1. Introduction
Common bread wheat (
Triticum aestivum L.) is one of the most important cereals, and provides the basis for the nutrition of the Earth’s population. More than a quarter of the world’s grain production is provided by the cultivation of this cereal, which is a food source for more than one-fifth of the world’s population [
1]. In addition, grains and their products provide more than 20% of calories and protein for the world’s population [
2].
To effectively create and evaluate new highly productive varieties and lines of wheat resistant to biotic and abiotic stresses, it is necessary to understand the subtle mechanisms of kernel formation, while taking into account its qualitative characteristics. Wheat grain, like other cereals, is a dry one-seeded fruit (kernel). To evaluate the effectiveness of fertilizers, stimulants, and protectants, wheat grains (kernels) are a traditional model object. However, the apparent simplicity and ideality of this model often turns out to be imaginary. In our recent work, it was demonstrated that the hypothetical idealized model of wheat seedling development was far from ideal, which became especially noticeable when exposed to adverse factors such as salinity [
3]. It was found that in most cases the root system developed unevenly and showed clear signs of developmental asymmetry. The fruit of wheat, the kernel, was also capable of exhibiting varying degrees of asymmetry.
The grain quality of wheat equally depends on the varietal characteristics and manifestation of the genotype [
4], as well as the susceptibility of the genotype to adverse environmental conditions, such as drought, salinity, high and low temperatures [
5,
6,
7], and pests. All these factors can cause various disturbances in the ovule development, such as asymmetry, deformation, impaired ontogenesis, and partial or complete abortion of the ovule [
8,
9]. Work on the search for evaluation algorithms using neural networks opens up special prospects [
10]. In particular, in the study by Basati et al. (2018), 25 features were extracted, including 9 color features, 10 morphological features, and 6 text statistical features after preprocessing and image segmentation, to discriminate between healthy and insect-damaged grains of wheat cv. Azar using moisture levels and two-color lighting (yellow light and a combination of yellow and white light) [
11]. However, in order to develop the fundamental parameters for the evaluation and selection of valuable genotypes for breeding, it is necessary to carry out research and development without taking into account kernels that are diseased and damaged by pests by relying on the equivalence of kernels located in different parts of spikelet and spike. Therefore, the assessment of asymmetry as an indicator of the qualitative implementation of the generative development program seems to be decisive. One can assume various reasons for this effect, but the most obvious is the assumption that the seeds do not have the property of an ideal model; namely, they do not have ideal symmetry.
The reason for such an assumption is obvious. The plant itself and the spike of wheat, although they have a pronounced orderliness and at first glance can be assessed as mirror-symmetrical structures, in fact are more suitable for the description of spiral symmetry. It is especially important to understand these issues in the transition to the use of digital-assessment technologies necessary for crop forecasting and automation of the evaluation of breeding lines [
12]. In addition, the heterogeneity of kernels, which causes a decline in grain quality, especially when environmental conditions change and other developmental disorders occur, is not taken into account when assessing promising varieties and breeding lines. It seems that the mechanical properties of the covering tissues of spikes with an increase in temperature, a decrease or increase in air humidity, or a violation of turgor provoked by insufficient or excessive moisture during the formation and filling of kernels can significantly distort the expected properties and structure of the wheat yield. Evaluation of the stability of this indicator can contribute to effective breeding practices in the assessment of resistant genotypes in a changing climate with sharp changes in temperature and humidity and an increase in the number of unpredictable dry periods or flooding due to heavy rains.
It can be assumed that the structural features that cause disturbance at the level of the embryo, and subsequent manifestations of asymmetry in the development of seedlings are associated with physiological causes; in particular, with a change in hormonal regulation [
3,
13]. However, the reason may lie in the long-term selection of cereal genotypes that are “convenient” in cultivation and processing, in which the packing density and relatively stable attachment of kernels in spikelets prevented grain from spilling, which can cause a large loss in yield. Evolution in determining crop yields has moved from the ratio of seed harvested to seed sown to the modern measure of seed mass per unit area of cropland, and as a result, has favored less competitive phenotypes [
14]. Thus, we are dealing with a special rather stable structural organization of the commercial soft wheat ear, in which two lines of spikelets arranged with a shift provide uniform growth, sufficient strength and elasticity, as well as uniform distribution of nutrients from the plant to the growing embryos. Meanwhile, such an arrangement is clearly subject to the influence of many factors, including those related to the morphology of spikelets.
To test this assumption, we carried out a detailed study of the kernels, taking into account the location in the spikelets and spike, under the condition of controlling the location of the kernels relative to each other. The goal was to check the hypothesis on the unevenness of kernels and to examine the relationship between the asymmetry of kernels and their position in the spikelet and spike.
2. Materials and Methods
2.1. Plant Material
Spikes of soft wheat (Triticum aestivum L.) were the object of our research. Two spring wheat varieties (Agata and Zlata) and one winter wheat variety (Rubezhnaya) were grown in 2021 in the field plot of the Distant Hybridization Department in the N.V. Tsitsin Main Botanical Garden of the Russian Academy of Sciences. The variety Agata, var. lutescens has moderate drought and lodging tolerance. Grain of this variety is of high quality. The variety Zlata, var. lutescens has an average drought tolerance and is resistant to lodging. It is susceptible to some fungal diseases. The variety has a lower grain quality than Agata, but a higher yield. Variety Rubezhnaya, var. lutescens, has good drought and disease tolerance, and does not lodge.
In 2021, meteorological conditions were dry compared to long-term average observations, which had a corresponding effect on the grain yield and quality. The main spikes from mature wheat plants typical for the prevailing conditions in the phase of full ripeness were taken for research.
2.2. Preparing Material for Primary Analysis
Thirty spikes were randomly selected for analysis. A complex spike was disassembled into simple spikelets while retaining the order of location on the main axis. Next, the kernels were fixed with double-sided tape on a palette—a white sheet of A4 paper—with a 1 × 1 cm grid applied (
Supplementary Materials Figure S1). In order to prevent the free areas of adhesive tape on the palette from sticking to the scanner, they were treated with a thin layer of flour with an art brush. Images were scanned with a professional Epson Perfection V550 photo scanner with a resolution of 600 dpi. Image analysis and processing were carried out in the ImageJ program. The area, perimeter, values of the major and minor axes of the fit ellipse, and shape descriptors (circularity, aspect ratio, roundness, and solidity) were measured in 505 kernels according to the program instructions [
15]. To avoid image-processing distortions in ImageJ, it was necessary to place the kernel strictly in the middle of the square cell, so that even the shadow of the kernel did not fall on the grid lines (
Supplementary Materials Figure S2).
2.3. Preparation and Measuring of Kernel Sections
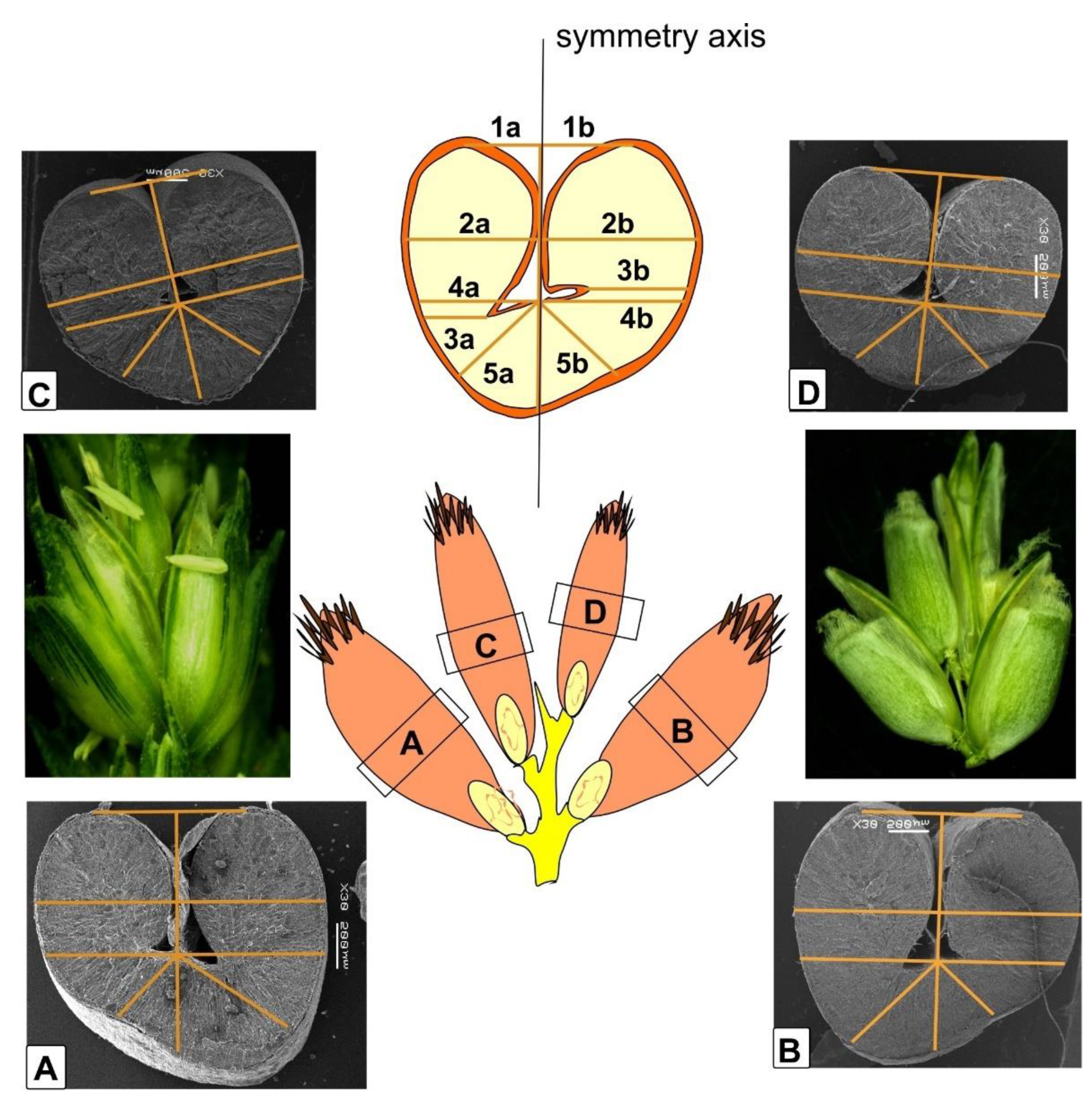
A total of 278 sections were prepared, of which 93 were from the far left kernels, 93 were from the far right kernels, and 93 sections were from the central kernels. Sections were taken from the middle part of kernel. To make give the light sections a higher contrast with the background, they were fixed on a matte black modeling board using Oasis Fix floral plasticine. Image was scanned with Epson Perfection V550 scanner at 600 dpi. Using the ImageJ program, the lengths of the central axis of kernel cut and the distance from the control points of the left and right sides of kernel to the axis were measured manually to establish the asymmetry indices. Index 1 was the ratio of the distance from the top of kernel to the central axis to the total distance between the tops. Index 2 was the ratio of the distance along the widest kernel part to the central axis to the total width. Index 3 was the ratio of the distance from the edge of kernel under the triangle to the central axis to the total distance. Index 4 was the ratio of a distance from the kernel edge along the line under the triangle to the corner of the triangle. Index 5 was the ratio of each segment that extended 45° from the axis of symmetry to the segment of length from the triangle to the bottom (
Figure 1;
Supplementary Materials Figures S3–S5).
2.4. Kernel Sample Preparation for Scanning Microscopy
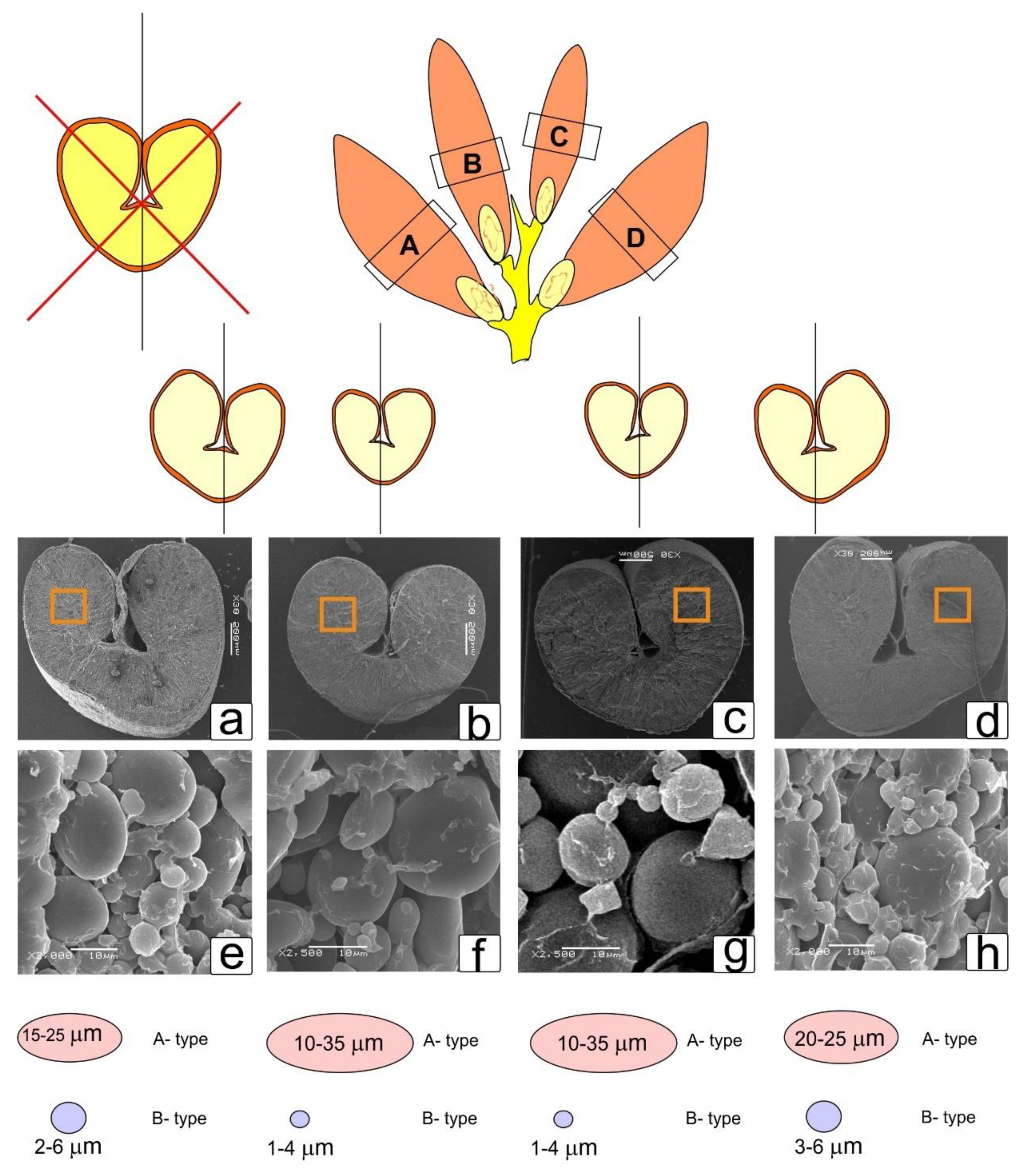
Kernels from the 5th and 6th spikelet located upwards from the base of the largest spikes of the Rubezhnaya variety were used for analysis. The choice of this zone and large spikes was due to the fact that in this zone, there were always four formed kernels (two upper and two lower) in the spikelet, two of which were located on the right and two on the left. Kernels from three spikes were analyzed using eight kernels from each spike. Kernels (having a moisture content of 14–15%) were placed crease (groove or ventral furrow) down on the cork surface. After that, a transverse cut was made at a distance of 2.5–3 mm from the embryo, pressing hard until the blade entered the cork. Next, another transverse cut at a distance of approximately 3 mm was made. Thus, the central part was cut out. To avoid errors in determining the analyzed kernel side, a longitudinal incision was made on the reverse side (closer to the embryo), visible on the section.
The samples of the kernel core were fixed in a 2.5% glutaraldehyde in 0.1 M Sorenson buffer at pH 7.2 and washed in buffer. The samples were dehydrated through an ethanol series (30% 30 min, 50% 30 min, 70% 30 min, 96% 30 min, 2 × 100% 30 min). Then, CO2 was added to the sample to be critical-point-dried with a Hitachi HCP-2 critical point dryer (Hitachi, Tokyo, Japan). Then, the dry seeds were mounted on a SEM stub and coated with gold and palladium using an Eiko IB-3 ion-coater (Eiko, Tokyo, Japan); the thickness of the Au layer was 20 nm. The slices were then observed under a Camscan-S2 SEM (Cambridge Instruments, Cambridge, UK) in the Laboratory of Electron Microscopy (Biological Faculty of Lomonosov Moscow State University). Magnifications from 30× to 2500× were used. The samples were determined using the SEM Control User Interface Version 7.11, copyright 2004, by JEOL Technics LTD. For analysis, photographs of at least 10 shots in each kernel were used. Selected photographs and data on a sample made according to the scheme shown in the corresponding image are given. Other results obtained during the study of these images are not presented in this work.
2.5. Statistical Analysis
All statistical calculations were carried out using the IBM SPSS Statistics 25 program. Compliance with the normal distribution was established by the Kolmogorov–Smirnov test. Differences were assessed, and spikelets typical of the variety were identified by multiple pairwise comparisons in ANOVA according to a Scheffe test (
p = 0.05). The variability of kernels was assessed on a scale on which coefficients of variation from 1 to 15% corresponded to a very low level of polymorphism, 16–25% to an average level, 26–35% to an increased level, 36–50% to a high level, and more than 50% to a very high level [
16]. To compare the identities of the left and right kernels in terms of area, perimeter, ellipse axes, and shape descriptors, the correspondence to a normal distribution of features was first established, and then the Student’s t-test for independent samples was conducted. A symmetry comparison of the left and right sides of the kernel was carried out in a similar way. To compare the mechanical influence of kernels on each other’s asymmetries, the value of relative asymmetry was determined as the ratio of the module of the difference in the length of the left and right sides of kernel to the length sum of the left and right sides of the kernel section. The overall fluctuating asymmetry index was calculated as the mean of the relative asymmetries of the kernels [
17]. Pairwise comparisons were made using the Kruskal–Wallis one-way analysis of variance (
p = 0.05). To develop an asymmetry scale, a ranking and breakdown into 5 classes of relative asymmetry values for 278 sections of kernels was carried out. Next, the location of the largest number of kernels with low asymmetry was clarified.
3. Results
Ears and spikelets of spring wheat can differ significantly, which can cause the formation of kernels of different sizes and shapes, depending on the full development of flowers in the spikelet, and the location of the kernels in the spikelet (
Figure 2). For this reason, we chose three varieties that differed in the parameter of the maximum number of seeds per spikelet: Agata had three grains (kernels) per spikelet (
Supplementary Materials Figure S3), Rubezhnaya had four grains per spikelet (
Supplementary Materials Figure S4), and Zlata had five grains per spikelet (
Supplementary Materials Figure S5). To study all the parameters, the Rubezhnaya variety was chosen as a model.
3.1. Assessment of Uniformity of Simple Spikelets According to the Average Parameters of Kernels in a Complex Spike
In a complex spike, the Rubezhnaya variety had an average of 19 simple spikelets, each of which included from one to four kernels, while the Zlata variety had five kernels maximum, and the Agata variety had three kernels in the spikelet. A total of 425 kernels were analyzed. The largest number of kernels (four pieces each) was found in spikelet numbers 4, 5, 6, 7, 8, 9, 10, and 11; while 1–2 kernels were more common in spikelet numbers 1, 14, 15, 16, 17, 18, and 19. Using ANOVA, it was possible to establish the presence of significant differences in the kernel area between spikelet number 1 and a group including spikelets 4, 5, 6, and 7, on average, by 4.8–5.4 mm2; spikelets 4, 5, 6, and 7 also differed from 15 and 18 (larger than the latter by 4.5–5.1 mm2); and spikelets 8 and 9 differed from spikelet 18. Significant pairwise differences in the group average values of the perimeter of kernels were not found. The kernels in spikelet numbers 1 and 18 had a significantly shorter length of the ellipse major axis than the kernels in spikelets 4, 5, 6, 7, 8, 9, and 10, on average, by 0.79–0.99 mm. All simple spikelets did not have significant differences in the average width (smaller axis of the ellipse) of kernels, except for spikelets 18 and 5 (the average width of the kernels in spikelet 5 was 0.79 mm larger). All this led to establishing that spikelet numbers 2 to 15 were typical for the variety, because they did not significantly differ from each other in the average values of the parameters studied.
Variation in the section area of kernels in spikelets was at a very low (up to 9–15%) or low level (16–18% in spikelets 11, 15, and 18) (
Figure 2 and
Figure 3). In addition, the bulk of the kernels had a very high evenness along their average perimeter of (variability is 9–14%), and the variability was at a level of 16–18% in spikelets 8, 9, 11, and 16. The Evenness of the kernels was very high (coefficients of variation were 5–13%) in length (major axis of the ellipse) and width (minor axis of the ellipse).
3.2. Assessment of Uniformity of Kernels in Simple Spikelets
In the distribution of the section area, the perimeter and the major and minor axes of the kernel ellipse in simple spikelets obeyed the law of normal distribution. Multiple pairwise comparisons established a significant effect of the kernel location in the spikelet on the cut area, perimeter, and minor axis. The far left and far right kernels did not differ from each other in these parameters, nor did the middle left and middle right kernels. However, the furthest outside kernels were significantly different from the middle kernels along the smaller axis. The far left kernel and the far right kernel were reliably identical with the middle right kernel in area and perimeter, but differed from the middle left kernel. A detailed comparison of the form descriptors of the far left and far right kernels using the t-test method showed that the angle between the major and minor axes of the kernel ellipse was the same, as well as parameters of the aspect ratio and roundness. However, there were significant differences in the circularity and solidity. The far right kernel was 0.027 mm more rounded than the far left one, and it also had 0.011 units more solidity.
To identify the causes of asymmetry in the circularity and solidity, the cuts of kernels were further considered.
3.3. Analysis of the Section Symmetry of the Kernel Left and Right Sides
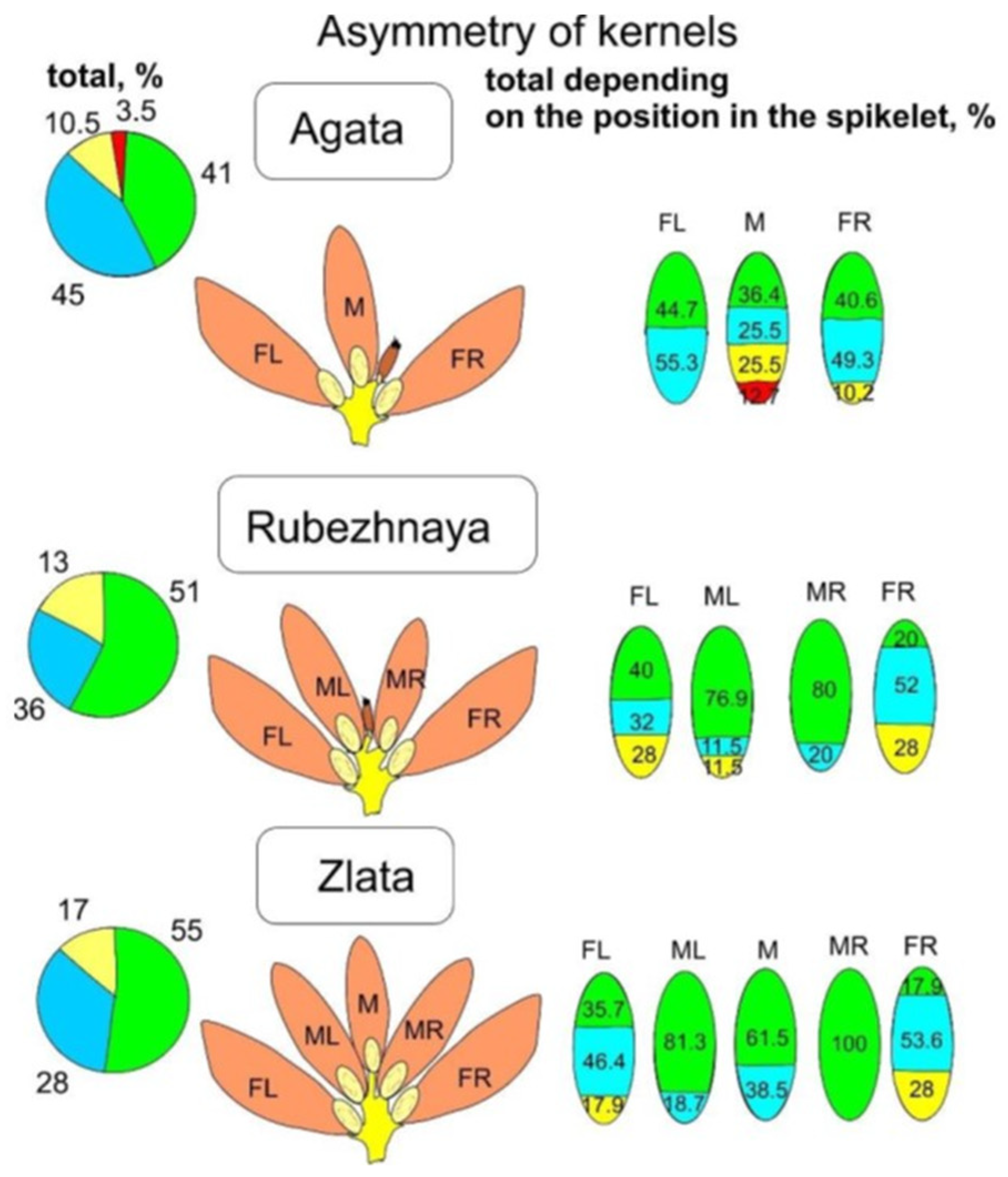
Comparison of the left and right sides of the section of the far left kernel according to five indices in three varieties rejected the hypotheses on the symmetry of the cut sides. The same asymmetry was observed in the far right kernel. However, mirror symmetry was found in the extreme grains. So, the left side of left kernel was a mirror symmetry of the right side of right kernel, and the right side of left kernel was mirrored to the left side of right kernel. The left kernel of the Rubezhnaya variety had a significant symmetry according to index 1 (the significance of the criterion was 0.353, with a threshold value of 0.05); the right kernel had an index of 5, but the excess of the threshold significance of 0.05 was only 0.13. The left kernel of the Agata variety had an asymmetry according to index 5 (significance of the criterion of 0.001), and the right kernel had an asymmetry according to index 1 (significance of 0.001). The variety Zlata had an asymmetry with the left kernel according to indices 3 (significance of 0.045) and 5 (0.027), and the right one according to the indices 1 and 2 (0.001).
In internal kernels, there was more symmetry in the structure of kernel. So, a significant symmetry of all indices was found in the middle left kernel, while the middle right kernel has symmetry in four indices, except for index 1. When assessing the mirroring of the left side of left kernel and the right side of right kernel, there was no asymmetry, as well as when establishing the same trait for the right side of left kernel and the left side of right kernel (
Table 1).
Since the proportion of asymmetric indices was significant, it was necessary to evaluate an influence of the mechanical action of kernels on each other.
3.4. Evaluation of the Influence of Mechanical Action on the Asymmetry of Indices of Kernel Section
The indicator of relative asymmetry 1 regarding the deviation of the upper points of cuts from the symmetry axis was the same for all kernels; i.e., for those where there were four kernels in a simple spikelet, and where a single kernel was freely formed in the spikelet. The same results were obtained in the deviation of the lengths of sections in the widest part (relative asymmetry 2). The indicator of relative asymmetry 3, showing the distance ratio under the middle triangle, demonstrated the differences between the left kernel and the middle kernel in spikelets with three kernels. Indicator of relative asymmetry 5 regarding the ratio of lengths of the diagonals extending at an angle of 45° from the base of the middle triangle of the kernel to the bottom of the kernel indicated that the middle kernel in a spikelet with three kernels differed from the far left and far right kernels in a spikelet with four kernels, from left and right kernels in a spikelet with three kernels, and from the left kernel in a spikelet with two kernels.
When considering the general asymmetry of the kernels, significant differences were found between the middle kernel in a spike of three kernels and the leftmost and rightmost kernel in a spikelet of four kernels. In general, this indicator showed that the asymmetry of the outside kernels in spikelets with 2, 3, or 4 kernels was statistically the same (
Table 2). In kernels in the middle of spike, the symmetry was higher.
3.5. Scale of Indicators of General Asymmetry
In a sample of 278 slices, the minimum value of relative asymmetry was 0.01, and the maximum range of variation reached 0.184. A breakdown of the range into five classes led to the following results (
Table 3).
In the Agata variety, low or very low degrees of relative asymmetry were noted in all left kernels. Among the right kernels, 10% of the samples had a middle degree of asymmetry, and in the middle kernels, a middle or very high degree of asymmetry was rare, and most were low or very low. The Zlata variety had a maximum of five kernels per spikelet, and the middle left and middle right kernels had a very low degree of asymmetry, while the central (upper) one had both a very low degree (in 59% of middle kernels) and a middle degree of asymmetry (in 41% of kernels). In the left and right kernels of the Zlata variety, a low or very low degree of asymmetry also prevailed; the average degree was found in 18% of the sample of left kernels and in 27% of the entire sample of right kernels. On the contrary, in the Rubezhnaya variety, a greater amount of low or very low variation was concentrated in the samples of the middle left and middle right kernels (up to 95% within these samples). Among the far left and far right kernels, there were up to 30% of kernels with a middle degree of variation (
Figure 4).
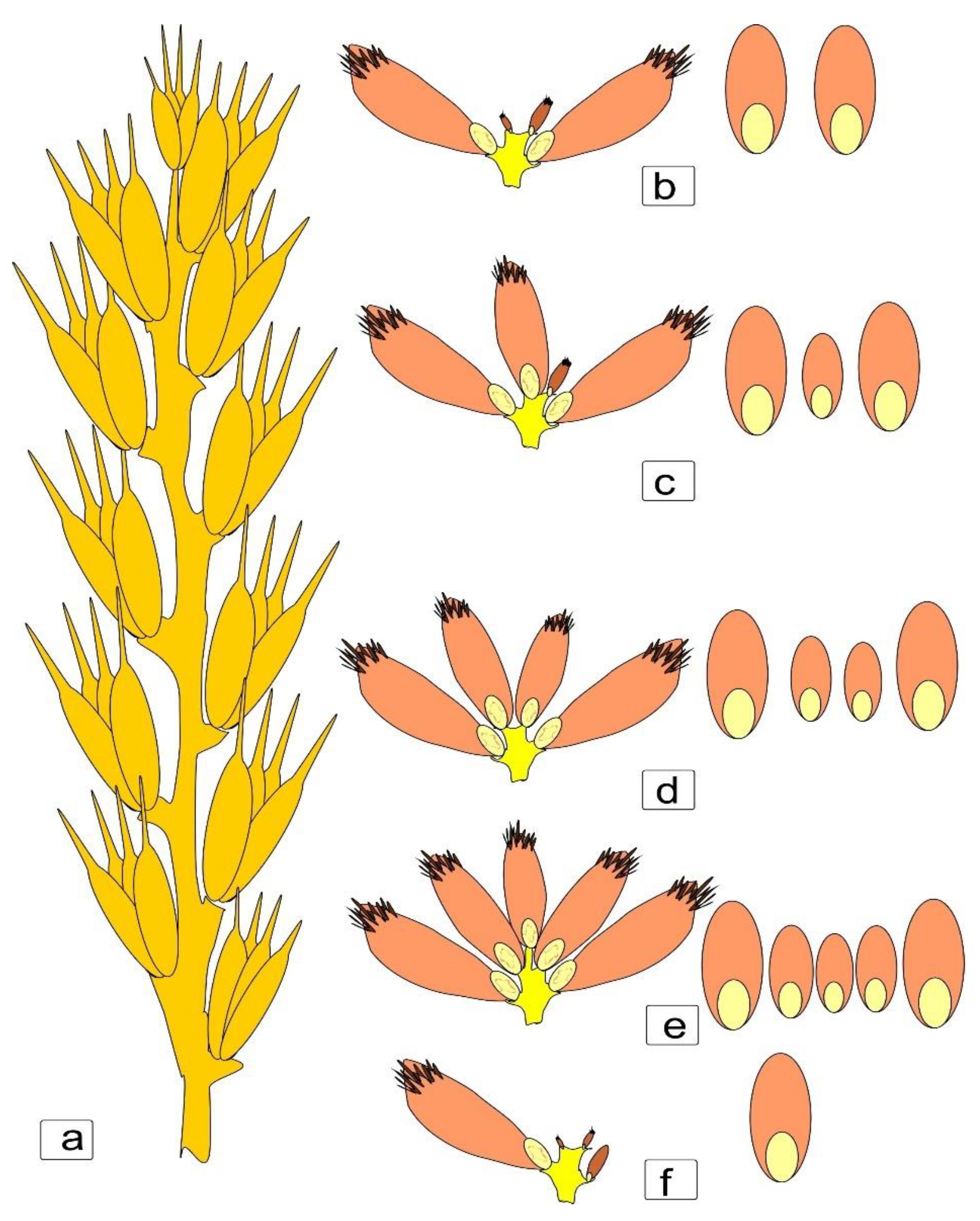
To understand the structural morphology of the formation of asymmetry in kernels, it seemed important to us to assess the aspect of shape change at an earlier stage. Here, we present kernels from the central part of the spike that had completed growth and were moving on to maturation (
Figure 5). Here it can be seen that when transferring the outline of the spikelet facing the central part, uneven development and a change in the shape of the kernel and crease were revealed (
Figure 5a,b,e,f), and at this stage, it already was noticeable that the right and left parts of the section of the kernel differed in the lower kernels of the larger ones in the upper kernels (
Figure 5c,d) to a lesser extent (
Figure 5g,h).
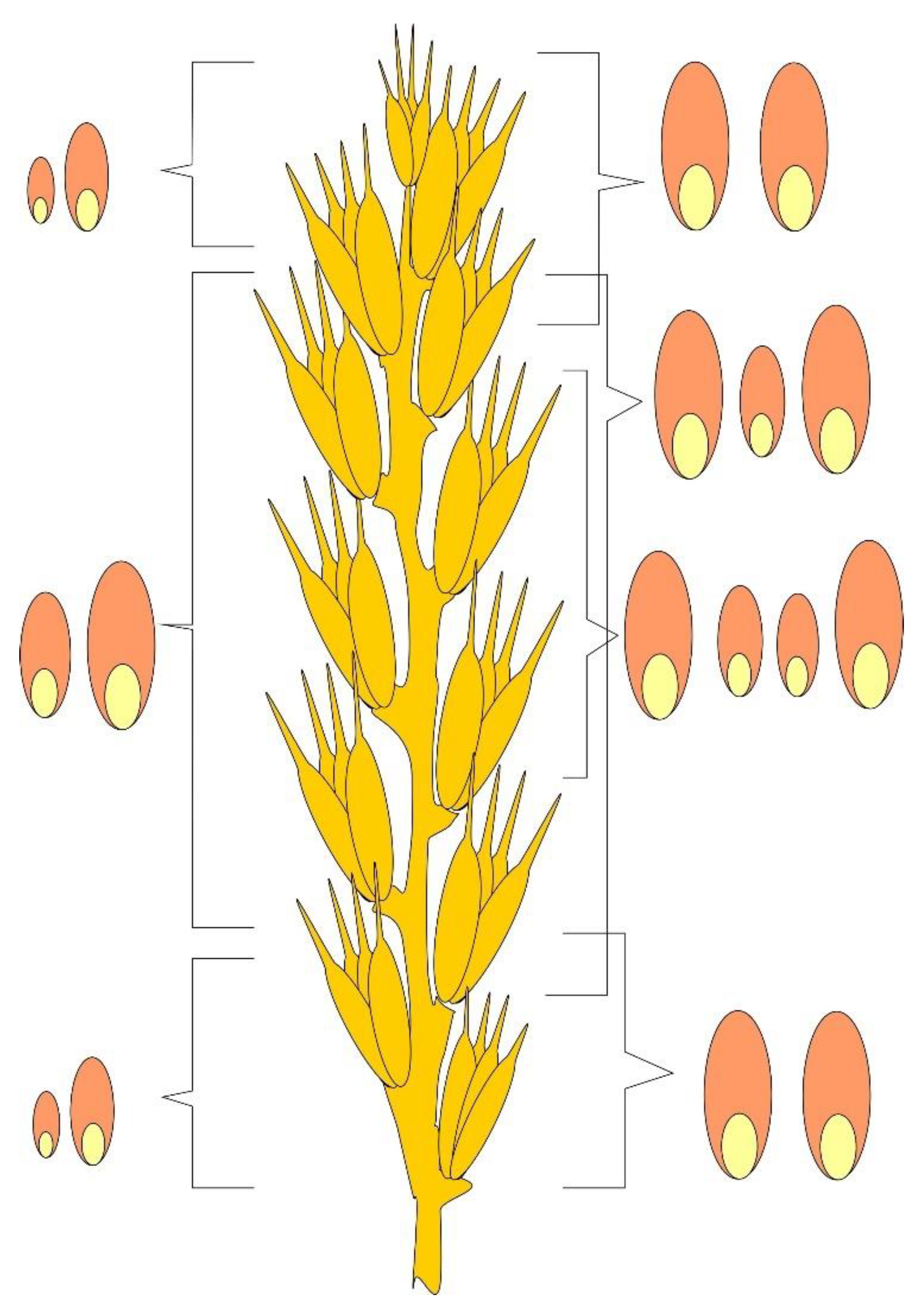
Examples of the formation of a spikelet and kernels with a different number of emerging kernels are shown in
Supplementary Materials Figure S6. In the horizontal rows, there are images of spikelets forming three (
Figure 6a–c), two (
Figure 6d–f), and one (
Figure 6g–i) kernels. The lower row shows underdeveloped spikelets in the upper and lower parts of the ear formed on a weakened plant and not forming anthers and ovaries (
Figure 6j–m).
3.6. Visual Assessment of Asymmetry in Starch Filling of the Left and Right Sides of Kernel Section
When studying cross sections of the central part of the kernels under a scanning microscope, the pronounced asymmetry effects that were characteristic of those located in the right and left parts of the spikelet were preserved (
Figure 6).
This was especially clearly reflected in the uneven formation of the part facing the center of the spike and the part facing outward, which was more pronounced in the lower kernels of the spikelet (a,d) compared to the upper kernels (c,d) and the asymmetry of the crease zone. It also was noted that there were significant differences in the starch grains of the lower (e,h) and upper kernels (f,g), which probably could affect the quality and properties of flour obtained from large lower and upper finer kernels. Therefore, an increase in the percentage of protein and a change in the properties of starch in smaller kernels could be expected.
4. Discussion
The important role of assessing the asymmetry of wheat grains was first proposed in 1918 [
18]. At present, it is obvious that a number of parameters that determine the predictability of the symmetry of the kernel sides and the equivalence of the quality of kernels, which are so valuable for humans, can be genetically determined without a significant spread between the upper/lower, right/left, and central ones. In the work of Boshnakian, as assumptions, it was suggested that the uneven development of kernels, which is undesirable, causes excessive rigidity of the protective coat (skin), the shape of the cavity in which the embryo develops, and the features of the attachment of caryopses to the spikelet and spikelets to the spike [
18]. Thus, as an explanation for the phenomenon of asymmetry, various physical conditions were proposed that were formed during the development of generative organs, which became the causes of purely mechanical restrictions that caused shape deformation and prevented the symmetrical development of kernels [
19]. It can be assumed that the analyzed seed accessions that had not yet passed the “filter” of the comprehended selection and the green revolution had a much more pronounced asymmetry, since it is known that other unfavorable and more noticeable consequences of uneven development were still quite often noted for wheat in those years, such as the germination of kernels in a spike [
20] or premature shedding of grain [
21]. Currently, approaches are being considered that imply the use of marker and phenotypic selection with computer modeling [
22]. The authors discussed the relationship between rainfall, grain wetting, and germination, and explored the nature of physiological and morphological controls over wetting and sprouting of the spike [
23]. It was argued that the scatter in the wettability of spikes in different varieties indicated acceptable genotypic differences that were suitable for breeding, even in terms of the properties of separate grains [
24]. In addition, to estimate the quality of valuable breeding lines and varieties in relation to the analysis of genetic characteristics, it was proposed to use the shape of the grain and its size as important phenotypic traits [
25]. Such properties have been found to be suitable for models for predicting grain quality [
26,
27]. This may acquire particular importance in connection with the modulation of these traits in varieties and breeding lines in a changing climate and an increase in the degree of damage from abiotic stress factors. So, it was shown that the quality of wheat grain depended on weather conditions and properties of the variety [
28].
The varietal differences we established are significant, and indicate the need for further research using phenotypic analysis to accompany the modern breeding process. Therefore, the proposed system for analyzing the totality of fluctuations in the cereal spike and individual kernel becomes necessary.
The initiation and formation of spikelets in wheat spikes begins after the transformation of the apical meristem, and is accompanied by changes when entering the tube up to the beginning of flowering. Thus, the laying of ovules and the structuring of spikelets and the spike as a whole occur at the stage when the plant is in the tillering [
29,
30]. Thus, a genetically determined potential of productivity is formed long before the structures of the male and female gametophyte are finally formed [
31]. However, there may be a number of key points that may cause a change in the most productive process that provides emergence of a new generation. So, damages can affect the very system of formation of both spikelets and flowers, and cause violations of the development of the ovule and stamens [
32]. In particular, changes in the morphology or disturbances in the formation of both the generative organs themselves and the generative cells (pollen and eggs) can occur. These processes are reflected primarily in the number of spikelets in the spike and the number of flowers in the spikelet, as well as in various types of violations of the processes of fertilization and embryo development [
33]. Disturbances in normal development can also affect the manifestation of the external parameters of the skin of wheat grains [
34,
35]. In our studies, such a picture was typical for examples for which a decrease in the maximum productivity of spikelets was observed (
Figure 3;
Supplementary Materials Figure S6). So, for wheat varieties, these indicators were different, and spikelets could produce from zero to five kernels (
Table 2;
Figure 2;
Supplementary Materials Figures S3–S6). For example, the variety Zlata could demonstrate high generative efficiency, expressed in the generation of up to five kernels per spike, in contrast to other varieties, for which the maximum number was four (
Supplementary Materials Figure S5). Meanwhile, it was obvious that under any adverse conditions, this maximum number would be reduced [
36]. The probability of the absence of fertilization or the arrest of the development of embryos was probably quite high, and could be observed under normal conditions of development, which could clearly be observed for the lower and upper spikelets in the spike, the maximum generative potential of which was equal to one or two kernels, and in a number of unfavorable cases, with underdeveloped flowers, it was absent (
Figure 3;
Supplementary Materials Figure S6).
Attempts to study the fluctuating asymmetry in wheat were carried out only on polycrystalline structures isolated from wheat [
17]. No studies have been devoted to the examination of fluctuating asymmetry of kernels; however, there are works on the effects of seed galls on
Inga laurina [
37] and the effects of salinity and light levels on asymmetry in
Sinapis arvensis (
Brassicaceae) [
38], as well as on fluctuating asymmetry in the population size of
Aquilegia thalictrifolia [
39]. In our study, the number of kernels per spike acted as a stress factor. The results of our study were consistent with the above-mentioned results: with an increase in the stress factor, the level of fluctuating (general) asymmetry increased. So, the values of fluctuating asymmetry in the far left and far right kernels were higher in spikelets of four kernels than in spikelets of two grains (
Table 2). The kernels that were closer to the top of the spikelet experienced the least stress (
Table 2), and the indicator of fluctuating (general) asymmetry was lower.
It was revealed that the spikelets in a complex spike were not symmetrical; the most typical part of the spike lay in the interval from the second to the 15th spikelet. The variability of kernels was at a low or very low level; i.e., the kernels of a complex spike were aligned. The kernels did not differ in cut area, perimeter, or length of the major and minor axes of the ellipse. However, on the cut of kernels, a pronounced asymmetry of the left and right sides was visible. The far left and far right kernels were mirror-symmetrical. The general asymmetry of the furthest outside kernels was 2 times higher than that of the middle kernels. The number of kernels in the spike did not affect the overall asymmetry of the outside kernels.
The aim of our study was the evaluation of the asymmetry expressiveness in wheat kernels in relation to the previously identified asymmetry in the development of seedlings [
3]. The assumption regarding the presence of asymmetry in the kernels and the relationship of this fact with the kernel position in the spike was confirmed for all varieties of soft wheat studied (
Table 2). Meanwhile, it was obvious that the expressiveness degree of this effect depended on the phenotypic and morphological features of the variety, which was especially noticeable for the Zlata variety (
Figure 4). On the other hand, the regularity of the existence of asymmetry between the right and left parts of the kernel, as well as the presence of differences in the composition and structure of starch grains between the lower and upper kernels, allowed us to assess the potential of these differences for practical application, since these kernels had a smaller weight and size, and could be separated from the larger kernels of the lower tier. In addition, if it is possible to identify among wheat varieties those in which these differences are minimal, the quality of the harvest and the properties of grain and flour resulting from it will become more predictable.
The revealed facts of asymmetry, mirror symmetry, and differences in the asymmetry between kernels of different origins depending on both the position in the spikelet and the position of the spikelet on the spike raised the issue of whether these factors should be taken into account when setting up fine experiments, in which wheat kernels are used as a habitual ideal model [
26,
40]. The wheat kernel is considered as a clear mathematical model in applied evaluation systems. The proposed model uses simple geometric data: length, width, and thickness and depth of the crease, but ignores both unevenness and asymmetry [
41]. In order to suggest how exactly this can affect the development of the seedling, it is necessary to take into account that the imbibition and swelling of seeds with different endosperm starch composition can occur in different ways [
23]. Both the activation of enzymes and the transport of nutrients through the scutellum, which are necessary for the growth of the embryo and the activation of meristems of the main and adventitious roots and apical meristem of the shoot, can be changed, especially under stress [
42]. Thus, when working with such a familiar and idealized model as a grain of wheat, it is better to take into account its lineage and/or use seeds taken from a spike of variety with control of their origin.


 ,
, 









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}