
Protein Helical Structures: Defining Handedness and Localization Features

 ,
,
Abstract
:1. Introduction
2. Computational Methodology
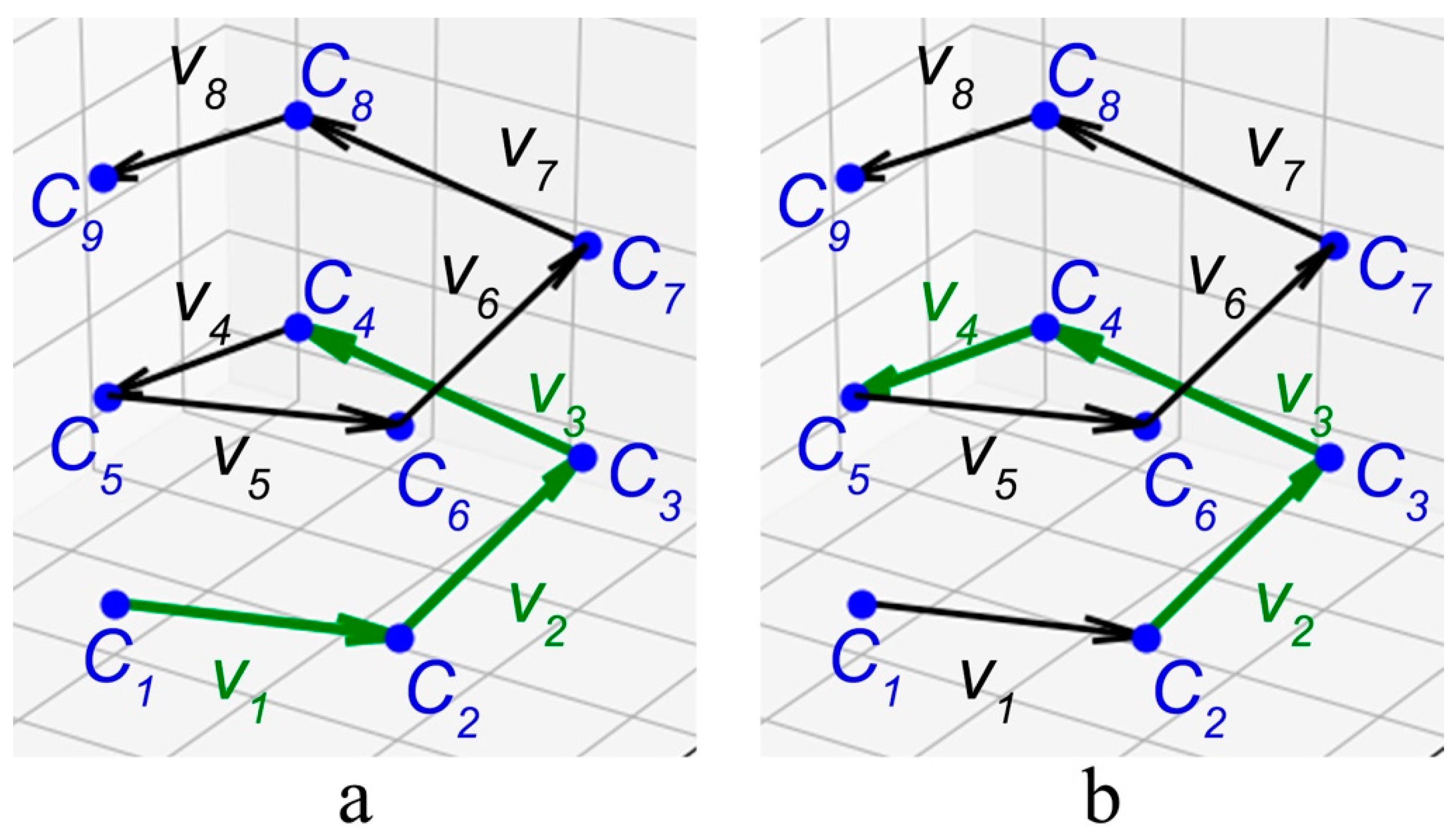
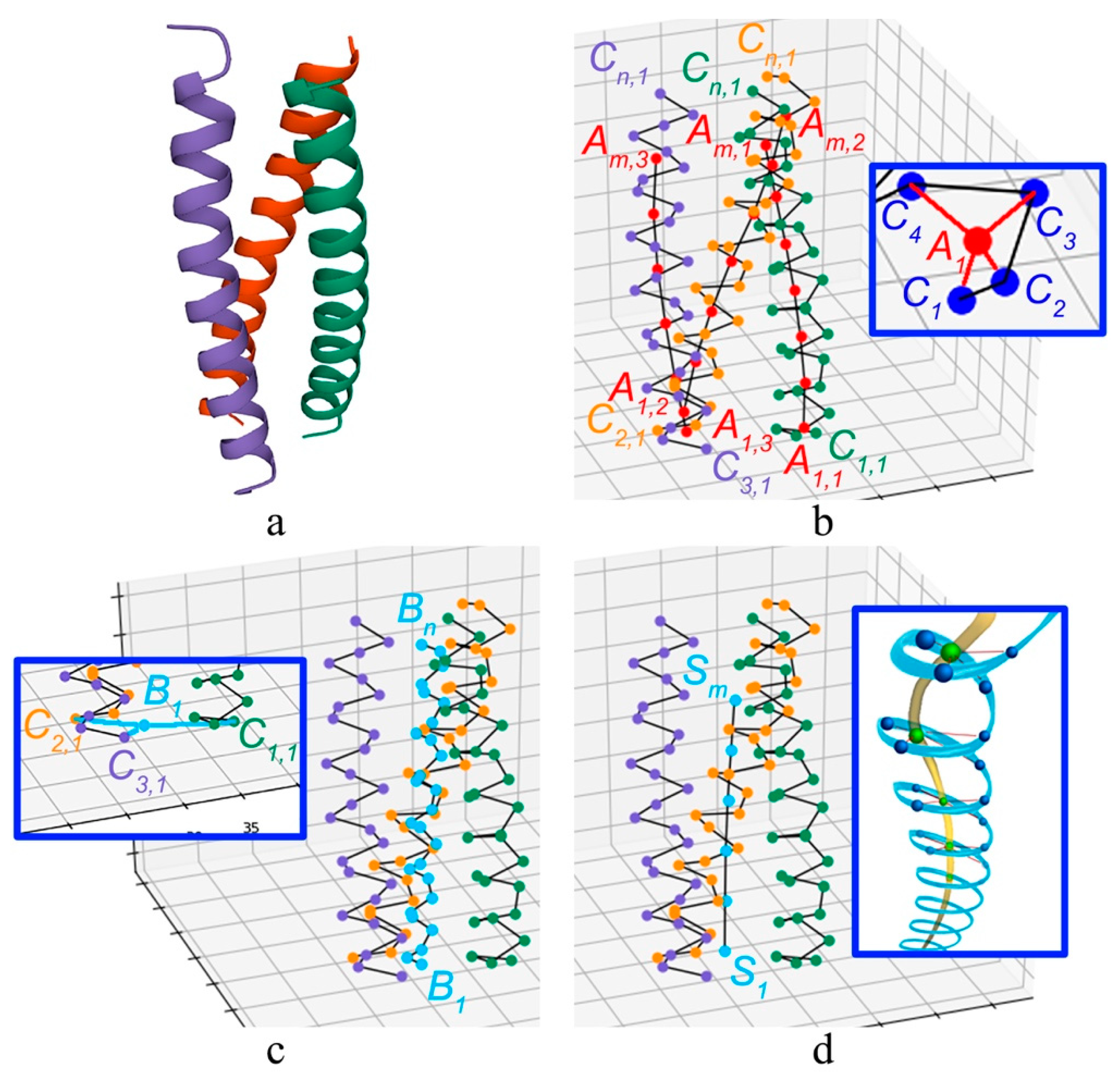
2.1. Method for Quantitative Analysis of the Chirality of Protein Helical Structures
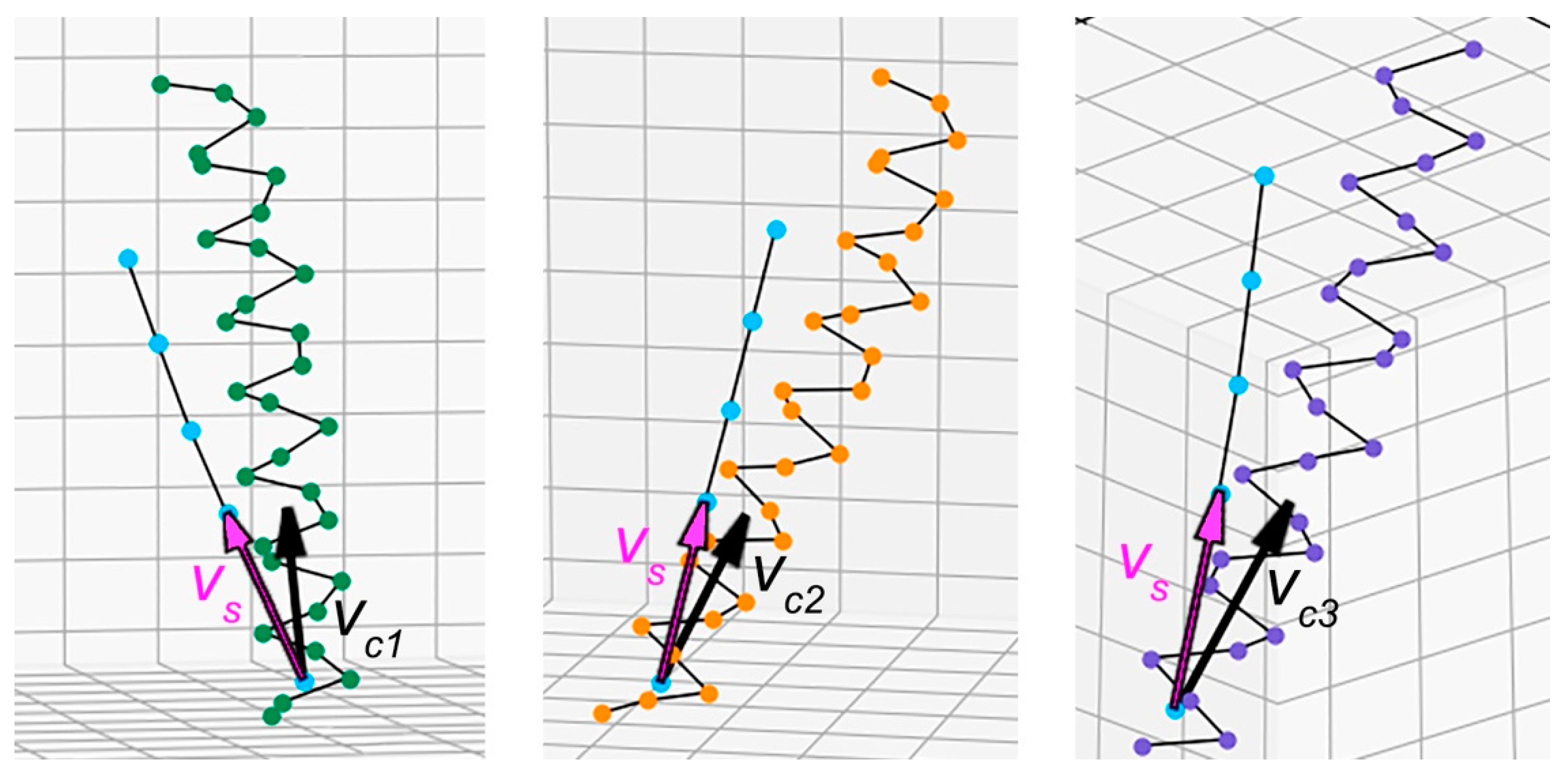
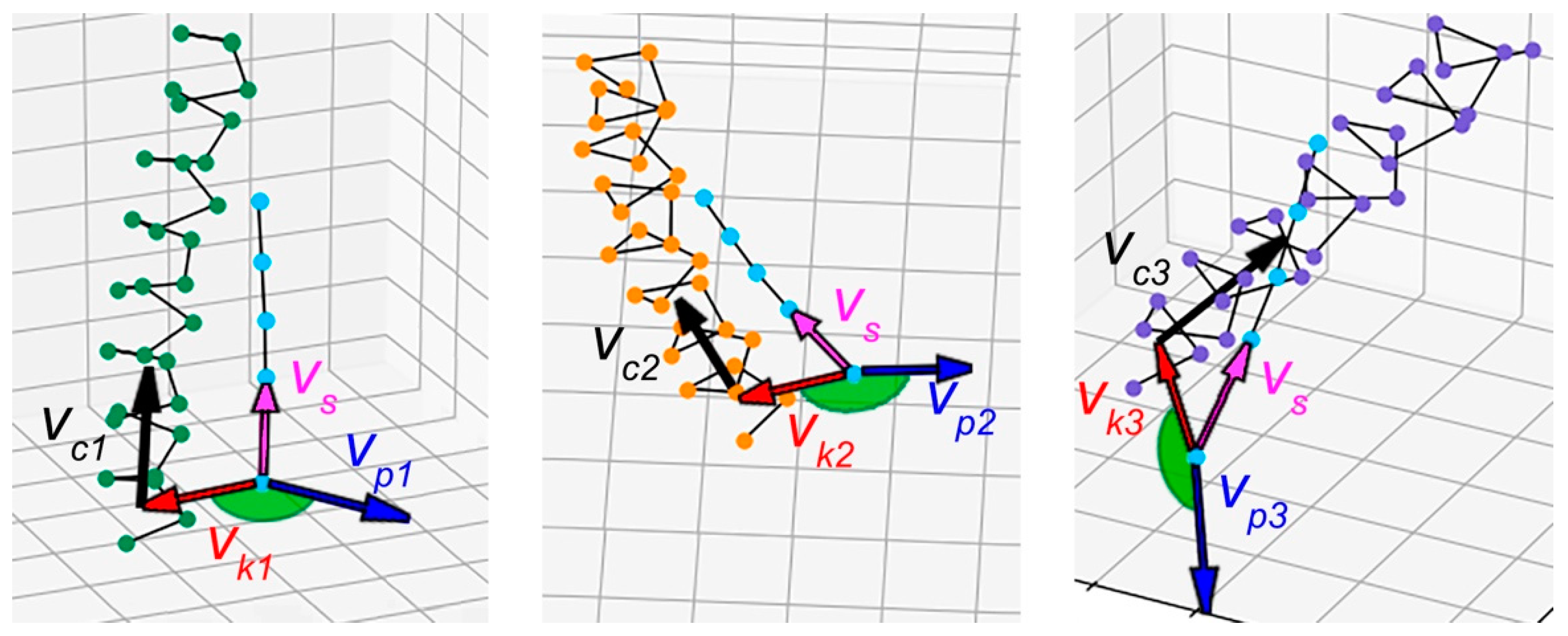
2.2. Method for Quantitative Analysis of the Chirality of Protein Superhelical Structures
3. Results
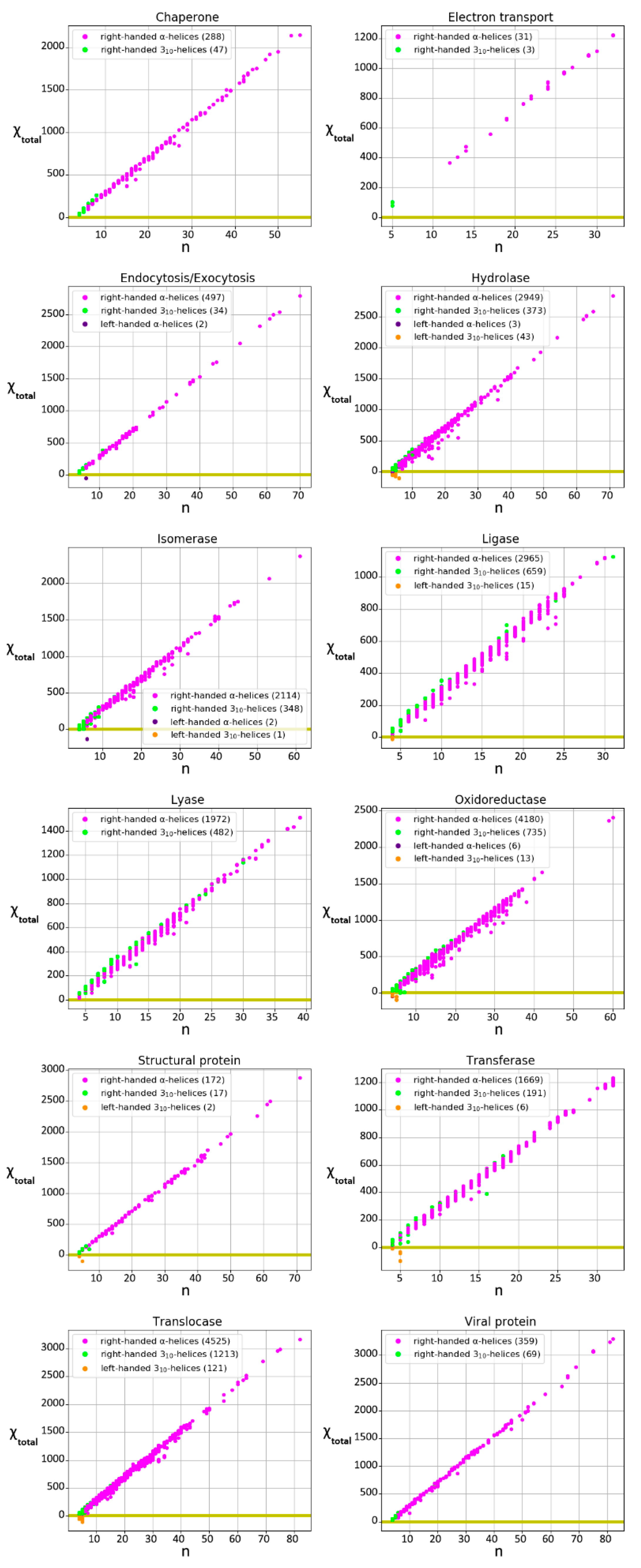
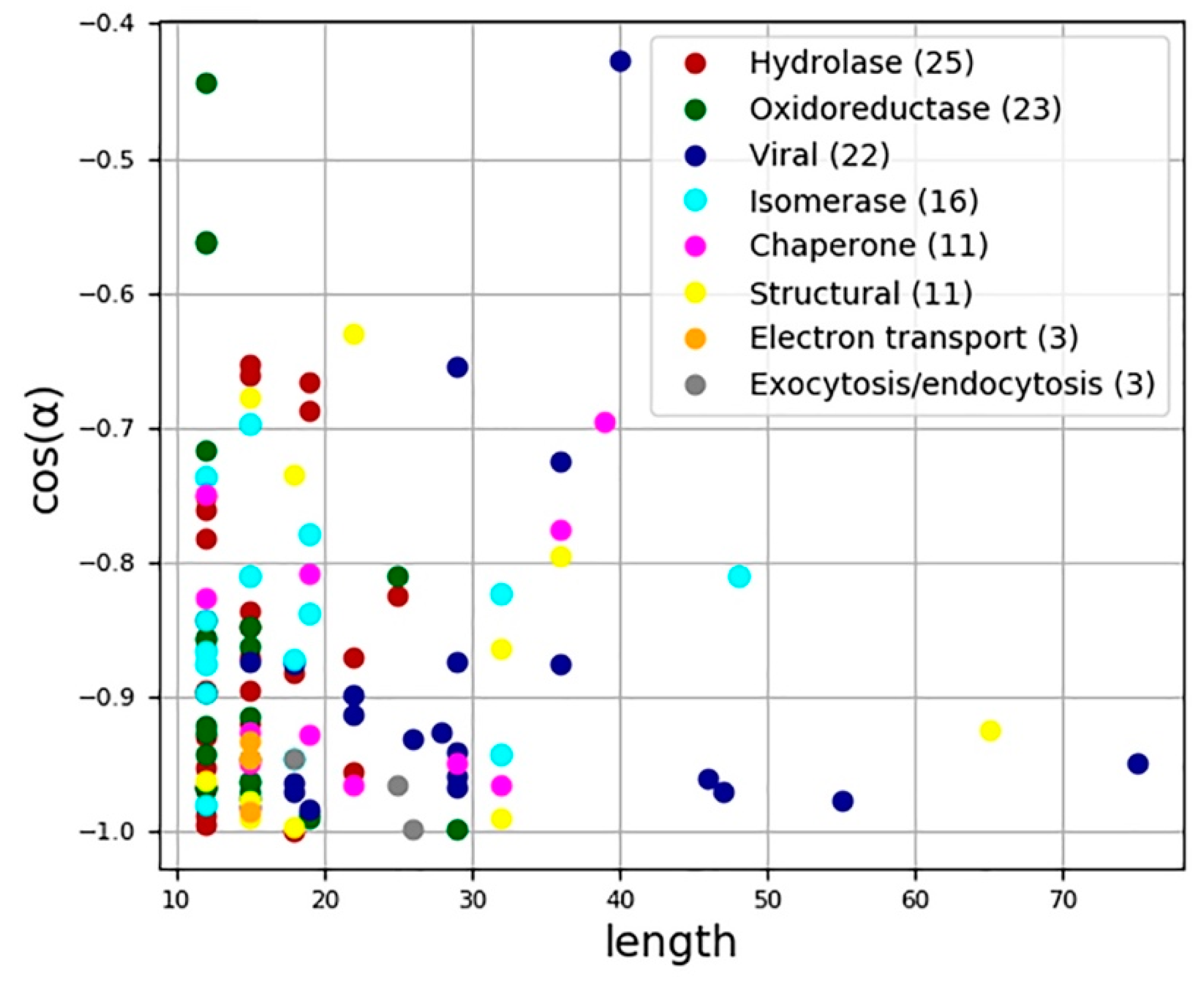
3.1. Characterization of the Chirality of Protein Helical Structures
3.2. Characterization of the Chirality of Protein Superhelical Structures
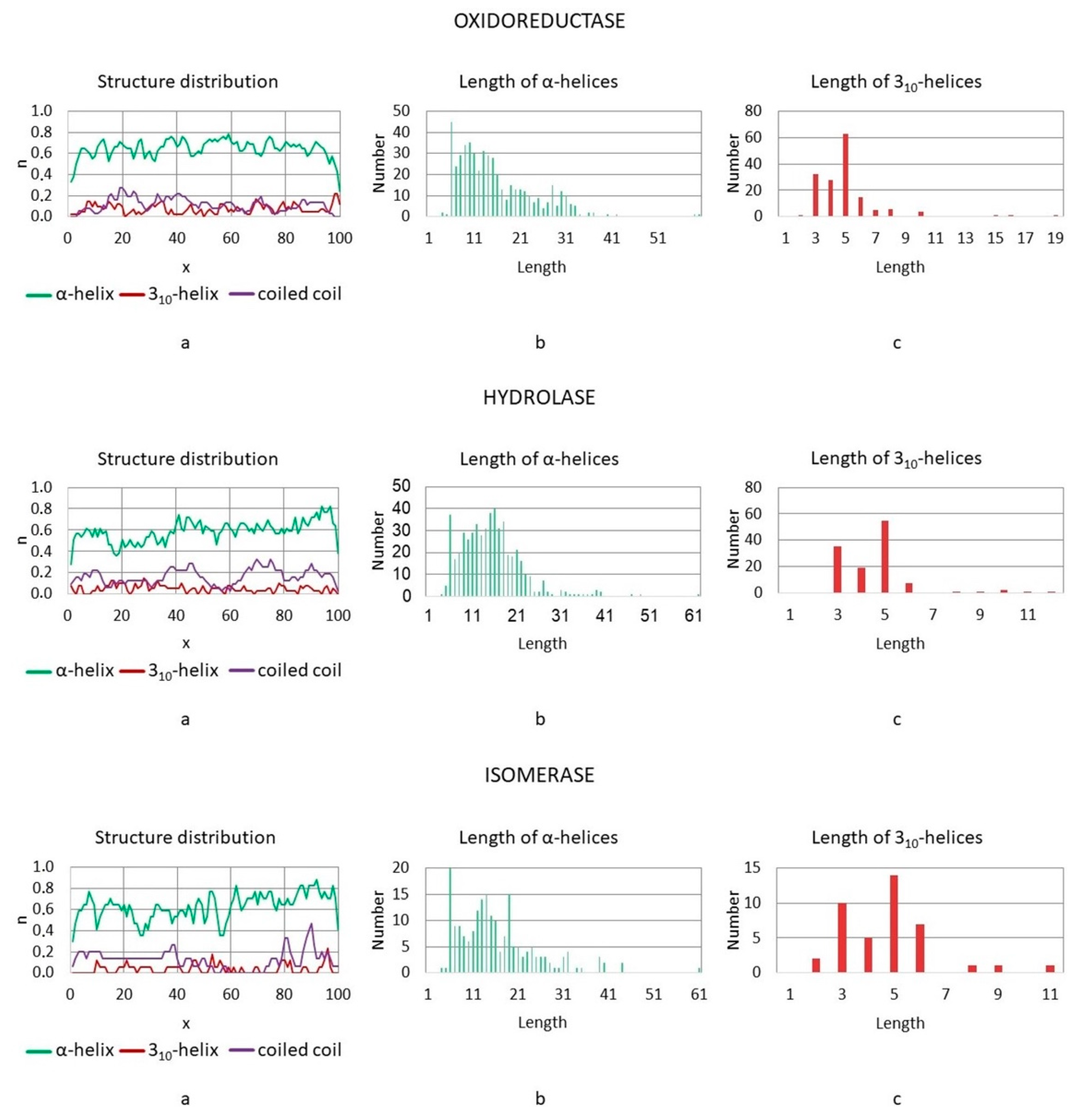
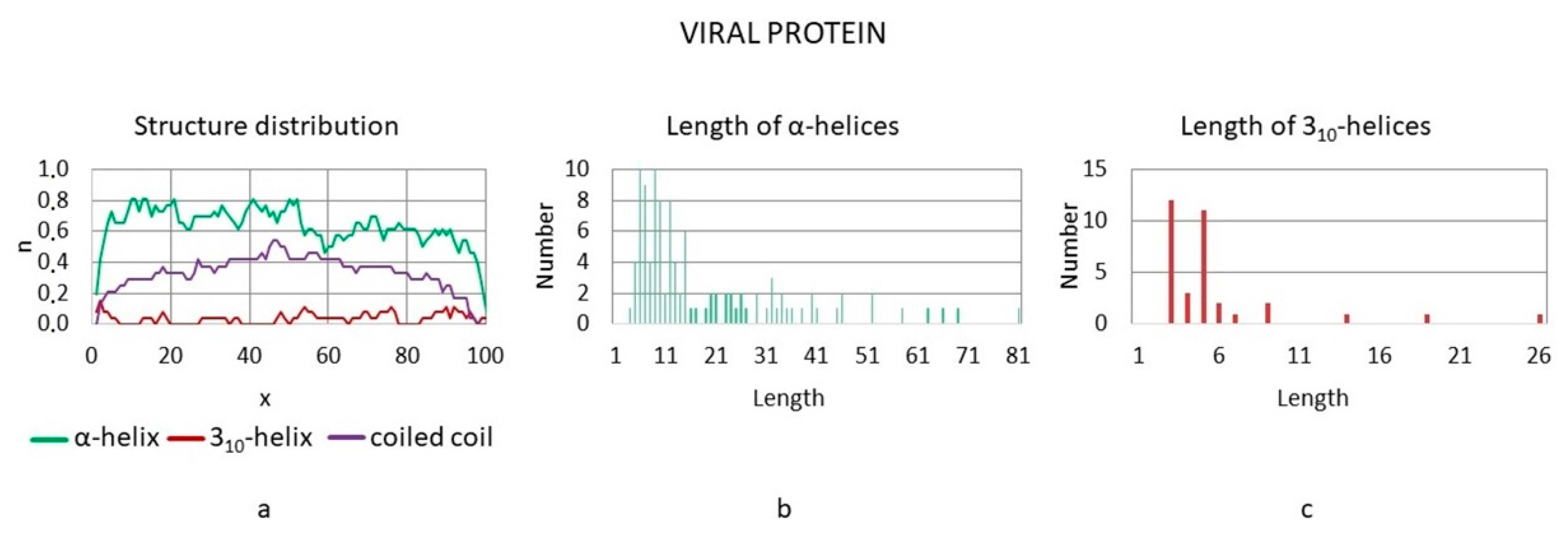
3.3. Distributions of Helical and Superhelical Structures in Polypeptide Chains
3.3.1. Enzymes
3.3.2. Viral Proteins
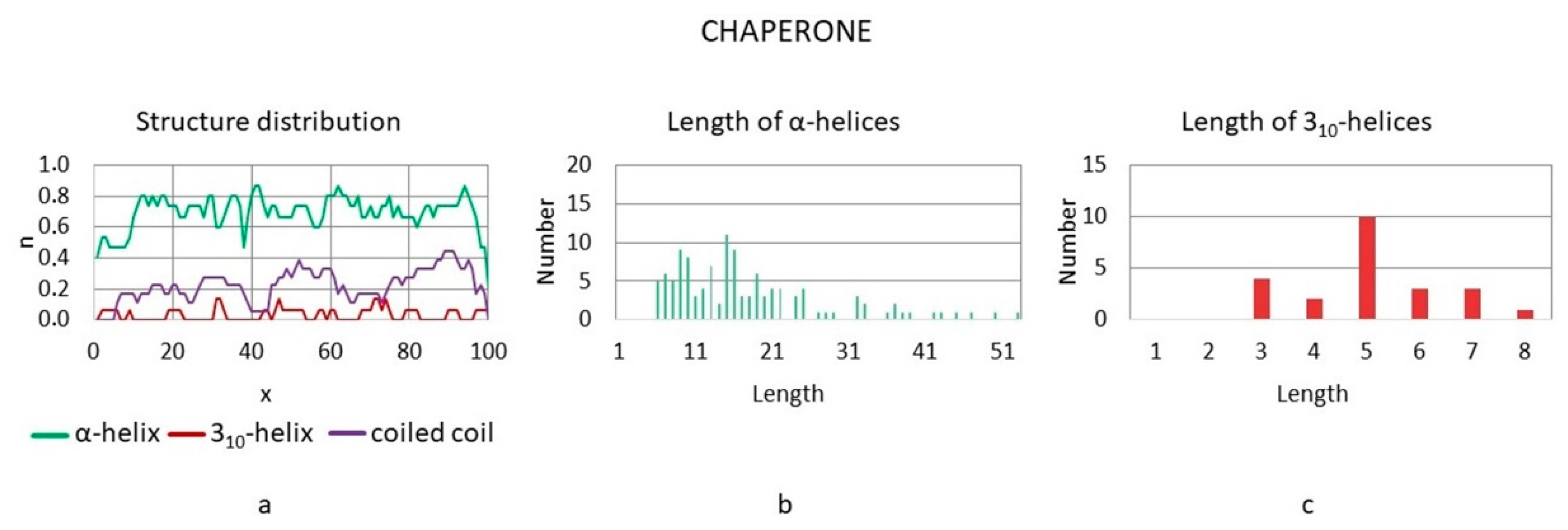
3.3.3. Chaperones
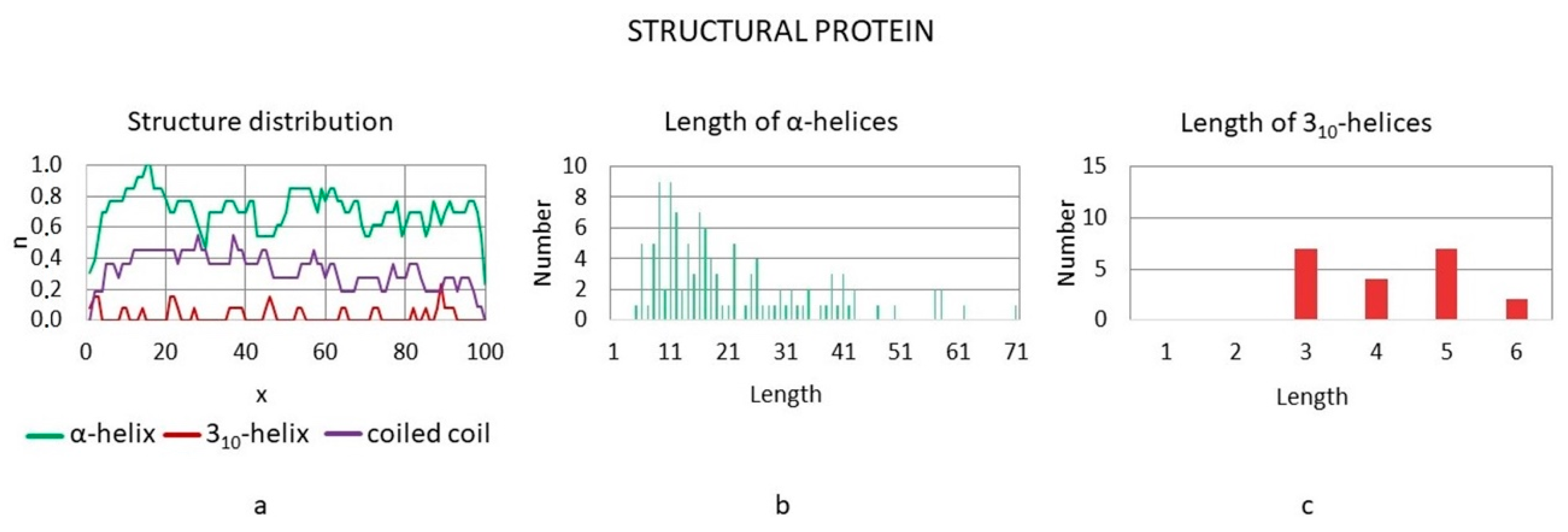
3.3.4. Structural Proteins
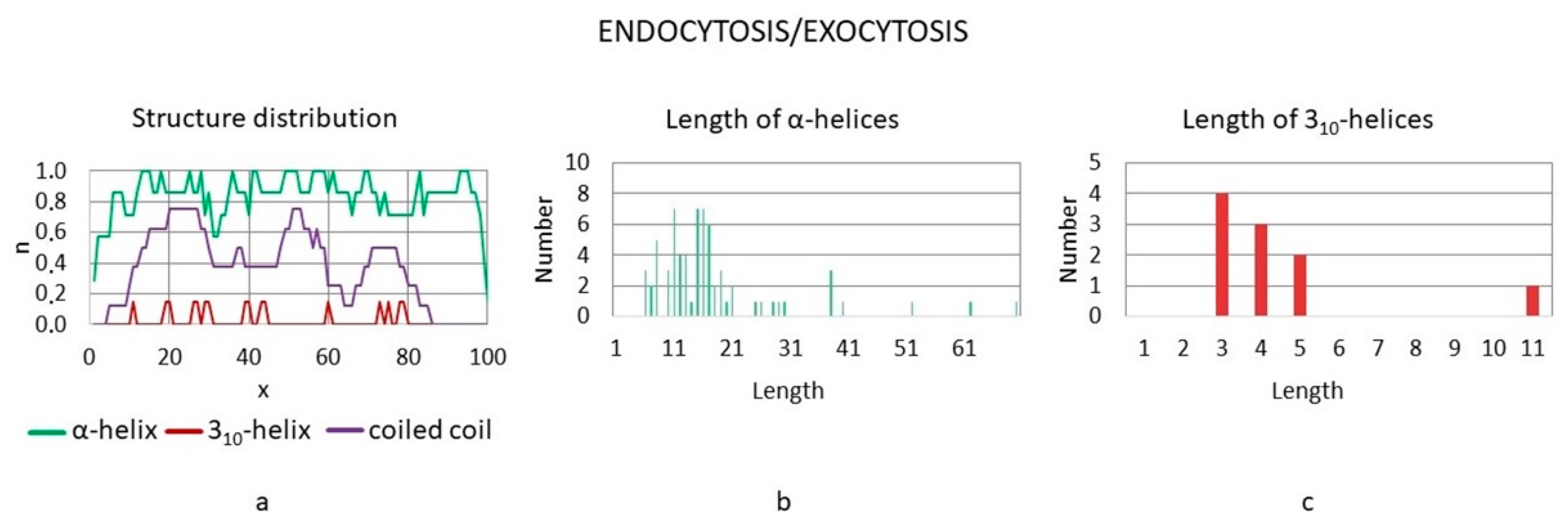
3.3.5. Exocytosis/Endocytosis Proteins
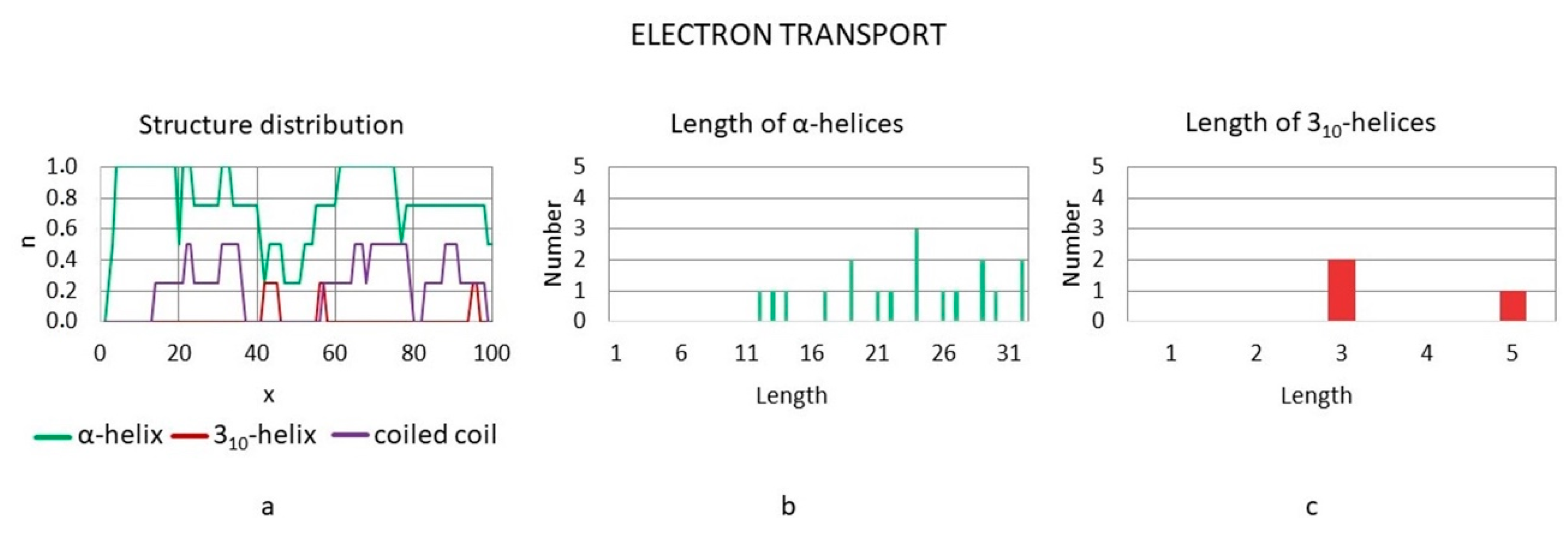
3.3.6. Electron Transport Proteins
4. Discussion
5. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Class | Protein PDB ID | Helix Quantity | Lengths of Helixes (A.a.) | Av. Cosine | Av. Angle | Chirality Sign (L–Left-Handed, R–Right-Handed) |
|---|---|---|---|---|---|---|
| Hydrolase | 1AQT | 2 | 15 | −0.653 | 130.762 | L |
| 1AYX | 2 | 15 | −0.836 | 146.684 | L | |
| 1FS0 | 2 | 25 | −0.825 | 145.588 | L | |
| 1H12 | 2 | 12 | −0.968 | 165.455 | L | |
| 1HUF | 2 | 12 | −0.896 | 153.688 | L | |
| 1K46 | 2 | 12 | −0.895 | 153.535 | L | |
| 1KS8 | 2 | 12 | −0.989 | 171.387 | L | |
| 1L1Y | 2 | 12 | −0.995 | 174.296 | L | |
| 1LF9 | 2 | 15 | −0.662 | 131.436 | L | |
| 1S4B | 2 | 18 | −0.882 | 151.825 | L | |
| 1WCH | 2 | 12 | −0.752 | 138.743 | L | |
| 1WP9 | 2 | 15 | −0.849 | 148.050 | L | |
| 1XZQ | 2 | 19 | −0.666 | 131.735 | L | |
| 1XZQ | 2 | 15 | −0.920 | 156.955 | L | |
| 1YVW | 2 | 18 | −0.999 | 177.619 | L | |
| 1YXB | 2 | 15 | −0.870 | 150.500 | L | |
| 2A8Z | 2 | 12 | −0.783 | 141.528 | L | |
| 2A9U | 2 | 12 | −0.761 | 139.538 | L | |
| 2C6F | 2 | 15 | −0.895 | 153.452 | L | |
| 2CBJ | 2 | 19 | −0.687 | 133.432 | L | |
| 2CFU | 2 | 12 | −0.930 | 158.467 | L | |
| 2CLY | 2 | 12 | −0.953 | 162.345 | L | |
| 2EWF | 2 | 22 | −0.955 | 162.815 | L | |
| 2GTQ | 2 | 12 | −0.843 | 147.444 | L | |
| 2HLD | 2 | 22 | −0.870 | 150.451 | L | |
| Oxidoreductase | 1AFR | 2 | 12 | −0.564 | 124.312 | L |
| 1EZV | 2 | 15 | −0.965 | 164.724 | L | |
| 1FYZ | 2 | 15 | −0.863 | 149.661 | L | |
| 1GU2 | 2 | 12 | −0.942 | 160.418 | L | |
| 1H0O | 2 | 15 | −0.975 | 167.093 | L | |
| 1IS2 | 2 | 12 | −0.857 | 149.013 | L | |
| 1JKU | 2 | 19 | −0.987 | 170.838 | L | |
| 1MHY | 2 | 15 | −0.963 | 164.298 | L | |
| 1NG3 | 2 | 12 | −0.855 | 148.812 | L | |
| 1OTK | 2 | 15 | −0.848 | 148.023 | L | |
| 1PHZ | 2 | 12 | −0.921 | 157.041 | L | |
| 1R2F | 2 | 12 | −0.561 | 124.148 | L | |
| 1U6I | 2 | 12 | −0.969 | 165.689 | L | |
| 1YQ9 | 2 | 12 | −0.967 | 165.141 | L | |
| 1YUX | 2 | 25 | −0.810 | 144.082 | L | |
| 1ZA0 | 2 | 19 | −0.991 | 172.268 | L | |
| 1ZOY | 2 | 12 | −0.718 | 135.855 | L | |
| 2B0T | 2 | 12 | −0.444 | 116.333 | L | |
| 2C2X | 2 | 15 | −0.971 | 166.152 | L | |
| 2CWL | 2 | 12 | −0.927 | 157.969 | L | |
| 2F1K | 2 | 15 | −0.946 | 161.114 | L | |
| 2FYN | 2 | 15 | −0.915 | 156.240 | L | |
| 2HKO | 2 | 29 | −0.998 | 176.265 | L | |
| Viral | 1EBO | 3 | 22 | −0.914 | 157.015 | L |
| 1EBO | 3 | 36 | −0.726 | 136.569 | L | |
| 1EIA | 2 | 18 | −0.964 | 164.508 | L | |
| 1EZJ | 4 | 40 | −0.426 | 115.268 | L | |
| 1G2C | 3 | 29 | −0.960 | 164.582 | L | |
| 1G5G | 3 | 47 | −0.970 | 169.015 | L | |
| 1HTM | 3 | 55 | −0.977 | 168.418 | L | |
| 1LJ2 | 2 | 18 | −0.970 | 165.910 | L | |
| 1LJ2 | 2 | 19 | −0.984 | 169.846 | L | |
| 1MG1 | 3 | 29 | −0.941 | 162.021 | L | |
| 1MOF | 3 | 29 | −0.967 | 166.447 | L | |
| 1NO4 | 2 | 36 | −0.876 | 151.169 | L | |
| 1SFK | 2 | 18 | −0.875 | 151.088 | L | |
| 1SLQ | 3 | 15 | −0.983 | 170.282 | L | |
| 1SVF | 3 | 46 | −0.960 | 165.161 | L | |
| 1WYY | 3 | 75 | −0.950 | 165.976 | L | |
| 1ZTM | 3 | 29 | −0.874 | 153.034 | L | |
| 1ZVB | 3 | 22 | −0.899 | 155.777 | L | |
| 2BEQ | 3 | 26 | −0.931 | 161.373 | L | |
| 2C9L | 2 | 29 | −0.655 | 130.887 | L | |
| 2FXP | 3 | 28 | −0.927 | 161.728 | L | |
| 2GUM | 3 | 15 | −0.874 | 152.780 | L | |
| Isomerase | 1A36 | 2 | 18 | −0.946 | 161.033 | L |
| 1FP3 | 2 | 18 | −0.872 | 150.673 | L | |
| 1IHG | 2 | 12 | −0.843 | 147.459 | L | |
| 1IIP | 2 | 48 | −0.811 | 144.150 | L | |
| 1N1B | 2 | 12 | −0.897 | 153.793 | L | |
| 1P5Q | 2 | 19 | −0.838 | 146.898 | L | |
| 1P5Q | 2 | 12 | −0.736 | 137.413 | L | |
| 1QZ2 | 2 | 19 | −0.808 | 143.890 | L | |
| 1QZ2 | 2 | 12 | −0.866 | 150.011 | L | |
| 1 × 94 | 2 | 12 | −0.980 | 168.509 | L | |
| 1XRS | 2 | 12 | −0.876 | 151.123 | L | |
| 1YBZ | 2 | 32 | −0.824 | 145.444 | L | |
| 1ZVU | 2 | 15 | −0.698 | 134.264 | L | |
| 2D8E | 2 | 32 | −0.943 | 160.643 | L | |
| 2FEL | 2 | 15 | −0.811 | 144.154 | L | |
| 2NOV | 2 | 19 | −0.780 | 141.220 | L | |
| Chaperone | 1ELR | 2 | 12 | −0.826 | 145.686 | L |
| 1FXK | 2 | 36 | −0.775 | 140.815 | L | |
| 1FXK | 2 | 39 | −0.696 | 134.088 | L | |
| 1I6Z | 2 | 22 | −0.966 | 165.006 | L | |
| 1ORJ | 2 | 19 | −0.927 | 158.032 | L | |
| 1OX3 | 3 | 32 | −0.966 | 167.239 | L | |
| 1QVR | 2 | 15 | −0.949 | 161.707 | L | |
| 1QZ2 | 2 | 19 | −0.808 | 143.890 | L | |
| 1SG2 | 2 | 15 | −0.927 | 157.908 | L | |
| 1XOU | 2 | 29 | −0.950 | 161.774 | L | |
| 1YT1 | 2 | 12 | −0.750 | 138.560 | L | |
| Structural | 1DEB | 2 | 36 | −0.796 | 142.716 | L |
| 1IO1 | 2 | 32 | −0.865 | 149.854 | L | |
| 1LUJ | 2 | 15 | −0.677 | 132.604 | L | |
| 1R0D | 2 | 15 | −0.977 | 167.644 | L | |
| 1U89 | 2 | 18 | −0.997 | 175.819 | L | |
| 1U89 | 2 | 12 | −0.962 | 164.187 | L | |
| 1WU9 | 2 | 32 | −0.989 | 171.630 | L | |
| 1 × 8Y | 2 | 65 | −0.925 | 157.610 | L | |
| 2D4Y | 2 | 22 | −0.630 | 129.025 | L | |
| 2D4Y | 2 | 15 | −0.990 | 171.794 | L | |
| 2F6H | 2 | 18 | −0.735 | 137.274 | L | |
| Electron transport | 1B71 | 2 | 15 | −0.946 | 161.105 | L |
| 1J30 | 2 | 15 | −0.985 | 169.947 | L | |
| 1LM3 | 2 | 15 | −0.932 | 158.825 | L | |
| Exocytosis/endocytosis | 1HVV | 2 | 26 | −0.999 | 177.304 | L |
| 1L4A | 2 | 25 | −0.966 | 164.976 | L | |
| 1URU | 2 | 18 | −0.946 | 161.156 | L |
References
- Pasteur, L. Recherches sur les relations qui peuvent exister entre la forme crystalline, la composition chimique et le sens de la polarisation rotatoire. Ann. Chim. Phys. 1848, 24, 442–459. [Google Scholar]
- Schwartz, A.W. Origin of life. The origin of macromolecular chirality. Curr. Biol. 1994, 4, 758–760. [Google Scholar] [CrossRef]
- Podlech, J. Origin of organic molecules and biomolecular homochirality. Cell Mol. Life Sci. 2001, 58, 44–60. [Google Scholar] [CrossRef] [PubMed]
- Hein, J.E.; Blackmond, D.G.; On Kojić-Prodić, B.; Štefanić, Z. Symmetry versus Asymmetry in the Molecules of Life: Homomeric Protein Assemblies. Symmetry 2010, 2, 884–906. [Google Scholar] [CrossRef] [Green Version]
- Hein, J.E.; Blackmond, D.G. On the origin of single chirality of amino acids and sugars in biogenesis. Acc. Chem. Res. 2012, 45, 2045–2054. [Google Scholar] [CrossRef] [PubMed]
- Inaki, M.; Liu, J.; Matsuno, K. Cell chirality: Its origin and roles in left-right asymmetric development. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2016, 371, 20150403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dorta-Urra, A.; Bargueño, P. Homochirality: A Perspective from Fundamental Physics. Symmetry 2019, 11, 661. [Google Scholar] [CrossRef] [Green Version]
- Blackmond, D.G. The Origin of Biological Homochirality. Cold Spring Harb. Perspect. Biol. 2019, 11, a032540. [Google Scholar] [CrossRef] [Green Version]
- Famiano, M.; Boyd, R.; Kajino, T.; Onaka, T.; Mo, Y. Astrophysical Sites that Can Produce Enantiomeric Amino Acids. Symmetry 2019, 11, 23. [Google Scholar] [CrossRef] [Green Version]
- Tverdislov, V.A. Chirality as a primary switch of hierarchical levels in molecular biological systems. Biophysics 2013, 58, 128–132. [Google Scholar] [CrossRef]
- Tverdislov, V.A.; Malyshko, E.V. On regularities of spontaneous formation of structural hierarchies in chiral systems of non-living and living nature. Phys. Uspekhi. 2019, 189, 375–385. [Google Scholar] [CrossRef]
- Tverdislov, V.A.; Malyshko, E.V. Chiral Dualism as an Instrument of Hierarchical Structure Formation in Molecular Biology. Symmetry 2020, 12, 587. [Google Scholar] [CrossRef] [Green Version]
- Pauling, L.; Corey, R.B. Two hydrogen-bonded spiral configurations of the polypeptide chain. J. Am. Chem. Soc. 1950, 72, 5349. [Google Scholar] [CrossRef]
- Pauling, L.; Corey, R.B. The structure of synthetic polypeptides. Proc. Natl. Acad. Sci. USA 1951, 37, 241–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pauling, L.; Corey, R.B. A proposed structure for the nucleic acids. Proc. Natl. Acad. Sci. USA 1953, 39, 84–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pauling, L.; Corey, R.B. The pleated sheet, a new layer configuration of polypeptide chains. Proc. Natl. Acad. Sci. USA 1951, 37, 2451–2456. [Google Scholar] [CrossRef] [Green Version]
- Branden, C.; Tooze, J. Introduction to Protein Structure, 2nd ed.; Garland Publishing: New York, NY, USA, 1999; 410p. [Google Scholar]
- Novotny, M.; Kleywegt, G.J. A Survey of Left-handed Helices in Protein Structures. J. Mol. Biol. 2005, 347, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Barlow, D.J.; Thornton, J.M. Helix geometry in proteins. J. Mol. Biol. 1988, 201, 601–619. [Google Scholar] [CrossRef]
- Herzberg, O.; Moult, J. Analysis of steric strain in the polypeptide backbone of protein models. Proteins 1991, 11, 223–229. [Google Scholar] [CrossRef]
- Petock, J.M.; Torshin, I.Y.; Weber, I.T.; Harrison, R.W. Analysis of protein structures reveals regions of rare backbone conformation at functional sites. Proteins 2003, 53, 872–879. [Google Scholar] [CrossRef]
- Crick, F.H.C. The packing of α-helices: Simple coiled-coils. Acta Cryst. 1953, 6, 689–697. [Google Scholar] [CrossRef] [Green Version]
- Lupas, A.; Gruber, M. The structure of alpha-helical coiled coils. Adv. Protein. Chem. 2005, 70, 37–78. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, M.D.; Mendler, C.T.; Bassler, J.; Karamichali, I.; Ridderbusch, O.; Lupas, A.N.; Alvarez, B.H. α/β Coiled coils. eLife 2016, 5, e11861. [Google Scholar] [CrossRef] [Green Version]
- Mortenson, D.E.; Steinkruger, J.D.; Kreitler, D.F.; Perroni, D.V.; Sorenson, G.P.; Huang, L.; Mittal, R.; Yun, H.G.; Travis, B.R.; Mahanthappa, M.K.; et al. High-resolution structures of a heterochiral coiled coil. Proc. Natl. Acad. Sci. USA 2015, 112, 13144–13149. [Google Scholar] [CrossRef] [Green Version]
- Mezey, P. Chirality Measures and Graph Representations. Comput. Math. Appl. 1997, 34, 105–112. [Google Scholar] [CrossRef] [Green Version]
- Zabrodsky, H.; Peleg, S.; Avnir, D. Continuous symmetry measures. J. Am. Chem. Soc. 1992, 114, 7843–7851. [Google Scholar] [CrossRef]
- Zabrodsky, H.; Peleg, S.; Avnir, D. Symmetry as a Continuous Feature. IEEE Trans. Pattern Anal. 1995, 17, 1154–1166. [Google Scholar] [CrossRef] [Green Version]
- Pinsky, M.; Dryzun, C.; Casanova, D.; Alemany, P.; Avnir, D. Analytical methods for calculating continuous symmetry measures and the chirality measure. J. Comput. Chem. 2008, 29, 2712–2721. [Google Scholar] [CrossRef]
- Luzanov, A.V. Simple one-electron invariants of molecular chirality. J. Math. Chem. 2006, 41, 417–435. [Google Scholar] [CrossRef]
- Randic, M.; Razinger, M. Molecular shapes and chirality. J. Chem. Inf. Comp. Sci. 1996, 36, 429–441. [Google Scholar] [CrossRef]
- Randic;, M. Graph theoretical descriptors of two-dimensional chirality with possible extension to three-dimensional chirality. J. Chem. Inf. Comput. Sci. 2001, 41, 639–649. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Zhang, Q.; Long, H.; Xu, L. Graph theoretical representation of atomic asymmetry and molecular chirality of benzenoids in two-dimensional space. PLoS ONE 2014, 9, e102043. [Google Scholar] [CrossRef]
- Gilat, G. Chiral coefficient-a measure of the amount of structural chirality. J. Phys. A Math. Gen. 1989, 22, L545–L550. [Google Scholar] [CrossRef]
- Kuz’min, V.E.; Stel’makh, B.; Bekker, M.B.; Pozigun, D.V. Quantitative aspects of chirality. I. Method of dissymmetry function. J. Phys. Org. Chem. 1992, 5, 295–298. [Google Scholar] [CrossRef]
- Ramachandran, G.N.; Ramakrishnan, C.; Sasisekharan, V. Stereochemistry of polypeptide chain configurations. J. Mol. Biol. 1963, 7, 95–99. [Google Scholar] [CrossRef]
- Ramachandran, G.N.; Sasiskharan, V. Conformation of polypeptides and proteins. Adv. Protein Chem. 1968, 23, 283–437. [Google Scholar] [CrossRef] [PubMed]
- Mannige, R. An exhaustive survey of regular peptide conformations using a new metric for backbone handedness (h). PeerJournal 2017, 5, e3327. [Google Scholar] [CrossRef]
- Petitjean, M. Chirality and Symmetry Measures: A Transdisciplinary Review. Entropy 2003, 5, 271–312. [Google Scholar] [CrossRef]
- Petitjean, M. On the root mean square quantitative chirality and quantitative symmetry measures. J. Math. Phys. 1999, 40, 4587–4595. [Google Scholar] [CrossRef] [Green Version]
- Petitjean, M. Chiral mixtures. J. Math. Phys. 2002, 43, 4147–4157. [Google Scholar] [CrossRef] [Green Version]
- Fraser, R.D.B.; MacRae, T.P. Conformation in Fibrous Proteins and Related Synthetic Polypeptides, 1st ed.; Academic Press: New York, NY, USA, 1973; 628p. [Google Scholar]
- Phillips, G.N. What is the pitch of the α-helical coiled coil? Proteins 1992, 14, 425–429. [Google Scholar] [CrossRef]
- Neukirch, S.; Goriely, A.; Hausrath, A.C. Chirality of coiled-coils: Elasticity matters. Phys. Rev. Lett. 2008, 100, 038105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- RCSB PDB. Available online: http://www.rcsb.org (accessed on 1 March 2021).
- CC+ Database. Available online: http://coiledcoils.chm.bris.ac.uk/ccplus/search/ (accessed on 1 March 2021).
- Sidorova, A.E.; Malyshko, E.V.; Kotov, A.R.; Tverdislov, V.A.; Ustinin, M.N. Quantitative Criteria of Chirality in Hierarchical Protein Structures. Biophysics 2019, 64, 155–166. [Google Scholar] [CrossRef]
- Nautiyal, S.; Alber, T. Crystal structure of a designed, thermostable, heterotrimeric coiled coil. Protein Sci. 1999, 8, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Malyshko, E.V.; Bagrova, O.E.; Tverdislov, V.A. The relationship between hierarchical chiral structures of proteins and their functions. Biophysics 2020, 65, 368–373. [Google Scholar] [CrossRef]
- Efimov, A. Chirality and Handedness of Protein Structures. Biochemistry 2018, 83, S103–S110. [Google Scholar] [CrossRef]
- Zayit, A.; Pinsky, M.; Elgavi, H.; Dryzun, C.; Avnir, D. A web site for calculating the degree of chirality. Chirality 2011, 23, 17–23. [Google Scholar] [CrossRef]
- Plazinska, A.; Plazinski, W. Chirality Effects in Biomolecular Systems: Calculation of the Relative Free Energies by Molecular Dynamics Simulations. J. Chem. Inf. Model. 2020, 60, 5424–5436. [Google Scholar] [CrossRef] [PubMed]
- Partovi, T. Calculation of the Chirality in Solutions. Chem. Lett. 2012, 41, 760–762. [Google Scholar] [CrossRef]
- Fowler, P. Quantification of chirality: Attempting the impossible. Symmetry Cult. Sci. 2003, 16, 321–334. [Google Scholar]












| Proteins | α-helices | |||||||
|---|---|---|---|---|---|---|---|---|
| Number of Right-Handed α-helices | Number of Left-Handed α-helices | -helices | ||||||
| Oxidoreductase | 4180 | 0.02–3.005 | 6 | (−0.054)– (−0.046) | 735 | 0.008–1.725 | 13 | (−0.115)– (−0.005) |
| Transferase | 1669 | 0.015–1.559 | - | - | 191 | 0.025–0.881 | 6 | (−0.122)– (−0.010) |
| Hydrolase | 2949 | 0.026–3.528 | 3 | (−0.047)– (−0.008) | 373 | 0.034–0.773 | 43 | (−0.132)– (−0.003) |
| Lyase | 1972 | 0.018–1.870 | - | - | 482 | 0.038–1.454 | - | - |
| Isomerase | 2114 | 0.030–2.991 | 2 | (−0.162)– (−0.161) | 348 | 0.004–1.263 | 1 | (−0.001) |
| Ligase | 2965 | 0.007–1.425 | - | - | 659 | 0.002–1.420 | 15 | (−0.015)– (−0.002) |
| Translocase | 4525 | 0.009–4.019 | - | - | 1213 | 0.012–1.258 | 121 | (−0.128)– (−0.005) |
| Chaperone | 288 | 0.122–2.708 | - | - | 47 | 0.042–0.324 | - | - |
| Viral protein | 359 | 0.043–4.137 | - | - | 69 | 0.038–1.227 | - | - |
| Structural protein | 172 | 0.097–3.532 | - | - | 17 | 0.044–0.180 | 2 | (−0.118), (−0.035) |
| Endo/exocytosis protein | 497 | 0.135–3.480 | 2 | (−0.067)– (−0.066) | 34 | 0.043–0.479 | - | - |
| Electron transport protein | 31 | 0.468–1.550 | - | - | 3 | 0.1–0.122 | - | - |
| Protein Class | Percentage of Left-Handed α-helices | |||
|---|---|---|---|---|
| Oxidoreductase | 4186/748 | 6/13 | 0.14 | 1.74 |
| Transferase | 1669/197 | -/6 | - | 3.04 |
| Hydrolase | 2952/416 | 3/43 | 0.1 | 10.34 |
| Lyase | 1972/482 | -/- | - | - |
| Isomerase | 2116/349 | 2/1 | 0.09 | 0.29 |
| Ligase | 2965/674 | -/15 | - | 2.23 |
| Translocase | 4525/1334 | -/121 | - | 9.07 |
| Chaperone | 288/47 | -/- | - | - |
| Viral protein | 359/69 | -/- | - | - |
| Structural protein | 172/19 | -/2 | - | 10.53 |
| Endo/exocytosis protein | 499/34 | 2/- | 0.4 | - |
| Electron transport protein | 31/3 | -/- | - | - |
| Total | 21374/4372 | 13/201 | 0.06 | 4.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sidorova, A.E.; Malyshko, E.V.; Lutsenko, A.O.; Shpigun, D.K.; Bagrova, O.E. Protein Helical Structures: Defining Handedness and Localization Features. Symmetry 2021, 13, 879. https://doi.org/10.3390/sym13050879
Sidorova AE, Malyshko EV, Lutsenko AO, Shpigun DK, Bagrova OE. Protein Helical Structures: Defining Handedness and Localization Features. Symmetry. 2021; 13(5):879. https://doi.org/10.3390/sym13050879
Chicago/Turabian StyleSidorova, Alla E., Ekaterina V. Malyshko, Aleksey O. Lutsenko, Denis K. Shpigun, and Olga E. Bagrova. 2021. "Protein Helical Structures: Defining Handedness and Localization Features" Symmetry 13, no. 5: 879. https://doi.org/10.3390/sym13050879
APA StyleSidorova, A. E., Malyshko, E. V., Lutsenko, A. O., Shpigun, D. K., & Bagrova, O. E. (2021). Protein Helical Structures: Defining Handedness and Localization Features. Symmetry, 13(5), 879. https://doi.org/10.3390/sym13050879