
Fluctuating Asymmetry of Craniological Features of Small Mammals as a Reflection of Heterogeneity of Natural Populations



Abstract
:1. Introduction
2. Materials and Methods
3. Results and Discussion
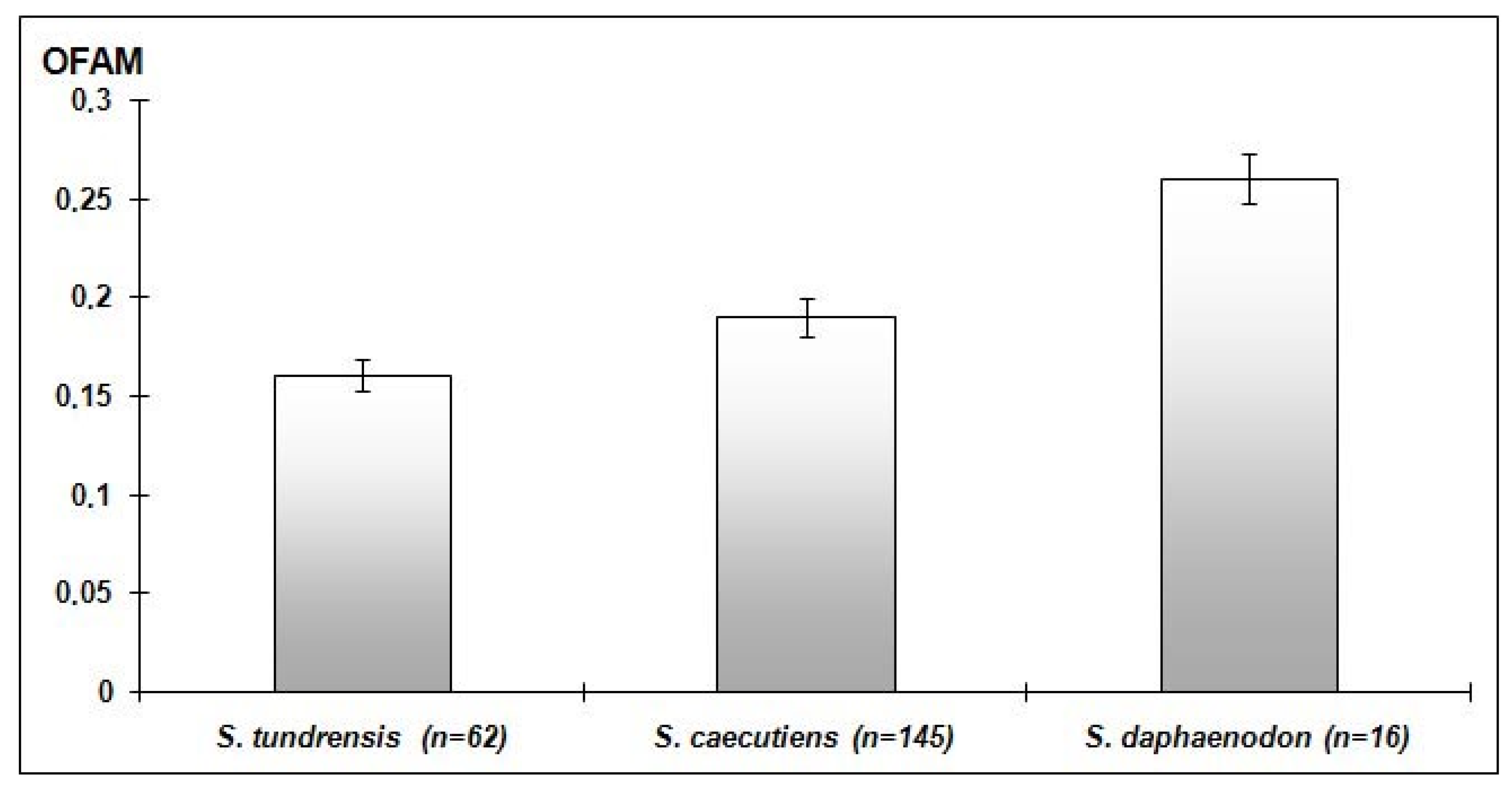
3.1. Geographical and Zonal Differences in FA Level
3.2. Intrapopulation Variability in Developmental Stability Values
3.2.1. Sex-Related Differences
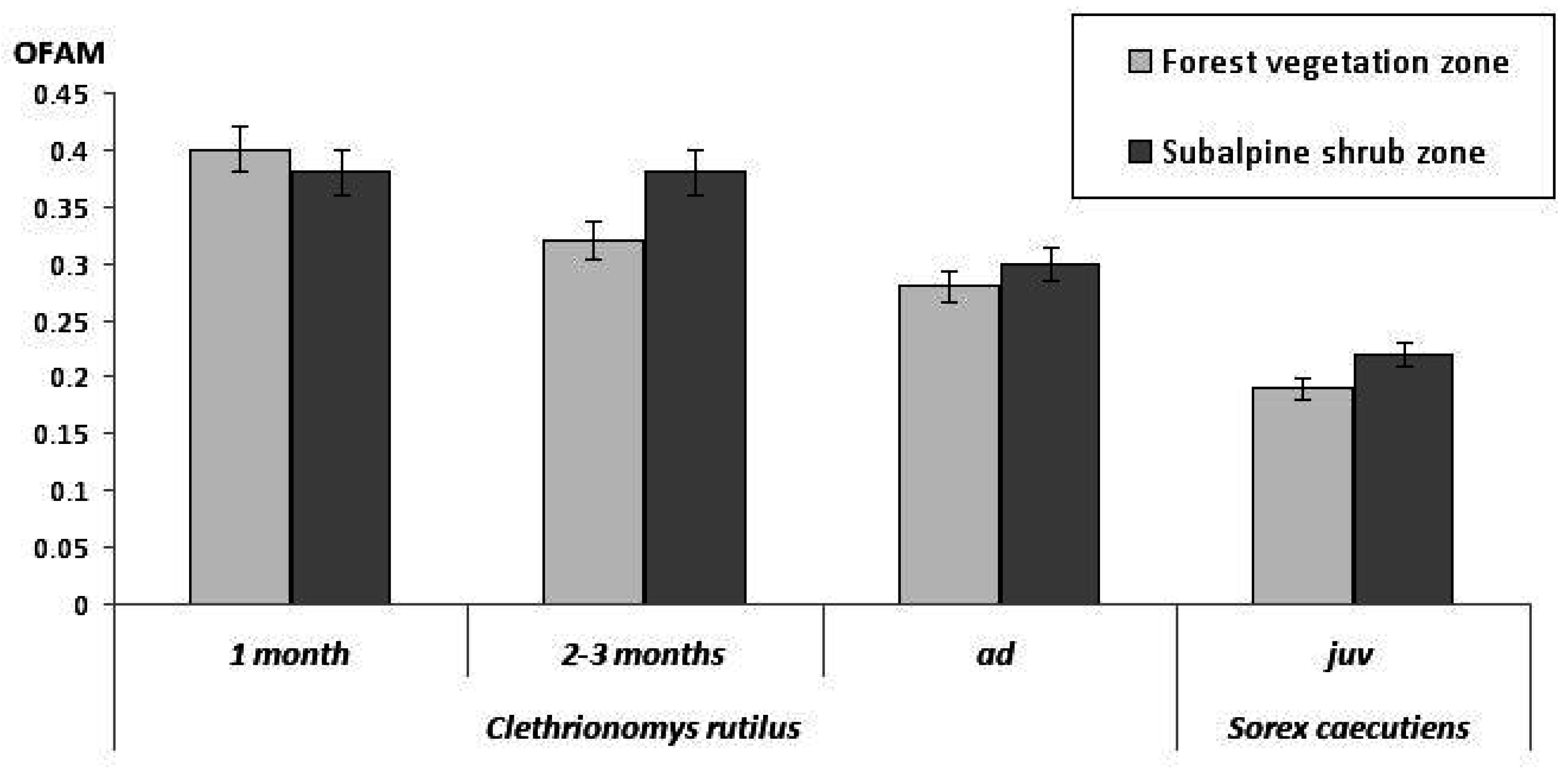
3.2.2. Age-Related Differences
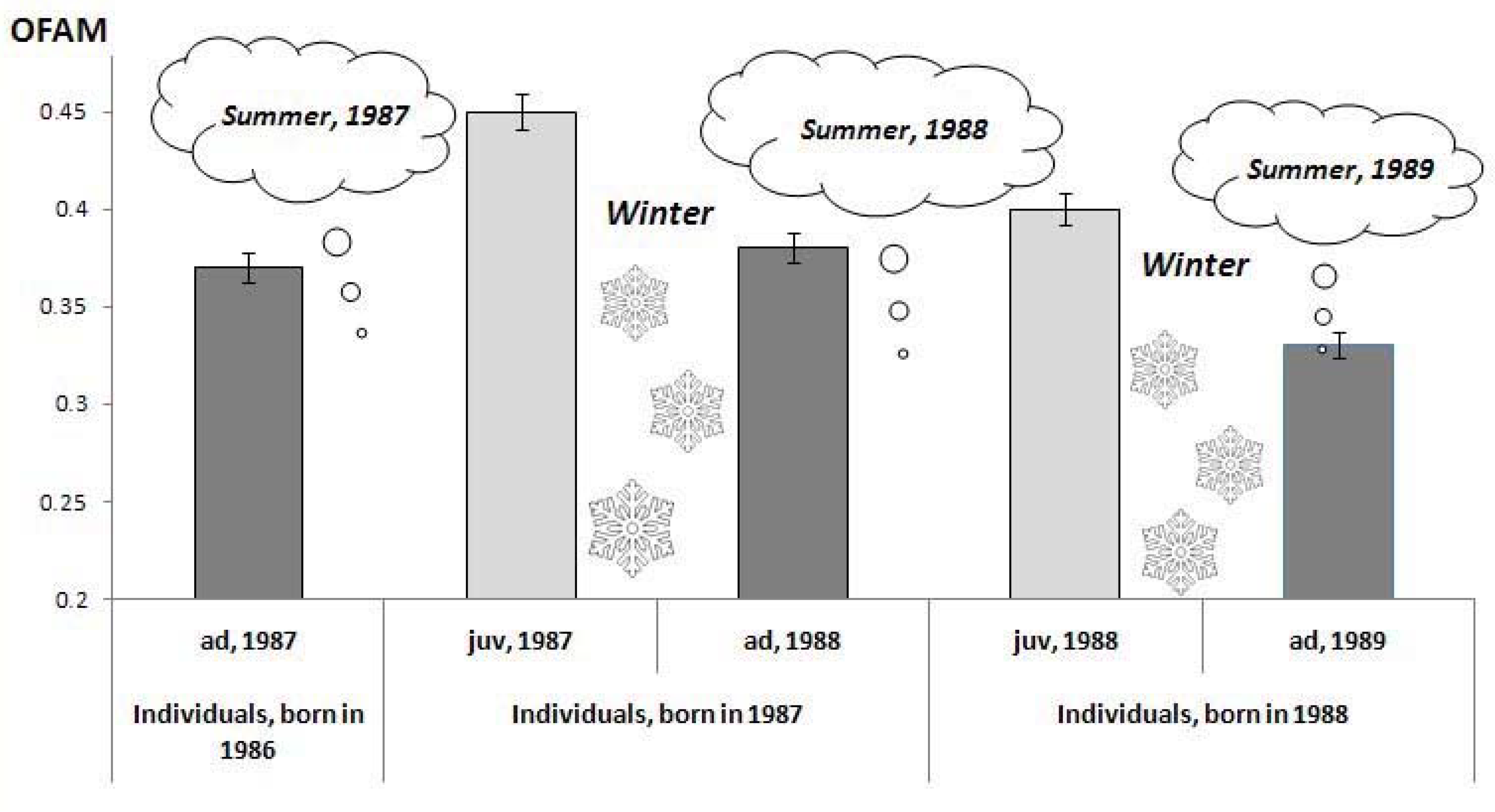
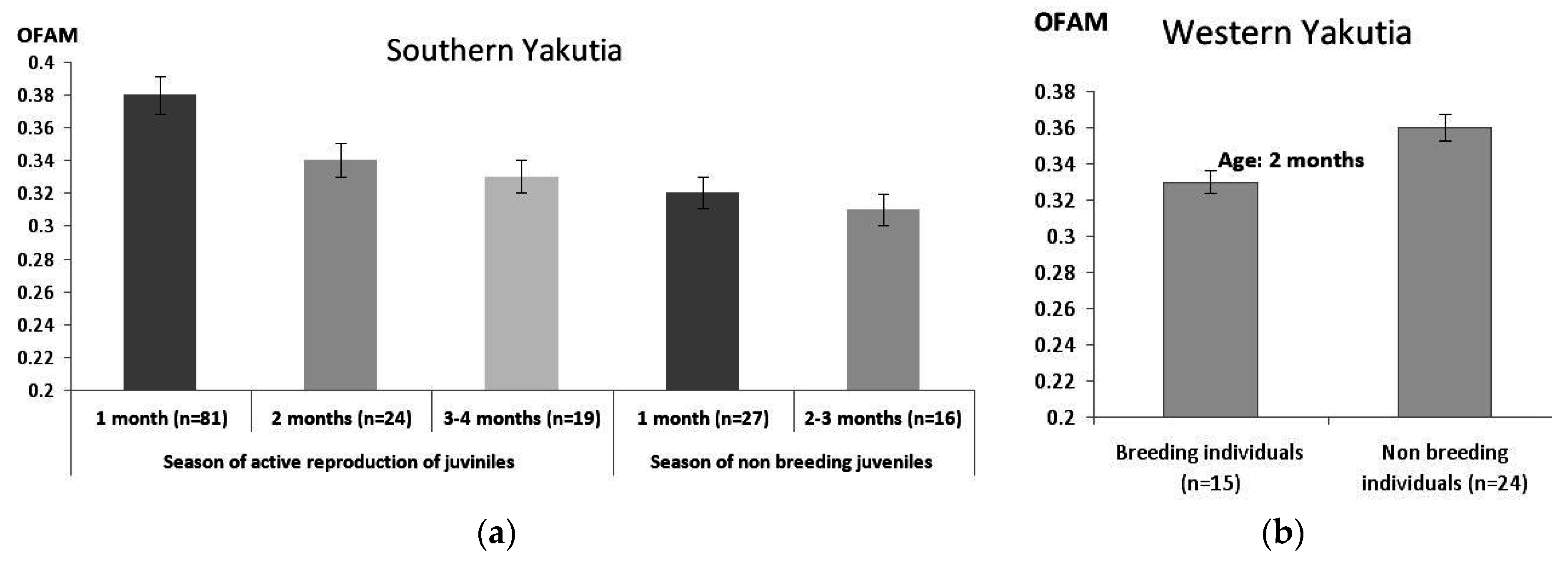
3.2.3. Changes in FA Level within One Generation Lifetime
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Van Valen, L. Study of fluctuating asymmetry. Evolution 1962, 16, 125–146. [Google Scholar] [CrossRef]
- Leary, R.F.; Allendorf, F.W. Fluctuating asymmetry as an indicator stress: Chance use in protect nature. Acta Zool. Fenica 1989, 4, 214–217. [Google Scholar]
- Parsons, P.A. Fluctuating asymmetry: An epigenetic measure of stress. Biol. Rev. 1990, 65, 131–145. [Google Scholar] [CrossRef] [PubMed]
- Parsons, P.A. Fluctuating asymmetry: A biological monitor of environmental and genomic stress. Heredity 1992, 68, 361–364. [Google Scholar] [CrossRef] [PubMed]
- Zakharov, V.M.; Zhdanova, N.P.; Kirik, E.F.; Shkil’, F.N. Ontogenesis and Population: Evaluation of Developmental Stability in Natural Populations. Russ. J. Dev. Biol. 2001, 32, 336–351. [Google Scholar] [CrossRef]
- Palmer, R.A.; Strobeck, C. Fluctuating assymmetry: A measurement, analysis, patterns. Annu. Rev. Ecol. Syst. 1986, 17, 391–421. [Google Scholar] [CrossRef]
- Zakharov, V.M. Animal Asymmetry: Population-Phenogenetic Approach (Asimmetriya Zhivotnykh: Populyatsionnofenogeneticheskii Podkhod); Nauka: Moscow, Russia, 1987; p. 216. (In Russian) [Google Scholar]
- Graham John, H.; Raz, S.H.; Hel-Or, H.; Nevo, E. Fluctuating Asymmetry: Methods, Theory, and Applications. Symmetry 2010, 2, 466–540. [Google Scholar] [CrossRef]
- Markow, T.A.; Ricker, J.P. Developmental stability in hybrids between the sibling species pair, Drosophila melanogaster and Drosophila simulans. Genetica 1991, 84, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Alibert, P.; Renaud, S.; Dod, B.; Bonhomme, F.; Auffray, J.C. Fluctuating asymmetry in the Mus musculus hybrid zone: A heterotic effect in disrupted coadapted genomes. Proc. R. Soc. Lond. B 1994, 258, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Auffray, J.-C.; Alibert, P.; Renaud, S.; Orth, A.; François Bonhomm, F. Fluctuating asymmetry in Mus musculus subspecific hybridization. In Traditional and Procrustes Comparative Approach, Advances in Morphometrics; Marcus, L.F., Corti, M., Loy, A., Naylor, G.J.P., Slice, D.E., Eds.; Plenum Press: New York, NY, USA, 1996; pp. 275–283. [Google Scholar]
- Hosken, D.J.; Blanckenhorn, W.U.; Ward, P.I. Developmental stability in yellow dung flies (Scathophaga stercoraria): Fluctuating asymmetry, heterozygosity and environmental stress. J. Evol. Biol. 2000, 13, 919–926. [Google Scholar] [CrossRef]
- Fowler, K.; Whitlock, M.C. Fluctuating asymmetry does not increase with moderate inbreeding in Drosophila melanogaster. Heredity 1994, 73, 373–376. [Google Scholar] [CrossRef] [PubMed]
- Zakharov, V.M.; Sikorski, M.D. Inbreeding and developmental stability in a laboratory strain of the bank vole Clethrionomys glareolus. Acta Theriol. 1997, 4, 73–78. [Google Scholar] [CrossRef]
- Borisov, V.I.; Valetsky, A.V.; Dmitrieva, I.L.; Krushinskaya, N.L.; Zakharov, V.M. Inbreeding and outbreeding impact on developmental stability of laboratory rat Rattus norvegicus. Acta Theriol. 1997, 4, 67–72. [Google Scholar] [CrossRef]
- Valentine, D.W.; Soule, M. Effect of p,p’-DDT on developmental stability of pectoral fin rays in the grunion Leuresthes tenuis. Nat. Mar. Fish. Serv. Fish. Bull. 1971, 71, 921–925. [Google Scholar]
- Pankakoski, E.; Koivisto, I.; Hyvärinen, H. Reduced developmental stability as an indicator of heavy metal pollution in the common shrew Sorex araneus. Acta Zool. Fenn. 1992, 191, 137–144. [Google Scholar]
- Graham, J.H.; Roe, K.E.; West, T.B. Effects of lead and benzene on developmental stability of Drosophila melanogaster. Ecotoxicology 1993, 2, 185–195. [Google Scholar] [CrossRef] [PubMed]
- Borisov, V.I.; Baranov, A.S.; Valetsky, A.V.; Zakharov, V.M. Developmental stability of the mink Mustela vison under the impact of PCB. IBID 1997, 4, 17–26. [Google Scholar] [CrossRef]
- Allenbach, D.M.; Sullivan, K.B.; Lyary, M.J. Higher Fluctuating asymmetry as a measure of susceptibility to Pesticides in fishes. Environ. Toxycol. Chem. 1999, 18, 899–905. [Google Scholar] [CrossRef]
- Allen, D.; Leamy, L.J. Effects of 2,3,4,8-tetrachloro-p-dioxin on directional and fluctuating asymmetry of mandible characters in mice. Ecotoxicology 2001, 10, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Michaelsen, S.A. Fluctuating Asymmetry of Menidia beryllina as a Measure of the Environmental Stress Caused by the 2010 Deepwater Horizon Oil Spill. Bachelor’s Thesis, The University of Southern Mississippi, Hattiesburg, MS, USA, 2013; p. 114. Available online: http://aquila.usm.edu/honors_theses/114 (accessed on 14 July 2016). [Google Scholar]
- Chubinishvili, A.T. Homeostasis in the populations of marsh frog (Rana ridibunda Pall) living in conditions of chemical pollution in the area of the Middle Volga. Russ. J. Ecol. 1998, 1, 71–74. (In Russian) [Google Scholar]
- Estes, E.C.; Katholi, C.R.; Angus, R.A. Elevated fluctuating asymmetry in eastern mosquitofish (Gambusia holbrooki) from a river receiving paper mill effluent. Environ. Toxicol. Chem. 2006, 25, 1026–1033. [Google Scholar] [CrossRef] [PubMed]
- Shadrina, E.G.; Vol’pert, Y.L. Developmental Instability of the Organism as a Result of Pessimization of Environment under Anthropogenic Transformation of Natural Landscapes. Russ. J. Dev. Biol. 2014, 45, 117–126. [Google Scholar] [CrossRef]
- Zhelev, Z.M.; Popgeorgiev, G.S.; Arnaudov, A.D.; Georgieva, K.N.; Mehterov, N.H. Fluctuating Asymmetry in Pelophylax Ridibundus (Amphibia: Ranidae) as a Response to Anthropogenic Pollution in South Bulgaria. Arch. Biol. Sci. 2015, 67, 1009–1023. [Google Scholar] [CrossRef]
- Zakharov, V.M.; Demin, D.V.; Baranov, A.S.; Borisov, V.I.; Valetsky, A.V.; Sheftel, B.I. Developmental stability and population dynamics of shrews Sorex in Central Siberia. Acta Theriol. 1997, 4, 41–48. [Google Scholar] [CrossRef]
- Leary, R.F.; Allendorf, F.W.; Knudsen, K.L. Genetic, environmental, and developmental causes of meristic varitation in rainbow trout. Acta Zool. Fennica 1992, 191, 79–95. [Google Scholar]
- Clarke, G.M.; McKenzie, J.A. Fluctuating Asymmetry as a Quality Control Indicator for Insect Mass Rearing Processes. Entomol. Soc. Am. 1992, 85, 1–6. [Google Scholar] [CrossRef]
- Jones, J.C.; Helliwell, P.; Beekman, M.; Maleszka, R.; Oldroyd, B.P. The effects of rearing temperature on developmental stability and learning and memory in the honey bee, Apis mellifera. J. Comp. Physiol. A 2005, 191. [Google Scholar] [CrossRef] [PubMed]
- Vishalakshi, C.; Singh, B.N. Effect of developmental temperature stress on fluctuating asymmetry in certain morphological traits in Drosophila ananassae. J. Therm. Biol. 2007, 33, 201–208. [Google Scholar] [CrossRef]
- Polak, M.; Kroeger, D.E.; Cartwright, I.L.; Ponce deLeon, C. Genotype-specific responses of fluctuating asymmetry and of preadult survival to the effects of lead and temperature stress in Drosophila melanogaster. Environ. Pollut. 2004, 127, 145–155. [Google Scholar] [CrossRef]
- Siegel, M.I.; Mooney, M.P.; Taylor, A.B. Dental and skeletal reduction as a consequence of environmental stress. Acta Zool. Fennica 1992, 191, 145–149. [Google Scholar]
- Anciães, M.; Marini, M.Ã. The effects of fragmentation on fluctuating asymmetry in passerine birds of Brazilian tropical forests. J. Appl. Ecol. 2000, 37, 1013–1028. [Google Scholar] [CrossRef]
- Cuervo, A.M.; Restrepo, C. Assemblage and population-level consequences of forest fragmentation on bilateral asymmetry in tropical montane birds. Biol. J. Linn. Soc. 2007, 92, 119–133. [Google Scholar] [CrossRef]
- Wojcik, J.M.; Polly, P.D.; Wojcik, A.M.; Sikorski, M.D. Epigenetic variation of the common shrew, Sorex araneus, in different habitats. Russ. J. Theriol. 2007, 6, 43–49. [Google Scholar]
- Stige, L.C.; Slagsvold, T.; Vøllestad, L.A. Individual fluctuating asymmetry in Pied Flycatchers (Ficedula hypoleuca) persists across moults, but is not heritable and not related to fitness. Evol. Ecol. Res. 2005, 7, 381–406. [Google Scholar]
- Bartoš, L.; Bahbouh, R.; Vach, M. Repeatability of size and fluctuating asymmetry of antler characteristics in red deer (Cervus elaphus) during ontogeny. Biol. J. Linn. Soc. 2007, 91, 215–226. [Google Scholar] [CrossRef]
- Mateos, C.; Alarcos, S.; Carranza, J.; Sánchez-Prieto, C.B.; Valencia, J. Fluctuating asymmetry of red deer antlers negatively relates to individual condition and proximity to prime age. Anim. Behav. 2008, 75, 1629–1640. [Google Scholar] [CrossRef]
- Naugler, C.T.; Leech, S.M. Fluctuating Asymmetry and Survival Ability in the Forest Tent Caterpillar Moth Malacosoma disstria: Implications for Pest Management. Entomol. Exp. Appl. 1994, 70, 295–298. [Google Scholar] [CrossRef]
- Moller, A.P. Fluctuating Asymmetry in Male Sexual Ornaments May Reliably Reveal Male Quality. Anim. Behav. 1990, 40, 1185–1187. [Google Scholar] [CrossRef]
- Muzlanov Yu, A. Otsenka sostoyaniya prirodnykh populyatsii po gomeostazu razvitiya na primere analiza raspredeleniya anomalii zhilkovaniya kryl’ev strekozy krasotki blestyashchei (Calopteryx splendens Harr). Russ. J. Ecol. 1997, 6, 442–446. (In Russian) [Google Scholar]
- Khirevich, E.A. Strukturno-Funktsional’nyi Analiz Populyatsii Obyknovennoi Polevki v Svyazi s Problemoi Izbiratel’nosti Otlova Gryzunov Sovami; Extended Abstract of the Dissertation for the Degree of Candidate of Science (Biology); Pemskii Gosudarstvennyi Universitat: Perm, Russia, 2002; p. 24. (In Russian) [Google Scholar]
- Johnstone, R.A. Female Preference for Symmetrical Males as a by-Product of Mate Recognition. Nature 1994, 372, 172–175. [Google Scholar] [CrossRef] [PubMed]
- Swaddle, J.P.; Cuthill, I.C. Preference for Symmetrical Males by Female Zebra Finches. Nature 1994, 367, 165–166. [Google Scholar] [CrossRef]
- Vol’pert, Y.L.; Shadrina, E.G. Small Mammals of the North-Eastern Siberia (Melkie Mlekopitayushchie Severo-Vostoka Sibiri); Nauka (Sib. Otdel.): Novosibirsk, Russia, 2002; p. 246, (In Russian). [Google Scholar] [CrossRef]
- Zakharov, V.M.; Baranov, A.S.; Borisov, V.I.; Valetskii, A.V.; Kryazheva, N.G.; Chistyakova, E.K.; Chubinishvili, A.T. Zdorov’e Sredy: Metodika Otsenki (Health of the Environment: Practical Evaluation); Tsentr Ekolog; Politiki Rossii: Moscow, Russia, 2000; p. 68. (In Russian) [Google Scholar]
- Shadrina, E.G.; Vol’pert, Y.L.; Danilov, V.A.; Shadrin, D.Y. Bioindication Impact of Mining on Terrestrial Ecosystems of the North (Morphogenetic Approach; Bioindikatsiya Vozdeistviya Gornodoby Vayushchei Promyshlennosti na Nazemnye Ekosistemy Severa (Morfogeneticheskii Podkhod); Nauka (Sib. Otdel.): Novosibirsk, Russia, 2003; p. 110, (In Russian). [Google Scholar] [CrossRef]
- Koshkina, T.V. Metod opredeleniya vozrasta ryzhikh polevok i opyt ego primeneniya. Russ. J. Zool. 1955, 34, 631–639. (In Russian) [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
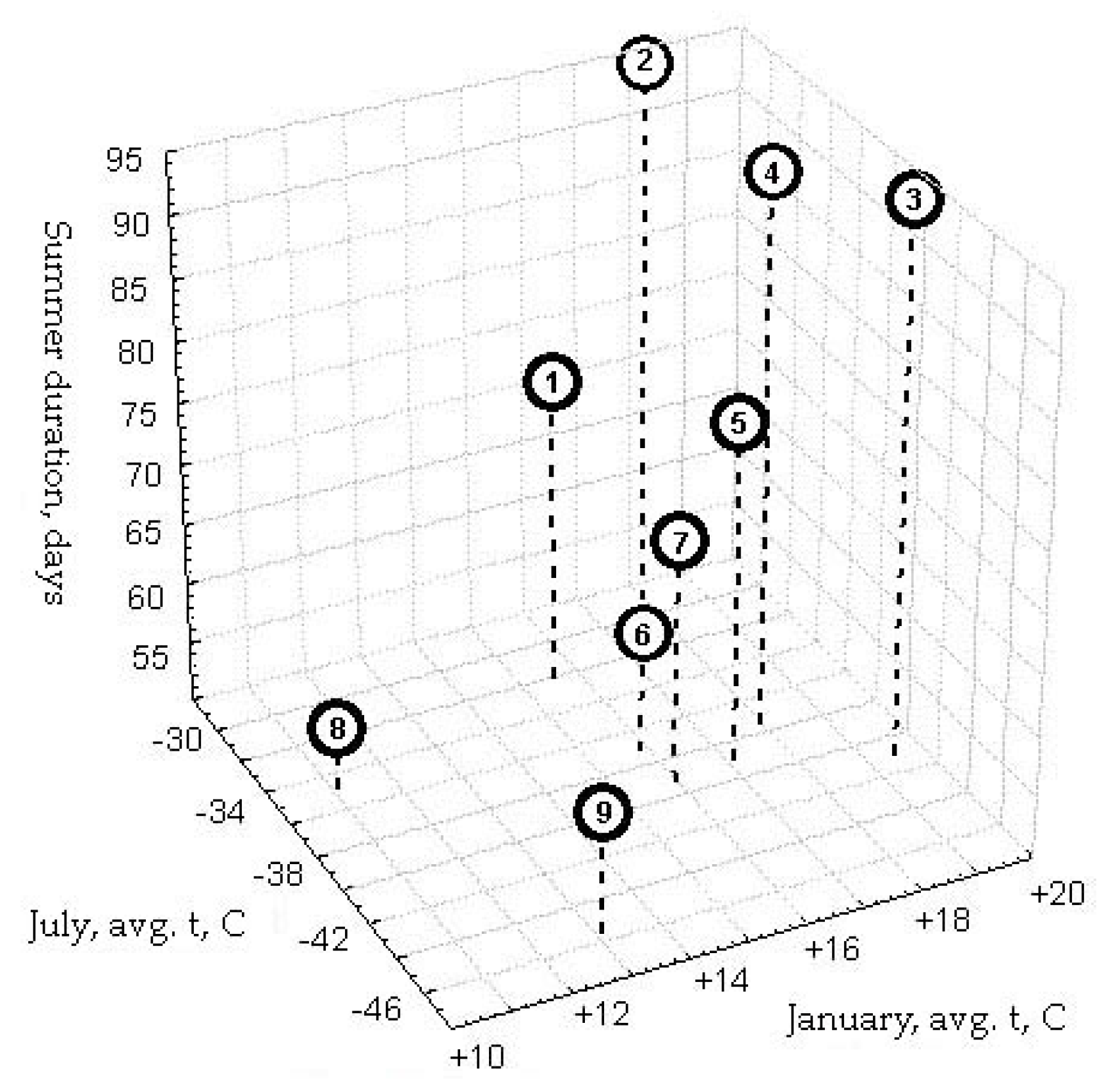
| No. | Point | Type of Vegetation | Climatic Zone | Summers, Period (Days) | Suitability for Small Mammals |
|---|---|---|---|---|---|
| 1 | Southern Yakutia | Middle taiga | Temperate, sharply continental climate | 75 | Near the optimum for the taiga species |
| 2 | Southern Yakutia | Middle taiga | 95 | ||
| 3 | Central Yakutia | Middle taiga | 95 | ||
| 4 | Western Yakutia | Middle taiga | 95 | ||
| 5 | Western Yakutia | Northern taiga | 78 | ||
| 6 | Western Yakutia | Tundra-forest | Subarctic climate | 60 | Northern periphery of the taiga species range |
| 7 | Northern Yakutia | Tundra-forest | 70 | ||
| 8 | Northeastern Yakutia | Tundra-forest | 55 | ||
| 9 | Central Verchoyanie | Mountain taiga | 60 | Ecological periphery of the taiga species range |
| No. | Region | Species | Study Goal * | n | The Number of: | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Ad | Juv | |||||||||
| G | S | A | ♂ | ♀ | ♂ | ♀ | ||||
| 1 | Southern Yakutia | Clethrionomys rutilus | + | + | 63 | 14 | 12 | 22 | 15 | |
| Cl. rufocanus | + | 7 | 1 | 1 | 2 | 3 | ||||
| Myopus schisticolor | + | + | + | 128 | 20 | 34 | 29 | 45 | ||
| Sorex caecutiens | + | 128 | 8 | 9 | 90 | 21 | ||||
| S. isodon | + | 12 | 1 | - | 8 | 3 | ||||
| S. daphaenodon | + | 6 | - | - | 4 | 2 | ||||
| 2 | Southern Yakutia | Clethrionomys rutilus | + | + | + | 125 | - | 1 | 65 | 59 |
| Microtus oeconomus | + | 53 | 11 | 6 | 21 | 15 | ||||
| Sorex caecutiens | + | 36 | 2 | - | 18 | 16 | ||||
| S. tundrensis | + | 16 | 1 | 1 | 5 | 9 | ||||
| 3 | Central Yakutia | Clethrionomys rutilus | + | + | 107 | 29 | 21 | 30 | 27 | |
| Microtus oeconomus | + | + | 48 | - | - | 28 | 20 | |||
| M. gregalis | + | 24 | - | - | 12 | 12 | ||||
| Sorex caecutiens | + | 12 | - | - | 7 | 5 | ||||
| S. tundrensis | + | 20 | - | - | 10 | 10 | ||||
| 4 | Western Yakutia | Clethrionomys rutilus | + | 16 | - | - | 8 | 6 | ||
| Microtus oeconomus | + | + | + | 180 | 28 | 23 | 74 | 55 | ||
| Microtus gregalis | + | 23 | - | - | 11 | 12 | ||||
| Sorex caecutiens | + | 18 | - | - | 10 | 8 | ||||
| S. tundrensis | + | 12 | - | - | 6 | 6 | ||||
| 5 | Western Yakutia | Clethrionomys rutilus | + | + | 51 | 5 | 4 | 21 | 21 | |
| Cl. rufocanus | + | 25 | - | - | 12 | 13 | ||||
| Microtus oeconomus | + | 30 | 1 | 2 | 13 | 12 | ||||
| Sorex caecutiens | + | + | 46 | 2 | - | 23 | 21 | |||
| S. tundrensis | + | + | 42 | - | - | 22 | 20 | |||
| S. daphaenodon | + | + | 49 | 1 | 1 | 25 | 22 | |||
| 6 | Western Yakutia | Clethrionomys rutilus | + | 28 | 2 | 1 | 15 | 10 | ||
| Cl. rufocanus | + | 14 | - | - | 8 | 6 | ||||
| Microtus oeconomus | + | 8 | - | - | 3 | 5 | ||||
| Sorex caecutiens | + | 12 | 1 | - | 8 | 3 | ||||
| 7 | Northern Yakutia | S. caecutiens | + | + | 39 | 1 | - | 24 | 14 | |
| S. tundrensis | + | 48 | - | - | 27 | 21 | ||||
| 8 | Northeastern Yakutia | Clethrionomys rutilus | + | + | + | 84 | 22 | 26 | 20 | 16 |
| Cl. rufocanus | + | 34 | 13 | 11 | 4 | 6 | ||||
| Myopus schisticolor | + | + | + | 173 | 72 | 75 | 10 | 16 | ||
| Microtus oeconomus | + | + | + | 208 | 53 | 37 | 61 | 57 | ||
| Sorex caecutiens | + | + | 145 | 8 | 3 | 78 | 56 | |||
| Sorex tundrensis | + | + | 62 | 3 | - | 35 | 24 | |||
| S. daphaenodon | + | 16 | - | - | 8 | 8 | ||||
| 9 | Central Verchoyanie | Clethrionomys rutilus | + | + | 68 | 10 | 17 | 20 | 21 | |
| Sorex caecutiens | + | 83 | 9 | 5 | 39 | 30 | ||||
| Species | Region | Age | OFAM (occurrence of the fluctuating asymmetry manifestations) | |||
|---|---|---|---|---|---|---|
| Males | Females | |||||
| n | M ± m | n | M ± m | |||
| Clethrinomys rutilus | Southern Yakutia | juv | 65 | 0.35 ± 0.01 | 59 | 0.35 ± 0.02 |
| Central Yakutia | ad | 29 | 0.25 ± 0.02 | 21 | 0.26 ± 0.02 | |
| juv | 30 | 0.31 ± 0.01 | 27 | 0.29 ± 0.02 | ||
| Western Yakutia | juv | 21 | 0.28 ± 0.03 | 21 | 0.34 ± 0.02 | |
| Northeastern Yakutia | ad | 22 | 0.34 ± 0.02 | 26 | 0.32 ± 0.03 | |
| Central Verchoyanye | juv | 20 | 0.38 ± 0.03 | 21 | 0.39 ± 0.03 | |
| Myopus schisticolor | Southern Yakutia | ad | 20 | 0.35 ± 0.01 | 34 | 0.34 ± 0.02 |
| juv | 29 | 0.36 ± 0.03 | 45 | 0.34 ± 0.03 | ||
| Northeastern Yakutia | ad | 57 | 0.36 ± 0.01 | 47 | 0.37 ± 0.02 | |
| Microtus oeconomus | Central Yakutia | juv | 28 | 0.27 ± 0.02 | 20 | 0.25 ± 0.03 |
| Western Yakutia | ad | 28 | 0.29 ± 0.02 | 23 | 0.30 ± 0.04 | |
| juv | 74 | 0.31 ± 0.03 | 55 | 0.32 ± 0.02 | ||
| Northeastern Yakutia | ad | 53 | 0.35 ± 0.02 | 37 | 0.34 ± 0.01 | |
| juv | 61 | 0.44 ± 0.04 | 57 | 0.41 ± 0.03 | ||
| Sorex caecutiens | Southern Yakutia | juv | 90 | 0.15 ± 0.01 | 21 | 0.16 ± 0.02 |
| Western Yakutia | juv | 23 | 0.15 ± 0.03 | 21 | 0.15 ± 0.04 | |
| Northeastern Yakutia | juv | 78 | 0.20 ± 0.03 | 56 | 0.18 ± 0.01 | |
| Central Verchoyanye | juv | 39 | 0.22 ± 0.03 | 30 | 0.20 ± 0.02 | |
| Sorex tundrensis | Western Yakutia | juv | 22 | 0.18 ± 0.03 | 20 | 0.20 ± 0.02 |
| Northeastern Yakutia | juv | 27 | 0.16 ± 0.02 | 21 | 0.19 ± 0.03 | |
| Sorex daphaenodon | Western Yakutia | juv | 25 | 0.18 ± 0.03 | 22 | 0.20 ± 0.01 |
| Region | Species | OFAM, in Age Group | t-Test, Significance of Difference | |||
|---|---|---|---|---|---|---|
| Juv | Ad | |||||
| n | M ± m | n | M ± m | |||
| Middle taiga | Clethrionomys rutilus | 57 | 0.29 ± 0.02 | 50 | 0.25 ± 0.02 | 0–1.46 * |
| Myopus schisticolor | 74 | 0.35 ± 0.02 | 54 | 0.35 ± 0.01 | ||
| Microtus oeconomus | 129 | 0.32 ± 0.02 | 51 | 0.29 ± 0.02 | ||
| Sorex caecutiens | 56 | 0.17 ± 0.02 | 11 | 0.15 ± 0.02 | ||
| Mountain taiga | Clethrionomys rutilus | 41 | 0.38 ± 0.01 | 27 | 0,29 ± 0,02 | 4.02 (p < 0.01) |
| Sorex caecutiens | 69 | 0.21 ± 0.01 | 14 | 0.17 ± 0.02 | 1.79 * | |
| Forest-tundra | Clethrionomys rutilus | 36 | 0.37 ± 0.01 | 48 | 0.33 ± 0.01 | 2.83 (p < 0.01) |
| Clethrionomys rufocanus | 10 | 0.43 ± 0.03 | 24 | 0.37 ± 0.02 | 1.66 * | |
| Myopus schisticolor | 26 | 0.45 ± 0.01 | 147 | 0.37 ± 0.03 | 2.53 (p < 0.05) | |
| Microtus oeconomus | 118 | 0.42 ± 0.02 | 90 | 0.35 ± 0.02 | 2.47 (p < 0.05) | |
| Sorex caecutiens | 134 | 0.20 ± 0.01 | 11 | 0.17 ± 0.01 | 2.12 (p < 0.05) | |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shadrina, E.; Vol’pert, Y. Fluctuating Asymmetry of Craniological Features of Small Mammals as a Reflection of Heterogeneity of Natural Populations. Symmetry 2016, 8, 142. https://doi.org/10.3390/sym8120142
Shadrina E, Vol’pert Y. Fluctuating Asymmetry of Craniological Features of Small Mammals as a Reflection of Heterogeneity of Natural Populations. Symmetry. 2016; 8(12):142. https://doi.org/10.3390/sym8120142
Chicago/Turabian StyleShadrina, Elena, and Yakov Vol’pert. 2016. "Fluctuating Asymmetry of Craniological Features of Small Mammals as a Reflection of Heterogeneity of Natural Populations" Symmetry 8, no. 12: 142. https://doi.org/10.3390/sym8120142
APA StyleShadrina, E., & Vol’pert, Y. (2016). Fluctuating Asymmetry of Craniological Features of Small Mammals as a Reflection of Heterogeneity of Natural Populations. Symmetry, 8(12), 142. https://doi.org/10.3390/sym8120142