1. Introduction
Linear chains, such as proteins and nucleic acids, demonstrate an immense structural diversity owing, in part, to a myriad of possible chain configurations which appear as various knots [
1], slip-knots [
2], and loops, and are believed to be of relevance to biological function of these molecules [
3,
4]. Advances in chemistry enabled synthesis of artificial molecular knots with various physicochemical properties [
5,
6,
7]. A three-dimensional structure of linear molecular chains is commonly described in terms of knot theory [
8], which is a powerful and rigorous mathematical concept. The approach is generic and applicable to any linear chain, not limited to biological molecules. In terms of knot theory, a knot is a one-dimensional topological circle embedded into three-dimensional space; it is a continuous structure without free ends. In other words, in order to turn a linear chain into a knot, one has to join the chain ends. While discussing chains and knots, it might be convenient to think of a rope, which we will tie and tangle. The most basic, “undecomposable” knots are called prime knots. Some of them are shown in
Figure 1. To refer to a knot, we use the Alexander-Briggs notation, which is common in knot theory, where the capital number specifies the number of crossings in the minimal crossing projection and the subscript is assigned in order to distinguish between knots with the same number of crossings. In
Figure 1, the number of crossings in each knot cannot be decreased, but can easily be increased by, for example, twisting some loops or threading the rope through a loop. Knots do not change upon deformations that do not break the rope, i.e., that do not break the continuity of the knot. Such deformations can be expressed via a sequence of specific deformations performed on a knot projection, which are called Reidemeister moves. The resulting structure could look very different from the original primary knot, but is perceived as equivalent by knot theory. This is one of the major ideological differences between knot theory and molecular engineering. Knot theory is designed for other purposes, namely to capture topological invariance under ambient isotopy (i.e., whether two knots can be deformed into each other); while in the case of molecular engineering, even minor changes in the shape of the chain might matter. For example, slip-knots—which are very common in proteins and crucially important for their proper functioning—are ignored by knot theory. However, the nature of slip-knots is geometric rather than topological, and therefore knot theory has to ignore them. In our work, we aim to develop a theory that would serve for molecular engineering. All basic molecular engineering operations should have a clear and intuitive representations. Hence, having the basic structural units to build up a chain seems to be convenient. For example, it is well known that by cutting the loop of a slip-knot, we get the slip-knot vanished and the chain reduced to a knot, e.g., to a trefoil. Our theory should (and will) give a clear
analytical visualization of this process and a prediction of what knot we will end up with. For more information on molecular knots and the related basics of knot theory, see the comprehensive review by Fielden et al. [
9].
In this paper, we will consider chains with different geometrical shapes. In order to make sure that two chains cannot be deformed into each other, we will join their ends to form a mathematical knot and then we calculate the so-called Alexander polynomial. Its definition can be found in any knot theory textbook [
10]. There is a rigorous mathematical algorithm for how to calculate it. In our previous paper [
11], we demonstrate it step-by-step for the objects most relevant in our approach and discussed in the next section. If two knots have different Alexander polynomials, then these knots must be different. The inverse statement is usually correct, but not always.
An application of knot theory to proteins is centered around a search for prime knots in a spatial protein structure. The only knots that have so far been found in proteins [
12] are
,
,
, and
. What is a fundamental reason for this choice? What property exactly separates these knots from other knots? The answer to the second question is known. The knots found in proteins can be formed following the so-called twisted hairpin folding mechanism [
14] outlined below. This mechanism is rather a phenomenological explanation that does not provide a fundamental difference between knots in terms of knot theory. In principle, a topological theory alone is not able to provide such a reason because physical properties of the chain must matter. In our theory, the twisted hairpin mechanism appears naturally as part of the formalism. A few years ago, the concept of circuit topology was suggested in order to account for intra-chain contacts [
15,
16,
17], which are also very important for proteins. Very recently, circuit topology was generalized to account for chain entanglement as well, focusing on applicability to real-life molecules [
11]. The new framework is still in its development stage and lacks certain rigorousness, especially in comparison to very-well developed knot theory. In this study, we attempt to strengthen the foundation of generalized circuit topology and demonstrate that this theory appeals to the “natural and inherent” language describing entanglement. To demonstrate this, we will, among others, re-discover some known results, which appear smoothly as an internal part of circuit topology. We believe it will be useful for molecular engineering and will help puzzle out the design principle of naturally evolved protein knots.
2. S-Contacts
We are looking for basic structural units which would comprise a molecular knot and be invariant to the knot structure, i.e., the “smallest piece of entanglement” that would not change upon a knot deformation. By a deformation, we understand any manipulation with a chain as long as the chain is not broken. One should keep in mind that any chain is an open structure, i.e., it has two ends. By passing ends through knots, we can tie or untie anything. Therefore, the ends must not participate in any manipulations, so that we do not change the topological structure of the chain. Additionally, we recognize the fact that molecules are three-dimensional, hence our formalism is built in 3D, so for the most part, we do not consider projections. On the other hand, it makes it harder to describe the mutual position of different segments of a chain. Unlike in 2D, in 3D there are no crossings of a chain and the corresponding segments of a chain might be distant in space, hence one cannot strictly define a loop. In what follows, these terms should be understood merely as references to certain segments or configurations of a chain in 3D, and as a tribute to the fact that all drawings are inevitably flat, i.e., 2D. A single loop, i.e., one twist of a chain (or a rope) that leads to only one crossing (or no crossings at all in certain projections), is not stable in the sense that it can easily be undone (untied) by stretching the chain, and hence it cannot serve as a structural unit. Indeed, a structural unit cannot disappear: once it is found, it must always be there. To make a loop stable, one should “fix” the loop by threading the chain through it.
Figure 2a shows all four possible resulting structures drawn in the way to visually single out the loop and to easily trace how the loop gets fixed. Keep in mind that we are looking for the basic structural units, i.e., we are fixing the loop in the simplest possible way. In principle, the chain can wind around the loop several times before threading the loop, or it can participate in another piece of entanglement. All these effects lead to a creation of several structural units connected in some specific way. This will be considered in the next section.
Each basic structural unit is called an s-contact, or a soft contact, and cannot by untied by stretching the chain. The term “contact” was coined to stress similarity to intra-chain, non-entanglement contacts, which can also be treated by circuit topology [
11], but are not considered in the present paper. Each contact must have two sites. (In case of intra-chain contacts, contact sites are the two chain segments that are linked together). An s-contact is supposed to be contained between its contact sites, i.e., contact sites are the boundaries of the entangled piece of the chain, as viewed while moving along the chain. We define that contact sites are located where the chain passes through the loops, so that the entangled segment is located in-between. What happens to the chain outside to the structural unit (i.e., on either side from the contact sites) is irrelevant. Since s-contacts, and real molecular chains, should be considered as three-dimensional entities, the exact position of contact sites depends on many parameters [
11], e.g., on the knot tightness, but it always represents the knot structure. Additionally, contact sites can migrate along the chain during a chain deformation. This uncertainty is essential in order to be able to catch the fixed topological structure of a flexible, mobile chain. If the chain were solid, incapable of any movements, or in case of a static chain projection, s-contact sites would not be movable. Contact sites are depicted with a red ball on several structures shown in
Figure 2.
Figure 2 shows several equivalent representations of s-contacts. Each column represents only one structure, i.e., all four structures in each column can be deformed one into another. It might not be immediately obvious, therefore we put colored strips to ease the tracking of corresponded deformations. Let us consider the first column in
Figure 2. Panel “a” is designed to highlight the loop being fixed by the chain end passing through it. If we move the chain segment at the top to the left to make it more symmetric, we will end up with panel “b”. If we move the red and blue strips from panel “a” up, we will get panel “c”. Therefore, even though the structures within one column are perceived by an eye as different, a 3D transition between them requires just a minor deformation. Such deformations are irrelevant and occur all the time in real molecules. In our theory, we do not want to notice these minor deformations because they do not make any physics contribution. We do not want to be concerned with them because they do not change the properties of the molecule. This is why we use such a flexible definition of s-contacts and s-contacts’ site, i.e., we make them sensitive to the molecule’s structure, which cannot be changed, but ignorant of all irrelevant details that in real molecules are nothing but noise. While threading the loop, we have to go around one segment of the chain, thereby creating another loop. Therefore, each of the four structures is symmetric: each consists of two identical loops hooked together. Following the chain in either direction, left-to-right or right-to-left, we see the same structure, which is most noticeable in panel “c”. The representation in panel “d” shows that each s-contacts consists of two loops. This representation is topologically identical to the other representations, but is more distinct from them. A transition to it requires a major deformation. This representation can for convenience be considered flat as a limiting case of 3D, which will be useful in the next sections. This “flat” representation resembles projections used by knot theory and can be useful in building a link between knot theory and circuit topology.
Contacts should be given names. We usually use capital letters, such as contact A, contact B, etc. In
Figure 2, each chain has one contact. If we move along the chain from any end and write down contact sites we encounter, we will get AA, which is a code for one contact. How to distinguish between the four different s-contact shown in the figure?
Figure 2d shows the same s-contact from
Figure 2a in a different representation where loops are easier to spot. The both representations are topologically equivalent and one can continuously transition from one to another without breaking the chain. Each s-contact is a connection of 2 loops, which are easy to recognize in
Figure 2d. Each connection has 4 crossings, which can naturally be split into 2 independent pairs: the crossings forming the loops (located on the sides) and the crossings forming the connection between the loops (located in the middle). The crossings in each pair are not independent. If one of them is flipped, the s-contact unties, as should be clear from the illustration. Each independent crossing can take two values, depending on which chain segment is on top. Two values of two independent crossings give rise to 4 different structures. It means that there are only 4 s-contacts possible, and hence our set of s-contacts is complete. The pair of crossings defining the s-contact chirality (i.e., forming the loops) is depicted on the left-most s-contact. The chirality of each loop is defined by the conventional right-hand rule. If the loops have different chirality, then none of these loops have been fixed by threading the chain through them, which means this structure will untie. In other words, despite consisting of two loops, s-contacts can only be either positive, A
A, or negative, A
A.
The other independent pair of crossings, shown on the right-most s-contact in
Figure 2d, defines how the two loops are hooked together. If the chain passes through the loop in the same direction as the chain shifts in each loop at the crossings defining chirality, such an s-contact is called “even”, A
eA. If the chain passes through the loop in the opposite direction, the s-contact is called “odd”, A
oA. Hence, the only 4 possible s-contacts are A
A, A
A, A
A, and A
A. This notation is called the string notation of circuit topology. It codes a chain entanglement as a string of letters. One of the advantages of this notation is the ability to apply combinatorial analysis directly to a description of entanglement. Additionally, note that the attributes introduced above are universal and do not depend on the chain orientation, i.e., in which direction we move along the chain, left-to-right or right-to-left.
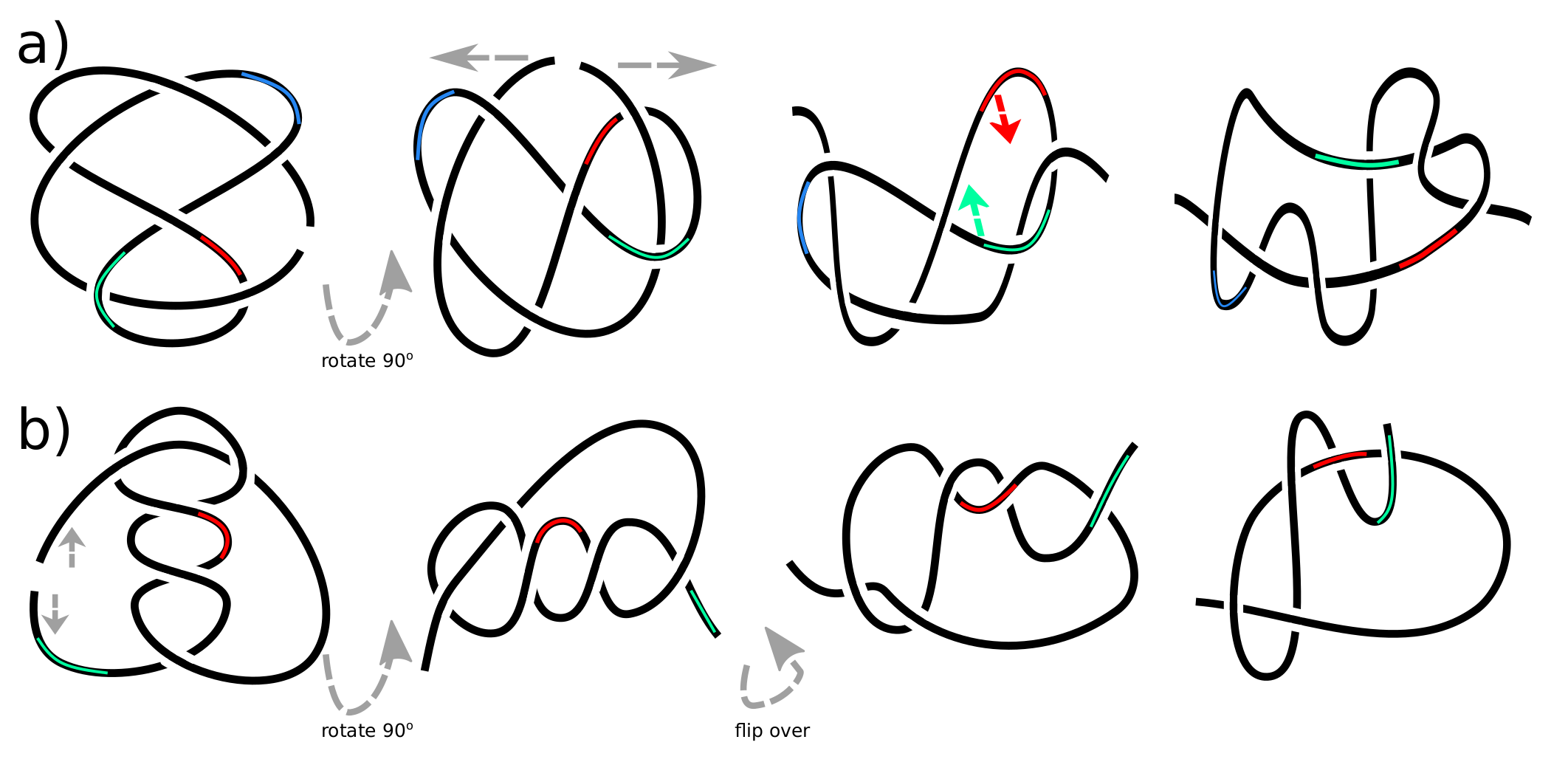
Let us consider s-contacts in the view of knot theory. To form a knot from a rope, one has to join its ends. To make a rope form a knot, one has to cut the knot somewhere. A
eA corresponds to
. This knot can be right-handed, as in
Figure 1, or left-handed if all the crossings are flipped. A
oA corresponds to
, see
Figure 3a for a visualization of the sequence of corresponding deformations. This knot is known to be achiral (amphichiral), i.e.,
, i.e., A
A and A
A can be deformed into each other. The sequence of corresponding moves is shown in
Figure 3b. However, both A
A and A
A should be kept and considered as separate s-contacts because, as will be shown below, they comprise different knots in a presence of other s-contacts. Can we distinguish A
A and A
A when they are along? Topologically speaking, we cannot, and knot theory is clear about this. Geometrically speaking, it boils down to the notion of stability, similar to retaining or quenching slip-knots. Namely, the deformation needed for a transition between A
A and A
A costs energy. If the cost is low, the transition can occur spontaneously. Otherwise, it will not occur, rendering the molecular knot stable. We will discuss it in
Section 4.2. However, note in
Figure 2b that A
A and A
A look like a mirror reflection. Such a flip of symmetry matters in proteins, so we must retain it for molecular engineering purposes.
So far, we identified 4 stable “basic structural units” of chain entanglement, and called them s-contacts. Note that if we flip only one crossing in any chain from
Figure 2, the s-contact will disappear and that chain will untie. Any messy blob of a rope is held together by loops hooking to each other, which is the essence of entanglement and the definition of s-contacts. In the next section, we will consider how s-contacts can be connected to each other. S-contacts might not be easy to spot in illustrations, even in such a simple case of
knot. Where exactly are s-contacts in the prime knots shown in
Figure 1? We will break it down below, but at the current state of our theory, it is easier to go in the opposite direction, i.e., to tie s-contacts on a rope and then identify the resulting knot. In the present paper, we tie s-contacts, we tie knots and we want to see how it works. Here, we focus on developing the formalism of s-contacts. When it comes to analyzing real folded molecules, the procedure is the opposite: we have to identify s-contacts based on the input from experimental data, for example based on the positions of all atoms in the molecule. This is a separate problem, which should be treated numerically and will be consider in another paper. So far, a protocol, along with a computer code, has been developed to treat intra-chain contacts [
18].
3. SPX Configurations of S-Contacts
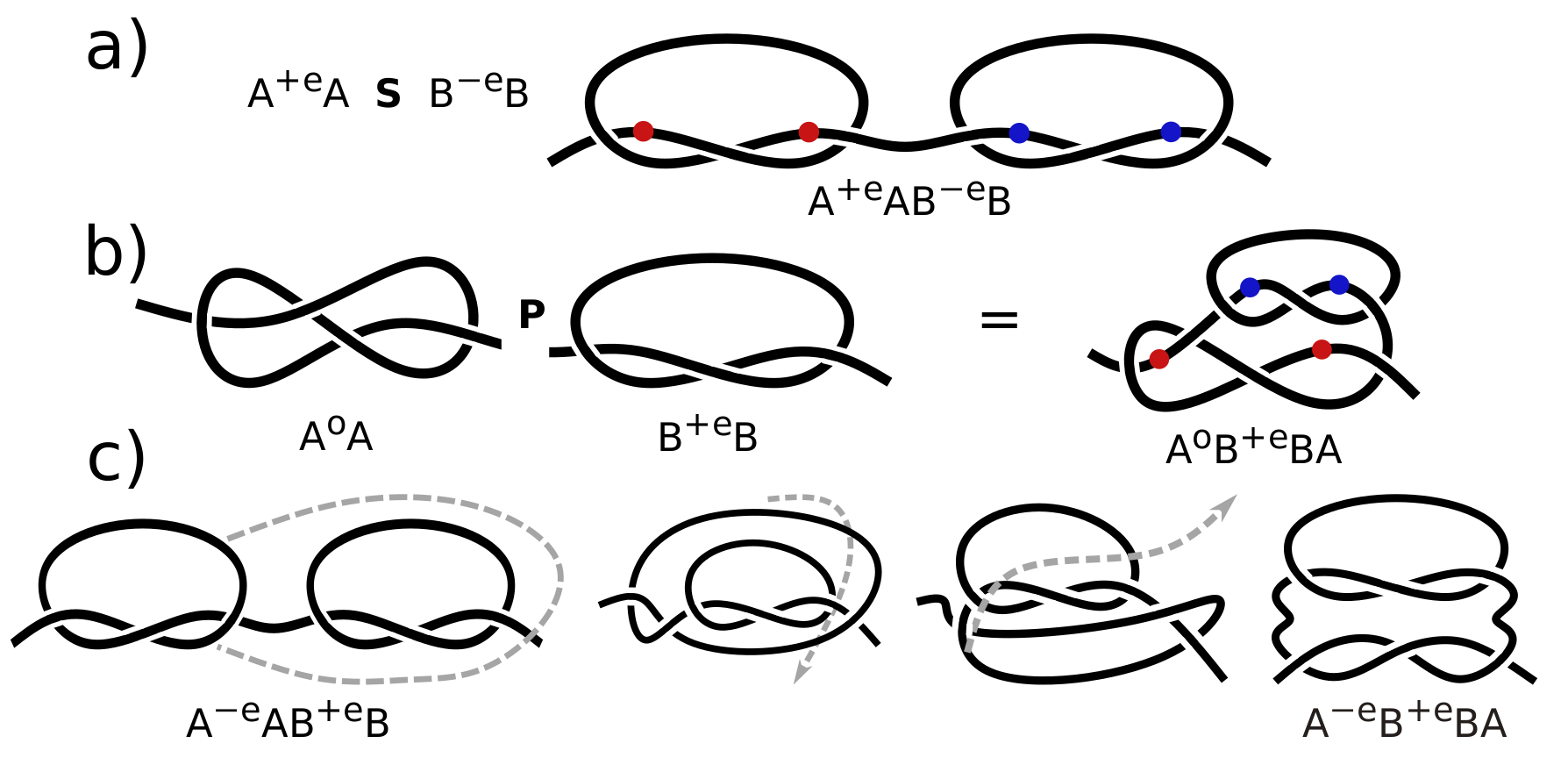
One s-contact has two contact sites and appears in the string notation as AA where each letter signifies this contact’s sites. Two s-contacts, AA and BB, can occur in three different configurations defined by permutations of two pairs of letters: AABB, ABBA, and ABAB. Because s-contacts can have any name, the configurations ABAB and BABA are identical. These three configurations are called series (S), parallel (P), and cross (X), and comprise the SPX relations. Regardless how many s-contacts a knot consists of, these pair-wise relations always hold and are shown unambiguously by the string notation. For example, in AACDCBDB, all relations are immediately obvious, e.g., the contacts A and B are in series, and contacts C and D are in cross, etc. Therefore, a consideration of all pairs of s-contacts is sufficient to describe a chain entanglement. Indeed, the string notation lists all contact sites as they appear along the chain and can be unambiguously deduced from the relative positions of all pairs of contact; thereby it completely specifies (or codes) the chain entanglement in terms of s-contacts.
Series and parallel configurations are easy to visualize.
Figure 4a shows two s-contacts in series;
Figure 4b shows two other s-contacts in parallel; and contact sites are marked with colored balls. Note that in
Figure 4b the internal contact, marked with blue balls, can be placed in different loops of the external contact marked with red balls. However, blue balls will always be between red balls, i.e., the contacts are always in parallel with the same string notation (ABBA). Here, we do not specify the symmetry and chirality because this rationale works for any kind of s-contact. SP configurations with other types of s-contacts can be drawn in a similar manner. Note that chains from
Figure 4a,b look very different not only because they consist of different s-contacts, but also because of the different relations between s-contacts, i.e., series and parallel. However, the relations of s-contacts can be swapped between each other by a sequence of moves shown in
Figure 4c. Notice that contact B is not altered in any way during the deformation. Hence, it can be replaced by any other kind of s-contact or any arrangement of s-contacts. Contact A throws out a loop and gets deformed. A similar deformation can be applied to any kind of s-contact. Applying this deformation to one s-contact after another, one can push s-contacts inside other s-contacts or pull them outside, thereby dragging an s-contact along the string in string notation. For example, one can turn A
eA B
B C
eC into A
eA C
eC B
B (i.e.,
to
). In general, because a pair-wise consideration is sufficient, any set of s-contacts consisted of only SP configurations can have any fraction of S and P relations as long as their total number is constant. As limiting cases, such a set can be deformed into all s-contacts in series or all s-contacts in parallel. Because in this case s-contact can be singled out, A
A and A
A are indistinguishable in case of series and parallel configurations (since they correspond to
knot which is achiral). However, A
A and A
A cannot be deformed into other s-contacts (since
knot is not achiral). Two entangled chains consisted only of SP configurations can be deformed into each other only if they contain the same number of s-contacts of each kind. For example, the chains in SP configurations from
Figure 4a,b contain different kinds of s-contacts, and therefore cannot be deformed into each other.
The transition from
Figure 4c is important in the context of protein folding and has been studied in the literature [
19]. Knot theory cannot distinguish these two configurations because they correspond to the same knot [
11]. However, in real molecules, such a transition between these configurations requires energy and once again comes down to the question of stability; the transition might be very probable or might never happen, depending on the physical properties of the chain. Circuit topology aims at addressing this question and provides consideration of different levels of structural stability.
SP configurations are similar to the notion of a connected sum used in knot theory. To form a connected sum of two knots, one should cut each knot and merge the resulting ends together, which is the procedure demonstrated in
Figure 4. Consequently, based on the connected sum properties, the Alexander polynomial of several s-contacts in series or in parallel is a product of Alexander polynomials of each s-contact.
Cross (X) configurations are very different from SP configurations. Each s-contact consists of two loops.
Figure 5 shows all possible arrangements of two pairs of loops. Indeed, the loops in this “flat” representation can be counted similarly to counting s-contact sites: the loops can be in series, in parallel, and in cross. The S operation is obtained by joining the red and blue s-contacts together at one endpoint of each. The connection between the two contacts is shown in black. The P operation is obtained by cutting open the bottom arc of the red s-contact and joining the endpoints of the blue s-contact to the endpoints of the cut open part of the red s-contact. The X operation requires altering the s-contact sites (and hence, the loops) as in ABAB. It is obtained by first cutting open the bottom arc of both the red and the blue s-contacts to obtain red arcs
and
and blue arcs
and
, and turning the pictures so that
and
are above
and
, respectively. Next, one endpoint of
is attached to one endpoint of
and the other endpoint of
is attached to one endpoint of
, and one endpoint of
is attached to an endpoint of
. This gives us a single arc, alternating between red and blue, which has one endpoint on
and the other endpoint on
. The case when one loop is shared by a pair, i.e., C configuration, is considered in the next section. This “flat” representation is convenient for listing and counting cases because it retains the number of loops, but one should keep in mind that these flat structures can always be deformed into 3D. In S and P configurations, the loops belonging to the same s-contacts are hooked together. Each pair of loops can be separated and cut off the whole chain. It is not possible to do in X configuration where loops belonging to different s-contacts are connected. Contacts A
A and A
A are identical, i.e., can be deformed into each other, only as long as their loops are free to move as is the case in SP configurations, see
Figure 3. In X configuration, loops from one s-contact are connected to loops from another s-contact, and hence are not free. Therefore, contacts A
A and A
A are not identical and lead to different knots when they are parts of X configuration.
Let us consider the transition between S and P configurations by looking at the illustrations in
Figure 5 and see again that it does not work for X configuration (C configuration will be considered in the next section). Additionally, the single deformation shown in
Figure 4c is obvious only in case of two s-contacts. What if there are more s-contacts? What is the general rule? Topologically, we can stretch the chain but we should not break it. When two loops are joined into an s-contact, they are connected and cannot be separated. In contrast to loops, one single s-contact can be moved along the chain freely. So, we take the blue contact from the right top corner of
Figure 5 and move it to the left. It passes through the left loop of the red contact and then moves down to the bottom of the red contact. By this manipulation, we turned S configuration into P. In other words, we are only allowed to move the whole contact along the chain, but not contact sites. The transition AABB ⟶ ABBA should be understood as the whole contact B moves to the left, but not as one contact site of contact A moves to the right. Following the same logic, let us add contact C to the left of X configuration in
Figure 5. The resulting string is CC ABAB. Contact C as one entity can move through the loop of contact A to form A CC BAB or further to ABCCAB; but it can never lead to CACBAB or ACBCAB. As a consequence, X configuration cannot be turned into SP. Here, the crossings in
Figure 5 make no difference in the rationale, hence this conclusion is applicable to any kind of s-contacts.
There are 4 s-contacts (A
A, A
A, A
A, A
A), which gives rise to
configurations of 2 s-contact in cross, which can be written as a 4 × 4 table. This table is symmetric owing to the left-right symmetry, i.e., which direction we move along the rope, e.g., A
B
AB vs. B
A
BA. This leaves 4 configurations on the diagonal and 6 configurations above (or below) the diagonal, i.e., 4 + 6 = 10 configurations. As shown in
Figure 2, a change of chirality in a single s-contact, i.e., a change of all signs in the string notation, leads to a flipping of all crossings. This property holds for any combination of s-contacts. Indeed, flipping
all crossings means a mirror reflection of the whole knot. Due to this symmetry, the number of configurations to consider can be further reduced. On the diagonal, A
B
AB and A
B
AB can be treated (or drawn) as one configuration. Similarly, A
B
AB and A
B
AB. The same holds for two non-diagonal configurations: A
B
AB vs. A
B
AB and A
B
AB vs. A
B
AB. It leaves us with 10 − 4 = 6 configurations which are shown in
Figure 6. Let us count these 6 configurations again, but this time geometrically. First, we tie contact A whose contact sites are marked with red balls. We made one of the loops corresponding to contact A the largest in the illustration in order to make it easier to spot contact A. This loop can always be shrunk without changing the overall topology. After contact A is “fixed”, i.e., after the second red ball, where can the rope go? It can go away, which would create SP configurations. Or the rope can pass through this large loop again, thereby creating another s-contact in cross with contact A.
Figure 6 shows all possible route of the rope leading to another s-contact. If the rope keeps passing though the large loop, it will just create more and more s-contacts. If the rope keeps winding around the large loop, it will lead to the relation considered in the next section. Note that A
B
AB and A
B
AB look very different and their Alexander polynomials are different, though the only difference between them is the chirality of contact B. It is a rigorous proof that even though B
B and B
B are identical while standing along, i.e., while being in series or in parallel to other s-contacts, in cross configurations they lead to different composite structures. Hence, all 4 s-contacts should be retained.
In order to easily verify and visualize the correctness of strings provided in
Figure 6, one can either calculate the Alexander polynomial or untie one of the s-contacts. For example, in A
B
AB one can unhook the left site of contact A (red balls), so that contact A disappears and only B
B is left. In the drawing, this procedure means that the left-most crossing is flipped, so that the left red ball no longer passes through the loop. Then, the rope can be deformed into the configuration from
Figure 2a by reducing the loop freed by the flipping. The same holds for the right site of contact B (blue balls). Additionally, consider A
B
AB. After the second red ball, the rope wraps around the large loop and gets “fixed” by passing through the large loop at the location of the second blue ball. If the rope does not pass through the large loop, so there is no blue ball, then no matter how many times the rope wraps around the large loop, the second contact will not be formed. Indeed, this “spiral” around the large loop will not be stable and will be easily untied by pulling the rope ends apart.
Each of the four s-contacts has an Alexander polynomial degree 2. Two s-contacts of any kind in any of SPX configurations have Alexander polynomial degree 4 (these polynomials are provided in the corresponding figures).
n s-contacts of any kind in any SP configuration have Alexander polynomial degree
. Indeed, SP configurations correspond to a connected sum in knot theory, which proves that the Alexander polynomial of the sum equals a product of the Alexander polynomials of single connected knots [
10], in our case s-contacts. It is reasonable to expect that the Alexander polynomial degree scales the same for X configurations as well. Indeed,
Figure 6 shows a clear pattern of Alexander polynomials, depending on the kind of s-contacts in cross. The easiest pattern appears for positive even s-contacts. The Alexander polynomial of A
A is
; A
B
AB corresponds to
, see
Figure 6. One can predict that A
B
C
ABC has the Alexander polynomial
. The corresponding prime knot,
, is not drawn here, but can be easily deduced from the pattern, and it indeed has this Alexander polynomial. Let us formulate a recipe for how to construct
knot. A
A is shown in
Figure 2a. Then, the right end of the chain makes one circle around the horizontal segment and forms A
B
AB from
Figure 6. Another similar circle around the horizontal segment will lead to A
B
C
ABC. It would be interesting to investigate further on the relationship between Alexander polynomials and s-contacts, but it is beyond the scope of the present paper. Here, we only hypothesize that such a relation exists. One should, however, note that the chains in
Figure 6 have a different number of crossings, correspond to prime knots, which also have another number of crossings; yet they all have Alexander polynomials of the same degree. We attribute it to the pattern we outlined.
The cross configuration of chains in different forms in
Figure 6 look nothing alike, yet they are topologically identical. One can verify it by calculating Alexander polynomials provided in
Figure 6. To change to another chain in the “flat” representation of
Figure 5c, one has to flip just a few crossings (consult
Figure 2d). Therefore, all the different-looking chains from
Figure 6 look extremely similar when presented as loops. What about a 3D shape of real molecules? It depends on the physical and chemical properties of the molecule. If these properties require a minimization of the bending energy of the chain, the 3D shape would resemble a distorted version of the “non-flat” chains in
Figure 6. The “flat” representation is just convenient for theoretical studies, visualization of the formalism and tying simple knots. Despite having the same Alexander polynomials, the configurations from
Figure 6 do not visually resemble the prime knots from
Figure 1. One can deform them one into another. However, such manipulations require many-step, major deformations that are hard to follow and, quite frankly, tedious to draw. More importantly, this would have no practical use. Indeed, we aim at describing proteins and other linear macromolecules. In principle, the 4 s-contacts (
,
,
,
) are supposed to be found and identified automatically by a computer, not by a naked eye. Yet, it does look suspicious that s-contacts are not obvious in prime knots.
Figure 7a shows the equivalence of
knot from
Figure 1 and A
B
AB form
Figure 6. Surprisingly, it requires only a minor deformation.
and
can be treated similarly.
and
will be discussed in the next section.
5. Circuits
We found in
Section 3 that contacts as a whole can be dragged along the string, which explains the transition between S and P. What would happen if other relations were present? Let us consider ACABCB. Contacts A and B are in series, but they can never become in parallel because they cannot be dragged along the string. The dragging is blocked by contact C, which is in cross with contacts A and B. Indeed, if we want to drag contact A, we would also have to drag everything locked between the letters “A”, i.e., the letter “C”. However, it is not the whole contact, hence such a drag is forbidden. In our previous work [
11], we introduced the notion of circuits. A circuit is a segment of a string, which consists only of pairs of letters and subscripts of the same letters. In other words, a circuit can be isolated from other contacts. By “isolated”, we mean “can be put in series”. Circuits can be dragged along the string. Obviously, circuits can consist of several circuits, e.g., AABCBC consists of AA and BCBC. The number of possible prime knots for a given number of crossings is still unknown [
20,
21] and our theory might help find it. It would be interesting to further investigate this algebra of s-contacts and the detailed construction of prime knots out of circuits. For example, we said that SP looks similar to a connected sum. Why? It is because the circuit AABB consists of smaller circuits, namely AA and BB, and hence AABB is not a prime knot, which implies that it must be a connected sum of prime knots. In principle, coding entanglement as a string of letters offers an advantage of being able to apply combinatorial analysis (even before considering the algebra of circuit topology operations). In this paper, we employed it in a very mild proportion in order to count the number and kind of possible s-contacts (A
A, A
A, A
A, A
A), see
Figure 2, and all possible configurations of pairs of s-contacts,
Figure 5. Indeed, two s-contacts cannot have more then 2 + 2 = 4 loops, and we considered all configurations consisted of 2, 3, and 4 loops. This pair-wise consideration is sufficient to code entanglement, i.e., to specify the unique string corresponding to the chain, but, as we just saw by ACABCB, the dynamics of the chain, i.e., the mobility of s-contacts, can be affected by other contacts, so that larger scale structures such as circuits have to be considered.
In this paper, we demonstrated how circuit topology can be used to describe simple knots consisted of just a few s-contacts. However, how many are “just a few” in practice? This chain we considered, ACABCB, if s-contacts are assigned symmetry and chirality, leads to configurations. Half of these configurations are chirality symmetric (i.e., all crossings flipped; left-hand/right-hand symmetry). Additionally, configurations are left-right symmetric (i.e., the kinds of s-contacts A and B coincide). Half of these configurations are chirality symmetric as well and we do not want to count them twice. Therefore, independent configuration of the string ACABCB are possible. How many prime knots can we get from here? To turn a chain into a knot, one has to join its ends. In reverse, depending on where we cut the prime knot, we will end up with different-looking chains, which can be deformed into each other. In our example, ACABCB, the left letter “A” and the right letter “B” should be connected to form a knot. Then, we cut the resulting ring of letters at different spots, which gives rise to several strings (chains): ACABCB, BACABC, and CBACAB. The strings are the same up to a cyclic permutation. These chains look different, but they correspond to the same prime knot and can be deformed one into another. Because there are only three such permutations, there are different prime knots described by the chain ACABCB. Analyzing these cyclic permutations by combinatorics, we can easily see which chains can be deformed one into another. There are other strings, apart from ACABCB, made of three s-contacts. The number of different configurations grows fast with the number of s-contacts. Hence, all practically manageable chains involving reasonably complex prime knots are made of three or maximum four s-contacts (and subscripts). We believe it can be useful for engineering new molecular chains, which can be compiled from a small set of these basic structural units.
All the illustrations shown so far came from the pursuit to consider all possible configurations consisting of one and two s-contacts. It is a bottom-up approach when we combine the “basic units” and see which knots we end up with. Let us now go in the opposite direction. We will consider a fairly complicated knot and break it down to s-contacts. As mentioned above, it is a tedious procedure, which should be done by a computer, not by a naked eye. On the other hand, it helps to visualize and appreciate how s-contacts work in real life. We chose to consider knot
because it has the same Alexander polynomial as knot
from
Figure 1. In this paper, we use Alexander polynomials only to distinguish between knots while developing our approach. Alexander polynomials work very well, but fail in some rare cases. Let us see if our circuit topology can catch the different between
and
.
Figure 10a shows a sequence of moves to deform
to a more eye-friendly representation with one large loop. All the moves are in 3D. The string notation is (A
C
)B
ABC. Note that it is a circuit.
Figure 10b color-codes the s-contacts. Every rope segment trapped by a loop restricting its motion, gives rise to an s-contact site or to a subscript. Notice the use of the simplified notation for C configuration and the treatment of the subscripts originated from the loop passing through contact B (marked in dash). So, circuit topology clearly differentiates between
and
. Note that
contains the same s-contact, A
A, which comprises
. In addition, note that
and
contain a different number of s-contacts, hence the scaling of Alexander polynomial with the number of s-contacts does not hold in this case. The main reason for this is the presence of subscripts that are not a part of knot theory (see
Figure 9 where the pattern is broken as well). Whether chains with mixed operations (SP and X and C) follow the same scaling is unclear.
Table 1 shows Alexander polynomials and string notation for prime knots with up to seven crossings and two other knots. It is difficult to notice any patterns in the Alexander polynomials, while the strings look very consistent: first single s-contacts, then two s-contacts, then three s-contacts with all the combinations of concerted s-contacts. S-contacts with different symmetry and chirality offer knots with more crossings. Moreover, s-contacts can somewhat explain the Alexander polynomials. The polynomial power is double the number of contacts, where concerted contacts are counted as one. For example, knots
and
. A tricky case is knot
. Contacts A and B are concerted, so we count them simultaneously as 1. Contacts B and C are concerted, therefore we count them simultaneously as well, so contact C does not change the count. Hence, all three contacts are counted as 1. This kind of counting the number of contacts does not always work. It fails for knot
considered above. However, Alexander polynomials also have troubles with this knot. Another pattern to notice is that the leading coefficient of Alexander polynomials coincides with the number of letters in the parenthesis in the string notation. These patterns fail sometimes, but they also work very often and it would be interesting to study them further.
So far, we have considered only chains consisted of a small number of s-contacts. It might be sufficient when it comes to molecular engineering since all the knots so far found in proteins consist only of 1 or 2 s-contacts,
Figure 1. While listing these knots, we did not specify their chirality because it does not lead to any topological distinction, but only flips all the crossings in the knot. However, sometimes in the literature, their chirality is reported [
12], namely the knots in proteins are A
A, A
A, A
oA, A
A, and A
A, which are
,
,
,
, and
. As said above, A
oA or
is achiral, hence one cannot specify its sign as long as it is not in cross with other s-contacts. So, this list contains all single s-contacts (A
A, A
A, A
oA) and two s-contacts concerted (A
A, A
A). Why does this list not contain A
A, A
A? It has been agreed upon [
14] that topology cannot answer this question because it is related to the chemical structure of a protein chain. Additionally, it might be the case that these two configurations do exist, and just have not been found yet. All five of these found knots consist of concerted s-contacts only (single s-contacts are considered as a limiting case of concerted). The physical reason behind this is still unknown and lies beyond pure topology and the scope of this paper, though some speculations can be made. In order to tie a concerted structure, one has to thread a chain through a loop only once; whereas other configurations (knots) require two events of threading, thereby making them more complicated to tie. Another reason might be related to the 3D shape of the chain. To tie a concerted structure, one has to twist the chain a few times in order to form the spiral-like shape, see
Figure 8. Such a shape might be natural for proteins and induce less stress on the chain than other shapes. In other words, the twisting motion can be done automatically by the chain itself in order to attain the preferable spiral-like shape. Circuit topology might be a convenient approach to work with such problems because it can be naturally generalized to account for relevant physical properties. Indeed, circuit topology differentiates between stable configurations (s-contacts), meta-stable configurations (subscripts, i.e., slip-knots), and not-stable configurations (single loops). Each kind of s-contact possesses its own energy; and a transition between s-contacts requires some energy (maybe in a form of entropy penalty). By building up knots out of s-contacts, one can analytically estimate the energetical complexity of various transitions.















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}