
Model Membrane Systems Used to Study Plasma Membrane Lipid Asymmetry

 ,
,  , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Techniques Used to Prepare Asymmetric Membranes In Vitro
2.1. Asymmetric Large Unilamellar Vesicles
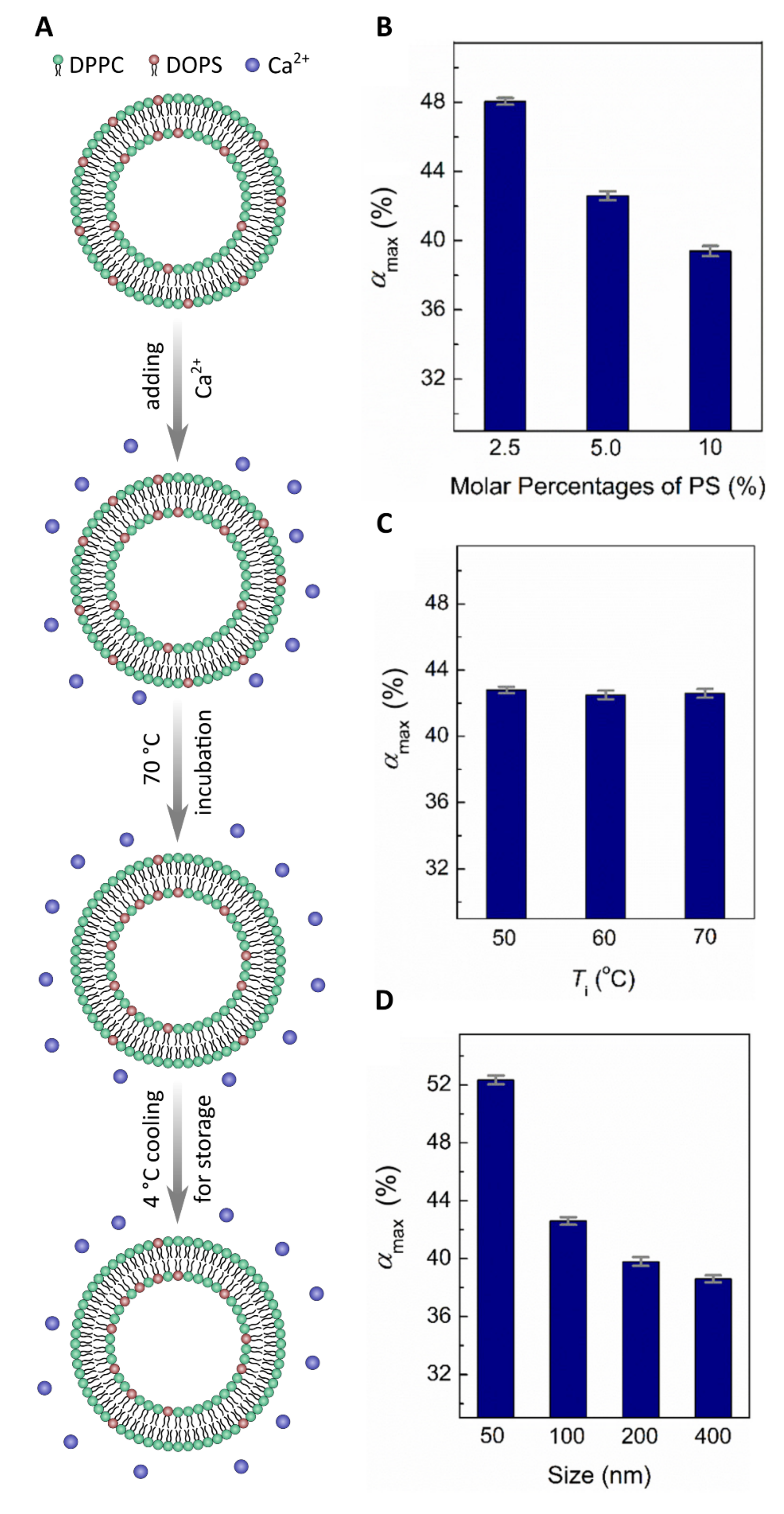
2.1.1. Ca2+ Ions Induce Asymmetry of PS Lipids in Membranes
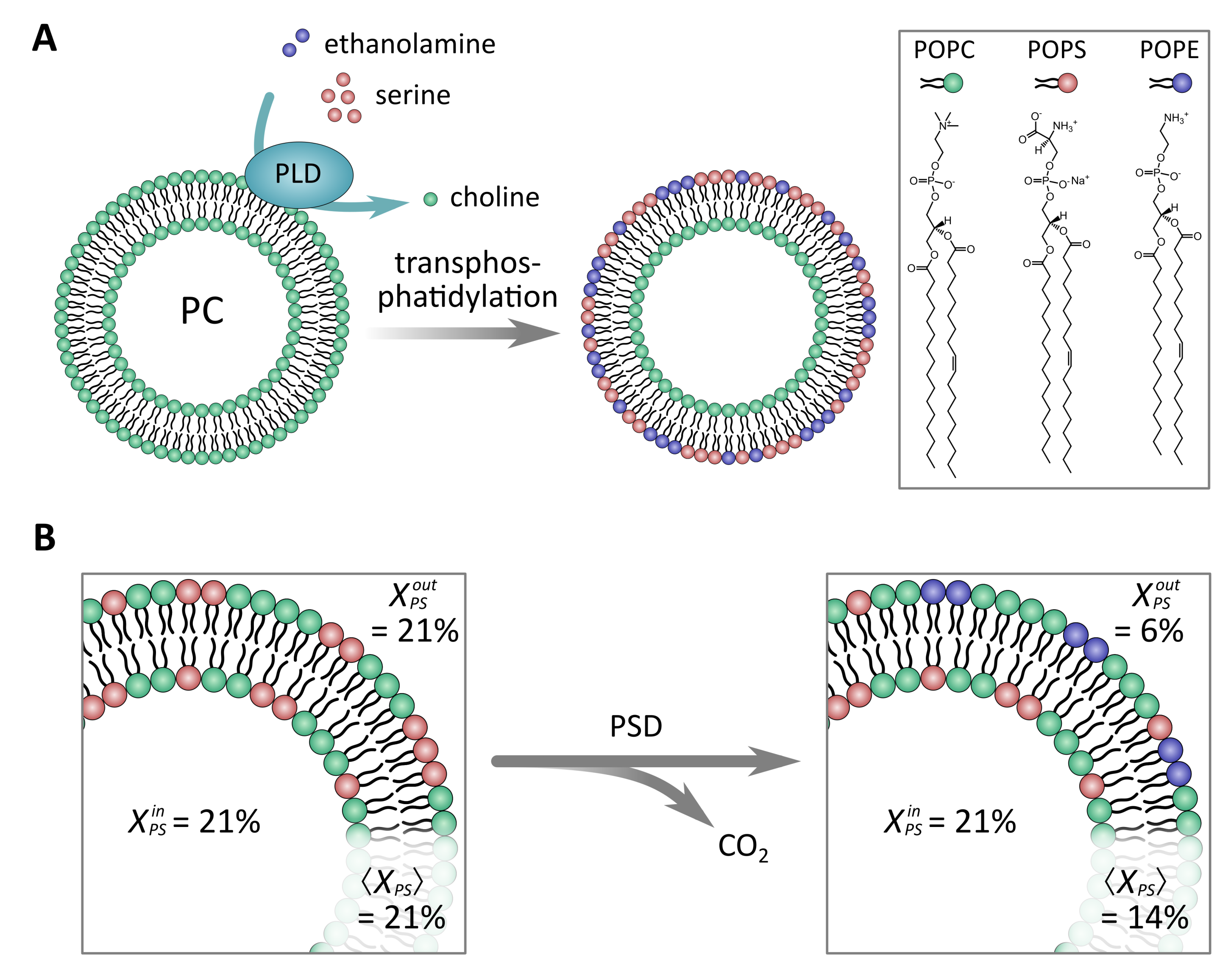
2.1.2. Enzymes as Tools to Generate Asymmetric Vesicles
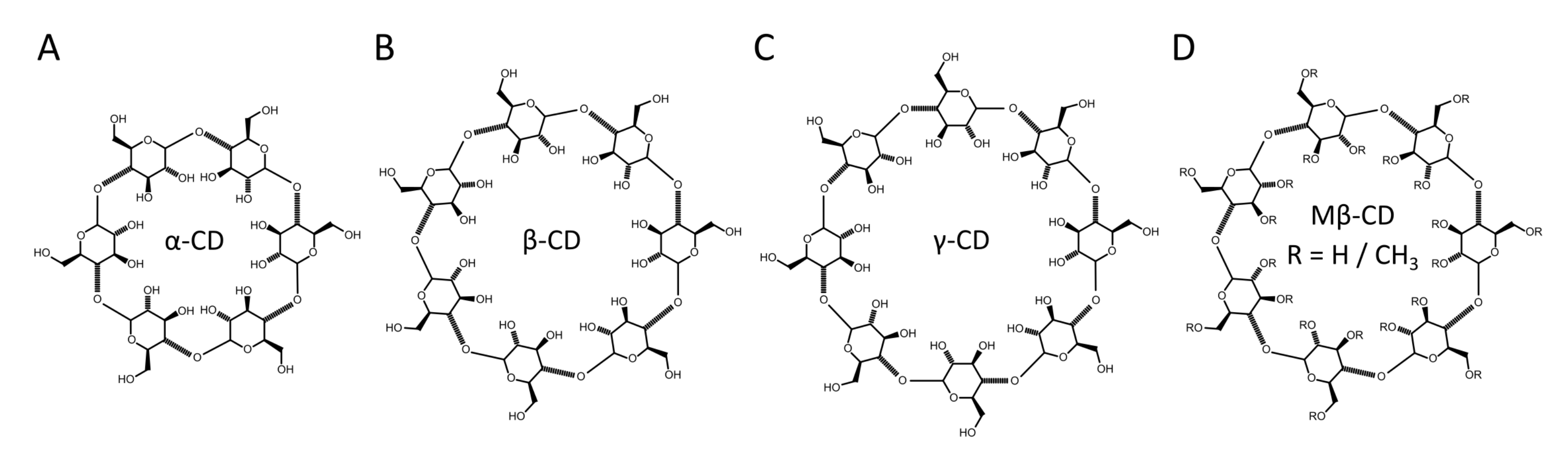
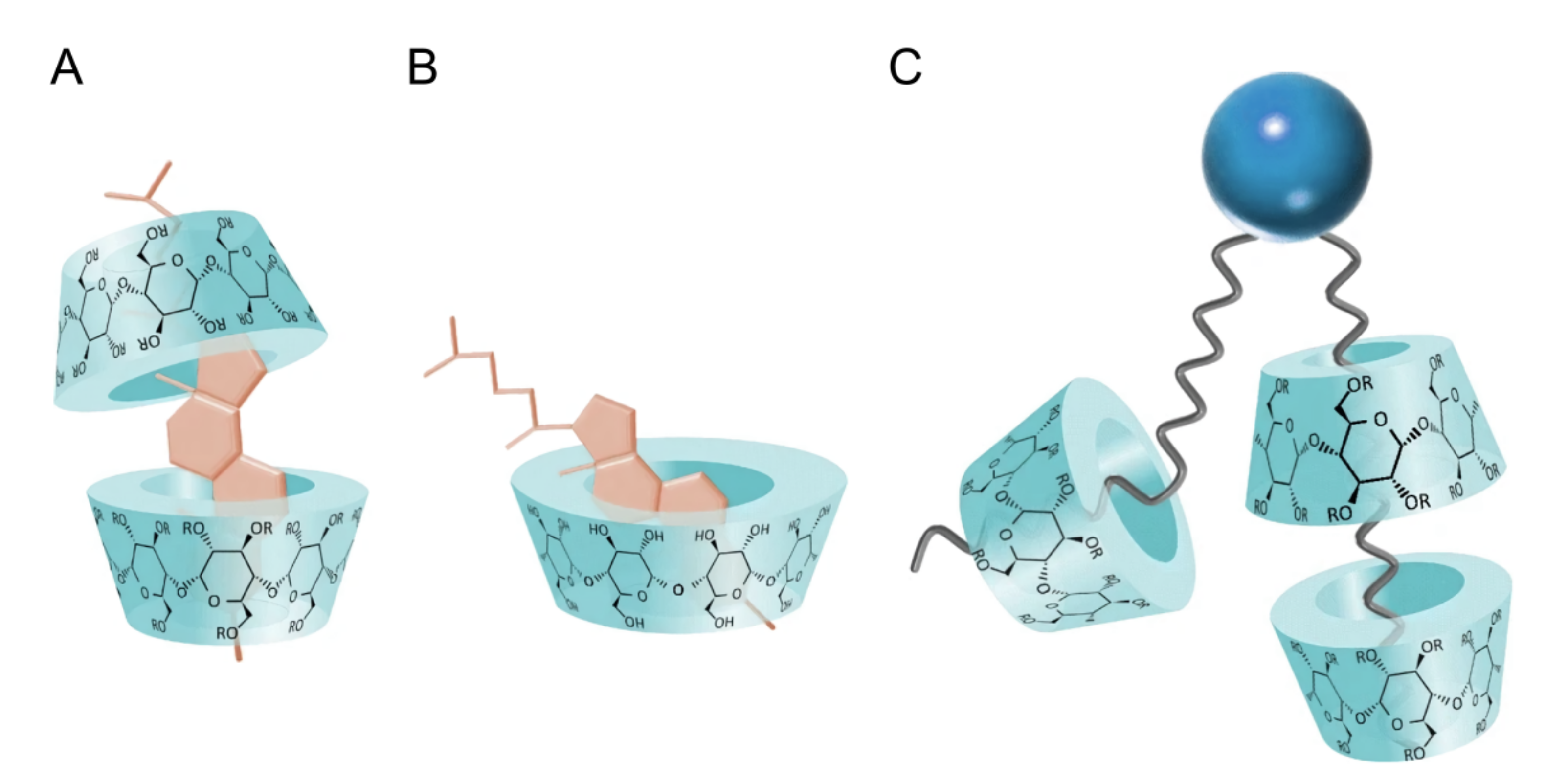
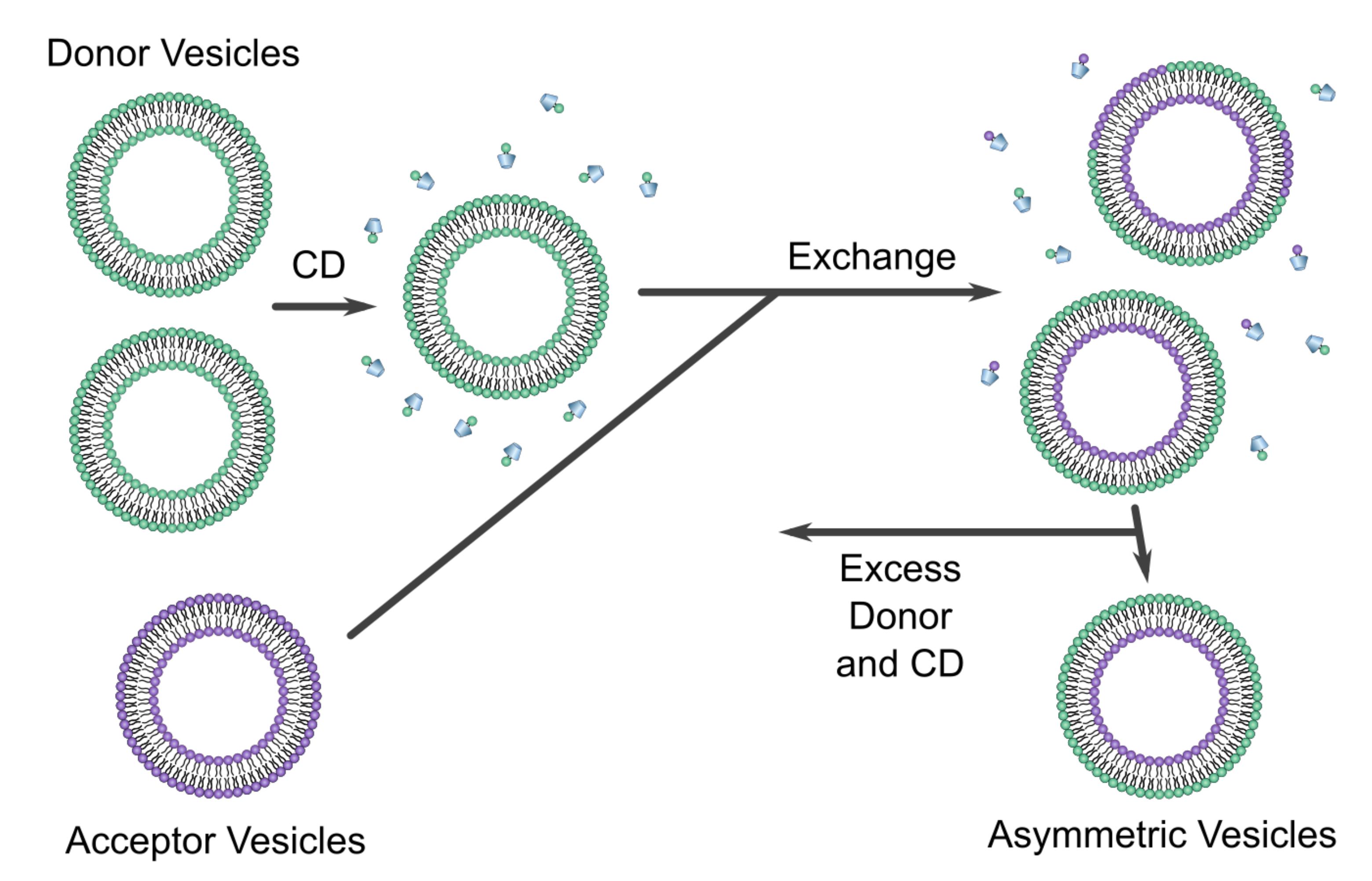
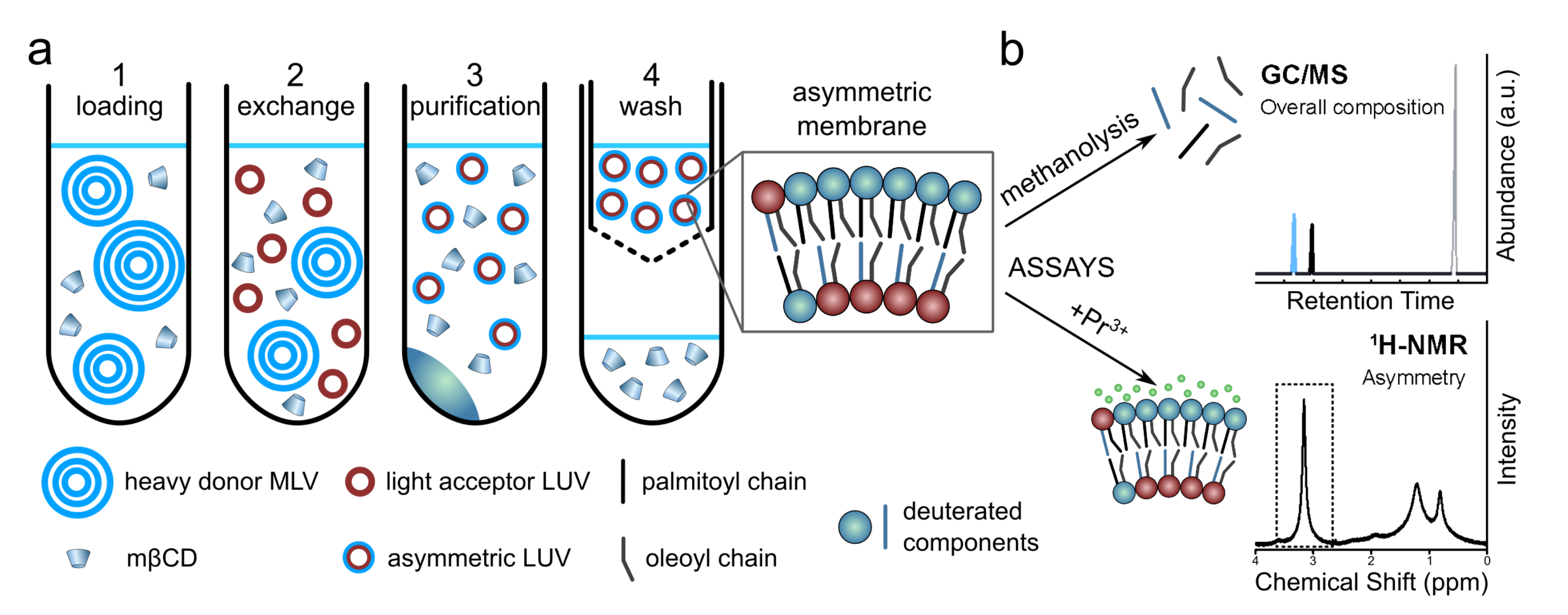
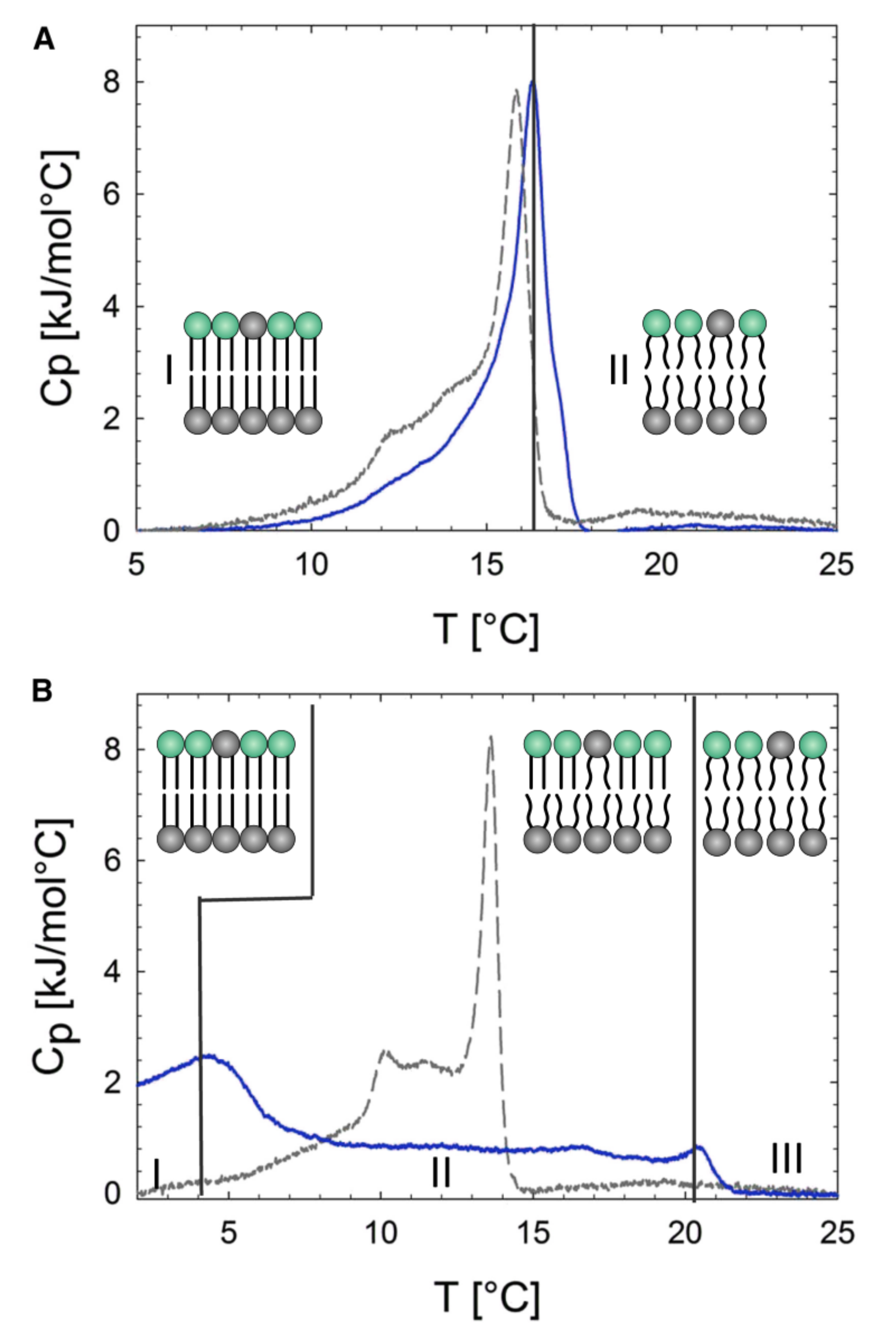
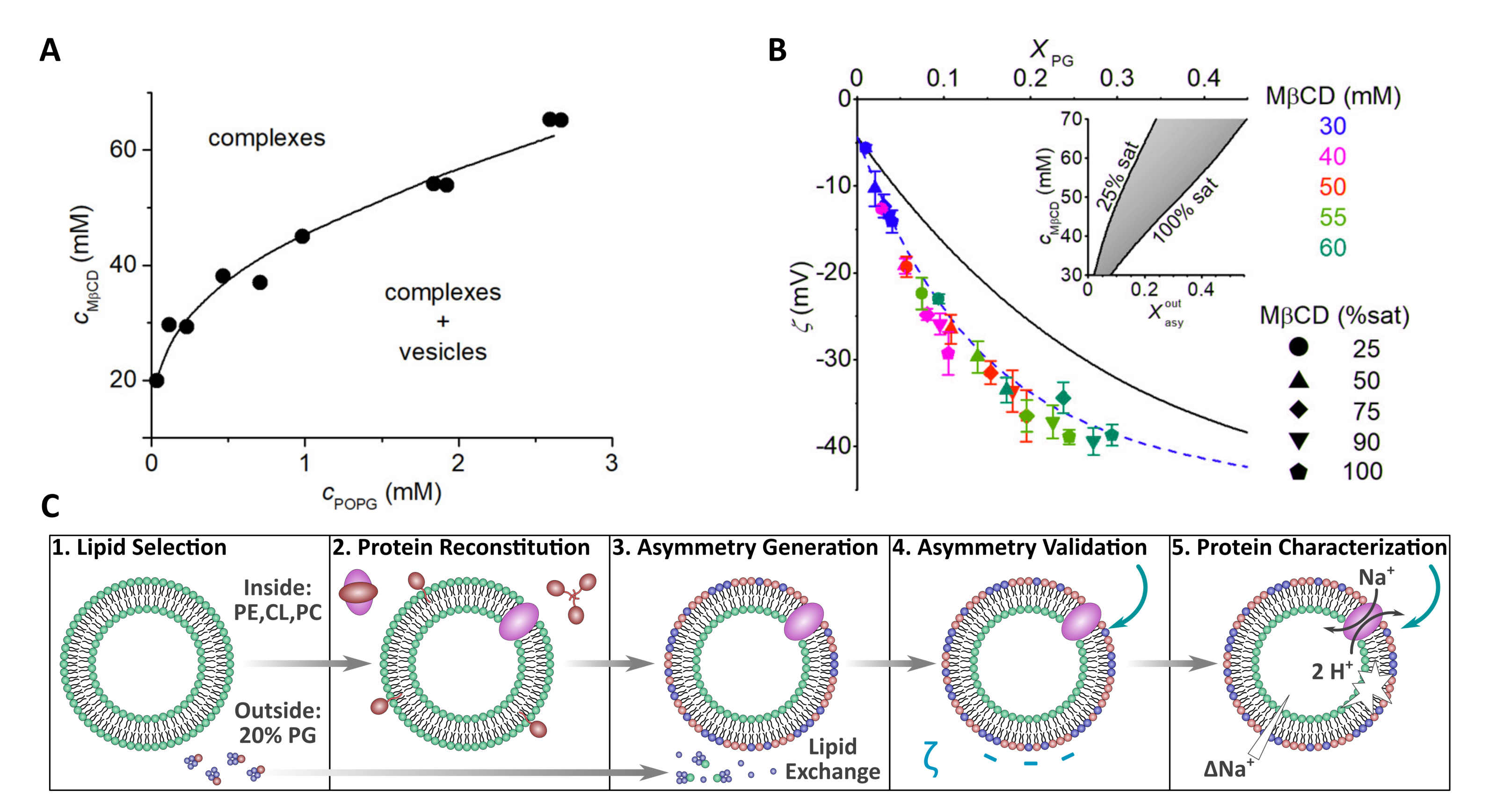
2.1.3. Using Cyclodextrins to Generate Asymmetric Vesicles
2.2. Giant Unilamellar Vesicles and Membrane Asymmetry
2.2.1. Preparation of aGUVs by MCD-Mediated Lipid Exchange
2.2.2. Cholesterol-Rich aGUVs Made Using MCD-Mediated Lipid Exchange
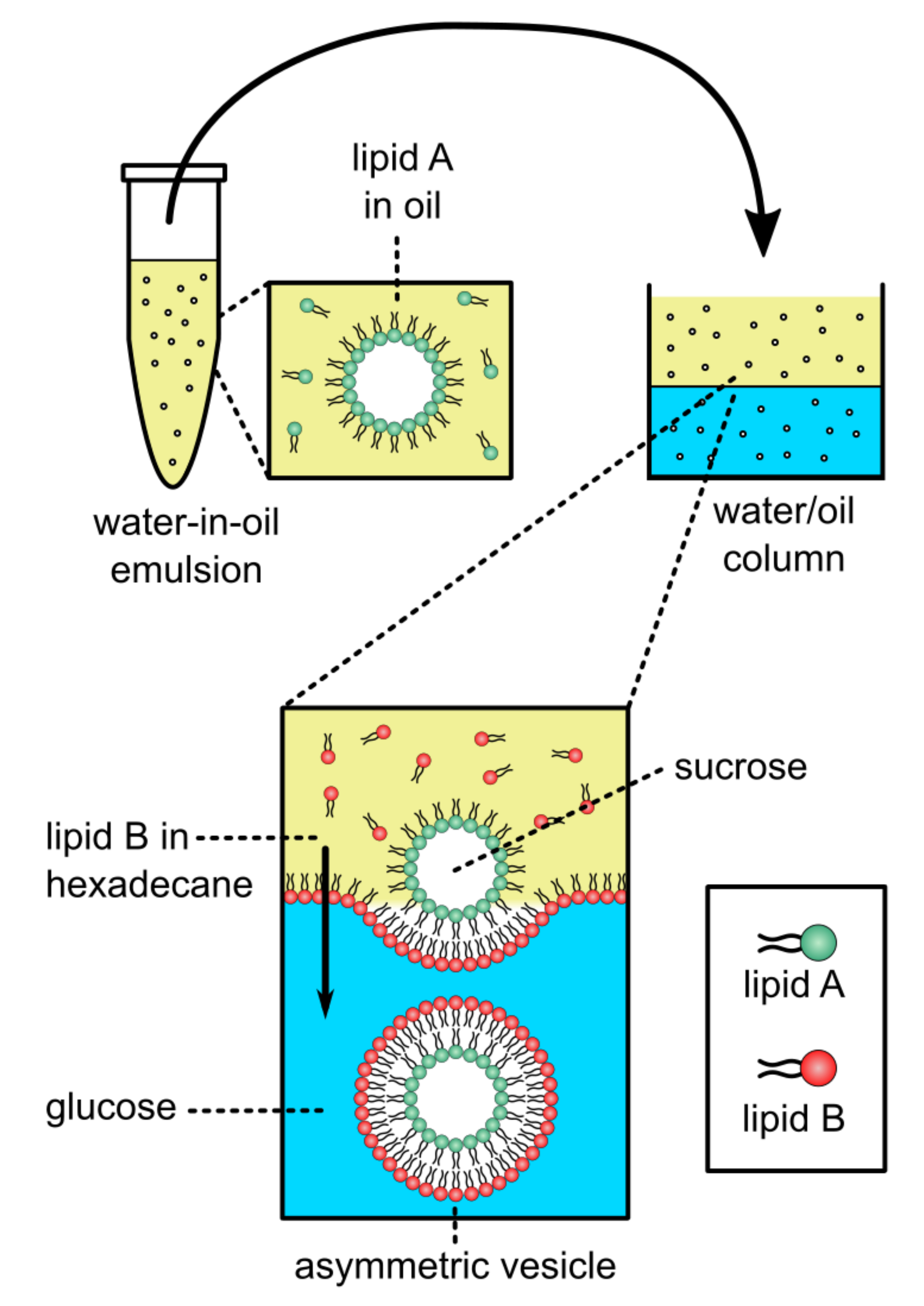
2.2.3. Phase Transfer, Water-Oil, Approaches Used in Generating aGUVs
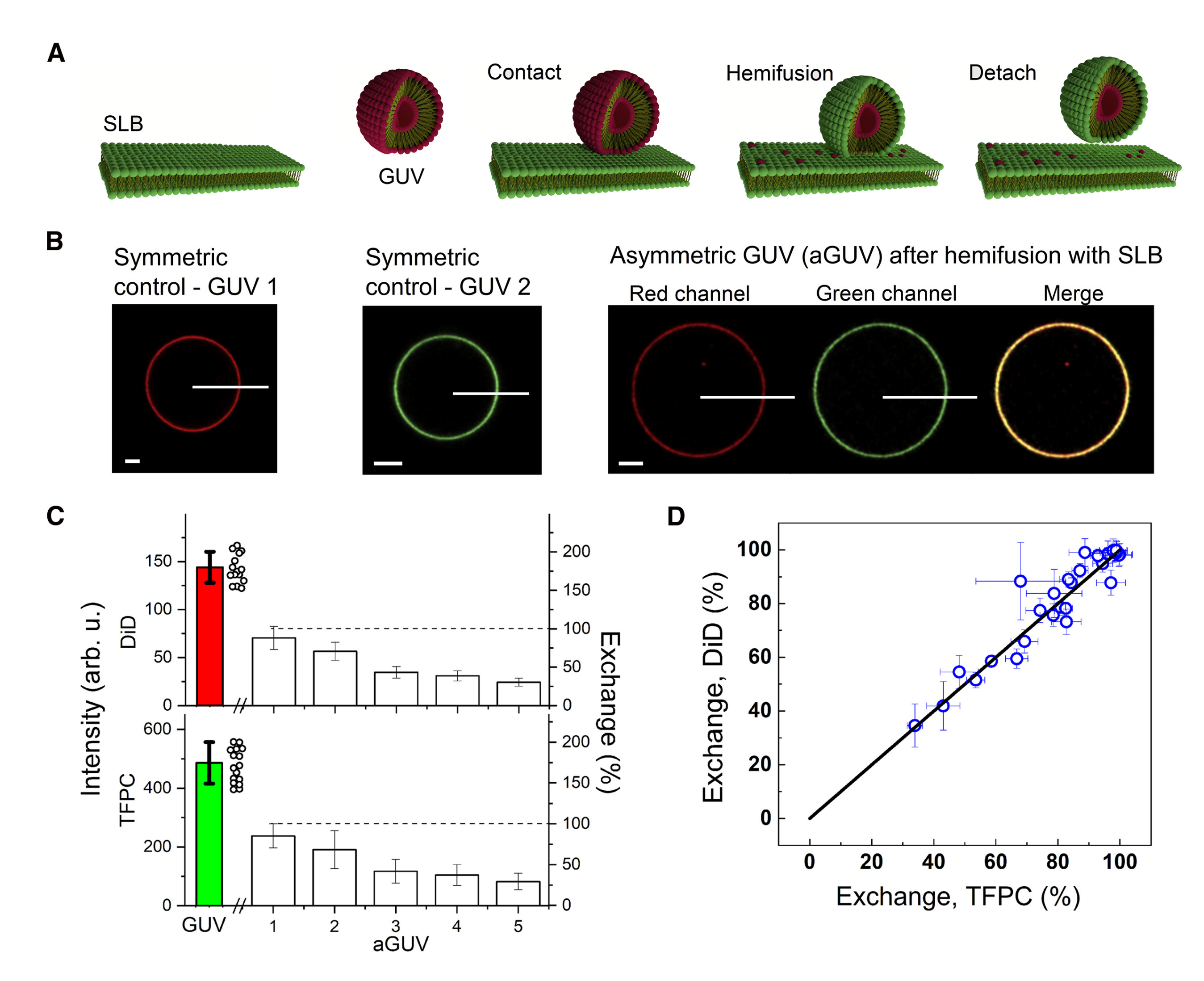
2.2.4. Hemifusion and aGUVs
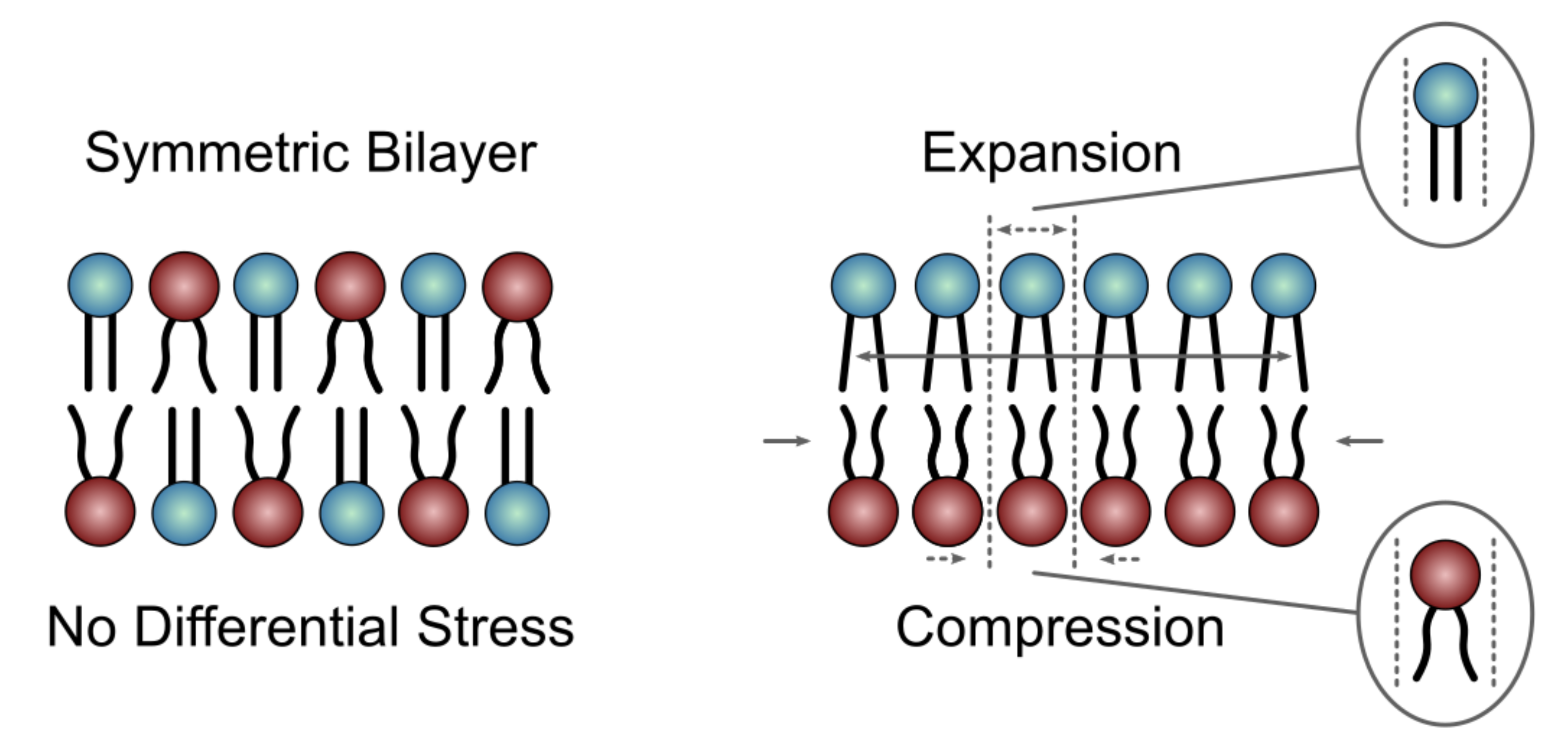
3. Molecular Dynamics (MD) Simulations Used to Model Asymmetric Membranes
4. Concluding Remarks
Author Contributions
Funding
Conflicts of Interest
References
- Bretscher, M.S. Asymmetrical lipid bilayer structure for biological membranes. Nat. New Biol. 1972, 236, 11–12. [Google Scholar] [CrossRef]
- Verkleij, A.; Zwaal, R.; Roelofsen, B.; Comfurius, P.; Kastelijn, D.; Van Deenen, L. The asymmetric distribution of phospholipids in the human red cell membrane. A combined study using phospholipases and freeze-etch electron microscopy. Biochim. Biophys. Acta Biomembr. 1973, 323, 178–193. [Google Scholar] [CrossRef]
- Op den Kamp, J. Lipid asymmetry in membranes. Annu. Rev. Biochem. 1979, 48, 47–71. [Google Scholar] [CrossRef] [PubMed]
- Van Meer, G.; Voelker, D.R.; Feigenson, G.W. Membrane lipids: Where they are and how they behave. Nat. Rev. Mol. 2008, 9, 112–124. [Google Scholar] [CrossRef] [PubMed]
- Gurtovenko, A.A.; Vattulainen, I. Lipid transmembrane asymmetry and intrinsic membrane potential: Two sides of the same coin. J. Am. Chem. Soc. 2007, 129, 5358–5359. [Google Scholar] [CrossRef]
- Entova, S.; Billod, J.-M.; Swiecicki, J.-M.; Martin-Santamaria, S.; Imperiali, B. Insights into the key determinants of membrane protein topology enable the identification of new monotopic folds. Elife 2018, 7, e40889. [Google Scholar] [CrossRef]
- Lorent, J.; Levental, K.; Ganesan, L.; Rivera-Longsworth, G.; Sezgin, E.; Doktorova, M.; Lyman, E.; Levental, I. Plasma membranes are asymmetric in lipid unsaturation, packing and protein shape. Nat. Chem. Biol. 2020, 16, 644–652. [Google Scholar] [CrossRef]
- Barenholz, Y.; Thompson, T. Sphingomyelins in bilayers and biological membranes. Biochim. Biophys. Acta Biomembr. 1980, 604, 129–158. [Google Scholar] [CrossRef]
- May, S. Trans-monolayer coupling of fluid domains in lipid bilayers. Soft Matter 2009, 5, 3148–3156. [Google Scholar] [CrossRef]
- Collins, M.D.; Keller, S.L. Tuning lipid mixtures to induce or suppress domain formation across leaflets of unsupported asymmetric bilayers. Proc. Natl. Acad. Sci. USA 2008, 105, 124–128. [Google Scholar] [CrossRef] [Green Version]
- Wagner, A.J.; Loew, S.; May, S. Influence of monolayer-monolayer coupling on the phase behavior of a fluid lipid bilayer. Biophys. J. 2007, 93, 4268–4277. [Google Scholar] [CrossRef] [Green Version]
- Chiantia, S.; London, E. Acyl chain length and saturation modulate interleaflet coupling in asymmetric bilayers: Effects on dynamics and structural order. Biophys. J. 2012, 103, 2311–2319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heberle, F.A.; Marquardt, D.; Doktorova, M.; Geier, B.; Standaert, R.F.; Heftberger, P.; Kollmitzer, B.; Nickels, J.D.; Dick, R.A.; Feigenson, G.W.; et al. Subnanometer structure of an asymmetric model membrane: Interleaflet coupling influences domain properties. Langmuir 2016, 32, 5195–5200. [Google Scholar] [CrossRef]
- Seigneuret, M.; Devaux, P.F. ATP-dependent asymmetric distribution of spin-labeled phospholipids in the erythrocyte membrane: Relation to shape changes. Proc. Natl. Acad. Sci. USA 1984, 81, 3751–3755. [Google Scholar] [CrossRef] [Green Version]
- Manno, S.; Takakuwa, Y.; Mohandas, N. Identification of a functional role for lipid asymmetry in biological membranes: Phosphatidylserine-skeletal protein interactions modulate membrane stability. Proc. Natl. Acad. Sci. USA 2002, 99, 1943–1948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, B.D.; Lambert, T.N. Molecular ferries: Membrane carriers that promote phospholipid flip-flop and chloride transport. Chem. Commun. 2003, 2261–2268. [Google Scholar] [CrossRef] [PubMed]
- Van Helvoort, A.; Smith, A.J.; Sprong, H.; Fritzsche, I.; Schinkel, A.H.; Borst, P.; van Meer, G. MDR1 P-glycoprotein is a lipid translocase of broad specificity, while MDR3 P-glycoprotein specifically translocates phosphatidylcholine. Cell 1996, 87, 507–517. [Google Scholar] [CrossRef] [Green Version]
- Rousselet, A.; Guthmann, C.; Matricon, J.; Bienvenue, A.; Devaux, P.F. Study of the transverse diffusion of spin labelled phospholipids in biological membranes. I. Human red blood cells. Biochim. Biophys. Acta Biomembr. 1976, 426, 357–371. [Google Scholar] [CrossRef]
- Van den Eijnde, S.; Boshart, L.; Baehrecke, E.; De Zeeuw, C.; Reutelingsperger, C.; Vermeij-Keers, C. Cell surface exposure of phosphatidylserine during apoptosis is phylogenetically conserved. Apoptosis 1998, 3, 9–16. [Google Scholar] [CrossRef]
- Fadeel, B.; Xue, D. The ins and outs of phospholipid asymmetry in the plasma membrane: Roles in health and disease. Crit. Rev. Biochem. Mol. 2009, 44, 264–277. [Google Scholar] [CrossRef] [PubMed]
- Gurtovenko, A.A.; Vattulainen, I. Membrane potential and electrostatics of phospholipid bilayers with asymmetric transmembrane distribution of anionic lipids. J. Phys. Chem. B. 2008, 112, 4629–4634. [Google Scholar] [CrossRef] [Green Version]
- Williamson, P.; Christie, A.; Kohlin, T.; Schlegel, R.A.; Comfurius, P.; Harmsma, M.; Zwaal, R.F.; Bevers, E.M. Phospholipid scramblase activation pathways in lymphocytes. Biochemistry 2001, 40, 8065–8072. [Google Scholar] [CrossRef] [PubMed]
- Shlomovitz, I.; Speir, M.; Gerlic, M. Flipping the dogma–phosphatidylserine in non-apoptotic cell death. Cell Commun. Signal 2019, 17, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doktorova, M.; Symons, J.L.; Levental, I. Structural and functional consequences of reversible lipid asymmetry in living membranes. Nat. Chem. Biol. 2020, 16, 1321–1330. [Google Scholar] [CrossRef]
- Kodigepalli, K.M.; Bowers, K.; Sharp, A.; Nanjundan, M. Roles and regulation of phospholipid scramblases. FEBS Lett. 2015, 589, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Crane, J.M.; Kiessling, V.; Tamm, L.K. Measuring lipid asymmetry in planar supported bilayers by fluorescence interference contrast microscopy. Langmuir 2005, 21, 1377–1388. [Google Scholar] [CrossRef] [PubMed]
- Pautot, S.; Frisken, B.J.; Weitz, D. Engineering asymmetric vesicles. Proc. Natl. Acad. Sci. USA 2003, 100, 10718–10721. [Google Scholar] [CrossRef] [Green Version]
- Drechsler, C.; Markones, M.; Choi, J.-Y.; Frieling, N.; Fiedler, S.; Voelker, D.R.; Schubert, R.; Heerklotz, H. Preparation of asymmetric liposomes using a phosphatidylserine decarboxylase. Biophys. J. 2018, 115, 1509–1517. [Google Scholar] [CrossRef] [Green Version]
- Takaoka, R.; Kurosaki, H.; Nakao, H.; Ikeda, K.; Nakano, M. Formation of asymmetric vesicles via phospholipase D-mediated transphosphatidylation. Biochim. Biophys. Acta Biomembr. 2018, 1860, 245–249. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.-Y.; Deng, G.; Jiang, Y.-W.; Zhou, Y.; Xu, J.; Wu, F.-G.; Yu, Z.-W. Controllable engineering of asymmetric phosphatidylserine-containing lipid vesicles using calcium cations. Chem. Commun. 2017, 53, 12762–12765. [Google Scholar] [CrossRef]
- Guo, H.-Y.; Sun, H.-Y.; Deng, G.; Xu, J.; Wu, F.-G.; Yu, Z.-W. Fabrication of Asymmetric Phosphatidylserine-Containing Lipid Vesicles: A Study on the Effects of Size, Temperature, and Lipid Composition. Langmuir 2020, 36, 12684–12691. [Google Scholar] [CrossRef] [PubMed]
- Enoki, T.A.; Feigenson, G.W. Asymmetric bilayers by hemifusion: Method and leaflet behaviors. Biophys. J. 2019, 117, 1037–1050. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.-T.; London, E. Preparation and properties of asymmetric vesicles that mimic cell membranes effect upon lipid raft formation and transmembrane helix orientation. J. Biol. Chem. 2009, 284, 6079–6092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, H.-T.; London, E. Preparation and properties of asymmetric large unilamellar vesicles: Interleaflet coupling in asymmetric vesicles is dependent on temperature but not curvature. Biophys. J. 2011, 100, 2671–2678. [Google Scholar] [CrossRef] [Green Version]
- Lin, Q.; London, E. Preparation of artificial plasma membrane mimicking vesicles with lipid asymmetry. PLoS ONE 2014, 9, e87903. [Google Scholar] [CrossRef]
- Doktorova, M.; Heberle, F.A.; Eicher, B.; Standaert, R.F.; Katsaras, J.; London, E.; Pabst, G.; Marquardt, D. Preparation of asymmetric phospholipid vesicles for use as cell membrane models. Nat. Protoc. 2018, 13, 2086–2101. [Google Scholar] [CrossRef]
- Marquardt, D.; Heberle, F.A.; Miti, T.; Eicher, B.; London, E.; Katsaras, J.; Pabst, G. 1H NMR shows slow phospholipid flip-flop in gel and fluid bilayers. Langmuir 2017, 33, 3731–3741. [Google Scholar] [CrossRef]
- Nguyen, M.H.; DiPasquale, M.; Rickeard, B.W.; Doktorova, M.; Heberle, F.A.; Scott, H.L.; Barrera, F.N.; Taylor, G.; Collier, C.P.; Stanley, C.B.; et al. Peptide-induced lipid flip-flop in asymmetric liposomes measured by small angle neutron scattering. Langmuir 2019, 35, 11735–11744. [Google Scholar] [CrossRef]
- Doktorova, M.; Heberle, F.A.; Marquardt, D.; Rusinova, R.; Sanford, R.L.; Peyear, T.A.; Katsaras, J.; Feigenson, G.W.; Weinstein, H.; Andersen, O.S. Gramicidin increases lipid flip-flop in symmetric and asymmetric lipid vesicles. Biophys. J. 2019, 116, 860–873. [Google Scholar] [CrossRef] [Green Version]
- Scott, H.L.; Heberle, F.A.; Katsaras, J.; Barrera, F.N. Phosphatidylserine asymmetry promotes the membrane insertion of a transmembrane helix. Biophys. J. 2019, 116, 1495–1506. [Google Scholar] [CrossRef] [PubMed]
- Clair, J.R.S.; Wang, Q.; Li, G.; London, E. The Biophysics of Cell Membranes; Springer: Berlin, Germany, 2017; pp. 1–27. [Google Scholar]
- Fenyvesi, E.; Zemlényi, C.; Orgoványi, J.; Oláh, E.; Szente, L. Can conversion mixture substitute beta-cyclodextrin in encapsulation of essential oils and their components? J. Incl. Phenom. Macrocycl. Chem. 2016, 86, 55–66. [Google Scholar] [CrossRef]
- Szente, L.; Fenyvesi, É. Cyclodextrin-lipid complexes: Cavity size matters. Struct. Chem. 2017, 28, 479–492. [Google Scholar] [CrossRef]
- Yao, J.; Ho, D.; Calingasan, N.Y.; Pipalia, N.H.; Lin, M.T.; Beal, M.F. Neuroprotection by cyclodextrin in cell and mouse models of Alzheimer disease. J. Exp. Med. 2012, 209, 2501–2513. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z.; London, E. Effect of cyclodextrin and membrane lipid structure upon cyclodextrin–lipid interaction. Langmuir 2013, 29, 14631–14638. [Google Scholar] [CrossRef] [PubMed]
- Miyajima, K.; Tomita, K.; Nakagaki, M. Complex formation between di-and monophosphatidylcholines and cyclodextrins in water. Chem. Pharm. Bull. 1985, 33, 2587–2590. [Google Scholar] [CrossRef] [Green Version]
- Anderson, T.G.; Tan, A.; Ganz, P.; Seelig, J. Calorimetric measurement of phospholipid interaction with methyl-β-cyclodextrin. Biochemistry 2004, 43, 2251–2261. [Google Scholar] [CrossRef]
- Ravichandran, R.; Divakar, S. Inclusion of ring A of cholesterol inside the β-cyclodextrin cavity: Evidence from oxidation reactions and structural studies. J. Incl. Phenom. Mol. Recognit. Chem. 1998, 30, 253–270. [Google Scholar] [CrossRef]
- Köhler, J.; Hohla, M.; Söllner, R.; Amann, M. The difference between cholesterol-and glycyrrhizin-γ-cyclodextrin complexes—An analysis by MD simulations in vacuo and in aquo and the calculation of solvation free energies with AMSOL. Supramol. Sci. 1998, 5, 117–137. [Google Scholar] [CrossRef]
- London, E. Membrane Structure–Function Insights from Asymmetric Lipid Vesicles. Acc. Chem. Res. 2019, 52, 2382–2391. [Google Scholar] [CrossRef]
- Lin, Q.; London, E. The influence of natural lipid asymmetry upon the conformation of a membrane-inserted protein (perfringolysin O). J. Biol. Chem. 2014, 289, 5467–5478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; London, E. Lipid structure and composition control consequences of interleaflet coupling in asymmetric vesicles. Biophys. J. 2018, 115, 664–678. [Google Scholar] [CrossRef] [PubMed]
- Perly, B.; Smith, I.; Hughes, L.; Burton, G.; Ingold, K. Estimation of the location of natural α-tocopherol in lipid bilayers by 13C-NMR spectroscopy. Biochim. Biophys. Acta Biomembr. 1985, 819, 131–135. [Google Scholar] [CrossRef]
- Eicher, B.; Marquardt, D.; Heberle, F.A.; Letofsky-Papst, I.; Rechberger, G.N.; Appavou, M.-S.; Katsaras, J.; Pabst, G. Intrinsic curvature-mediated transbilayer coupling in asymmetric lipid vesicles. Biophys. J. 2018, 114, 146–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scott, H.L.; Nguyen, V.P.; Alves, D.S.; Davis, F.L.; Booth, K.R.; Bryner, J.; Barrera, F.N. The negative charge of the membrane has opposite effects on the membrane entry and exit of pH-low insertion peptide. Biochemistry 2015, 54, 1709–1712. [Google Scholar] [CrossRef] [PubMed]
- Markones, M.; Drechsler, C.; Kaiser, M.; Kalie, L.; Heerklotz, H.; Fiedler, S. Engineering Asymmetric Lipid Vesicles: Accurate and Convenient Control of the Outer Leaflet Lipid Composition. Langmuir 2018, 34, 1999–2005. [Google Scholar] [CrossRef] [PubMed]
- Markones, M.; Fippel, A.; Kaiser, M.; Drechsler, C.; Hunte, C.; Heerklotz, H. Stairway to Asymmetry: Five Steps to Lipid-Asymmetric Proteoliposomes. Biophys. J. 2020, 118, 294–302. [Google Scholar] [CrossRef]
- Doktorova, M. On the Long and Winding Road to a Perfect Membrane Model. Biophys. J. 2020, 118, 273–275. [Google Scholar] [CrossRef]
- Chiantia, S.; Schwille, P.; Klymchenko, A.S.; London, E. Asymmetric GUVs prepared by MβCD-mediated lipid exchange: An FCS study. Biophys. J. 2011, 100, L1–L3. [Google Scholar] [CrossRef] [Green Version]
- Kahya, N.; Scherfeld, D.; Bacia, K.; Poolman, B.; Schwille, P. Probing lipid mobility of raft-exhibiting model membranes by fluorescence correlation spectroscopy. J. Biol. Chem. 2003, 278, 28109–28115. [Google Scholar] [CrossRef] [Green Version]
- Edholm, O.; Nagle, J.F. Areas of molecules in membranes consisting of mixtures. Biophys. J. 2005, 89, 1827–1832. [Google Scholar] [CrossRef] [Green Version]
- Alwarawrah, M.; Dai, J.; Huang, J. A molecular view of the cholesterol condensing effect in DOPC lipid bilayers. J. Phys. Chem. B 2010, 114, 7516–7523. [Google Scholar] [CrossRef] [Green Version]
- Sun, W.; Tristram-Nagle, S.; Suter, R.; Nagle, J. Structure of gel phase saturated lecithin bilayers: Temperature and chain length dependence. Biophys. J. 1996, 71, 885–891. [Google Scholar] [CrossRef] [Green Version]
- Weinberger, R. Practical Capillary Electrophoresis; Elsevier: Amsterdam, The Netherlands, 2000. [Google Scholar]
- Bozelli Jr, J.C.; Hou, Y.H.; Epand, R.M. Thermodynamics of methyl-β-cyclodextrin-induced lipid vesicle solubilization: Effect of lipid headgroup and backbone. Langmuir 2017, 33, 13882–13891. [Google Scholar] [CrossRef] [PubMed]
- Lin, Q.; London, E. Ordered raft domains induced by outer leaflet sphingomyelin in cholesterol-rich asymmetric vesicles. Biophys. J. 2015, 108, 2212–2222. [Google Scholar] [CrossRef] [Green Version]
- Elani, Y.; Purushothaman, S.; Booth, P.J.; Seddon, J.M.; Brooks, N.J.; Law, R.V.; Ces, O. Measurements of the effect of membrane asymmetry on the mechanical properties of lipid bilayers. Chem. Commun. 2015, 51, 6976–6979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charalambous, K.; Miller, D.; Curnow, P.; Booth, P.J. Lipid bilayer composition influences small multidrug transporters. BMC Biochem. 2008, 9, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Hu, P.C.; Li, S.; Malmstadt, N. Microfluidic fabrication of asymmetric giant lipid vesicles. ACS Appl. Mater. Interfaces 2011, 3, 1434–1440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamada, T.; Yoshikawa, K. Cell-sized liposomes and droplets: Real-world modeling of living cells. Materials 2012, 5, 2292–2305. [Google Scholar] [CrossRef]
- Lu, L.; Doak, W.J.; Schertzer, J.W.; Chiarot, P.R. Membrane mechanical properties of synthetic asymmetric phospholipid vesicles. Soft Matter 2016, 12, 7521–7528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hossein, A.; Deserno, M. Spontaneous curvature, differential stress, and bending modulus of asymmetric lipid membranes. Biophys. J. 2020, 118, 624–642. [Google Scholar] [CrossRef] [PubMed]
- Angelova, M.I.; Dimitrov, D.S. Liposome electroformation. Faraday Discuss. Chem. Soc. 1986, 81, 303–311. [Google Scholar] [CrossRef]
- Usery, R.D.; Enoki, T.A.; Wickramasinghe, S.P.; Weiner, M.D.; Tsai, W.-C.; Kim, M.B.; Wang, S.; Torng, T.L.; Ackerman, D.G.; Heberle, F.A.; et al. Line tension controls liquid-disordered+ liquid-ordered domain size transition in lipid bilayers. Biophys. J. 2017, 112, 1431–1443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enoki, T.A.; Heberle, F.A.; Feigenson, G.W. FRET detects the size of nanodomains for coexisting liquid-disordered and liquid-ordered phases. Biophys. J. 2018, 114, 1921–1935. [Google Scholar] [CrossRef] [Green Version]
- Heberle, F.A.; Petruzielo, R.S.; Pan, J.; Drazba, P.; Kučerka, N.; Standaert, R.F.; Feigenson, G.W.; Katsaras, J. Bilayer thickness mismatch controls domain size in model membranes. J. Am. Chem. Soc. 2013, 135, 6853–6859. [Google Scholar] [CrossRef] [PubMed]
- Heberle, F.A.; Doktorova, M.; Scott, H.L.; Skinkle, A.D.; Waxham, M.N.; Levental, I. Direct label-free imaging of nanodomains in biomimetic and biological membranes by cryogenic electron microscopy. Proc. Natl. Acad. Sci. USA 2020, 117, 19943–19952. [Google Scholar] [CrossRef]
- Enoki, T.A.; Wu, J.; Heberle, F.A.; Feigenson, G.W. Investigation of the domain line tension in asymmetric vesicles prepared via hemifusion. Biochim. Biophys. Acta Biomembr. 2021, 1863, 183586. [Google Scholar] [CrossRef] [PubMed]
- Perlmutter, J.D.; Sachs, J.N. Interleaflet interaction and asymmetry in phase separated lipid bilayers: Molecular dynamics simulations. J. Am. Chem. Soc. 2011, 133, 6563–6577. [Google Scholar] [CrossRef] [PubMed]
- Koldsø, H.; Shorthouse, D.; Hélie, J.; Sansom, M.S. Lipid clustering correlates with membrane curvature as revealed by molecular simulations of complex lipid bilayers. PLoS Comput. Biol. 2014, 10, e1003911. [Google Scholar] [CrossRef]
- Marrink, S.J.; Corradi, V.; Souza, P.C.; Ingólfsson, H.I.; Tieleman, D.P.; Sansom, M.S. Computational modeling of realistic cell membranes. Chem. Rev. 2019, 119, 6184–6226. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.; Kim, B.N.; Stansfeld, P.J.; Sansom, M.S.; Lindau, M. A coarse grained model for a lipid membrane with physiological composition and leaflet asymmetry. PLoS ONE 2015, 10, e0144814. [Google Scholar] [CrossRef] [Green Version]
- Pezeshkian, W.; Marrink, S.J. Simulating realistic membrane shapes. Curr. Opin. Cell Biol. 2021, 71, 103–111. [Google Scholar] [CrossRef]
- Ingólfsson, H.I.; Melo, M.N.; Van Eerden, F.J.; Arnarez, C.; Lopez, C.A.; Wassenaar, T.A.; Periole, X.; De Vries, A.H.; Tieleman, D.P.; Marrink, S.J. Lipid organization of the plasma membrane. J. Am. Chem. Soc. 2014, 136, 14554–14559. [Google Scholar] [CrossRef]
- Mohideen, N.; Weiner, M.D.; Feigenson, G.W. Bilayer compositional asymmetry influences the nanoscopic to macroscopic phase domain size transition. Chem. Phys. Lipids 2020, 232, 104972. [Google Scholar] [CrossRef]
- Shahane, G.; Ding, W.; Palaiokostas, M.; Orsi, M. Physical properties of model biological lipid bilayers: Insights from all-atom molecular dynamics simulations. J. Mol. Model. 2019, 25, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Aghaaminiha, M.; Farnoud, A.M.; Sharma, S. Quantitative relationship between cholesterol distribution and ordering of lipids in asymmetric lipid bilayers. Soft Matter 2021, 17, 2742–2752. [Google Scholar] [CrossRef] [PubMed]
- Falkovich, S.G.; Martinez-Seara, H.; Nesterenko, A.M.; Vattulainen, I.; Gurtovenko, A.A. What can we learn about cholesterol’s transmembrane distribution based on cholesterol-induced changes in membrane dipole potential? J. Phys. Chem. Lett. 2016, 7, 4585–4590. [Google Scholar] [CrossRef]
- Vácha, R.; Berkowitz, M.L.; Jungwirth, P. Molecular model of a cell plasma membrane with an asymmetric multicomponent composition: Water permeation and ion effects. Biophys. J. 2009, 96, 4493–4501. [Google Scholar] [CrossRef] [Green Version]
- Polley, A.; Mayor, S.; Rao, M. Bilayer registry in a multicomponent asymmetric membrane: Dependence on lipid composition and chain length. J. Chem. Phys. 2014, 141, 064903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blumer, M.; Harris, S.; Li, M.; Martinez, L.; Untereiner, M.; Saeta, P.N.; Carpenter, T.S.; Ingólfsson, H.I.; Bennett, W. Simulations of Asymmetric Membranes Illustrate Cooperative Leaflet Coupling and Lipid Adaptability. Front. Cell Dev. Biol. 2020, 8, 575. [Google Scholar] [CrossRef]
- Shearer, J.; Khalid, S. Communication between the leaflets of asymmetric membranes revealed from coarse-grain molecular dynamics simulations. Sci. Rep. 2018, 8, 1805. [Google Scholar] [CrossRef] [PubMed]
- Weiner, M.D.; Feigenson, G.W. Molecular dynamics simulations reveal leaflet coupling in compositionally asymmetric phase-separated lipid membranes. J. Phys. Chem. B 2019, 123, 3968–3975. [Google Scholar] [CrossRef] [PubMed]
- Esteban-Martín, S.; Risselada, H.J.; Salgado, J.; Marrink, S.J. Stability of asymmetric lipid bilayers assessed by molecular dynamics simulations. J. Am. Chem. Soc. 2009, 131, 15194–15202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.; Beaven, A.H.; Klauda, J.B.; Im, W. How tolerant are membrane simulations with mismatch in area per lipid between leaflets? J. Chem. Theory Comput. 2015, 11, 3466–3477. [Google Scholar] [CrossRef] [PubMed]
- Levadnyy, V.; Hasan, M.; Saha, S.K.; Yamazaki, M. Effect of Transmembrane Asymmetric Distribution of Lipids and Peptides on Lipid Bilayers. J. Phys. Chem. B. 2019, 123, 4645–4652. [Google Scholar] [CrossRef] [PubMed]
- Doktorova, M.; Weinstein, H. Accurate in silico modeling of asymmetric bilayers based on biophysical principles. Biophys. J. 2018, 115, 1638–1643. [Google Scholar] [CrossRef] [Green Version]
- Ollila, O.S.; Vattulainen, I. Lateral pressure profiles in lipid membranes: Dependence on molecular composition. Mol. Simul. Biomembr. Biophys. Funct. 2010, 20, 26. [Google Scholar]
- Hossein, A.; Deserno, M. Stiffening transition in asymmetric lipid bilayers: The role of highly ordered domains and the effect of temperature and size. J. Chem. Phys. 2021, 154, 014704. [Google Scholar] [CrossRef]
- Foley, S.; Deserno, M. Stabilizing Leaflet Asymmetry under Differential Stress in a Highly Coarse-Grained Lipid Membrane Model. J. Chem. Theory Comput. 2020, 16, 7195–7206. [Google Scholar] [CrossRef]
- Rickeard, B.W.; Nguyen, M.H.; DiPasquale, M.; Yip, C.G.; Baker, H.; Heberle, F.A.; Zuo, X.; Kelley, E.G.; Nagao, M.; Marquardt, D. Transverse lipid organization dictates bending fluctuations in model plasma membranes. Nanoscale 2020, 12, 1438–1447. [Google Scholar] [CrossRef]
- Levental, I.; Levental, K.R.; Heberle, F.A. Lipid rafts: Controversies resolved, mysteries remain. Trends Cell Biol. 2020, 30, 341–353. [Google Scholar] [CrossRef]
- Simons, K.; Ikonen, E. Functional rafts in cell membranes. Nature 1997, 387, 569–572. [Google Scholar] [CrossRef] [PubMed]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scott, H.L.; Kennison, K.B.; Enoki, T.A.; Doktorova, M.; Kinnun, J.J.; Heberle, F.A.; Katsaras, J. Model Membrane Systems Used to Study Plasma Membrane Lipid Asymmetry. Symmetry 2021, 13, 1356. https://doi.org/10.3390/sym13081356
Scott HL, Kennison KB, Enoki TA, Doktorova M, Kinnun JJ, Heberle FA, Katsaras J. Model Membrane Systems Used to Study Plasma Membrane Lipid Asymmetry. Symmetry. 2021; 13(8):1356. https://doi.org/10.3390/sym13081356
Chicago/Turabian StyleScott, Haden L., Kristen B. Kennison, Thais A. Enoki, Milka Doktorova, Jacob J. Kinnun, Frederick A. Heberle, and John Katsaras. 2021. "Model Membrane Systems Used to Study Plasma Membrane Lipid Asymmetry" Symmetry 13, no. 8: 1356. https://doi.org/10.3390/sym13081356
APA StyleScott, H. L., Kennison, K. B., Enoki, T. A., Doktorova, M., Kinnun, J. J., Heberle, F. A., & Katsaras, J. (2021). Model Membrane Systems Used to Study Plasma Membrane Lipid Asymmetry. Symmetry, 13(8), 1356. https://doi.org/10.3390/sym13081356