
Dynamical Simulation of Effective Stem Cell Transplantation for Modulation of Microglia Responses in Stroke Treatment


Abstract
:1. Introduction
2. Mathematical Representation of the SDRPA Model
3. The SDRPA Model’s Equilibrium Points
4. The Equilibrium Points’ Stability of the SDRPA Model
- Theorem 1 indicates that the damage, D, can penetrate the SDRPA model, if .
- Theorem 2 indicates that the damage, , penetrated the brain.
- Theorem 3 indicates that stem cell transplantation, , modulates the inflammatory environment in a stroke, .
- The SDRPA model is considered stable when the immunomodulation from transplanted stem cells can be one of the mechanisms of post-stroke recovery.
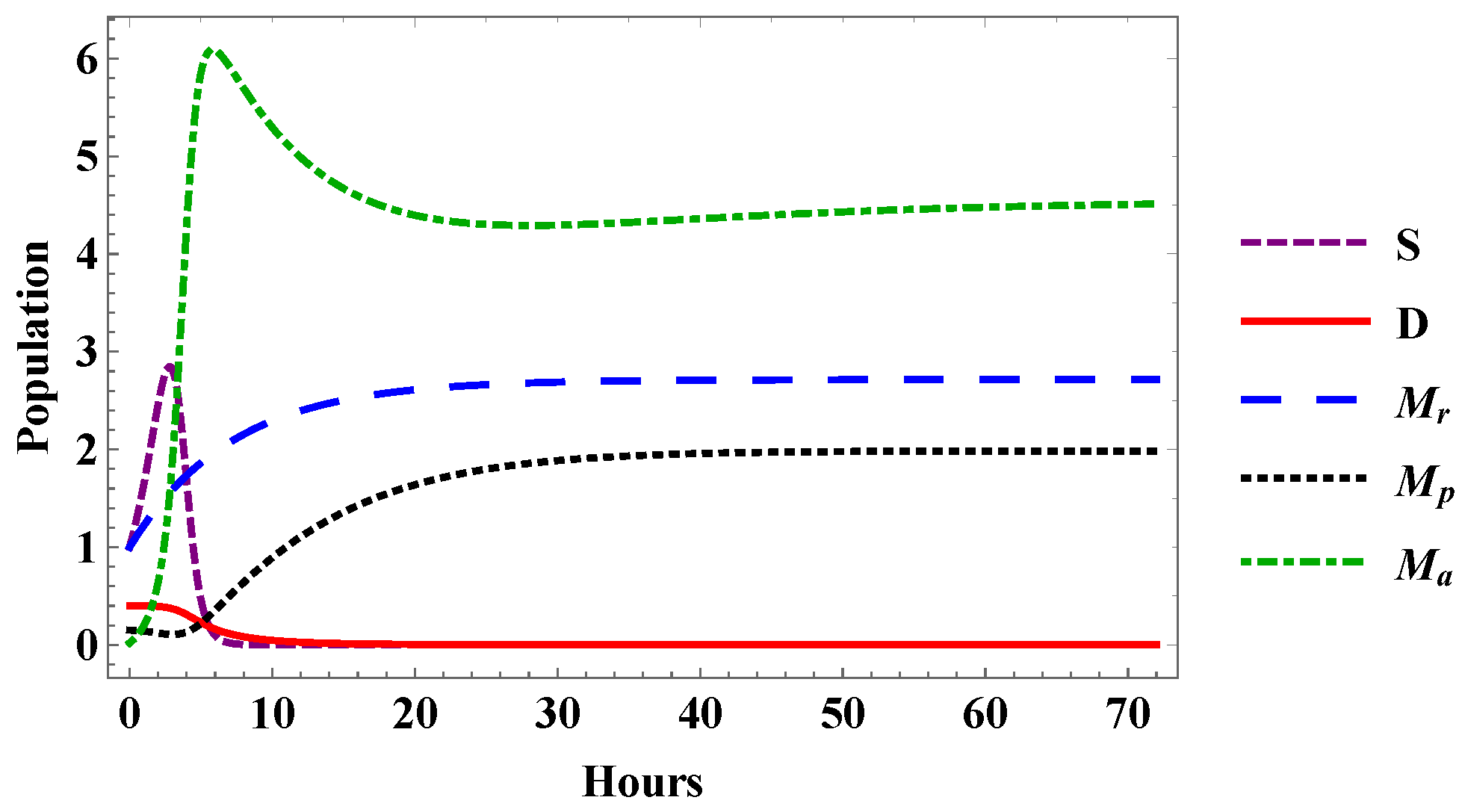
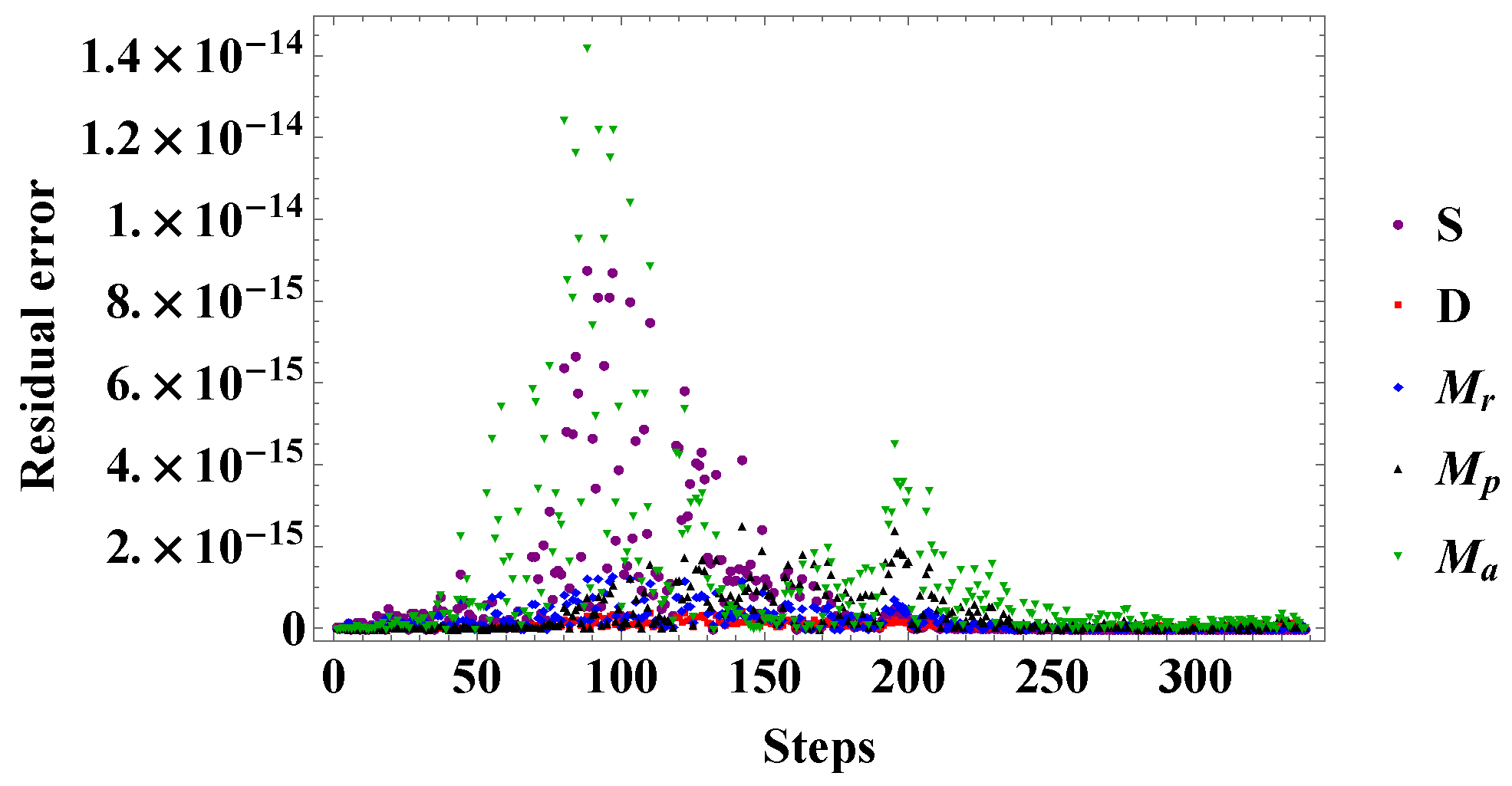
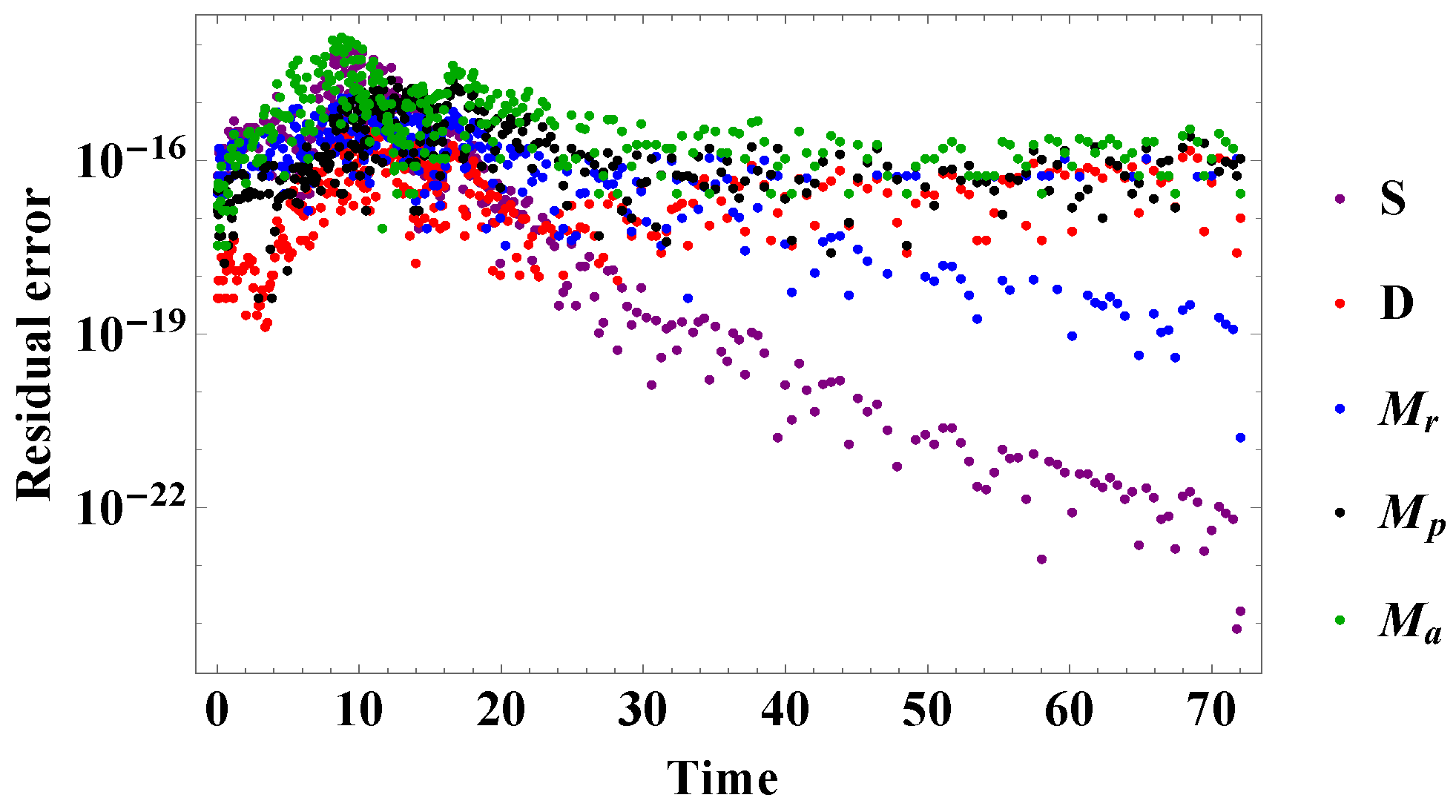
5. Numerical Results and Analysis
5.1. Determination of Parameters
5.2. Comparison of Experimental Results
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Marei, H.E.; Hasan, A.; Rizzi, R.; Althani, A.; Afifi, N.; Cenciarelli, C.; Shuaib, A. Potential of Stem Cell-Based Therapy for Ischemic Stroke. Front. Neurol. 2018, 9, 34. [Google Scholar] [CrossRef]
- Nordin, N.A.M.; Aljunid, S.M.; Aziz, N.A.; Nur, A.M.; Sulong, S. Direct medical cost of stroke: Findings from a tertiary hospital in malaysia. Med. J. Malays. 2012, 67, 473–477. [Google Scholar]
- Chouw, A.; Triana, R.; Dewi, N.M.; Darmayanti, S.; Rahman, M.N.; Susanto, A.; Putera, B.W.; Sartika, C.R. Ischemic Stroke: New Neuron Recovery Approach with Mesenchymal and Neural Stem Cells. Mol. Cell. Biomed. Sci. 2018, 2, 48–54. [Google Scholar] [CrossRef]
- Alqarni, A.J.; Rambely, A.S.; Hashim, I. Dynamic Modelling of Interactions between Microglia and Endogenous Neural Stem Cells in the Brain during a Stroke. Mathematics 2020, 8, 132. [Google Scholar] [CrossRef] [Green Version]
- Russo, C.D.; Lagaert, J.-B.; Chapuisat, G.; Dronne, M.-A. A mathematical model of inflammation during ischemic stroke. ESAIM Proc. 2010, 30, 15–33. [Google Scholar] [CrossRef]
- Hui-Yin, Y.; Ahmad, N.; Azmi, N.; Makmor-Bakry, M. Curcumin: The molecular mechanisms of action in inflammation and cell death during kainate-induced epileptogenesis. Indian J. Pharm. Educ. Res. 2018, 52, 32–41. [Google Scholar] [CrossRef] [Green Version]
- Vay, S.U.; Flitsch, L.J.; Rabenstein, M.; Rogall, R.; Blaschke, S.; Kleinhaus, J.; Reinert, N.; Bach, A.; Fink, G.R.; Schroeter, M.; et al. The plasticity of primary microglia and their multifaceted effects on endogenous neural stem cells in vitro and in vivo. J. Neuroinflamm. 2018, 15, 226. [Google Scholar] [CrossRef] [PubMed]
- Rajab, N.F.; Musa, S.M.; Munawar, M.A.; Mun, L.L.; Yen, H.K.; Ibrahim, F.W.; Meng, C.K. Anti-neuroinflammatory Effects of Hibiscus sabdariffa Linn. (Roselle) on Lipopolysaccharides-induced Microglia and Neuroblastoma Cells. J. Sains Kesihat. Malays. 2016, 14, 111–117. [Google Scholar] [CrossRef] [Green Version]
- Galloway, D.A.; Phillips, A.E.M.; Owen, D.R.J.; Moore, C.S. Phagocytosis in the brain: Homeostasis and disease. Front. Immunol. 2019, 10, 790. [Google Scholar] [CrossRef] [Green Version]
- Tobin, M.K.; Bonds, J.A.; Minshall, R.D.; Pelligrino, D.A.; Testai, F.D.; Lazarov, O. Neurogenesis and inflammation after ischemic stroke: What is known and where we go from here. J. Cereb. Blood Flow Metab. 2014, 34, 1573–1584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boshuizen, M.C.S.; Steinberg, G.K. Stem Cell–Based Immunomodulation After Stroke: Effects on Brain Repair Processes. Stroke 2018, 49, 1563–1570. [Google Scholar] [CrossRef] [PubMed]
- Cherry, J.D.; Olschowka, J.A.; O’Banion, M.K. Neuroinflammation and M2 microglia: The good, the bad, and the inflamed. J. Neuroinflamm. 2014, 11, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Häke, I.; Schönenberger, S.; Neumann, J.; Franke, K.; Paulsen-Merker, K.; Reymann, K.; Ismail, G.; Din, L.; Said, I.M.; Latiff, A.; et al. Neuroprotection and enhanced neurogenesis by extract from the tropical plant Knema laurina after inflammatory damage in living brain tissue. J. Neuroimmunol. 2009, 206, 91–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boese, A.C.; Le, Q.-S.E.; Pham, D.; Hamblin, M.H.; Lee, J.-P. Neural stem cell therapy for subacute and chronic ischemic stroke. Stem Cell Res. Ther. 2018, 9, 154. [Google Scholar] [CrossRef] [PubMed]
- Guerra-Crespo, M.; De la Herran-Arita, A.K.; Boronat-Garcia, A.; Maya-Espinosa, G.; Garcia-Montes, J.R.; Fallon, J.H.; Drucker-Colín, R. Neural Stem Cells: Exogenous and Endogenous Promising Therapies for Stroke. Neural Stem Cells Ther. 2012, 297–342. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Wang, J.; Wang, Y.; Yang, G.-Y. The biphasic function of microglia in ischemic stroke. Prog. Neurobiol. 2017, 157, 247–272. [Google Scholar] [CrossRef]
- Ariffin, S.H.Z.; Wahab, R.M.A.; Ismail, I.; Mahadi, N.M.; Ariffin, Z.Z. Stem cells, cytokines and their receptors. Asia-Pac. J. Mol. Biol. Biotechnol. 2005, 13, 1–13. [Google Scholar]
- Hu, X.; Li, P.; Guo, Y.; Wang, H.; Leak, R.K.; Chen, S.; Chen, J. Microglia/Macrophage Polarization Dynamics Reveal Novel Mechanism of Injury Expansion After Focal Cerebral Ischemia. Stroke 2012, 43, 3063–3070. [Google Scholar] [CrossRef] [Green Version]
- Kizil, C.; Kyritsis, N.; Brand, M. Effects of inflammation on stem cells: Together they strive? EMBO Rep. 2015, 16, 416–426. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, A.; Rubin, J.; Clermont, G.; Day, J.; Vodovotz, Y.; Ermentrout, G.B. A reduced mathematical model of the acute inflammatory response: I. Derivation of model and analysis of anti-inflammation. J. Theor. Biol. 2006, 242, 220–236. [Google Scholar] [CrossRef]
- Kumar, R.; Clermont, G.; Vodovotz, Y.; Chow, C.C. The dynamics of acute inflammation. J. Theor. Biol. 2004, 230, 145–155. [Google Scholar] [CrossRef] [Green Version]
- Ma, B.F.; Liu, X.M.; Zhang, A.X.; Wang, P.; Zhang, X.M.; Li, S.N.; Lahn, B.T.; Xiang, A.P. Mathematical models for the proliferation of neural stem/progenitor cells in clonogenic culture. Rejuvenation Res. 2007, 10, 205–214. [Google Scholar] [CrossRef]
- Ziebell, F.; Martin-Villalba, A.; Marciniak-Czochra, A. Mathematical modelling of adult hippocampal neurogenesis: Effects of altered stem cell dynamics on cell counts and bromodeoxyuridine-labelled cells. J. R. Soc. Interface 2014, 11, 20140144. [Google Scholar] [CrossRef] [Green Version]
- Cacao, E.; Cucinotta, F.A. Modeling Impaired Hippocampal Neurogenesis after Radiation Exposure. Radiat. Res. 2016, 185, 319–331. [Google Scholar] [CrossRef]
- Nakata, Y.; Getto, P.; Marciniak-Czochra, A.; Alarcón, T. Stability analysis of multi-compartment models for cell production systems. J. Biol Dyn. 2012, 6, 2–18. [Google Scholar] [CrossRef] [Green Version]
- Alharbi, S.A.; Rambely, A.S. Dynamic Simulation for Analyzing the Effects of the Intervention of Vitamins on Delaying the Growth of Tumor Cells. IEEE Access 2019, 7, 128816–128827. [Google Scholar] [CrossRef]
- Alharbi, S.; Rambely, A. A dynamic simulation of the immune system response to inhibit and eliminate abnormal cells. Symmetry 2019, 11, 572. [Google Scholar] [CrossRef] [Green Version]
- Alharbi, S.A.; Rambely, A.S. Dynamic behaviour and stabilisation to boost the immune system by complex interaction between tumour cells and vitamins intervention. Adv. Differ. Equ. 2020, 1, 1–18. [Google Scholar] [CrossRef]
- Alharbi, S.A.; Rambely, A.S. A New ODE-Based Model for Tumor Cells and Immune System Competition. Mathematics 2020, 8, 1285. [Google Scholar] [CrossRef]
- Stone, L.L.H.; Grande, A.W.; Low, W.C. Neural Repair and Neuroprotection with Stem Cells in Ischemic Stroke. Brain Sci. 2013, 3, 599–614. [Google Scholar] [CrossRef]
- Alqudah, M.A. Cancer treatment by stem cells and chemotherapy as a mathematical model with numerical simulations. Alex. Eng. J. 2020, 59, 1953–1957. [Google Scholar] [CrossRef]
- Wei, L.; Wei, Z.Z.; Jiang, M.Q.; Mohamad, O.; Yu, S.P. Stem cell transplantation therapy for multifaceted therapeutic benefits after stroke. Prog. Neurobiol. 2017, 157, 49–78. [Google Scholar] [CrossRef]
- Zhang, G.-L.; Zhu, Z.-H.; Wang, Y.-Z. Neural stem cell transplantation therapy for brain ischemic stroke: Review and perspectives. World J. Stem Cells 2019, 11, 817–830. [Google Scholar] [CrossRef]
- Guruswamy, R.; ElAli, A. Complex roles of microglial cells in ischemic stroke pathobiology: New insights and future directions. Int. J. Mol. Sci. 2017, 18, 496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Xing, H.; Wan, L.; Jiang, X.; Wang, C.; Wu, Y. Treatment targets for M2 microglia polarization in ischemic stroke. Biomed. Pharmacother. 2018, 105, 518–525. [Google Scholar] [CrossRef]
- Kim, J.Y.; Kim, N.; Yenari, M.A. Mechanisms and potential therapeutic applications of microglial activation after brain injury. CNS Neurosci. Ther. 2019, 21, 309–319. [Google Scholar] [CrossRef]
- Bliss, T.M.; Andres, R.H.; Steinberg, G.K. Optimizing the success of cell transplantation therapy for stroke. Neurobiol. Dis. 2010, 37, 275–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartman, P. Ordinary Differential Equations; John Wiley & Sons: New York, NY, USA; London, UK; Sydney, Australia, 1964. [Google Scholar]
- Gantmacher, F.R. The Theory of Matrices; American Mathematical Society: Providence, RI, USA, 1959; Volume 2. [Google Scholar]
- Liao, L.-Y.; Lau, B.W.-M.; Sánchez-Vidaña, D.I.; Gao, Q. Exogenous neural stem cell transplantation for cerebral ischemia. Neural Regen. Res. 2019, 14, 1129–1137. [Google Scholar] [PubMed]
- Baker, E.W.; Kinder, H.A.; West, F.D. Neural stem cell therapy for stroke: A multimechanistic approach to restoring neurological function. Brain Behav. 2019, 9, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnston, M.D.; Edwards, C.M.; Bodmer, W.F.; Maini, P.K.; Chapman, S.J. Examples of mathematical modeling: Tales from the crypt. Cell Cycle 2007, 6, 2106–2112. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Parameters | Values | Descriptions | Sources |
|---|---|---|---|
| 1 | SCs initial concentration | [31] | |
| 0.4 | damage initial concentration | [4] | |
| 1 | resting microglia initial concentration | [4] | |
| 0.1415 | pro-inflammation initial concentration | [4] | |
| 0.02 | anti-inflammation initial concentration | [4] | |
| 0.69 | the reproduction rate of stem cells | [42] | |
| 0.38 | the resting microglia source | [4] | |
| 0.12 | activation rate of into | [4] | |
| 0.017 | activation rate of into | [4] | |
| 0.11 | the rate transference from to | [4] | |
| 0.2854 | the cytotoxic effects due to | [5] | |
| 0.1 | the death rate of brain cells due to | [5] | |
| 0.1 | the natural death rate of S | [42] | |
| 0.003 | the natural death rate of | [4] | |
| 0.06 | the natural death rate of | [4] | |
| 0.05 | the natural death rate of | [4] | |
| 0.05 | the decay rate of concentration of the D by | [4] | |
| 0.0125 | the decay rate of concentration of the D by | [4] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alqarni, A.J.; Rambely, A.S.; Hashim, I. Dynamical Simulation of Effective Stem Cell Transplantation for Modulation of Microglia Responses in Stroke Treatment. Symmetry 2021, 13, 404. https://doi.org/10.3390/sym13030404
Alqarni AJ, Rambely AS, Hashim I. Dynamical Simulation of Effective Stem Cell Transplantation for Modulation of Microglia Responses in Stroke Treatment. Symmetry. 2021; 13(3):404. https://doi.org/10.3390/sym13030404
Chicago/Turabian StyleAlqarni, Awatif Jahman, Azmin Sham Rambely, and Ishak Hashim. 2021. "Dynamical Simulation of Effective Stem Cell Transplantation for Modulation of Microglia Responses in Stroke Treatment" Symmetry 13, no. 3: 404. https://doi.org/10.3390/sym13030404
APA StyleAlqarni, A. J., Rambely, A. S., & Hashim, I. (2021). Dynamical Simulation of Effective Stem Cell Transplantation for Modulation of Microglia Responses in Stroke Treatment. Symmetry, 13(3), 404. https://doi.org/10.3390/sym13030404