Chemical Basis of Biological Homochirality during the Abiotic Evolution Stages on Earth


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
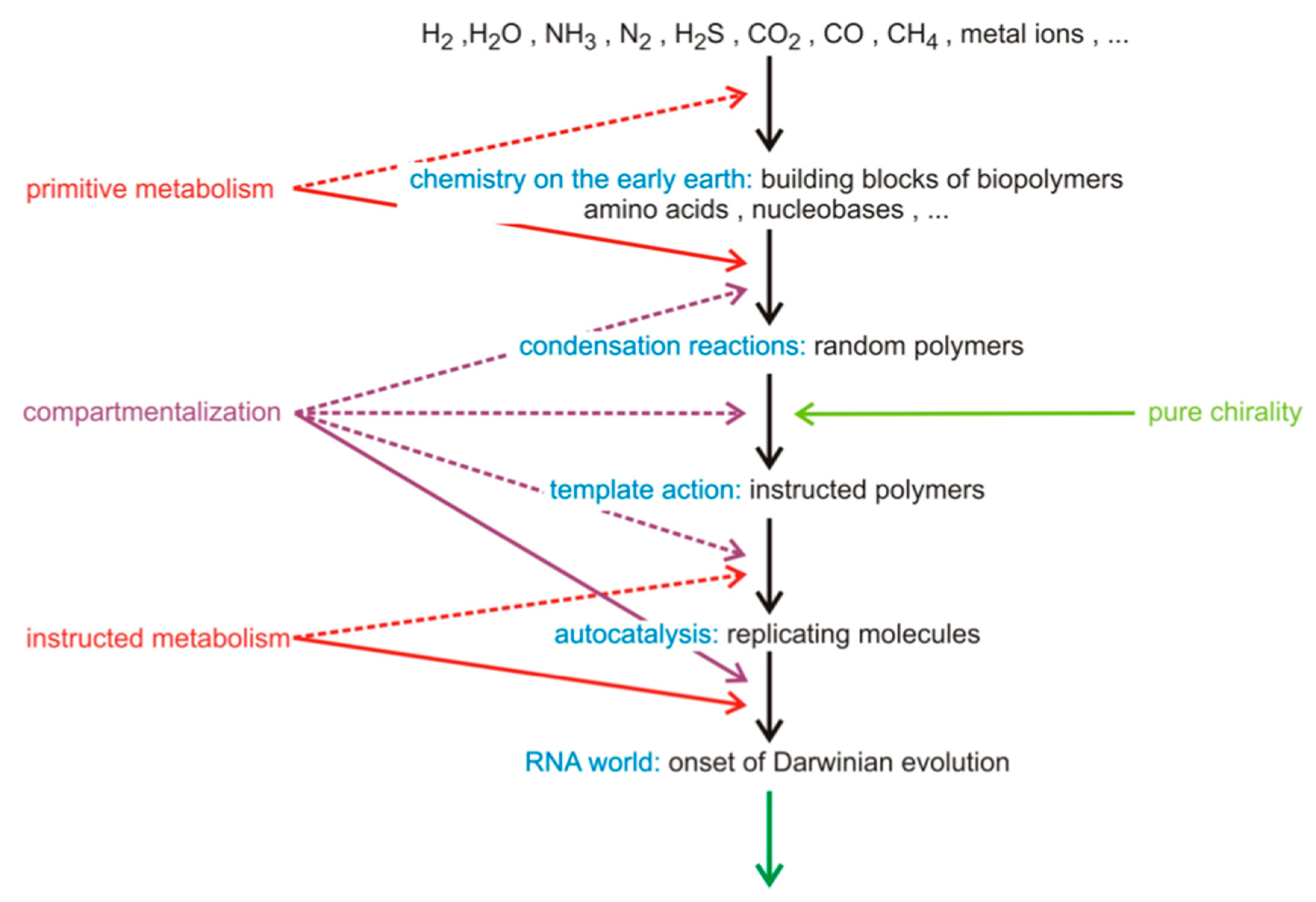
:1. Introduction
2. Racemization and Racemic Mixtures
3. Stable and Unstable Non-Equilibrium Stationary States (NESS) in Enantioselective Reactions
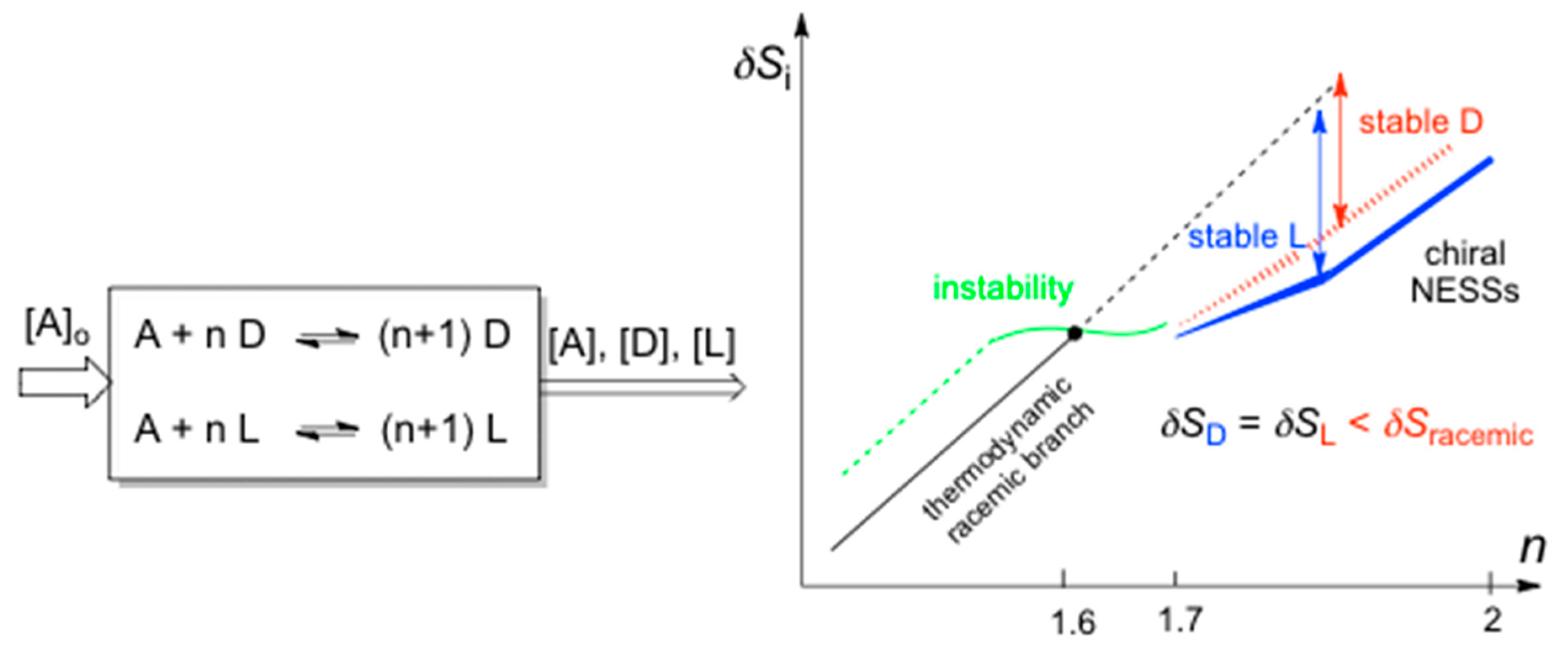
3.1. Entropy Production and Balance in Open Systems
3.2. Potential Reaction Networks Able to Yield SMSB.
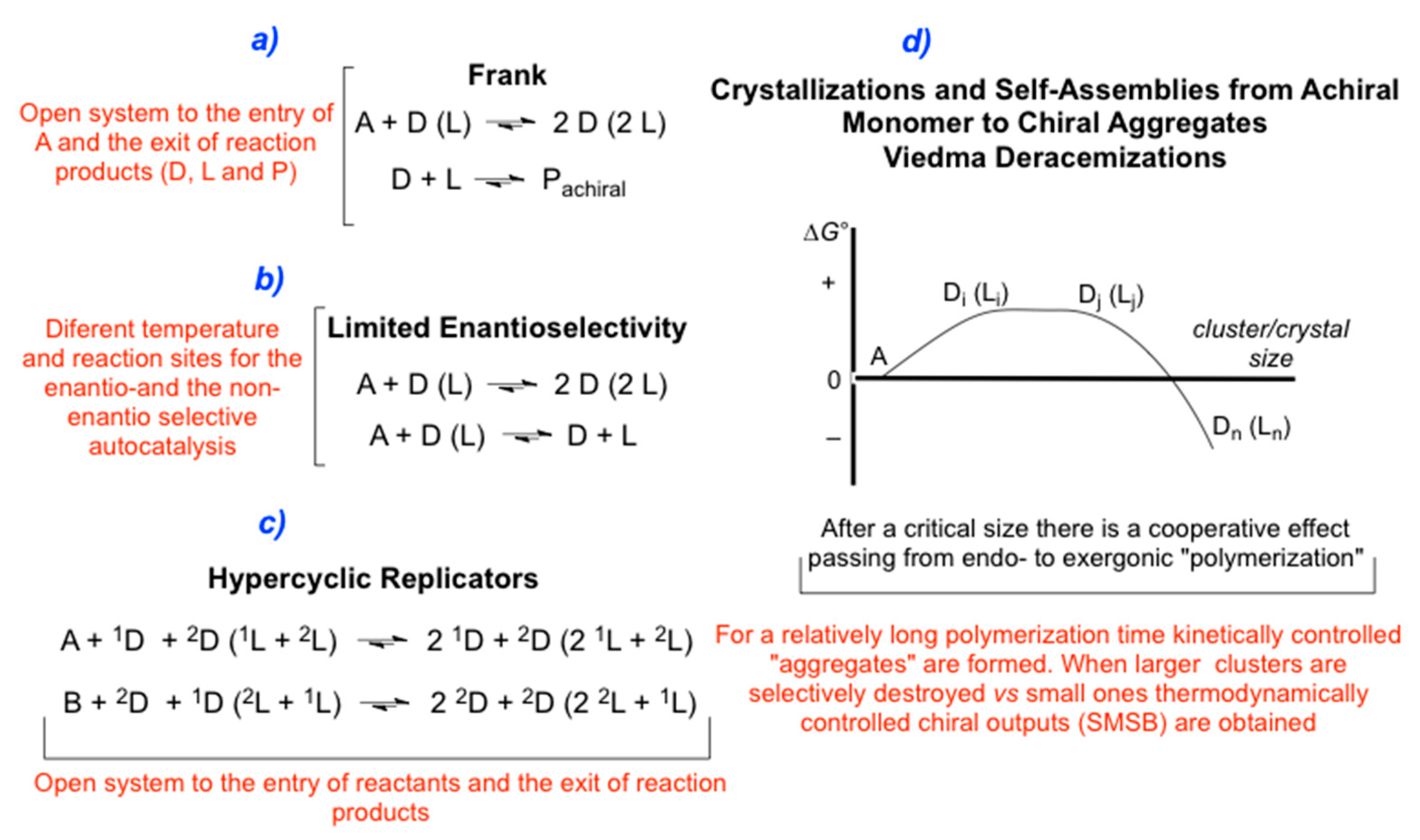
3.2.1. Frank-like Models
3.2.2. Limited Enantioselective Model
3.2.3. Enantioselective Hypercyclic Replicators
3.2.4. SMSB in Enantioselective Autocatalytic Polymerization/Depolymerization
- (a)
- In spite of being a closed system, the energy input is given selectively to only some of the species of the system. In other words, the mechanical grinding (the energy input) affects only the largest crystals.
- (b)
- The higher solubility of the smaller crystals obtained by grinding creates supersaturated solutions for the larger crystals and in consequence, a constant and permanent cycle of solubilization and crystal growth is maintained.
- (c)
- Homochiral cluster-to-cluster growth (this is equivalent—through its dynamic signature—to first order enantioselective autocatalysis). Notice that in saturated solutions monomer-to-cluster exchange between solution and crystal/cluster is not autocatalytic and therefore is unable to lead to any chiral amplification, nor to racemization, of the crystal mixture ee value.
- (d)
- Additional growth dynamics to first order autocatalysis of the cluster-to-cluster growth in (c) that increase the dynamic growth signature are provided by a mutualistic effect in the growth of homochiral material; each cluster coming from the fragmentation of large crystals can react with crystals of many different sizes leading to the formation of larger cluster/crystals of many different sizes, but of the same chiral sign. Furthermore, the free energy profile of the polymer formation, which is endergonic during the first “polymerization” steps, with cooperative growth beyond a critical size as well the intermediate sizes of the clusters coming from the breakage of the large ones, are decisive features for the achievement of the growth dynamics leading to SMSB [70].
3.2.5. On the Detection of SMSB in Low Exergonic Reactions in Solution
4. BH Based on SMSB Requires Previous Asymmetric Synthetic Scenarios: Stochastic vs. Deterministic Chiral Signs in SMSB
5. Hypothesis on the Emergence of BH
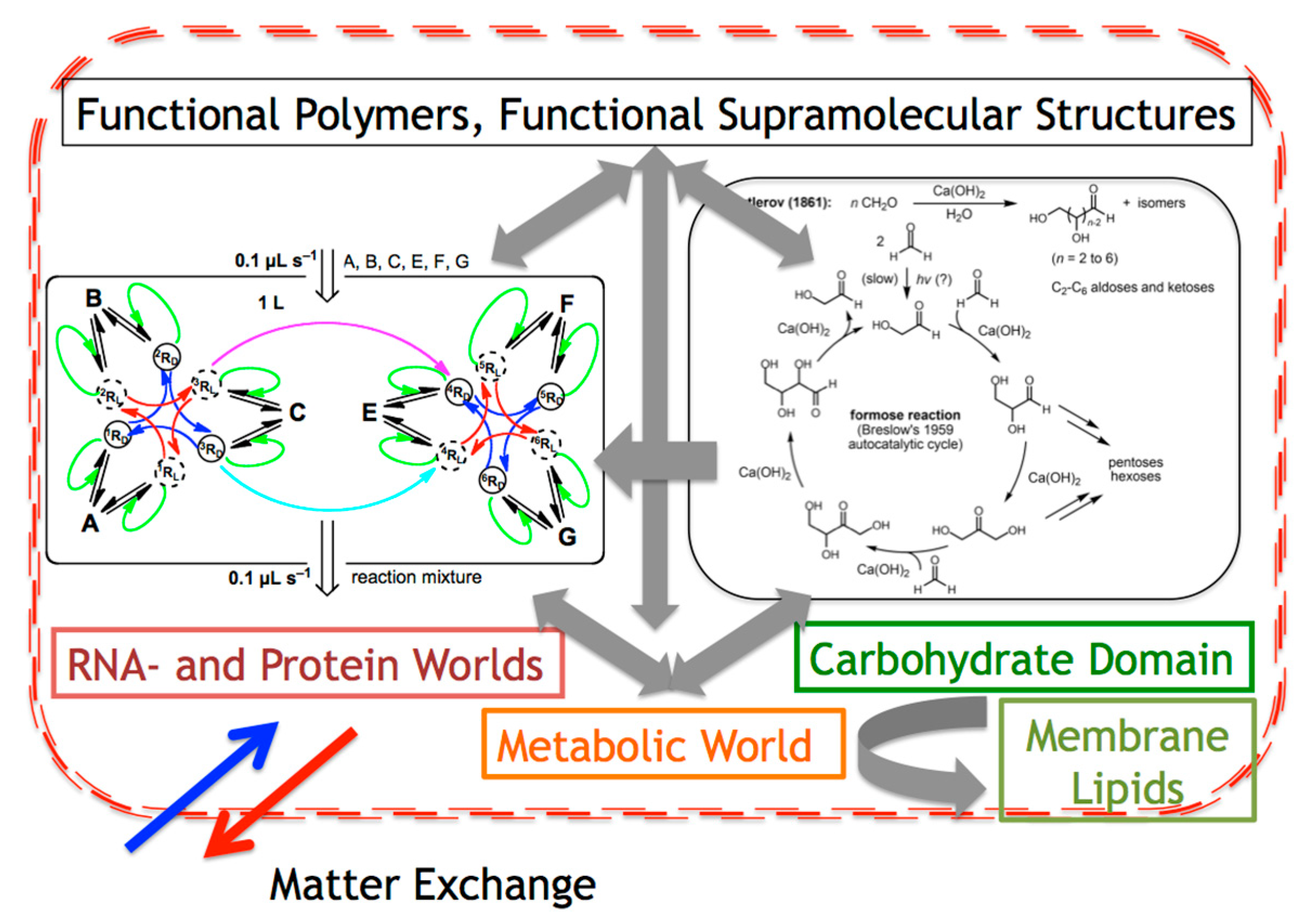
5.1. Is Carbohydrate Synthesis the Third Leg of a Tripodal SMSB Scenario?
5.2. Enantioselective Linear Reactions in the Chiral Machinery of Life
6. Conclusions
- (a)
- BH has its physico-chemical basis in enantioselective autocatalytic reaction networks operating in open dissipative systems. This thermodynamic scenario is similar to that proposed for the compartmentalization stages of chemical evolution where the question of chirality has been avoided up to the present. Moreover, the reaction networks proposed for the formation of pre-RNA worlds and peptides showing catalytic and autocatalytic functionalities and able to develop Darwinian evolution, also possess the ability for SMSB, i.e. of the selection between enantiomers.
- (b)
- Asymmetric inductions originating through natural chiral forces during previous stages of chemical evolution, as for example in astrophysical scenarios, could provide the necessary chiral polarization to transform, at later stages of chemical evolution, the stochastic chiral sign outcome of the SMSB processes into a deterministic one. This is in spite of the unavoidable racemization processes acting during the long-time interval of chemical evolution.
- (c)
- SMSB autocatalytic cycles provide the resilience against racemization characteristic of BH and would drive the rest of the enantioselective reactions towards a definite chiral sign.
- (d)
- Such a general scenario of BH may be applied to all current models, which aim to find the links between prebiotic chemistry and biological chemistry [97]. The emergence of BH is concurrent with the emergence of autocatalytic sets, proto-cells, etc., and it probably represents an energetic advantage (lower entropy production) [51], with respect to the racemic outcome.
- (e)
- Homochirality has been considered as an inherent property of matter [98]. From a chemical point of view, it is probably also a necessary condition for life because of the informational entropy advantage of asymmetry in molecular recognition, that is, in the emergence of catalytic and autocatalytic functionalities. Once the reasonable existence of SMSB in the decisive stages of chemical evolution is recognized, in our opinion, then the models concerning the emergence of autocatalytic sets, proto-cells, etc., cannot avoid the question of the enantioselectivity of the processes involved. Note that this also has direct consequences for the applied biotechnological fields of new metabolic cycles and of artificial cells.
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Guijarro, L.; Yus, M. The Origin of Chirality in the Molecules of Life: A Revision from Awareness to the Current; RSC Publishing: Cambridge, UK, 2009. [Google Scholar]
- Gal, J.; Cintas, P. Early History of the Recognition of Molecular Biochirality. In Biochirality; Springer: Berlin/Heidelberg, Germany, 2013; Volume 333, pp. 1–40. [Google Scholar]
- Blackmond, D.G. The origin of biological homochirality. Cold Spring Harb Perspect. Biol. 2010, 2, a002147. [Google Scholar] [CrossRef]
- Davankov, V.A. Biological homochirality on the earth, or in the universe? A selective review. Symmetry 2018, 10, 749. [Google Scholar] [CrossRef]
- An Evolutionary Mystery: Mirror Asymmetry in Life and in Space. Available online: https://www.acs.org/content/acs/en/acs-webinars/popular-chemistry/homochirality.html (accessed on 20 February 2019).
- Origin of Life. The Chirality Problem. Available online: https://creation.com/origin-of-life-the-chirality-problem (accessed on 20 February 2019).
- Soai, K.; Kawasaki, T. Asymmetric autocatalysis with amplification of chirality. In Amplification of Chirality; Springer: Berlin/Heidelberg, Germany, 2008; Volume 284, pp. 1–33. [Google Scholar]
- Kawasaki, T.; Matsumura, Y.; Tsutsumi, T.; Suzuki, K.; Ito, M.; Soai, K. Asymmetric autocatalysis triggered by carbon Isotope (13C/12C) chirality. Science 2009, 324, 492–495. [Google Scholar] [CrossRef]
- Viedma, C. Chiral symmetry breaking during crystallization: Complete chiral purity induced by non-linear autocatalysis. Phys. Rev. Lett. 2005, 94, 065504. [Google Scholar] [CrossRef]
- Glansdorff, P.; Prigogine, I. Thermodynamic Theory of Structure, Stability and Fluctuations; Wiley-Interscience: London, UK, 1971. [Google Scholar]
- Nicolis, G.; Prigogine, I. Self-Organization in Nonequilibrium Systems; Wiley: New York, NY, USA, 1977. [Google Scholar]
- Ruiz-Mirazo, K.; Briones, C.; de la Escosura, A. Prebiotic systems chemistry: New perspectives for the origins of life. Chem. Rev. 2014, 114, 285–366. [Google Scholar] [CrossRef]
- Kauffman, S. Autocatalytic sets of proteins. J. Theor. Biol. 1986, 119, 1–24. [Google Scholar] [CrossRef]
- Eigen, M.; Schuster, P. The Hypercycle: A Principle of Natural Self-Organization; Springer: Berlin, Germany, 1979. [Google Scholar]
- Hordijk, W.; Hein, J.; Steel, M. Autocatalytic sets and the origin of life. Entropy 2010, 12, 1733–1742. [Google Scholar] [CrossRef]
- Lifson, S. On the crucial stages in the origin of animate matter. J. Molec. Evol. 1997, 44, 1–8. [Google Scholar] [CrossRef]
- Schneider, E.D.; Kay, J.J. Life as manifestation of the second law of thermodynamics. Math. Comp. Model. 1994, 19, 25–48. [Google Scholar] [CrossRef]
- Mavelli, F.; Ruiz-Mirazo, K. Stochastic simulations of minimal self-reproducting cellular systems. Phil Trans. Royal Soc.: Biol. Sci. 2007, 362. [Google Scholar] [CrossRef]
- Rasmussen, S.; Chen, L.; Deamer, D.; Krakauer, D.C.; Packard, N.H.; Stadler, P.F.; Bedau, M.A. Transitions from Nonliving to Living Matter. Science 2004, 303, 963–965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, H.; Beard, D.A. Thermodynamic of stoichiometric biochemical networks in living system far from equilibrium. Biophys. Chem. 2005, 114, 213–220. [Google Scholar] [CrossRef]
- Schmiedl, T.; Seifert, U. Stochastic thermodynamics of chemical reaction networks. J. Chem. Phys. 2007, 126, 044101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michaelian, K. Thermodynamic dissipation theory for the origin of life. Earth Syst. Dynam. 2011, 2, 37–51. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Mirazo, K.; Briones, C.; de la Escosura, A. Chemial roots of biological evolution: The origins of life as a process of development of autonomous functional systems. Open Biol. 2017, 7, 170050. [Google Scholar] [CrossRef] [PubMed]
- Schuster, P.; Stadler, P.F. Networks in chemical evolution. Complexity 2001, 8, 34–42. [Google Scholar] [CrossRef]
- Polettini, M.; Esposito, M. Irreversible thermodynamics of open chemical networks. J. Chem. Phys. 2014, 141, 024117. [Google Scholar] [CrossRef]
- Eschenmoser, A.; Kisakürek, M.V. Chemistry and the Origin of Life. Helv. Chim. Acta 1996, 79, 1249–1259. [Google Scholar] [CrossRef]
- Barron, L.D. Symmetry and molecular chirality. Chem. Soc. Rev. 1986, 15, 189–223. [Google Scholar] [CrossRef]
- Avalos, M.; Babiano, R.; Cintas, P.; Jimenez, J.L.; Palacios, J.C.; Barron, L. Absolute asymmetric synthesis under physical fields: Facts and fictions. Chem. Rev. 1998, 98, 2391–2404. [Google Scholar] [CrossRef]
- Feringa, B.L.; van Delden, R.A. Absolute asymmetric synthesis: The origin, control, and amplification of chirality. Angew. Chem. Int. Ed. 1999, 38, 3418–3438. [Google Scholar] [CrossRef]
- Gu, H.; Nakamura, Y.; Sato, T.; Teramoto, A.; Green, M.M.; Jha, S.K.; Andreola, C.; Reidy, M.P.; Mark, H.F. Optical Rotation of Random Copolyisocyanates of Chiral and Achiral Monomers: Sergeant and Soldier Copolymers. Macromolecules 1998, 31, 6362–6368. [Google Scholar] [CrossRef]
- Guillaneux, D.; Zhao, S.H.; Samuel, O.; Rainford, D.; Kagan, E.B. Nonlinear Effects in Asymmetric Catalysis. J. Am. Chem. Soc. 1994, 116, 9430–9439. [Google Scholar] [CrossRef]
- Root-Bernstein, R. Simultaneous origin of homochirality, the genetic code and its directionality. BioEssays 2007, 29, 689–698. [Google Scholar] [CrossRef]
- Wu, M.; Walker, S.I.; Higgs, P.G. Autocatalytic replication and homochirality in biopolymers: Is homochirality a requierment of life or a result of it? Astrobiology 2012, 12, 818–829. [Google Scholar] [CrossRef] [PubMed]
- Schuster, P. Germany-Japan round table. Heidelberg. 2011. Available online: https://www.tbi.univie.ac.at/~pks/ (accessed on 30 May 2019).
- Cronin, J.R.; Pizzarello, S. Enantiomeric excesses in meteoric amino acids. Science 1997, 275, 951–955. [Google Scholar] [CrossRef]
- McGuire, A.B.; Carroll, P.B.; Loomis, R.A.; Finneran, I.A.; Jewell, P.R.; Remijan, A.J.; Blake, G.A. Discovery of the interstellar chiral molecule propylene oxide (CH3CHCH2O). Science 2016, 352, 1449–1452. [Google Scholar] [CrossRef]
- Oró, J.; Mills, T.; Lazcano, A. Comets and the formation of biochemical compounds on the primitive Earth–A review. Orig. Life Evol. Biosph. 1991, 21, 267–277. [Google Scholar] [CrossRef]
- Kempe, S.; Kazmierczak, J. Biogenesis and early life on Earth and Europa: Favored by an alkaline ocean? Astrobiology 2002, 2, 123–130. [Google Scholar] [CrossRef]
- Brack, A. Clay minerals and the origin of life. In Handbook of Clay Science, Developments in Clay Science; Bergaya, F., Theng, B.K.G., Lagaly, G., Eds.; Elsevier: Oxford, UK, 2006; Volume 1, pp. 379–391. [Google Scholar]
- Mislow, K. Absolute asymmetric synthesis: A commentary. Collect. Czech. Chem. Commun. 2003, 68, 849–864. [Google Scholar] [CrossRef]
- Mills, W.H. Some aspects of stereochemsitry. Chem. Ind. (Lond.) 1932, 51, 750–759. [Google Scholar] [CrossRef]
- Barabás, B.; Kurdi, R.; Pályi, G. Natural abundance isotopic chirality in the reagents of the soai reaction. Symmetry 2016, 8, 2. [Google Scholar] [CrossRef]
- Bolli, M.; Micura, R.; Eschenmoser, A. Pyranosyl-RNA: Chiroselective self-assembly of base sequences by ligative oligomerization of tetranucleotide-2′,3′-cyclophosphates (with a commentary concerning the origin of biomolecular homochirality). Chem. Biol. 1997, 4, 309–320. [Google Scholar] [CrossRef]
- Ben-Naim, B. Entropy Demystified: The Second Law of Thermodynamics Reduced to Plain Common Sense; World Scientific: Singapore, 2015. [Google Scholar]
- Plasson, R.; Kondepudi, D.K.; Bersini, H.; Commeyras, A.; Asakura, K. Emergence of homochirality in far-from-equilibrium systems: Mechanisms and role in prebiotic chemistry. Chirality 2007, 19, 589–600. [Google Scholar] [CrossRef] [PubMed]
- Ribo, J.M.; Blanco, C.; Crusats, J.; El-Hachemi, Z.; Hochberg, D.; Moyano, A. Absolute asymmetric synthesis in enantioselective autocatalytic reaction networks: Theoretical games, speculations on chemical evolution and perhaps a synthetic option. Chem. Eur. J. 2014, 20, 17250–17271. [Google Scholar] [CrossRef]
- Ross, J.; Vlad, M. Exact solutions for the entropy production rate of several irreversible processes. J. Phys. Chem. A 2005, 109, 10607–10612. [Google Scholar] [CrossRef]
- Hochberg, D.; Ribó, J.M. Stochiometric network analysis of entropy production in reaction networks. Phys. Chem. Chem. Phys. 2018, 20, 23726–23739. [Google Scholar] [CrossRef]
- Hochberg, D.; Ribó, J.M. Entropic analysis of mirror symmetry breaking in chiral hypercycles. Life 2019, 9, 28. [Google Scholar] [CrossRef]
- Werth, H. Über Irreversibilität, Naturprozesse und Zeitstrukture. In Offene Systeme I; Weizsäcker, E., Ed.; Klett-Cotta: Stuttgard, Germany, 1974; pp. 114–199. [Google Scholar]
- Kondepudi, D.; Kapcha, K. Entropy production in chiral symmetry breaking transitions. Chirality 2008, 20, 524–528. [Google Scholar] [CrossRef]
- Kondepudi, D.; Prigogine, I. Modern Thermodynamics, 2nd ed.; John Wiley & Sons: Chichester, UK, 2015. [Google Scholar]
- Plasson, R.; Bersini, H. Energetic analysis of mirror symmetry breaking processes in a recycled microreversible chemical system. J. Phys. Chem. B 2008, 113, 3477–3490. [Google Scholar] [CrossRef]
- Mauksch, M.; Tsogoeva, S.B. Spontaneous Emergence of Homochirality via Coherently Coupled Antagonistic and Reversible Reaction Cycles. ChemPhysChem 2008, 9, 2359–2371. [Google Scholar] [CrossRef] [PubMed]
- Plasson, R.; Brandenburg, A.; Jullient, L.; Bersini, H. Autocatalyses. J. Phys. Chem. A 2011, 115, 8073–8085. [Google Scholar] [CrossRef] [Green Version]
- Hochberg, D.; Bourdon García, R.D.; Ägreda Bastidas, J.A.; Ribó, J.M. Stoichiometric network analysis of spontaneous mirror symmetry breaking in chemical reactions. Phys. Chem. Chem. Phys. 2017, 19, 17618–17636. [Google Scholar] [CrossRef] [PubMed]
- Gellman, A.J.; Ernst, K.-H. Chiral autocatalysis and mirror symmetry breaking. Catal. Lett. 2018, 148, 1610. [Google Scholar] [CrossRef]
- Ribó, J.M.; Crusats, J.; El-Hachemi, Z.; Moyano, A.; Hochberg, D. Spontaneous mirror symmetry breaking in heterocatalytically coupled enantioselective replicators. Chem. Sci. 2017, 8, 763–769. [Google Scholar]
- Szathmary, E. The origin of replicators and reproducers. Philos. Trans. R. Soc. Lond. Ser. B 2006, 361, 1761–1776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frank, F.C. On spontaneous asymmetric synthesis. Biochim. Biophys. Acta 1953, 11, 459–463. [Google Scholar] [CrossRef]
- Ribó, J.M.; Hochberg, D. Competitive exclusion principle in ecology and absolute asymmetric synthesis in chemistry. Chirality 2015, 27, 722–727. [Google Scholar] [CrossRef]
- Rivera Islas, J.; Lavabre, D.; Grevy, J.-M.; Hernández Lamoneda, R.; Rojas Cabrera, K.; Micheau, J.C.; Buhse, T. Mirror-symmetry breaking in the Soai reaction: A kinetic understanding. Proc. Natl. Acad. Sci. USA 2005, 102, 13743–13748. [Google Scholar] [CrossRef] [Green Version]
- Lavabre, D.; Micheau, J.-C.; Islas, J.R.; Buhse, T. Kinetic Insight into specific features of the autocatalytic Soai reaction. In Amplification of Chirality; Springer: Berlin, Germany, 2008; Volume 284, pp. 67–96. [Google Scholar]
- Crusats, J.; Hochberg, D.; Moyano, A.; Ribó, J.M. Frank models and spontaneous emergence of chirality in closed systems. ChemPhysChem 2009, 10, 2123–2131. [Google Scholar] [CrossRef]
- Avetisov, V.; Goldanskii, V. Mirror symmetry breaking at the molecular level. Proc. Natl. Acad. Sci. USA 1996, 93, 11435–11442. [Google Scholar] [CrossRef] [PubMed]
- Ribó, J.M.; Hochberg, D. Stability of racemic and chiral steady states in open and closed chemical systems. Phys. Lett. A 2008, 373, 111–122. [Google Scholar] [CrossRef] [Green Version]
- Blanco, C.; Crusats, J.; El-Hachemi, Z.; Moyano, A.; Hochberg, D.; Ribó, J.M. Spontaneous Emergence of Chirality in the Limited Enantioselectivity Model: Autocatalytic Cycle Driven by an External Reagent. ChemPhysChem 2013, 14, 2432–2440. [Google Scholar] [CrossRef] [PubMed]
- Ribó, J.M.; Crusats, J.; El-Hachemi, Z.; Moyano, A.; Blanco, C.; Hochberg, D. Spontaneous Mirror Symmetry Breaking in the Limited Enantioselective Autocatalysis Model: Abyssal Hydrothermal Vents as Scenario for the Emergence of Chirality in Prebiotic Chemistry. Astrobiology 2013, 13, 132–142. [Google Scholar] [CrossRef] [Green Version]
- Blanco, C.; Crusats, J.; El-Hachemi, Z.; Moyano, A.; Veintemillas-Verdaguer, S.; Hochberg, D.; Ribó, J.M. The Viedma Deracemization of Racemic Conglomerate Mixtures as a Paradigm of Spontaneous Mirror Symmetry Breaking in Aggregation and Polymerization. ChemPhysChem 2013, 14, 3982–3993. [Google Scholar] [CrossRef]
- El-Hachemi, Z.; Crusats, Z.; Ribó, J.M.; Veintemillas-Verdaguer, S. Spontaneous transition toward chirality in the NaClO3 crystallization in boiling solutions. Cryst. Growth Des. 2009, 9, 4802–4806. [Google Scholar] [CrossRef]
- Cintas, P.; Viedma, C. Homochirality beyond grinding: Deracemizing chiral crystals by temperature gradient under boiling. Chem. Commun. 2011, 47, 12786–12788. [Google Scholar] [CrossRef]
- Crusats, J.; Veintemillas-Verdaguer, S.; Ribó, J.M. Homochirality as a Consequence of Thermodynamic Equilibrium? Chem. Eur. J. 2006, 12, 7776–7781. [Google Scholar] [CrossRef]
- El-Hachemi, Z.; Arteaga, O.; Canillas, A.; Crusats, J.; Sorrenti, A.; Veintemillas-Verdaguer, S. Achiral-to-chiral transition in benzil solidification: Analogies with racemic conglomerates systems showing deracemization. Chirality 2013, 25, 393–399. [Google Scholar] [CrossRef]
- Viedma, C. Fighting fire with fire: Racemization drives deracemization. In Proceedings of the Chiral Symmetry Breaking at Molecular Level, Solvay Workshop, Brussels, Belgium, 28–30 November 2018. [Google Scholar]
- Kondepudi, D.K.; Kauffman, R.; Singh, N. Chiral symmetry breaking in sodium chlorate crystallization. Science 1990, 250, 975–976. [Google Scholar] [CrossRef]
- Kondepudi, D.K.; Asakura, K.; Laudadio, J. Chiral symmetry breaking in stirred crystallization of 1,1′-binaphthyl melt. J. Am. Chem. Soc. 1999, 121, 1448–1451. [Google Scholar] [CrossRef]
- Ribó, J.M.; El-Hachemi, Z.; Arteaga, O.; Canillas, A.; Crusats, J. Hydrodynamic Effects in Soft-Matter Self-Assembly: The Case of J-Aggregates of Amphiphilic Porphyrins. Chem. Record. 2017, 17, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Lv, B.; Duan, P.; Liu, M.; Yin, M. Stoichiometry-controlled inversion of circulaly polarized luminiscence in co-assembly of chiral gelators with achiral tetraphenylethylene derivative. Chem. Commun. 2019, 55, 2194–2197. [Google Scholar] [CrossRef] [PubMed]
- Karunakaran, S.C.; Cafferty, B.J.; Weigert-Muñoz, A.; Schuster, G.B.; Hud, N.V. Spontaneous Symmetry Breaking in the Formation of Supramolecular Polymers: Implications for the Origin of Biological Homochirality. Angew. Chem. Int. Ed. 2019, 58, 1453–1457. [Google Scholar] [CrossRef] [PubMed]
- Blanco, C.; Stich, M.; Hochberg, D. Mechanically induced homochirality in nucleated enantioselective polymerization. J. Phys. Chem. B 2017, 121, 942–955. [Google Scholar] [CrossRef] [PubMed]
- Saghatelian, A.; Yokobayashi, Y.; Soltaini, K.; Gadhiri, M.R. A chiroselective peptide replicator. Nature 2001, 409, 797–801. [Google Scholar] [CrossRef] [PubMed]
- Eschenmoser, A. The search of the chemistry of life’s origin. Tetrahedron 2001, 63, 12821–12844. [Google Scholar] [CrossRef]
- Monnard, P.-A.; Walde, P. Current ideas about prebiological compartmentalization. Life 2015, 5, 1239–1262. [Google Scholar] [CrossRef]
- Kauffman, S. At Home in the Universe; Oxford University Press: Oxford, UK, 1995. [Google Scholar]
- Kondepudi, D.K.; Prigogine, I.; Nelson, G. Sensitivity of branch selection in nonequilibrium systems. Phys. Lett. A 1985, 111, 29–32. [Google Scholar] [CrossRef]
- Avetisov, V.A.; Kuz’min, V.V.; Anikin, S.A. Sensitivity of chemical chiral systems to weak asymmetric factors. Chem. Phys. 1987, 112, 179–187. [Google Scholar] [CrossRef]
- Pitsch, S.; Wendeborn, S.; Jaun, B.; Eschenmoser, A. Why Pentose- and Not Hexose-Nucleic Acids? Helvetica Chim. Acta 1993, 76, 2161–2183. [Google Scholar] [CrossRef]
- Shapiro, R. Prebiotic ribose synthesis: A critical analysis. Orig. Life Evol. Biosph. 1988, 18, 71–85. [Google Scholar] [CrossRef] [PubMed]
- Stern, R.; Jedrzejas, M.J. Carbohydrate polymers at the center of life’s origin. Chem. Rev. 2008, 108, 50161–55085. [Google Scholar] [CrossRef] [PubMed]
- Pestunova, O.P.; Simonov, A.N.; Suytnikov, V.N.; Parmon, V.N. Prebiotic carbohydrates and their derivatives. In Biosphere Origin and Evolution; Dobretsov, N., Kolchanov, N., Rozanov, A., Zavarzin, G., Eds.; Springer: New York, NY, USA, 2008; pp. 103–118. [Google Scholar]
- Orgel, L.E. Self-organized metabolic cycles. Proc. Natl. Acad. Sci. USA 2000, 97, 12503–12507. [Google Scholar] [CrossRef] [PubMed]
- Sel’kov, E.E. Self-oscillations in glycolysis. Eur. J. Biochem. 1968, 4, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Plasson, R.; Bersini, H.; Commeyras, A. Recycling Frank: Spontaneous emergence of homochirality in noncatalytic systems. Proc. Natl. Acad. Sci. USA 2004, 101, 16733–16738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stich, M.; Blanco, C.; Hochberg, C. Chiral and chemical oscillations in a simple dimerization model. Phys. Chem. Chem. Phys. 2013, 15, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Schmitt-Kopplin, P.; Gabelica, Z.; Gougeon, R.D.; Fekete, A.; Kanawati, B.; Harir, M.; Gebefuegi, I.; Eckel, G.; Hertkorn, N. High molecular diversity of extraterrestrial organic matter in Murchison meteorite revealed 40 years after its fall. Proc. Natl. Acad. Sci. USA 2010, 107, 2763–2768. [Google Scholar] [CrossRef] [Green Version]
- Krishnamurthy, R. Life’s Biological Chemistry: A Destiny or destination starting from prebiotic chemistry? Chem. Eur. J. 2018, 24, 16708–16715. [Google Scholar] [CrossRef]
- Peretó, J.; López-García, P.; Moreira, D. Ancestral lipid biosynthesis and early membrane evolution. Trends Biochem. Sci. 2004, 29, 469–477. [Google Scholar] [CrossRef]
- Davankov, V. The chirality as an inherent general property of matter. Chirality 2006, 18, 459–461. [Google Scholar] [CrossRef] [PubMed]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ribó, J.M.; Hochberg, D. Chemical Basis of Biological Homochirality during the Abiotic Evolution Stages on Earth. Symmetry 2019, 11, 814. https://doi.org/10.3390/sym11060814
Ribó JM, Hochberg D. Chemical Basis of Biological Homochirality during the Abiotic Evolution Stages on Earth. Symmetry. 2019; 11(6):814. https://doi.org/10.3390/sym11060814
Chicago/Turabian StyleRibó, Josep M., and David Hochberg. 2019. "Chemical Basis of Biological Homochirality during the Abiotic Evolution Stages on Earth" Symmetry 11, no. 6: 814. https://doi.org/10.3390/sym11060814
APA StyleRibó, J. M., & Hochberg, D. (2019). Chemical Basis of Biological Homochirality during the Abiotic Evolution Stages on Earth. Symmetry, 11(6), 814. https://doi.org/10.3390/sym11060814