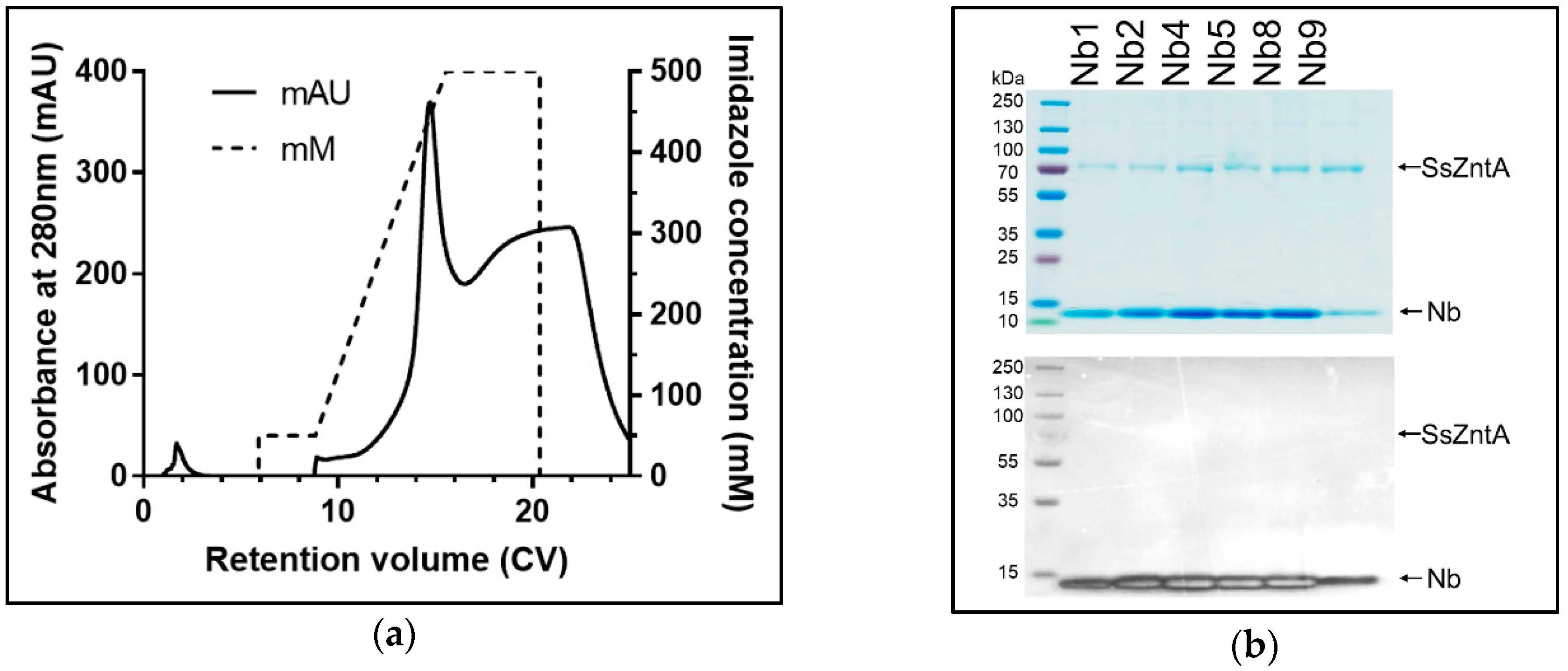
3.2. Isolation of Nanobodies
3.3. Size-Exclusion Chromatography (SEC) Co-Elution
3.4. Affinity Purification Co-Elution
3.5. Nb Effect on SsZntA Functionality
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Bublitz, M.; Morth, J.P.; Nissen, P. P-type ATPases at a glance. J. Cell Sci. 2011, 124, 2515–2519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Axelsen, K.B.; Palmgren, M.G. Evolution of substrate specificities in the P-type ATPase superfamily. J. Mol. Evol. 1998, 46, 84–101. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Sitsel, O.; Meloni, G.; Autzen, H.E.; Andersson, M.; Klymchuk, T.; Nielsen, A.M.; Rees, D.C.; Nissen, P.; Gourdon, P. Structure and mechanism of Zn2+-transporting P-type ATPases. Nature 2014, 514, 518. [Google Scholar] [CrossRef] [PubMed]
- Kühlbrandt, W. Biology, structure and mechanism of P-type ATPases. Nat. Rev. Mol. Cell. Biol. 2004, 5, 282. [Google Scholar] [CrossRef] [PubMed]
- Sitsel, O.; Duelli, A.; Gourdon, P. Zinc-Transporting P-Type ATPases. In Encyclopedia of Inorganic and Bioinorganic Chemistry; Scott, R.A., Ed.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2016. [Google Scholar]
- Post, R.L.; Sen, A.K.; Rosenthal, A.S. A phosphorylated intermediate in adenosine triphosphate-dependent sodium and potassium transport across kidney membranes. J. Biol. Chem. 1965, 240, 1437–1445. [Google Scholar] [PubMed]
- Albers, R.W. Biochemical Aspects of Active Transport. Annu. Rev. Biochem. 1967, 36, 727–756. [Google Scholar] [CrossRef] [PubMed]
- Apell, H.J. How do P-type ATPases transport ions? Bioelectrochemistry 2004, 63, 149–156. [Google Scholar] [CrossRef] [PubMed]
- De Meyer, T.; Muyldermans, S.; Depicker, A. Nanobody-based products as research and diagnostic tools. Trends Biotechnol. 2014, 32, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Hamers-Casterman, C.; Atarhouch, T.; Muyldermans, S.; Robinson, G.; Hamers, C.; Songa, E.B.; Bendahman, N.; Hamers, R. Naturally occurring antibodies devoid of light chains. Nature 1993, 363, 446–448. [Google Scholar] [CrossRef] [PubMed]
- Arbabi Ghahroudi, M.; Desmyter, A.; Wyns, L.; Hamers, R.; Muyldermans, S. Selection and identification of single domain antibody fragments from camel heavy-chain antibodies. FEBS Lett. 1997, 414, 521–526. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, T.G.M.; Skerra, A. The Strep-tag system for one-step purification and high-affinity detection or capturing of proteins. Nat. Protoc. 2007, 2, 1528. [Google Scholar] [CrossRef] [PubMed]
- Hansen, S.B.; Laursen, N.S.; Andersen, G.R.; Andersen, K.R. Introducing site-specific cysteines into nanobodies for mercury labelling allows de novo phasing of their crystal structures. Acta Crystallogr. D Struct. Biol. 2017, 73, 804–813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gouy, M.; Guindon, S.; Gascuel, O. SeaView Version 4: A Multiplatform Graphical User Interface for Sequence Alignment and Phylogenetic Tree Building. Mol. Biol. Evol. 2010, 27, 221–224. [Google Scholar] [CrossRef] [PubMed]
- Stothard, P. The Sequence Manipulation Suite: JavaScript Programs for Analyzing and Formatting Protein and DNA Sequences. BioTechniques 2000, 28, 1102–1104. [Google Scholar] [CrossRef] [PubMed]
- Leem, J.; Dunbar, J.; Georges, G.; Shi, J.; Deane, C.M. ABodyBuilder: Automated antibody structure prediction with data–driven accuracy estimation. mAbs 2016, 8, 1259–1268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuo, J.; Li, J.; Zhang, R.; Xu, L.; Chen, H.; Jia, X.; Su, Z.; Zhao, L.; Huang, X.; Xie, W. Institute collection and analysis of Nanobodies (iCAN): A comprehensive database and analysis platform for nanobodies. BMC Genomics 2017, 18, 797. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; Rensing, C.; Rosen, B.P.; Mitra, B. The ATP Hydrolytic Activity of Purified ZntA, a Pb(II)/Cd(II)/Zn(II)-translocating ATPase from Escherichia coli. J. Biol. Chem. 2000, 275, 3873–3878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, D.; Bruschweiler-Li, L.; Zhang, F.; Brüschweiler, R. Modulation and Functional Role of the Orientations of the N- and P-Domains of Cu+-Transporting ATPase along the Ion Transport Cycle. Biochemistry 2015, 54, 5095–5102. [Google Scholar] [CrossRef] [PubMed]
- Krasteva, M.; Barth, A. Structures of the Ca2+—ATPase complexes with ATP, AMPPCP and AMPPNP. An FTIR study. Biochim. Biophys. Acta. 2007, 1767, 114–123. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.; Wang, W.; Kim, R.; Yokota, H.; Damo, S.; Kim, S.-H.; Wemmer, D.; Kustu, S.; Yan, D. BeF3- acts as a phosphate analog in proteins phosphorylated on aspartate: Structure of a BeF3-complex with phosphoserine phosphatase. Proc. Nati. Acad. Sci. USA 2001, 98, 8525–8530. [Google Scholar] [CrossRef] [PubMed]
- Beghein, E.; Gettemans, J. Nanobody Technology: A Versatile Toolkit for Microscopic Imaging, Protein–Protein Interaction Analysis, and Protein Function Exploration. Front. Immunol. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Gutmann, D.A.P.; Mizohata, E.; Newstead, S.; Ferrandon, S.; Henderson, P.J.F.; van Veen, H.W.; Byrne, B. A high-throughput method for membrane protein solubility screening: The ultracentrifugation dispersity sedimentation assay. Protein Sci. 2007, 16, 1422–1428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersson, M.; Mattle, D.; Sitsel, O.; Klymchuk, T.; Nielsen, A.M.; Møller, L.B.; White, S.H.; Nissen, P.; Gourdon, P. Copper-transporting P-type ATPases use a unique ion-release pathway. Nat. Struct. Mol. Biol. 2014, 21, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Olesen, C.; Picard, M.; Winther, A.-M.L.; Gyrup, C.; Morth, J.P.; Oxvig, C.; Møller, J.V.; Nissen, P. The structural basis of calcium transport by the calcium pump. Nature 2007, 450, 1036. [Google Scholar] [CrossRef] [PubMed]
- Morth, J.P.; Pedersen, B.P.; Toustrup-Jensen, M.S.; Sørensen, T.L.M.; Petersen, J.; Andersen, J.P.; Vilsen, B.; Nissen, P. Crystal structure of the sodium–potassium pump. Nature 2007, 450, 1043. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tested Nb | Binding | Precipitation |
|---|---|---|
| Nb1 | + | - |
| Nb2 | + | - |
| Nb4 | + | - |
| Nb5 | + | - |
| Nb6 | N.D. | + |
| Nb7 | N.D. | + |
| Nb8 | + | - |
| Nb9 | + | - |
| Nb10 | N.D. | + |
| Nb11 | N.D. | + |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Longhin, E.; Grønberg, C.; Hu, Q.; Duelli, A.S.; Andersen, K.R.; Laursen, N.S.; Gourdon, P. Isolation and Characterization of Nanobodies against a Zinc-Transporting P-Type ATPase. Antibodies 2018, 7, 39. https://doi.org/10.3390/antib7040039
Longhin E, Grønberg C, Hu Q, Duelli AS, Andersen KR, Laursen NS, Gourdon P. Isolation and Characterization of Nanobodies against a Zinc-Transporting P-Type ATPase. Antibodies. 2018; 7(4):39. https://doi.org/10.3390/antib7040039
Chicago/Turabian StyleLonghin, Elena, Christina Grønberg, Qiaoxia Hu, Annette Susanne Duelli, Kasper Røjkjær Andersen, Nick Stub Laursen, and Pontus Gourdon. 2018. "Isolation and Characterization of Nanobodies against a Zinc-Transporting P-Type ATPase" Antibodies 7, no. 4: 39. https://doi.org/10.3390/antib7040039
APA StyleLonghin, E., Grønberg, C., Hu, Q., Duelli, A. S., Andersen, K. R., Laursen, N. S., & Gourdon, P. (2018). Isolation and Characterization of Nanobodies against a Zinc-Transporting P-Type ATPase. Antibodies, 7(4), 39. https://doi.org/10.3390/antib7040039