Construction of Antibody Phage Libraries and Their Application in Veterinary Immunovirology


Abstract
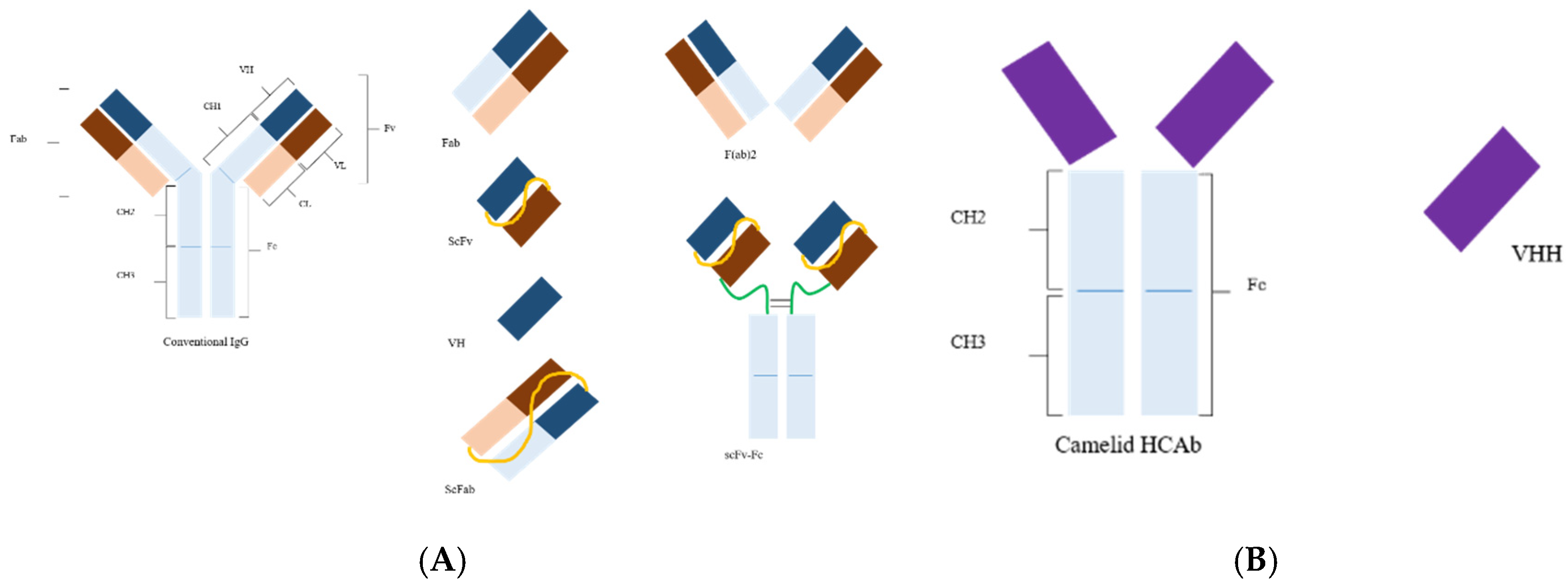
:1. Introduction
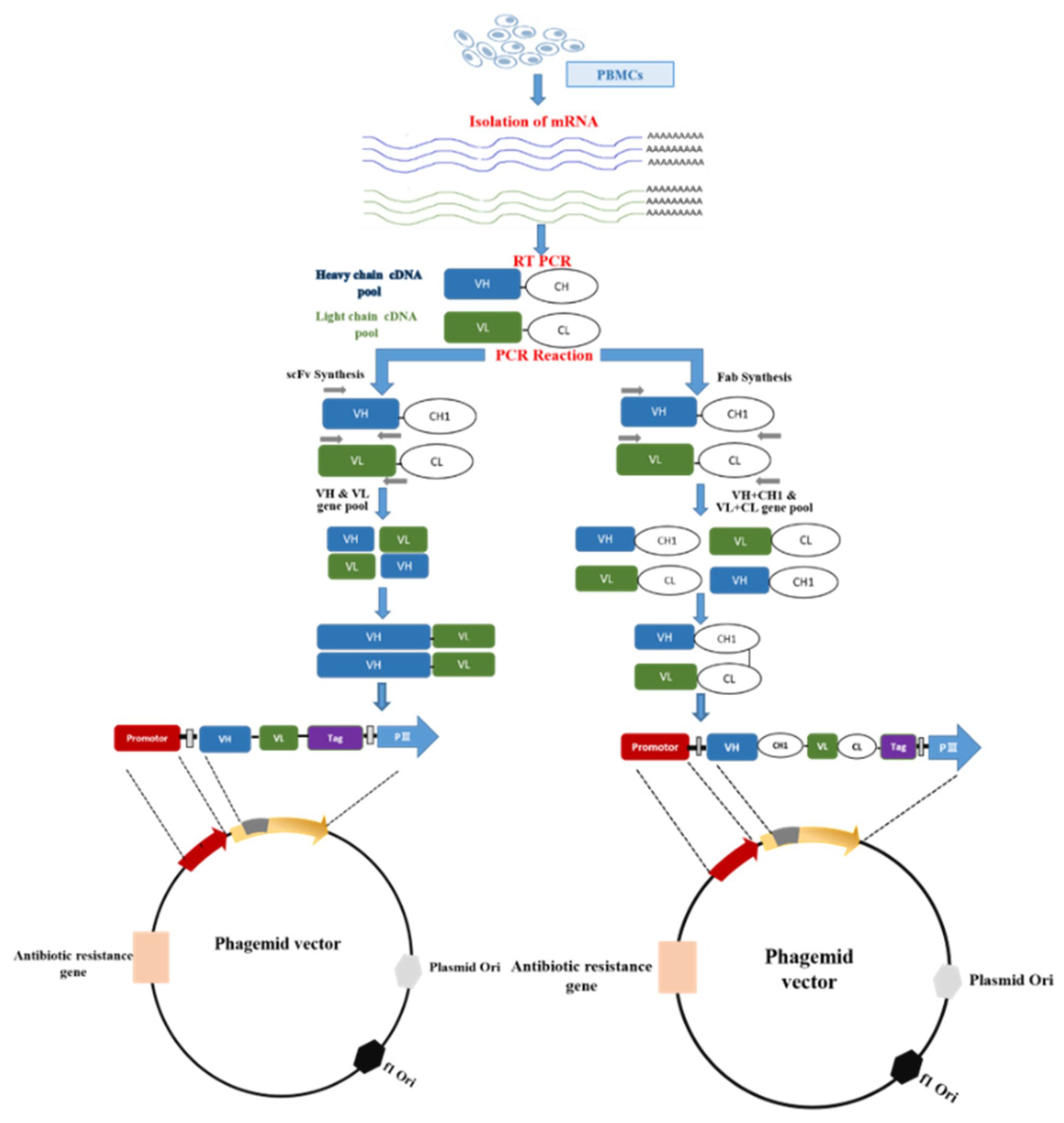
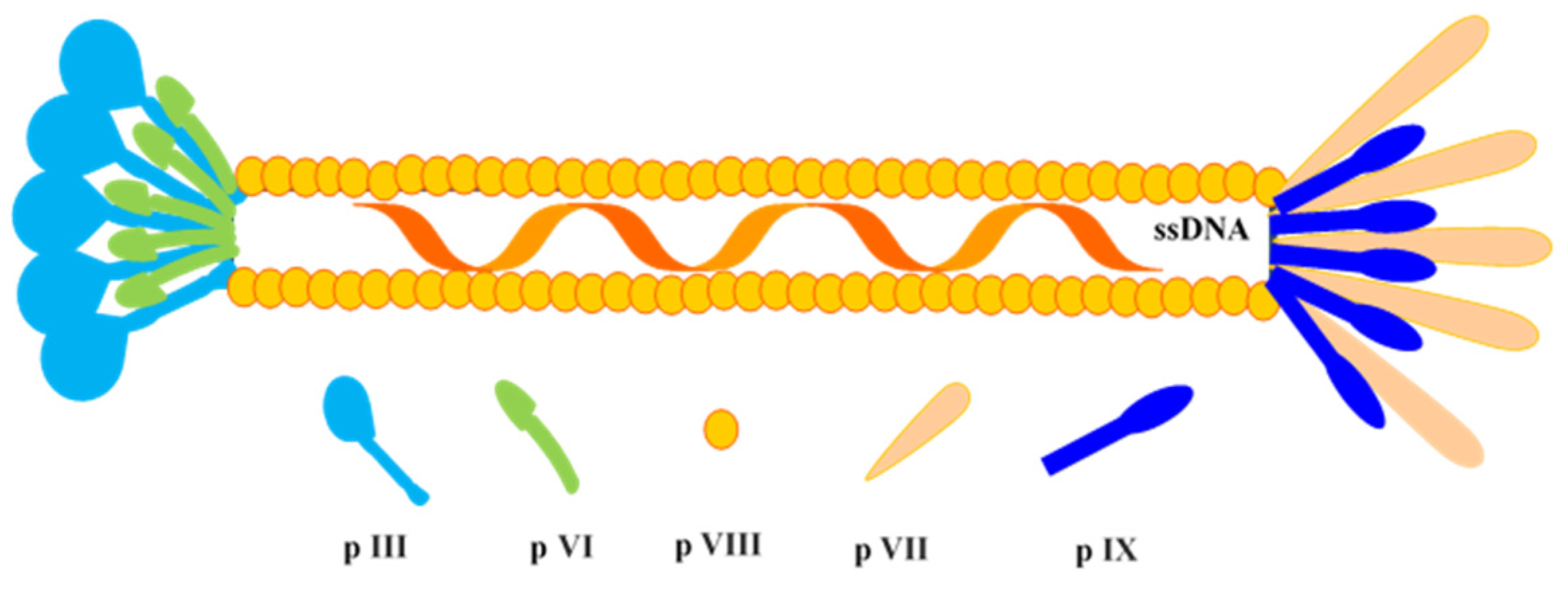
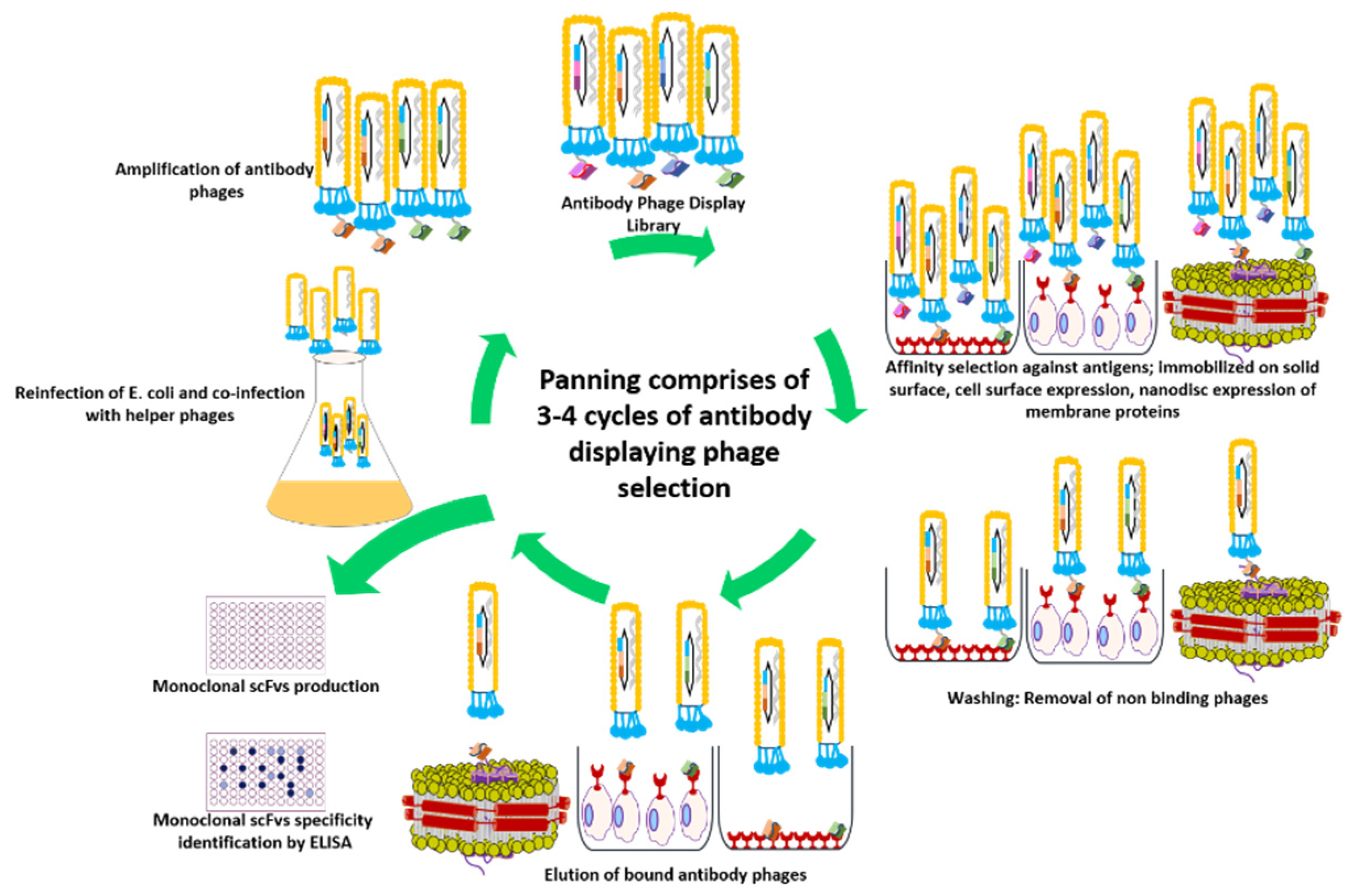
2. Antibody Phage Display Library Construction and Biopanning
Quality Assessment of an Antibody Library
3. Application of APD in Veterinary Immunovirology
3.1. Veterinary Diagnosis
3.2. Veterinary Therapeutics
3.3. Veterinary Immunoprophylaxis
4. Conclusions and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Hoogenboom, H.R.; De Brune, A.P.; Hufton, S.E.; Hoet, R.M.; Arends, J.W.; Roovers, R.C. Antibody phage display technology and its applications. Immunotechnology 1998, 4, 1–20. [Google Scholar] [CrossRef]
- Ahmad, Z.A.; Yeap, S.K.; Ali, A.M.; Ho, W.Y.; Alitheen, N.B.M.; Hamid, M. ScFv antibody: Principles and clinical application. Clin. Dev. Immunol. 2012, 2012, 1–15. [Google Scholar]
- Kuhn, P.; Fühner, V.; Unkauf, T.; Moreira, G.M.S.G.; Frenzel, A.; Miethe, S.; Hust, M. Recombinant antibodies for diagnostics and therapy against pathogens and toxins generated by phage display. Proteom.-Clin. Appl. 2016, 10, 922–948. [Google Scholar]
- Hairul Bahara, N.H.; Tye, G.J.; Choong, Y.S.; Ong, E.B.B.; Ismail, A.; Lim, T.S. Phage display antibodies for diagnostic applications. Biologicals 2013, 41, 209–216. [Google Scholar] [CrossRef]
- Burkovitz, A.; Ofran, Y. Understanding differences between synthetic and natural antibodies can help improve antibody engineering. MAbs 2016, 8, 278–287. [Google Scholar] [CrossRef] [Green Version]
- Tohidkia, M.R.; Barar, J.; Asadi, F.; Omidi, Y. Molecular considerations for development of phage antibody libraries. J. Drug Target. 2012, 20, 195–208. [Google Scholar] [CrossRef]
- Smith, G.P. Preface. Surface display and peptide libraries. Gene 1993, 128, 1–144. [Google Scholar] [CrossRef]
- Feng, J.N.; Model, P.; Russel, M. A trans-envelope protein complex needed for filamentous phage assembly and export. Mol. Microbiol. 1999, 34, 745–755. [Google Scholar] [CrossRef]
- Carlsson, F.; Trilling, M.; Perez, F.; Ohlin, M. A dimerized single-chain variable fragment system for the assessment of neutralizing activity of phage display-selected antibody fragments specific for cytomegalovirus. J. Immunol. Methods 2012, 376, 68–78. [Google Scholar] [CrossRef]
- Wen, K.; Nölke, G.; Schillberg, S.; Wang, Z.; Zhang, S.; Wu, C.; Jiang, H.; Meng, H.; Shen, J. Improved fluoroquinolone detection in ELISA through engineering of a broad-specific single-chain variable fragment binding simultaneously to 20 fluoroquinolones. Anal. Bioanal. Chem. 2012, 403, 2771–2783. [Google Scholar] [CrossRef]
- Kawasaki, T.; Onodera, K.; Kamijo, S. Identification of novel short peptide inhibitors of soluble 37/48 kDa oligomers of amyloid β42. Biosci. Biotechnol. Biochem. 2011, 75, 1496–1501. [Google Scholar] [CrossRef] [PubMed]
- Shimazaki, K.; Lepin, E.J.; Wei, B.; Nagy, A.K.; Coulam, C.P.; Mareninov, S.; Fu, M.; Wu, A.M.; Marks, J.D.; Braun, J.; et al. Diabodies targeting epithelial membrane protein 2 reduce tumorigenicity of human endometrial cancer cell lines. Clin. Cancer Res. 2008, 14, 7367–7377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azzazy, H.M.E.; Highsmith, W.E. Phage display technology: Clinical applications and recent innovations. Clin. Biochem. 2002, 35, 425–445. [Google Scholar] [CrossRef]
- Carmen, S.; Jermutus, L. Concepts in antibody phage display. Brief. Funct. Genom. Proteom. 2002, 1, 189–203. [Google Scholar] [CrossRef] [PubMed]
- Yuan, B.; Schulz, P.; Liu, R.; Sierks, M.R. Improved affinity selection using phage display technology and off-rate based selection. Electron. J. Biotechnol. 2006, 9. [Google Scholar] [CrossRef] [Green Version]
- Knappik, A.; Ge, L.; Honegger, A.; Pack, P.; Fischer, M.; Wellnhofer, G.; Hoess, A.; Wölle, J.; Plückthun, A.; Virnekäs, B. Fully synthetic human combinatorial antibody libraries (HuCAL) based on modular consensus frameworks and CDRs randomized with trinucleotides. J. Mol. Biol. 2000, 296, 57–86. [Google Scholar] [CrossRef]
- Sidhu, S.S.; Li, B.; Chen, Y.; Fellouse, F.A.; Eigenbrot, C.; Fuh, G. Phage-displayed antibody libraries of synthetic heavy chain complementarity determining regions. J. Mol. Biol. 2004, 338, 299–310. [Google Scholar] [CrossRef]
- McWhirter, J.R.; Kretz-Rommel, A.; Saven, A.; Maruyama, T.; Potter, K.M.; Mockridge, C.I.; Ravey, E.P.; Qin, F.; Bowdish, K.S. Antibodies selected from combinatorial libraries block a tumor antigen that plays a key role in immunomodulation. Proc. Natl. Acad. Sci. USA 2006, 103, 1041–1046. [Google Scholar] [CrossRef] [Green Version]
- Lomonosova, A.V.; Laman, A.G.; Fursova, K.K.; Shepelyakovskaya, A.O.; Vertiev, Y.V.; Brovko, F.A.; Grishin, E.V. Generation of scFv phages specific to Staphylococcus enterotoxin C1 by panning on related antigens. MAbs 2011, 3, 513–516. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.; Samanta, M.; Crawford, S.E.; Estes, M.K.; Neill, F.H.; Atmar, R.L.; Palzkill, T.; Burton, D. Identification of human single-chain antibodies with broad reactivity for noroviruses. Protein Eng. Des. Sel. 2014, 27, 339–349. [Google Scholar] [CrossRef]
- Blazek, D.; Celer, V.; Navrátilová, I.; Skládal, P. Generation and characterization of single-chain antibody fragments specific against transmembrane envelope glycoprotein gp46 of maedi-visna virus. J. Virol. Methods 2004, 115, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Osbourn, J.K.; Derbyshire, E.J.; Vaughan, T.J.; Field, A.W.; Johnson, K.S. Pathfinder selection: In situ isolation of novel antibodies. Immunotechnology 1998, 3, 293–302. [Google Scholar] [CrossRef]
- Hoogenboom, H.R.; Lutgerink, J.T.; Pelsers, M.M.A.L.; Rousch, M.J.M.M.; Coote, J.; Van Neer, N.; De Bruïne, A.; Van Nieuwenhoven, F.A.; Glatz, J.F.C.; Arends, J.W. Selection-dominant and nonaccessible epitopes on cell-surface receptors revealed by cell-panning with a large phage antibody library. Eur. J. Biochem. 1999, 260, 774–784. [Google Scholar] [CrossRef] [PubMed]
- Dominik, P.K.; Borowska, M.T.; Dalmas, O.; Kim, S.S.; Perozo, E.; Keenan, R.J.; Kossiakoff, A.A. Conformational Chaperones for Structural Studies of Membrane Proteins Using Antibody Phage Display with Nanodiscs. Structure 2016, 24, 300–309. [Google Scholar] [CrossRef] [Green Version]
- Pavlidou, M.; Hänel, K.; Möckel, L.; Willbold, D. Nanodiscs Allow Phage Display Selection for Ligands to Non-Linear Epitopes on Membrane Proteins. PLoS ONE 2013, 8, e72272. [Google Scholar] [CrossRef] [Green Version]
- Dominik, P.K.; Kossiakoff, A.A. Phage display selections for affinity reagents to membrane proteins in nanodiscs. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 2015. [Google Scholar]
- Jespersen, L.K.; Kuusinen, A.; Orellana, A.; Keinänen, K.; Engberg, J. Use of proteoliposomes to generate phage antibodies against native AMPA receptor. Eur. J. Biochem. 2000, 267, 1382–1389. [Google Scholar] [CrossRef] [Green Version]
- Thrane, S.; Janitzek, C.M.; Agerbæk, M.; Ditlev, S.B.; Resende, M.; Nielsen, M.A.; Theander, T.G.; Salanti, A.; Sander, A.F. A novel virus-like particle based vaccine platform displaying the placental malaria antigen VAR2CSA. PLoS ONE 2015, 10, e0143071. [Google Scholar] [CrossRef] [Green Version]
- Glanville, J.; D’Angelo, S.; Khan, T.A.; Reddy, S.T.; Naranjo, L.; Ferrara, F.; Bradbury, A.R.M. Deep sequencing in library selection projects: What insight does it bring? Curr. Opin. Struct. Biol. 2015, 33, 146–160. [Google Scholar] [CrossRef] [Green Version]
- Fischer, N. Sequencing antibody repertoires: The next generation. In MAbs; Taylor & Francis: Milton Park, Milton, UK; Abingdon, UK, 2011. [Google Scholar]
- Meli, G.; Visintin, M.; Cannistraci, I.; Cattaneo, A. Direct in Vivo Intracellular Selection of Conformation-sensitive Antibody Domains Targeting Alzheimer’s Amyloid-β Oligomers. J. Mol. Biol. 2009, 387, 584–606. [Google Scholar] [CrossRef]
- Tanaka, T.; Rabbitts, T.H. Protocol for the selection of single-domain antibody fragments by third generation intracellular antibody capture. Nat. Protoc. 2010, 5, 67. [Google Scholar] [CrossRef]
- Fantini, M.; Pandolfini, L.; Lisi, S.; Chirichella, M.; Arisi, I.; Terrigno, M.; Goracci, M.; Cremisi, F.; Cattaneo, A. Assessment of antibody library diversity through next generation sequencing and technical error compensation. PLoS ONE 2017, 12, e0177574. [Google Scholar] [CrossRef] [PubMed]
- Zinkernagel, R.M. Immunology taught by viruses. Science (80-.) 1996, 271, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, D.; Shaikh, A.; Bhatia, S.; Pateriya, A.K.; Khandia, R.; Sood, R.; Prakash, A.; Pattnaik, B.; Pradhan, H.K. Development of single-chain Fv against the nucleoprotein of type A influenza virus and its use in ELISA. J. Virol. Methods 2014, 208, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, K.; Yu, W.L.; Lin, M.; Davis, S.A.N.; Elmgren, C.; MacKenzie, R.; Tanha, J.; Li, S.; Dubuc, G.; Brown, E.G.; et al. Prototype single step lateral flow technology for detection of avian influenza virus and chicken antibody to avian influenza virus. J. Immunoass. Immunochem. 2007, 28, 307–318. [Google Scholar] [CrossRef]
- Li, Y.; Collins, M.S.; Whitelam, G.C.; Alexander, D.J. Rapid pathotyping of Newcastle disease virus using a single-chain Fv displayed on phage against the C-terminal end of the F2 polypeptide. Arch. Virol. 2002, 147, 2025–2037. [Google Scholar] [CrossRef]
- Li, B.; Ye, J.; Lin, Y.; Wang, M.; Jia, R.; Zhu, J. Selection and characterization of single-chain recombinant antibodies against phosphoprotein of newcastle disease virus. Biologicals 2014, 42, 285–289. [Google Scholar] [CrossRef]
- Sapats, S.I.; Trinidad, L.; Gould, G.; Heine, H.G.; Van Den Berg, T.P.; Eterradossi, N.; Jackwood, D.; Parede, L.; Toquin, D.; Ignjatovic, J. Chicken recombinant antibodies specific for very virulent infectious bursal disease virus. Arch. Virol. 2006, 151, 1551–1566. [Google Scholar] [CrossRef]
- Sapats, S.; Gould, G.; Trinidad, L.; Parede, L.H.; David, C.; Ignjatovic, J. An ELISA for detection of infectious bursal disease virus and differentiation of very virulent strains based on single chain recombinant chicken antibodies. Avian Pathol. 2005, 34, 449–455. [Google Scholar] [CrossRef] [Green Version]
- Xue, W.; Zhao, Q.; Li, P.; Zhang, R.; Lan, J.; Wang, J.; Yang, X.; Xie, Z.; Jiang, S. Identification and characterization of a novel nanobody against duck hepatitis A virus type 1. Virology 2019, 528, 101–109. [Google Scholar] [CrossRef]
- ShengFeng, C.; Ping, L.; Tao, S.; Xin, W.; GuoFeng, W. Construction, expression, purification, refold and activity assay of a specific scFv fragment against foot and mouth disease virus. Vet. Res. Commun. 2003, 27, 243–256. [Google Scholar] [CrossRef]
- Sharma, G.K.; Mahajan, S.; Matura, R.; Subramaniam, S.; Mohapatra, J.K.; Pattnaik, B. Production and characterization of single-chain antibody (scFv) against 3ABC non-structural protein in Escherichia coli for sero-diagnosis of Foot and Mouth Disease virus. Biologicals 2014, 42, 339–345. [Google Scholar] [CrossRef]
- Foord, A.J.; Muller, J.D.; Yu, M.; Wang, L.F.; Heine, H.G. Production and application of recombinant antibodies to foot-and-mouth disease virus non-structural protein 3ABC. J. Immunol. Methods 2007, 321, 142–151. [Google Scholar] [CrossRef]
- Salem, R.; El-Kholy, A.A.; Ibrahim, M. Eight novel single chain antibody fragments recognising VP2 of foot-and-mouth disease virus serotypes A, O, and SAT 2. Virology 2019, 533, 145–154. [Google Scholar] [CrossRef]
- Bhatia, S.; Gangil, R.; Gupta, D.S.; Sood, R.; Pradhan, H.K.; Dubey, S.C. Single-chain fragment variable antibody against the capsid protein of bovine immunodeficiency virus and its use in ELISA. J. Virol. Methods 2010, 167, 68–73. [Google Scholar] [CrossRef]
- Yang, S.; Shang, Y.; Yin, S.; Wang, D.; Cai, J.; Gong, Z.; Serge, M.; Liu, X. A phage-displayed single domain antibody fused to alkaline phosphatase for detection of porcine circovirus type 2. J. Virol. Methods 2015, 213, 84–92. [Google Scholar] [CrossRef]
- Yang, S.; Li, L.; Yin, S.; Shang, Y.; Khan, M.U.Z.; He, X.; Yuan, L.; Gao, X.; Liu, X.; Cai, J. Single-domain antibodies as promising experimental tools in imaging and isolation of porcine epidemic diarrhea virus. Appl. Microbiol. Biotechnol. 2018, 102, 8931–8942. [Google Scholar] [CrossRef]
- Chen, S.; Li, S.; Sun, H.; Li, Y.; Ji, S.; Song, K.; Zhang, L.; Luo, Y.; Sun, Y.; Ma, J.; et al. Expression and characterization of a recombinant porcinized antibody against the E2 protein of classical swine fever virus. Appl. Microbiol. Biotechnol. 2018, 102, 961–970. [Google Scholar] [CrossRef]
- Du, T.; Zhu, G.; Wu, X.; Fang, J.; Zhou, E.M. Biotinylated single-domain antibody-based blocking ELISA for detection of antibodies against swine influenza virus. Int. J. Nanomed. 2019, 14, 9337. [Google Scholar] [CrossRef] [Green Version]
- Bao, F.; Wang, L.; Zhao, X.; Lu, T.; Na, A.M.; Wang, X.; Cao, J.; Du, Y. Preparation and characterization of a single-domain antibody specific for the porcine epidemic diarrhea virus spike protein. AMB Express 2019, 9, 104. [Google Scholar] [CrossRef] [Green Version]
- Ma, Z.; Wang, T.; Li, Z.; Guo, X.; Tian, Y.; Li, Y.; Xiao, S. A novel biotinylated nanobody-based blocking ELISA for the rapid and sensitive clinical detection of porcine epidemic diarrhea virus. J. Nanobiotechnol. 2019, 17, 96. [Google Scholar] [CrossRef] [Green Version]
- Braganza, A.; Wallace, K.; Pell, L.; Parrish, C.R.; Siegel, D.L.; Mason, N.J. Generation and validation of canine single chain variable fragment phage display libraries. Vet. Immunol. Immunopathol. 2011, 139, 27–40. [Google Scholar] [CrossRef] [PubMed]
- Long, M.C.; Jager, S.; Mah, D.C.W.; Jebailey, L.; Mah, M.A.; Masri, S.A.; Nagata, L.P. Construction and characterization of a novel recombinant single-chain variable fragment antibody against Western equine encephalitis virus. Hybridoma 2000, 19, 1–3. [Google Scholar] [CrossRef]
- Duggan, J.M.; Coates, D.M.; Ulaeto, D.O. Isolation of single-chain antibody fragments against Venezuelan equine encephalomyelitis virus from two different immune sources. Viral Immunol. 2001, 14, 263–273. [Google Scholar] [CrossRef] [PubMed]
- Celer, V.; Blažek, D.; Navrátilová, I.; Skládal, P.; Blacklaws, B.; Bujdoso, R. Recombinant single-chain Fv antibodies that recognize the p25 protein of the maedi-visna virus. Folia Microbiol. (Praha) 2003, 48, 435. [Google Scholar] [CrossRef]
- Van Wyngaardt, W.; Malatji, T.; Mashau, C.; Fehrsen, J.; Jordaan, F.; Miltiadou, D.; du Plessis, D.H. A large semi-synthetic single-chain Fv phage display library based on chicken immunoglobulin genes. BMC Biotechnol. 2004, 4, 6. [Google Scholar] [CrossRef] [Green Version]
- Tutykhina, I.L.; Sedova, E.S.; Gribova, I.Y.; Ivanova, T.I.; Vasilev, L.A.; Rutovskaya, M.V.; Lysenko, A.A.; Shmarov, M.M.; Logunov, D.Y.; Naroditsky, B.S.; et al. Passive immunization with a recombinant adenovirus expressing an HA (H5)-specific single-domain antibody protects mice from lethal influenza infection. Antivir. Res. 2013, 97, 318–328. [Google Scholar] [CrossRef]
- Pitaksajjakul, P.; Lekcharoensuk, P.; Upragarin, N.; Barbas, C.F.; Ibrahim, M.S.; Ikuta, K.; Ramasoota, P. Fab MAbs specific to HA of influenza virus with H5N1 neutralizing activity selected from immunized chicken phage library. Biochem. Biophys. Res. Commun. 2010, 395, 496–501. [Google Scholar] [CrossRef]
- Wang, Y.; Cui, P.; Zhu, S.; Meng, T.; Hao, F.; Zhu, G.; Zuo, W. Preparation of single-chain antibody against VP3 protein of duck hepatitis virus type 1 by phage display technology. J. Virol. Methods 2018, 257, 73–78. [Google Scholar] [CrossRef]
- Duan, H.; Ma, Z.; Xu, L.; Zhang, A.; Li, Z.; Xiao, S. A novel intracellularly expressed NS5B-specific nanobody suppresses bovine viral diarrhea virus replication. Vet. Microbiol. 2019, 240, 108449. [Google Scholar] [CrossRef]
- Li, T.; Huang, M.; Xiao, H.; Zhang, G.; Ding, J.; Wu, P.; Zhang, H.; Sheng, J.; Chen, C. Selection and characterization of specific nanobody against bovine virus diarrhea virus (BVDV) E2 protein. PLoS ONE 2017, 12, e0178469. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Chen, Y.; Yang, L.; Zhu, J. Construction and characterization of porcine single-chain fragment variable antibodies that neutralize transmissible gastroenteritis virus in vitro. Arch. Virol. 2019, 164, 983–994. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Liang, C.; Duan, H.; Zhang, X.; Wang, X.; Xiao, S.; Zhou, E.M. Intracellularly expressed nanobodies against non-structural protein 4 of porcine reproductive and respiratory syndrome virus inhibit virus replication. Biotechnol. Lett. 2016, 38, 1081–1088. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wang, Y.; Duan, H.; Zhang, A.; Liang, C.; Gao, J.; Zhang, C.; Huang, B.; Li, Q.; Li, N.; et al. An intracellularly expressed Nsp9-specific nanobody in MARC-145 cells inhibits porcine reproductive and respiratory syndrome virus replication. Vet. Microbiol. 2015, 181, 252–260. [Google Scholar] [CrossRef] [PubMed]
- Boruah, B.M.; Liu, D.; Ye, D.; Gu, T.-j.; Jiang, C.-l.; Qu, M.; Wright, E.; Wang, W.; He, W.; Liu, C.; et al. Single Domain Antibody Multimers Confer Protection against Rabies Infection. PLoS ONE 2013, 8, e71383. [Google Scholar] [CrossRef]
- Molinková, D.; Skládal, P.; Celer, V. In vitro neutralization of equid herpesvirus 1 mediated by recombinant antibodies. J. Immunol. Methods 2008, 333, 186–191. [Google Scholar] [CrossRef]
- Harmsen, M.M.; van Solt, C.B.; Fijten, H.P.D.; van Keulen, L.; Rosalia, R.A.; Weerdmeester, K.; Cornelissen, A.H.M.; De Bruin, M.G.M.; Eblé, P.L.; Dekker, A. Passive immunization of guinea pigs with llama single-domain antibody fragments against foot-and-mouth disease. Vet. Microbiol. 2007, 120, 193–206. [Google Scholar] [CrossRef]
- Harmsen, M.M.; Fijten, H.P.D.; Engel, B.; Dekker, A.; Eblé, P.L. Passive immunization with llama single-domain antibody fragments reduces foot-and-mouth disease transmission between pigs. Vaccine 2009, 27, 1904–1911. [Google Scholar] [CrossRef]
- Zhang, F.; Chen, Y.; Ke, Y.; Zhang, L.; Zhang, B.; Yang, L.; Zhu, J. Single chain fragment variable (scFv) antibodies targeting the spike protein of porcine epidemic diarrhea virus provide protection against viral infection in piglets. Viruses 2019, 11, 58. [Google Scholar] [CrossRef] [Green Version]
- Griffiths, A.D.; Williams, S.C.; Hartley, O.; Tomlinson, I.M.; Waterhouse, P.; Crosby, W.L.; Kontermann, R.E.; Jones, P.T.; Low, N.M.; Allison, T.J. Isolation of high affinity human antibodies directly from large synthetic repertoires. EMBO J. 1994, 13, 3245–3260. [Google Scholar] [CrossRef] [Green Version]
- Jain, S.; Aresu, L.; Comazzi, S.; Shi, J.; Worrall, E.; Clayton, J.; Humphries, W.; Hemmington, S.; Davis, P.; Murray, E.; et al. The development of a recombinant scFv monoclonal antibody targeting canine CD20 for use in comparative medicine. PLoS ONE 2016, 11, e0148366. [Google Scholar] [CrossRef]
- Ecker, D.M.; Jones, S.D.; Levine, H.L. The therapeutic monoclonal antibody market. In MAbs; Taylor & Francis: Milton Park, Milton, UK; Abingdon, UK, 2015. [Google Scholar]
- Deal, C.E.; Balazs, A.B. Vectored antibody gene delivery for the prevention or treatment of HIV infection. Curr. Opin. HIV AIDS 2015, 10, 190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strohl, W.R. Current progress in innovative engineered antibodies. Protein Cell 2018, 9, 86–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Application of Antibody | Antibody Format | Source Species | Target | Reference |
|---|---|---|---|---|
| Diagnosis | scFv | Mouse | Recombinant NP protein of avian influenza virus in poultry | [35] |
| VHH | Camelid | NP protein of AIV in poultry | [36] | |
| scFv | Rabbit | F2 protein of Newcastle disease virus in poultry | [37] | |
| scFv | Chicken | Phosphoprotein of Newcastle disease virus in poultry | [38] | |
| scFv | Chicken | Whole infectious bursal disease virus in poultry | [39,40] | |
| VHH | Camelid | VP1 protein of duck hepatitis A virus-1 in ducks | [41] | |
| scFv | Mouse | Foot and mouth disease virus type O in cows | [42] | |
| scFv | Chicken | 3ABC protein of foot and mouth disease virus in cows | [43,44] | |
| scFv | Mouse | VP2 of protein of foot and mouth disease virus in cows | [45] | |
| scFv | Mouse | Capsid protein of bovine immunodeficiency virus in cows | [46] | |
| VHH | Camelid | Capsid protein of porcine circovirus type in pigs | [47] | |
| VHH | Camelid | Membrane proteins of porcine epidemic diarrhea virus in pigs | [48] | |
| scFv | Mouse | E2 protein of classical swine fever virus in pigs | [49] | |
| VHH | Camelid | P protein of, swine influenza virus in pigs | [50] | |
| VHH | Camelid | N protein and spike protein of porcine epidemic diarrhea virus | [51,52] | |
| scFv | Dog | Capsid protein of canine parvovirus in dogs | [53] | |
| scFv | Mouse | Whole Western equine encephalitis virus | [54] | |
| scFv | Mouse | Whole Venezuelan equine encephalitis virus | [55] | |
| scFv | Human * | transmembrane envelope glycoprotein gp46 of Maedi-visna virus in sheep | [21] | |
| scFv | Human * | P25 of Maedi-visna virus in sheep | [56] | |
| scFv | Chicken | Whole blue-tongue virus in sheep | [57] | |
| Therapy | VHH | Camelid | HA antigen of H5N1 in poultry | [58] |
| Fab | Chicken | HA0 hemagglutinin of H5N1 in poultry | [59] | |
| scFv | Mouse | VP3 protein duck hepatitis A virus-1 in ducks | [60] | |
| VHH | Camelid | NS5B protein of bovine viral diarrhea virus in cows | [61] | |
| VHH | Camelid | E2 protein bovine viral diarrhea virus in cows | [62] | |
| scFv | Pig | Whole transmissible gastroenteritis virus in pigs | [63] | |
| VHH | Camelid | non-structural protein 4 of porcine reproductive and respiratory syndrome virus in pigs | [64] | |
| VHH | Camelid | non-structural protein 9 of porcine reproductive and respiratory syndrome virus in pigs | [65] | |
| VHH | Camelid | Whole rabies virus in dogs | [66] | |
| scFv | Human * | gD protein of equid herpesvirus-1 in horses | [67] | |
| Immunoprophylaxis | VHH | Camelid | Whole food and mouth virus type O in cows | [68] |
| VHH | Camelid | Whole food and mouth virus in cows pigs | [69] | |
| scFv | Pig | Spike protein of porcine epidemic diarrhea virus in pigs | [70] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bashir, S.; Paeshuyse, J. Construction of Antibody Phage Libraries and Their Application in Veterinary Immunovirology. Antibodies 2020, 9, 21. https://doi.org/10.3390/antib9020021
Bashir S, Paeshuyse J. Construction of Antibody Phage Libraries and Their Application in Veterinary Immunovirology. Antibodies. 2020; 9(2):21. https://doi.org/10.3390/antib9020021
Chicago/Turabian StyleBashir, Shahbaz, and Jan Paeshuyse. 2020. "Construction of Antibody Phage Libraries and Their Application in Veterinary Immunovirology" Antibodies 9, no. 2: 21. https://doi.org/10.3390/antib9020021
APA StyleBashir, S., & Paeshuyse, J. (2020). Construction of Antibody Phage Libraries and Their Application in Veterinary Immunovirology. Antibodies, 9(2), 21. https://doi.org/10.3390/antib9020021