
Multiscale Microbial Preservation and Biogeochemical Signals in a Modern Hot-Spring Siliceous Sinter Rich in CO2 Emissions, Krýsuvík Geothermal Field, Iceland



Abstract
:1. Introduction
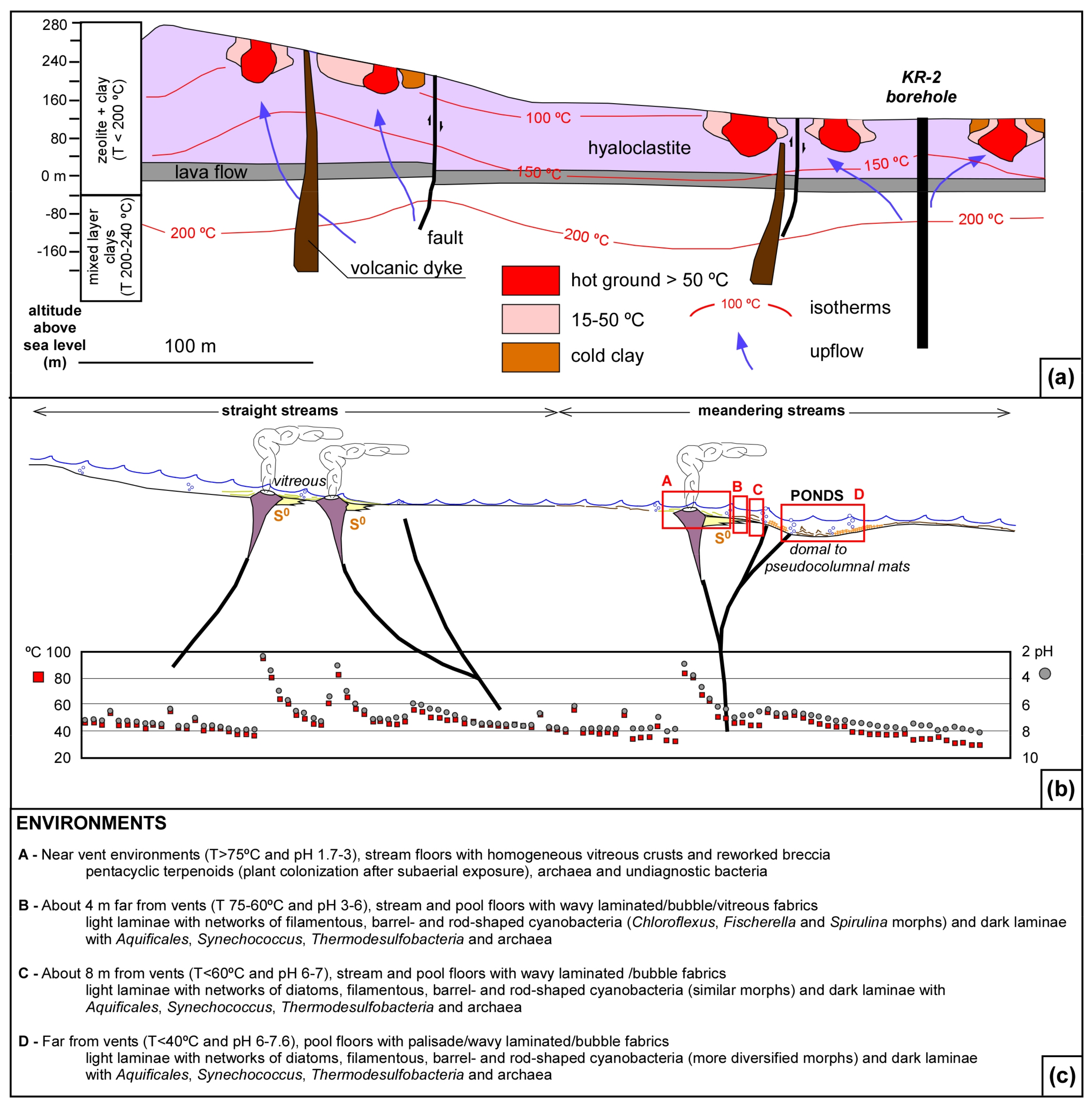
2. Field Setting
3. Material and Methods
3.1. Mineralogy
3.2. Lipid Biomarkers
4. Multiscale Structures of Sinter Samples
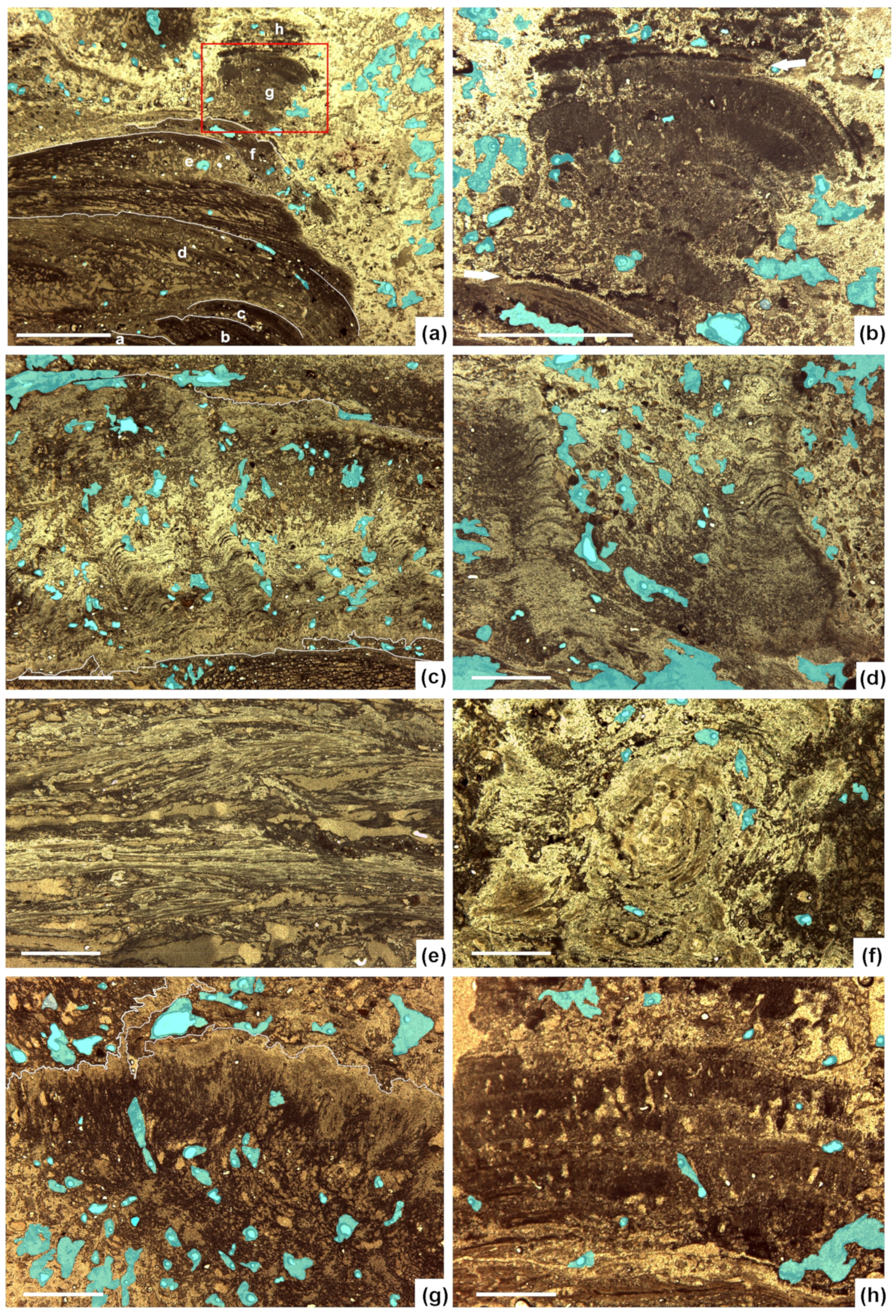
4.1. Sinter Fabrics (Mesoscale 1–100 mm)
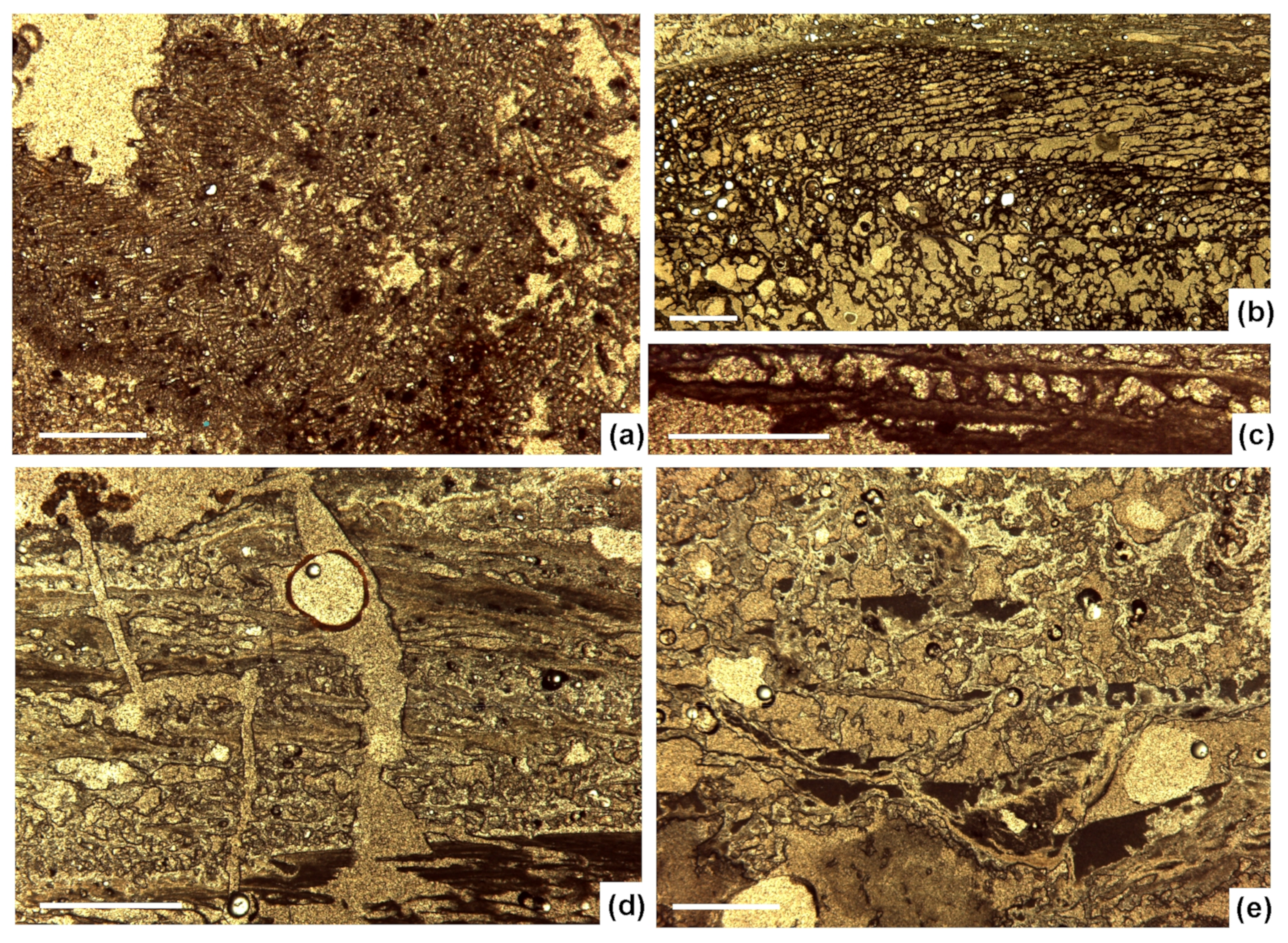
4.2. Sinter Textures (Microscale 10–1000 µm)
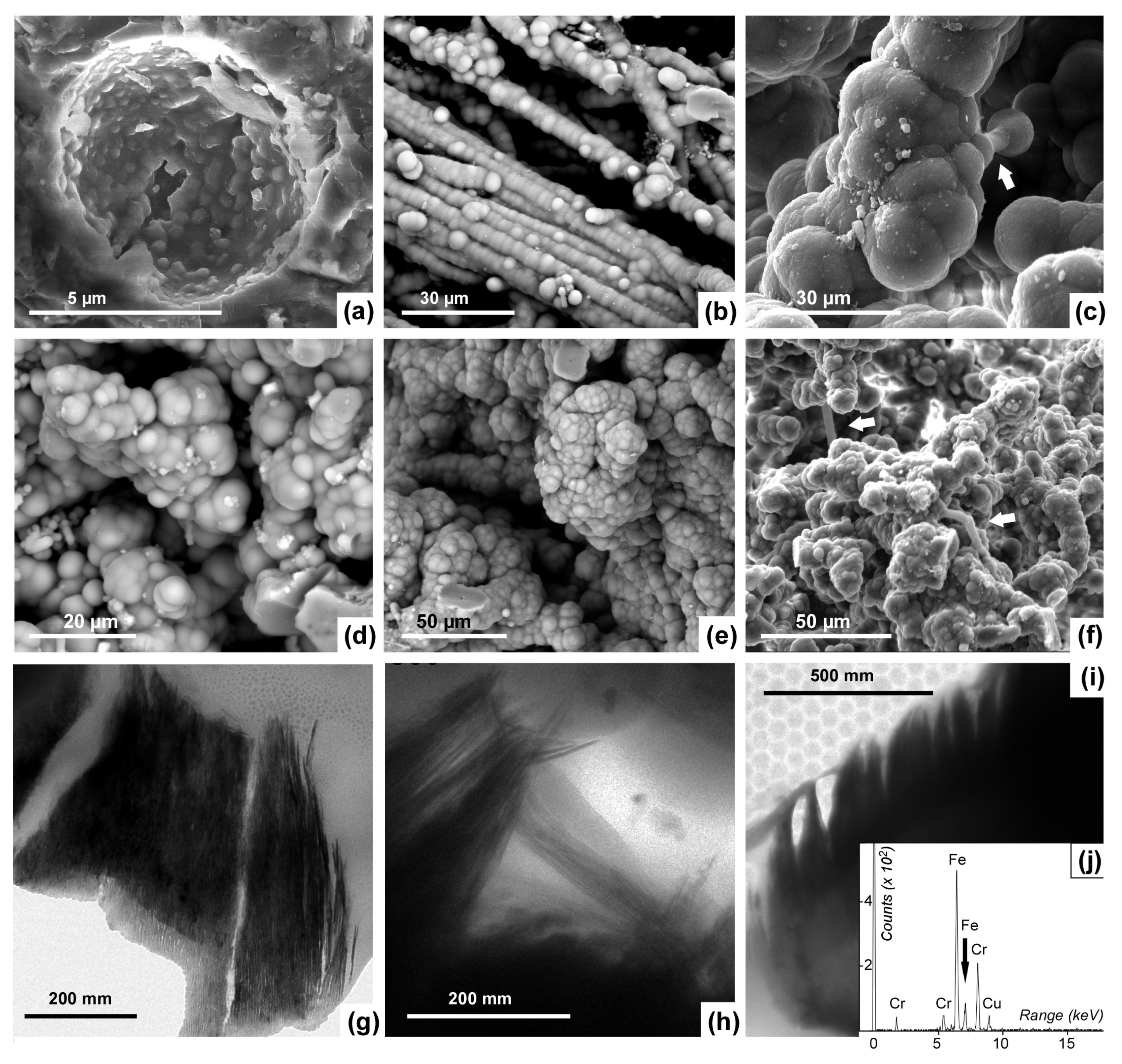
4.3. Silicified Microbes (Scale < 10 µm)
5. Iron-Rich Biominerals and Inferred Bacteria
6. Preservational Constraints on Silicified Microbes
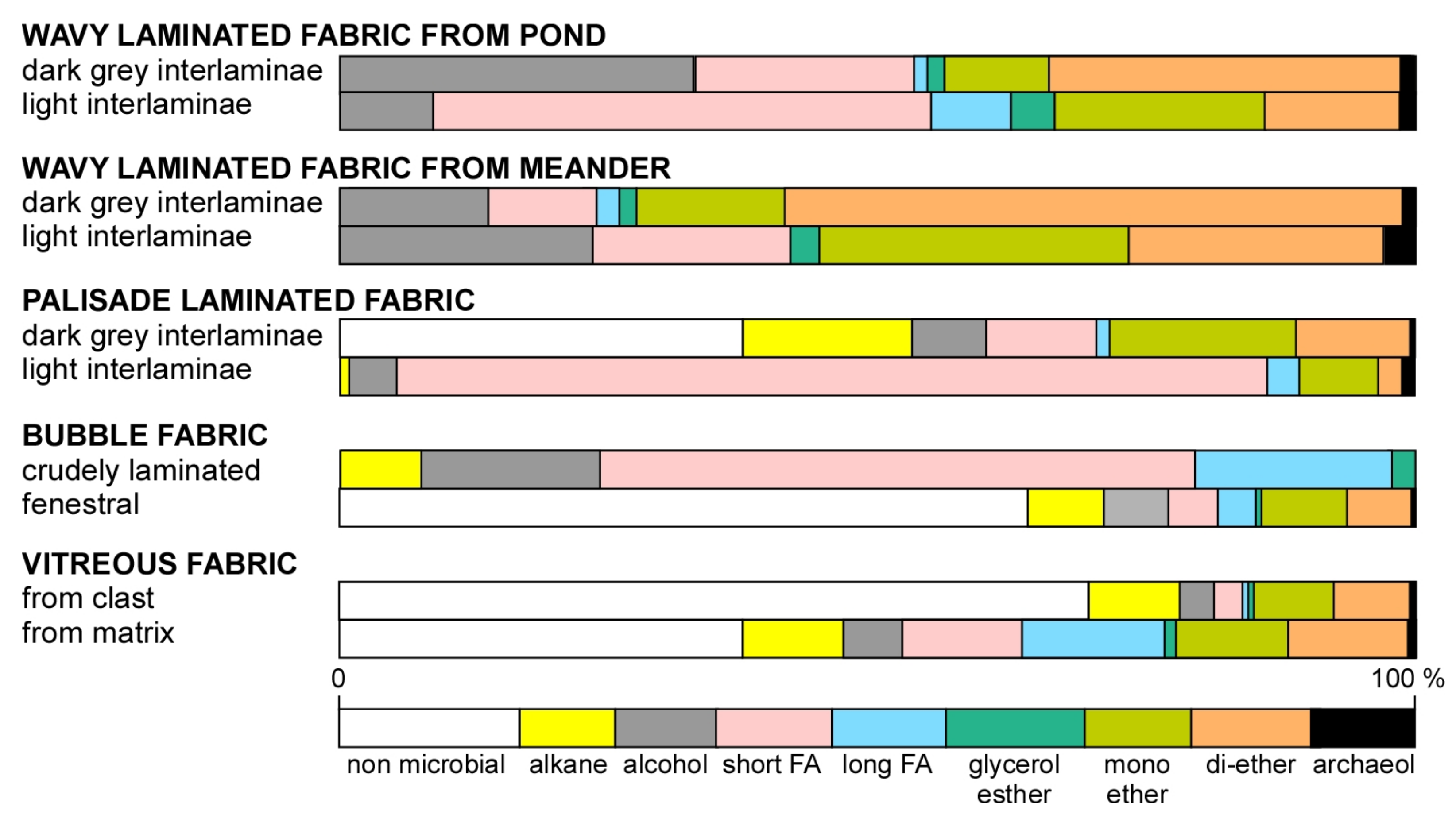
7. Biomarker Analysis
8. Carbon Isotopic Signatures of Lipid Biomarkers
9. Environmental Distribution
10. Astrobiological Implications
11. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Allwood, A.C.; Walter, M.R.; Kamber, B.S.; Marshall, C.P.; Burch, I.W. Stromatolite reef from the Early Archaean era of Australia. Nat. Cell Biol. 2006, 441, 714–718. [Google Scholar] [CrossRef] [PubMed]
- Noffke, N.; Eriksson, K.A.; Hazen, R.M.; Simpson, E.L. A new window into Early Archean life: Microbial mats in Earth’s oldest siliciclastic tidal deposits (3.2 Ga Moodies Group, South Africa). Geology 2006, 34, 253. [Google Scholar] [CrossRef]
- Schopf, J.W.; Kudryavtsev, A.B.; Czaja, A.D.; Tripathi, A.B. Evidence of Archean life: Stromatolites and microfossils. Precambrian Res. 2007, 158, 141–155. [Google Scholar] [CrossRef]
- Djokic, T.; Van Kranendonk, M.J.; Campbell, K.A.; Walter, M.R.; Ward, C.R. Earliest signs of life on land preserved in ca. 3.5 Ga hot spring deposits. Nat. Commun. 2017, 8, 15263. [Google Scholar] [CrossRef] [Green Version]
- Dodd, M.S.; Papineau, M.S.D.D.; Grenne, T.; Slack, J.F.; Rittner, M.S.D.D.P.M.; Pirajno, F.; O’Neil, J.; Little, C.T.S. Evidence for early life in Earth’s oldest hydrothermal vent precipitates. Nat. Cell Biol. 2017, 543, 60–64. [Google Scholar] [CrossRef]
- Squyres, S.W.; Arvidson, R.E.; Ruff, S.; Gellert, R.; Morris, R.V.; Ming, D.W.; Crumpler, L.; Farmer, J.D.; Marais, D.J.D.; Yen, A.; et al. Detection of Silica-Rich Deposits on Mars. Science 2008, 320, 1063–1067. [Google Scholar] [CrossRef]
- Ruff, S.W.; Farmer, J.D.; Calvin, W.M.; Herkenhoff, K.E.; Johnson, J.R.; Morris, R.V.; Rice, M.S.; Arvidson, R.E.; Bell, J.F.; Christensen, P.R.; et al. Characteristics, distribution, origin, and significance of opaline silica observed by the Spirit rover in Gusev crater, Mars. J. Geophys. Res. Space Phys. 2011, 116. [Google Scholar] [CrossRef]
- Damer, B.; Deamer, D. Coupled Phases and Combinatorial Selection in Fluctuating Hydrothermal Pools: A Scenario to Guide Experimental Approaches to the Origin of Cellular Life. Life 2015, 5, 872–887. [Google Scholar] [CrossRef] [Green Version]
- Ruff, S.W.; Farmer, J.D. Silica deposits on Mars with features resembling hot spring biosignatures at El Tatio in Chile. Nat. Commun. 2016, 7, 13554. [Google Scholar] [CrossRef] [PubMed]
- Konhauser, K.O.; Jones, B.; Reysenbach, A.-L.; Renaut, R.W. Hot spring sinters: Keys to understanding Earth’s earliest life forms. Can. J. Earth Sci. 2003, 40, 1713–1724. [Google Scholar] [CrossRef]
- Gangidine, A.; Havig, J.R.; Fike, D.A.; Jones, C.; Hamilton, T.L.; Czaja, A.D. Trace Element Concentrations in Hydrothermal Silica Deposits as a Potential Biosignature. Astrobiology 2020, 20, 525–536. [Google Scholar] [CrossRef] [Green Version]
- Gangidine, A.; Walter, M.R.; Havig, J.R.; Jones, C.; Sturmer, D.M.; Czaja, A.D. Trace Element Concentrations Associated with Mid-Paleozoic Microfossils as Biosignatures to Aid in the Search for Life. Life 2021, 11, 142. [Google Scholar] [CrossRef]
- Lindahl, T. Instability and decay of the primary structure of DNA. Nat. Cell Biol. 1993, 362, 709–715. [Google Scholar] [CrossRef]
- Jahnke, L.L.; Eder, W.; Huber, R.; Hope, J.M.; Hinrichs, K.-U.; Hayes, J.M.; Marais, D.J.D.; Cady, S.L.; Summons, R.E. Signature Lipids and Stable Carbon Isotope Analyses of Octopus Spring Hyperthermophilic Communities Compared with Those ofAquificales Representatives. Appl. Environ. Microbiol. 2001, 67, 5179–5189. [Google Scholar] [CrossRef] [Green Version]
- Pancost, R.D.; Pressley, S.; Coleman, J.M.; Benning, L.G.; Mountain, B.W. Lipid biomolecules in silica sinters: Indicators of microbial biodiversity. Environ. Microbiol. 2005, 7, 66–77. [Google Scholar] [CrossRef]
- Kaur, G.; Mountain, B.W.; Pancost, R.D. Microbial membrane lipids in active and inactive sinters from Champagne Pool, New Zealand: Elucidating past geothermal chemistry and microbiology. Org. Geochem. 2008, 39, 1024–1028. [Google Scholar] [CrossRef]
- Kaur, G.; Mountain, B.W.; Stott, M.B.; Hopmans, E.C.; Pancost, R.D. Temperature and pH control on lipid composition of silica sinters from diverse hot springs in the Taupo Volcanic Zone, New Zealand. Extremophiles 2015, 19, 327–344. [Google Scholar] [CrossRef] [PubMed]
- Schubotz, F.; Meyer-Dombard, D.R.; Bradley, A.S.; Fredricks, H.F.; Hinrichs, K.-U.; Shock, E.L.; Summons, R.E. Spatial and temporal variability of biomarkers and microbial diversity reveal metabolic and community flexibility in Streamer Biofilm Communities in the Lower Geyser Basin, Yellowstone National Park. Geobiology 2013, 11, 549–569. [Google Scholar] [CrossRef]
- Mata, S.A.; Harwood, C.L.; Corsetti, F.A.; Stork, N.J.; Eilers, K.; Berelson, W.M.; Spear, J.R. Influence of Gas Production and Filament Orientation on Stromatolite Microfabric. Palaios 2012, 27, 206–219. [Google Scholar] [CrossRef]
- Pepe-Ranney, C.; Berelson, W.M.; Corsetti, F.A.; Treants, M.; Spear, J.R. Cyanobacterial construction of hot spring siliceous stromatolites in Yellowstone National Park. Environ. Microbiol. 2012, 14, 1182–1197. [Google Scholar] [CrossRef] [PubMed]
- Bickford, M.E. The Web of Geological Sciences: Advances, Impacts, and Interactions. In Geological Society of America Special Papers; Geological Society of America: Boulder, CO, USA, 2013. [Google Scholar]
- Phoenix, V.R.; Adams, D.G.; Konhauser, K.O. Cyanobacterial viability during hydrothermal biomineralisation. Chem. Geol. 2000, 169, 329–338. [Google Scholar] [CrossRef]
- Yee, N.; Phoenix, V.R.; Konhauser, K.O.; Benning, L.G.; Ferris, F. The effect of cyanobacteria on silica precipitation at neutral pH: Implications for bacterial silicification in geothermal hot springs. Chem. Geol. 2003, 199, 83–90. [Google Scholar] [CrossRef]
- Menzel, P.; Gudbergsdóttir, S.R.; Rike, A.G.; Lin, L.; Zhang, Q.; Contursi, P.; Moracci, M.; Kristjansson, J.K.; Bolduc, B.; Gavrilov, S.; et al. Comparative Metagenomics of Eight Geographically Remote Terrestrial Hot Springs. Microb. Ecol. 2015, 70, 411–424. [Google Scholar] [CrossRef]
- Tobler, D.J.; Stefánsson, A.; Benning, L.G. In-situgrown silica sinters in Icelandic geothermal areas. Geobiology 2008, 6, 481–502. [Google Scholar] [CrossRef]
- Tobler, D.J.; Benning, L.G. Bacterial diversity in five Icelandic geothermal waters: Temperature and sinter growth rate effects. Extremophiles 2011, 15, 473–485. [Google Scholar] [CrossRef] [PubMed]
- Hersir, G.P.; Árnason, K.; Vilhjálmsson, A.M.; Saemundsson, K.; Ágústsdóttir, Þ.; Friðleifsson, G. Ómar Krýsuvík high temperature geothermal area in SW Iceland: Geological setting and 3D inversion of magnetotelluric (MT) resistivity data. J. Volcanol. Geotherm. Res. 2020, 391, 106500. [Google Scholar] [CrossRef]
- Franzson, H.; Zierenberg, R.A.; Schiffman, P. Chemical transport in geothermal systems in Iceland. J. Volcanol. Geotherm. Res. 2008, 173, 217–229. [Google Scholar] [CrossRef]
- Markússon, S.H.; Stefánsson, A. Geothermal surface alteration of basalts, Krýsuvík Iceland—Alteration mineralogy, water chemistry and the effects of acid supply on the alteration process. J. Volcanol. Geotherm. Res. 2011, 206, 46–59. [Google Scholar] [CrossRef]
- Mawejje, P. Geothermal exploration and geological mapping at Seltún, Krýsuvík geothermal field, Reykjanes Peninsula, SW-Iceland. Geotherm. Train. Programme Rep. 2007, 12, 1–276. [Google Scholar]
- Ármannsson, H. The fluid geochemistry of Icelandic high temperature geothermal areas. Appl. Geochem. 2016, 66, 14–64. [Google Scholar] [CrossRef]
- Gudjónsdóttir, S.R.; Ilyinskaya, E.; Hreinsdóttir, S.; Bergsson, B.; Pfeffer, M.A.; Michalczewska, K.; Aiuppa, A.; Óladóttir, A.A. Gas emissions and crustal deformation from the Krýsuvík high temperature geothermal system, Iceland. J. Volcanol. Geotherm. Res. 2020, 391, 106350. [Google Scholar] [CrossRef] [Green Version]
- Chung, A.P.; Rainey, F.A.; Valente, M.; Nobre, M.F.; Da Costa, M.S. Thermus igniterrae sp. nov. and Thermus antranikianii sp. nov., two new species from Iceland. Int. J. Syst. Evol. Microbiol. 2000, 50, 209–217. [Google Scholar] [CrossRef]
- Hjorleifsdottir, S.; Skirnisdottir, S.; Hreggvidsson, G.; Holst, O.; Kristjansson, J. Species Composition of Cultivated and Noncultivated Bacteria from Short Filaments in an Icelandic Hot Spring at 88 °C. Microb. Ecol. 2001, 42, 117–125. [Google Scholar] [CrossRef]
- Kvist, T.; Ahring, B.K.; Westermann, P. Archaeal diversity in Icelandic hot springs. FEMS Microbiol. Ecol. 2007, 59, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Schultze-Lam, S.; Ferris, F.G.; Konhauser, K.O.; Wiese, R.G. In situ silicification of an Icelandic hot spring microbial mat: Implications for microfossil formation. Can. J. Earth Sci. 1995, 32, 2021–2026. [Google Scholar] [CrossRef] [Green Version]
- Konhauser, K.O.; Ferris, F.G. Diversity of iron and silica precipitation by microbial mats in hydrothermal waters, Iceland: Implications for Precambrian iron formations. Geology 1996, 24, 323–326. [Google Scholar] [CrossRef] [Green Version]
- Konhauser, K.O.; Phoenix, V.R.; Bottrell, S.H.; Adams, D.G.; Head, I.M. Microbial-silica interactions in Icelandic hot spring sinter: Possible analogues for some Precambrian siliceous stromatolites. Sedimentology 2001, 48, 415–433. [Google Scholar] [CrossRef]
- Konhauser, K.O.; Jones, B.; Phoenix, V.R.; Ferris, G.; Renaut, R.W. The Microbial Role in Hot Spring Silicification. AMBIO 2004, 33, 552–558. [Google Scholar] [CrossRef]
- Petursdottir, S.K.; Hreggvidsson, G.O.; Da Costa, M.S.; Kristjansson, J.K. Genetic diversity analysis of Rhodothermus reflects geographical origin of the isolates. Extremophiles 2000, 4, 267–274. [Google Scholar] [CrossRef]
- Hreggvidsson, G.O.; Skirnisdottir, S.; Smit, B.; Hjörleifsdottir, S.; Marteinsson, V.T.; Petursdottir, S.; Kristjansson, J.K. Polyphasic analysis of Thermus isolates from geothermal areas in Iceland. Extremophiles 2006, 10, 563–575. [Google Scholar] [CrossRef]
- Sæmundsson, K.; Sigurgeirsson, M.Á.; Friðleifsson, G. Ómar Geology and structure of the Reykjanes volcanic system, Iceland. J. Volcanol. Geotherm. Res. 2020, 391, 106501. [Google Scholar] [CrossRef]
- Jóhannesson, H.; Sæmundsson, K. Geological Map of Iceland, 1:500,000; Icelandic Institute of Natural History: Reykjavík, Iceland, 1998. [Google Scholar]
- Pałgan, D.; Devey, C.W.; Yeo, I.A. Volcanism and hydrothermalism on a hotspot–influenced ridge: Comparing Reykjanes Peninsula and Reykjanes Ridge, Iceland. J. Volcanol. Geotherm. Res. 2017, 348, 62–81. [Google Scholar] [CrossRef] [Green Version]
- Fridriksson, T.; Kristjánsson, B.R.; Ármannsson, H.; Margrétardóttir, E.; Ólafsdóttir, S.; Chiodini, G. CO2 emissions and heat flow through soil, fumaroles, and steam heated mud pools at the Reykjanes geothermal area, SW Iceland. Appl. Geochem. 2006, 21, 1551–1569. [Google Scholar] [CrossRef]
- Ármannsson, H.; Fridriksson, T.; Kristjánsson, B.R. CO2 emissions from geothermal power plants and natural geothermal activity in Iceland. Geothermics 2005, 34, 286–296. [Google Scholar] [CrossRef]
- Stefánsson, A.; Hilton, D.R.; Sveinbjörnsdóttir, Á.E.; Torssander, P.; Heinemeier, J.; Barnes, J.D.; Ono, S.; Halldórsson, S.A.; Fiebig, J.; Arnórsson, S. Isotope systematics of Icelandic thermal fluids. J. Volcanol. Geotherm. Res. 2017, 337, 146–164. [Google Scholar] [CrossRef]
- Konhauser, K.O.; Phoenix, V.R.; Bottrell, S.H.; Adams, D.G.; Head, I.M. Microbial-silica interactions in modern hot spring sinter. In Geochemistry of the Earth Surface; Ármannsson, H., Ed.; Balkema: Rotterdam, The Netherlands, 1999; pp. 263–266. [Google Scholar]
- Herdianita, N.R.; Browne, P.R.L.; Rodgers, K.A.; Campbell, K.A. Mineralogical and textural changes accompanying ageing of silica sinter. Miner. Deposita 2000, 35, 48–62. [Google Scholar] [CrossRef]
- Lynne, B.Y.; Campbell, K.A. Diagenetic transformations (opal-A to quartz) of low- and mid-temperature microbial textures in siliceous hot-spring deposits, Taupo Volcanic Zone, New Zealand. Can. J. Earth Sci. 2003, 40, 1679–1696. [Google Scholar] [CrossRef]
- Lynne, B.Y.; Campbell, K.A. Morphologic and Mineralogic Transitions from Opal-A to Opal-CT in Low-Temperature Siliceous Sinter Diagenesis, Taupo Volcanic Zone, New Zealand. J. Sediment. Res. 2004, 74, 561–579. [Google Scholar] [CrossRef]
- Guidry, S.A.; Chafetz, H.S. Depositional Facies and Diagenetic Alteration in a Relict Siliceous Hot-Spring Accumulation: Examples from Yellowstone National Park, U.S.A. J. Sediment. Res. 2003, 73, 806–823. [Google Scholar] [CrossRef]
- Guidry, S.A.; Chafetz, H.S. Siliceous shrubs in hot springs from Yellowstone National Park, Wyoming, U.S.A. Can. J. Earth Sci. 2003, 40, 1571–1583. [Google Scholar] [CrossRef]
- Lynne, B.Y.; Campbell, K.A.; Moore, J.; Browne, P. Diagenesis of 1900-year-old siliceous sinter (opal-A to quartz) at Opal Mound, Roosevelt Hot Springs, Utah, U.S.A. Sediment. Geol. 2005, 179, 249–278. [Google Scholar] [CrossRef]
- Jones, B.; Renaut, R.W. Microstructural changes accompanying the opal-A to opal-CT transition: New evidence from the siliceous sinters of Geysir, Haukadalur, Iceland. Sedimentology 2007, 54, 921–948. [Google Scholar] [CrossRef]
- Lynne, B.Y.; Campbell, K.A.; James, B.J.; Browne, P.R.L.; Moore, J. Tracking crystallinity in siliceous hot-spring deposits. Am. J. Sci. 2007, 307, 612–641. [Google Scholar] [CrossRef]
- Brocks, J.J.; Buick, R.; Logan, G.A.; Summons, R.E. Composition and syngeneity of molecular fossils from the 2.78 to 2.45 billion-year-old Mount Bruce Supergroup, Pilbara Craton, Western Australia. Geochim. Cosmochim. Acta 2003, 67, 4289–4319. [Google Scholar] [CrossRef]
- Brocks, J.J.; Buick, R.; Summons, R.E.; Logan, G.A. A reconstruction of Archean biological diversity based on molecular fossils from the 2.78 to 2.45 billion-year-old Mount Bruce Supergroup, Hamersley Basin, Western Australia. Geochim. Cosmochim. Acta 2003, 67, 4321–4335. [Google Scholar] [CrossRef]
- Jones, B.; Renaut, R.W.; Rosen, M.R. Biogenicity of Silica Precipitation around Geysers and Hot-Spring Vents, North Island, New Zealand. J. Sediment. Res. 1997, 67, 88–104. [Google Scholar] [CrossRef]
- Jones, B.; Renaut, R.W.; Rosen, M.R. Stromatolites Forming in Acidic Hot-Spring Waters, North Island, New Zealand. Palaios 2000, 15, 450–475. [Google Scholar] [CrossRef]
- Jones, B.; Renaut, R.W.; Rosen, M.R. Silicified Microbes in a Geyser Mound: The Enigma of Low-Temperature Cyanobacteria in a High-Temperature Setting. PALAIOS 2003, 18, 87–109. [Google Scholar] [CrossRef]
- Jones, B.; De Ronde, C.E.J.; Renaut, R.W.; Owen, R.B. Siliceous sublacustrine spring deposits around hydrothermal vents in Lake Taupo, New Zealand. J. Geol. Soc. 2007, 164, 227–242. [Google Scholar] [CrossRef]
- Campbell, K.A.; Sannazzaro, K.; Rodgers, K.; Herdianita, N.; Browne, P. Sedimentary Facies and Mineralogy of the Late Pleistocene Umukuri Silica Sinter, Taupo Volcanic Zone, New Zealand. J. Sediment. Res. 2001, 71, 727–746. [Google Scholar] [CrossRef]
- Hinman, N.W.; Walter, M.R. Textural Preservation in Siliceous Hot Spring Deposits During Early Diagenesis: Examples from Yellowstone National Park and Nevada, USA. J. Sediment. Res. 2005, 75, 200–215. [Google Scholar] [CrossRef] [Green Version]
- Schinteie, R.; Campbell, K.A.; Browne, P.R.L. Microfacies of stromatolitic sinter from acid-sulphate-chloride springs at Parariki Stream, Rotokawa Geothermal Field, New Zealand. Palaeontol. Electron. 2007, 10, 33. [Google Scholar]
- Campbell, K.A.; Lynne, B.Y.; Handley, K.M.; Jordan, S.; Farmer, J.D.; Guido, D.M.; Foucher, F.; Turner, S.; Perry, R.S. Tracing Biosignature Preservation of Geothermally Silicified Microbial Textures into the Geological Record. Astrobiology 2015, 15, 858–882. [Google Scholar] [CrossRef]
- Wilmeth, D.T.; Corsetti, F.A.; Bisenic, N.; Dornbos, S.Q.; Oji, T.; Gonchigdorj, S. Punctuated Growth of Microbial Cones within Early Cambrian Oncoids, Bayan Gol Formation, Western Mongolia. Palaios 2015, 30, 836–845. [Google Scholar] [CrossRef]
- Hamilton, A.R.; Campbell, K.A.; Guido, D.M. Atlas of siliceous hot spring deposits (sinter) and other silicified surface manifestations in epithermal environments. GNS Sci. Rep. 2019, 1–56. [Google Scholar] [CrossRef]
- Jones, B.; Renaut, R.W.; Konhauser, K.O. Genesis of large siliceous stromatolites at Frying Pan Lake, Waimangu geothermal field, North Island, New Zealand. Sedimentology 2005, 52, 1229–1252. [Google Scholar] [CrossRef]
- Gong, J.; Myers, K.D.; Munoz-Saez, C.; Homann, M.; Rouillard, J.; Wirth, R.; Schreiber, A.; Van Zuilen, M.A. Formation and Preservation of Microbial Palisade Fabric in Silica Deposits from El Tatio, Chile. Astrobiology 2020, 20, 500–524. [Google Scholar] [CrossRef] [PubMed]
- Guidry, S.A.; Chafetz, H.S. Anatomy of siliceous hot springs: Examples from Yellowstone National Park, Wyoming, USA. Sediment. Geol. 2003, 157, 71–106. [Google Scholar] [CrossRef]
- Lynne, B.Y. Mapping vent to distal-apron hot spring paleo-flow pathways using siliceous sinter architecture. Geothermics 2012, 43, 3–24. [Google Scholar] [CrossRef]
- Ward, D.M.; Castenholz, R.W. Cyanobacteria in geothermal habitats. In Ecology of Cyanobacteria: Their Diversity in Time and Space; Whitton, B.A., Potts, M., Eds.; Kluwer: Dordrecht, The Netherlands, 2000; pp. 37–59. [Google Scholar]
- Doemel, W.N.; Brock, T.D. The Physiological Ecology of Cyanidium caldarium. J. Gen. Microbiol. 1971, 67, 17–32. [Google Scholar] [CrossRef] [Green Version]
- Hoiczyk, E. Structural and Biochemical Analysis of the Sheath of Phormidium uncinatum. J. Bacteriol. 1998, 180, 3923–3932. [Google Scholar] [CrossRef] [Green Version]
- Jones, B.; Renaut, R.W.; Rosen, M.R. Vertical Zonation of Biota in Microstromatolites Associated with Hot Springs, North Island, New Zealand. PALAIOS 1997, 12, 220. [Google Scholar] [CrossRef]
- Huber, R.; Eder, W.; Heldwein, S.; Wanner, G.; Huber, H.; Rachel, R.; Stetter, K.O. Thermocrinis ruber gen. nov., sp. nov., a Pink-Filament-Forming Hyperthermophilic Bacterium Isolated from Yellowstone National Park. Appl. Environ. Microbiol. 1998, 64, 3576–3583. [Google Scholar] [CrossRef] [Green Version]
- Inoue, N.; Emi, T.; Yamane, Y.; Kashino, Y.; Koike, H.; Satoh, K. Effects of High-Temperature Treatments on a Thermophilic Cyanobacterium Synechococcus vulcanus. Plant Cell Physiol. 2000, 41, 515–522. [Google Scholar] [CrossRef] [PubMed]
- Ciniglia, C.; Yoon, H.S.; Pollio, A.; Pinto, G.; Bhattacharya, D. Hidden biodiversity of the extremophilic Cyanidiales red algae. Mol. Ecol. 2004, 13, 1827–1838. [Google Scholar] [CrossRef]
- LaLonde, S.V.; Konhauser, K.O.; Reysenbach, A.-L.; Ferris, F.G. The experimental silicification of Aquificales and their role in hot spring sinter formation. Geobiology 2005, 3, 41–52. [Google Scholar] [CrossRef]
- Inagaki, F.; Hayashi, S.; Doi, K.; Motomura, Y.; Izawa, E.; Ogata, S. Microbial participation in the formation of siliceous deposits from geothermal water and analysis of the extremely thermophilic bacterial community. FEMS Microbiol. Ecol. 1997, 24, 41–48. [Google Scholar] [CrossRef]
- Yoon, H.S.; Ciniglia, C.; Wu, M.; Comeron, J.M.; Pinto, G.; Pollio, A.; Bhattacharya, D. Establishment of endolithic populations of extremophilic Cyanidiales (Rhodophyta). BMC Evol. Biol. 2006, 6, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flores, G.E.; Liu, Y.; Ferrera, I.; Beveridge, T.J.; Reysenbach, A.-L. Sulfurihydrogenibium kristjanssonii sp. nov., a hydrogen- and sulfur-oxidizing thermophile isolated from a terrestrial Icelandic hot spring. Int. J. Syst. Evol. Microbiol. 2008, 58, 1153–1158. [Google Scholar] [CrossRef] [PubMed]
- Reeb, V.; Bhattacharya, D. The Thermo-Acidophilic Cyanidiophyceae (Cyanidiales). In Red Algae in the Genomic Age; Springer International Publishing: Dordrecht, The Netherlands, 2010; Volume 13, pp. 409–426. [Google Scholar]
- Hugo, R.C.; Cady, S.L.; Smythe, W. The Role of Extracellular Polymeric Substances in the Silicification of Calothrix: Evidence from Microbial Mat Communities in Hot Springs at Yellowstone National Park, USA. Geomicrobiology 2011, 28, 667–675. [Google Scholar] [CrossRef]
- Skorupa, D.; Reeb, V.; Castenholz, R.; Bhattacharya, D.; McDermott, T. Cyanidiales diversity in Yellowstone National Park. Lett. Appl. Microbiol. 2013, 57, 459–466. [Google Scholar] [CrossRef] [Green Version]
- Smythe, W.F.; McAllister, S.M.; Hager, K.W.; Hager, K.R.; Tebo, B.M.; Moyer, C.L. Silica Biomineralization of Calothrix-Dominated Biofacies from Queen’s Laundry Hot-Spring, Yellowstone National Park, USA. Front. Environ. Sci. 2016, 4, 4. [Google Scholar] [CrossRef] [Green Version]
- Phoenix, V.R.; Martinez, R.E.; Konhauser, K.O.; Ferris, F.G. Characterization and Implications of the Cell Surface Reactivity of Calothrix sp. Strain KC97. Appl. Environ. Microbiol. 2002, 68, 4827–4834. [Google Scholar] [CrossRef] [Green Version]
- Takacs, C.D.; Ehringer, M.; Favre, R.; Cermola, M.; Eggertsson, G.; Palsdottir, A.; Reysenbach, A.-L.; Favre, R. Phylogenetic characterization of the blue filamentous bacterial community from an Icelandic geothermal spring. FEMS Microbiol. Ecol. 2001, 35, 123–128. [Google Scholar] [CrossRef]
- Blank, C.E.; Cady, S.L.; Pace, N.R. Microbial Composition of Near-Boiling Silica-Depositing Thermal Springs throughout Yellowstone National Park. Appl. Environ. Microbiol. 2002, 68, 5123–5135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castenholz, R.W.; McDermott, T.R. The Cyanidiales: Ecology, Biodiversity, and Biogeography. In Red Algae in the Genomic Age; Springer International Publishing: Dordrecht, The Netherlands, 2010; Volume 13, pp. 357–371. [Google Scholar]
- Konhauser, K.O. Bacterial iron biomineralisation in nature. FEMS Microbiol. Rev. 1997, 20, 315–326. [Google Scholar] [CrossRef]
- Konhauser, K.O. Diversity of bacterial iron mineralization. Earth Sci. Rev. 1998, 43, 91–121. [Google Scholar] [CrossRef]
- Phoenix, V.R.; Konhauser, K.O.; Ferris, F.G. Experimental study of iron and silica immobilization by bacteria in mixed Fe-Si systems: Implications for microbial silicification in hot springs. Can. J. Earth Sci. 2003, 40, 1669–1678. [Google Scholar] [CrossRef] [Green Version]
- Jones, B.; Renaut, R.W. Selective mineralization of microbes in Fe-rich precipitates (jarosite, hydrous ferric oxides) from acid hot springs in the Waiotapu geothermal area, North Island, New Zealand. Sediment. Geol. 2007, 194, 77–98. [Google Scholar] [CrossRef]
- Vollrath, S.; Behrends, T.; Van Cappellen, P. Oxygen Dependency of Neutrophilic Fe(II) Oxidation by Leptothrix Differs from Abiotic Reaction. Geomicrobiol. J. 2012, 29, 550–560. [Google Scholar] [CrossRef]
- Fleming, E.J.; Cetinić, I.; Chan, C.S.; King, D.W.; Emerson, D. Ecological succession among iron-oxidizing bacteria. ISME J. 2013, 8, 804–815. [Google Scholar] [CrossRef]
- Rodgers, K.; Browne, P.; Buddle, T.; Cook, K.; Greatrex, R.; Hampton, W.; Herdianita, N.; Holland, G.; Lynne, B.; Martin, R.; et al. Silica phases in sinters and residues from geothermal fields of New Zealand. Earth Sci. Rev. 2004, 66, 1–61. [Google Scholar] [CrossRef]
- Cady, S.L.; Farmer, J.D. Fossilization Processes in Siliceous Thermal Springs: Trends in Preservation Along Thermal Gradients. Novartis Foundation Symposia 2007, 202, 150–173. [Google Scholar] [CrossRef]
- Setiawan, F.A.; Rahayuningsih, E.; Petrus, H.T.B.M.; Nurpratama, M.I.; Perdana, I. Kinetics of silica precipitation in geothermal brine with seeds addition: Minimizing silica scaling in a cold re-injection system. Geotherm. Energy 2019, 7, 1–16. [Google Scholar] [CrossRef]
- Jones, B.; Konhauser, K.; Renaut, R.; Wheeler, R. Erratum to Microbial silicification in Iodine Pool, Waimangu geothermal area, North Island, New Zealand: Implications for recognition and identification of ancient silicified microbes. J. Geol. Soc. 2005, 162, 576. [Google Scholar] [CrossRef]
- Pancost, R.D.; Pressley, S.; Coleman, J.M.; Talbot, H.M.; Kelly, S.P.; Farrimond, P.; Schouten, S.; Benning, L.; Mountain, B.W. Composition and implications of diverse lipids in New Zealand Geothermal sinters. Geobiology 2006, 4, 71–92. [Google Scholar] [CrossRef]
- Hetzer, A.; Morgan, H.W.; McDonald, I.R.; Daughney, C.J. Microbial life in Champagne Pool, a geothermal spring in Waiotapu, New Zealand. Extremophiles 2007, 11, 605–614. [Google Scholar] [CrossRef]
- Kaur, G.; Mountain, B.W.; Hopmans, E.C.; Pancost, R.D. Relationship between lipid distribution and geochemical environment within Champagne Pool, Waiotapu, New Zealand. Org. Geochem. 2011, 42, 1203–1215. [Google Scholar] [CrossRef]
- Huber, R.; Wilharm, T.; Huber, D.; Trincone, A.; Burggraf, S.; König, H.; Reinhard, R.; Rockinger, I.; Fricke, H.; Stetter, K.O. Aquifex pyrophilus gen. nov. sp. nov., Represents a Novel Group of Marine Hyperthermophilic Hydrogen-Oxidizing Bacteria. Syst. Appl. Microbiol. 1992, 15, 340–351. [Google Scholar] [CrossRef]
- Huber, R.; Rossnagel, P.; Woese, C.; Rachel, R.; Langworthy, T.; Stetter, K. Formation of Ammonium from Nitrate During Chemolithoautotrophic Growth of the Extremely Thermophilic Bacterium Ammonifex degensii gen. nov. sp. nov. Syst. Appl. Microbiol. 1996, 19, 40–49. [Google Scholar] [CrossRef]
- Langworthy, T.A.; Holzer, G.; Zeikus, J.G.; Tornabene, T.G. Iso- and Anteiso-Branched Glycerol Diethers of the Thermophilic Anaerobe Thermodesulfotobacterium commune. Syst. Appl. Microbiol. 1983, 4, 1–17. [Google Scholar] [CrossRef]
- Knudsen, E.; Jantzen, E.; Bryn, K.; Ormerod, J.G. Quantitative and structural characteristics of lipids in Chlorobium and Chloroflexus. Arch. Microbiol. 1982, 132, 149–154. [Google Scholar] [CrossRef]
- Zelles, L. Identification of single cultured micro-organisms based on their whole-community fatty acid profiles, using an extended extraction procedure. Chemosphere 1999, 39, 665–682. [Google Scholar] [CrossRef]
- Jahnke, L.L.; Embaye, T.; Hope, J.; Turk, K.A.; Van Zuilen, M.; Marais, D.J.D.; Farmer, J.D.; Summons, R.E. Lipid biomarker and carbon isotopic signatures for stromatolite-forming, microbial mat communities and Phormidium cultures from Yellowstone National Park. Geobiology 2004, 2, 31–47. [Google Scholar] [CrossRef]
- Zelles, L. Phospholipid fatty acid profiles in selected members of soil microbial communities. Chemosphere 1997, 35, 275–294. [Google Scholar] [CrossRef]
- Grimalt, J.O.; De Wit, R.; Teixidor, P.; Albaigés, J. Lipid biogeochemistry of Phormidium and Microcoleus mats. Org. Geochem. 1992, 19, 509–530. [Google Scholar] [CrossRef]
- Zeng, Y.; Ward, D.M.; Brassell, S.C.; Eglinton, G. Biogeochemistry of hot spring environments. 2, Lipid compositions of Yellowstone (Wyoming, USA) cyanobacterial and Chloroflexus mats. Chem. Geol. 1992, 95, 327–345. [Google Scholar] [CrossRef]
- Zeng, Y.; Ward, D.M.; Brassell, S.C.; Eglinton, G. Biogeochemistry of hot spring environments. 3, Apolar and polar lipids in the biologically-active layers of a cyanobacterial mat. Chem. Geol. 1992, 95, 347–360. [Google Scholar] [CrossRef]
- Talbot, H.M.; Farrimond, P.; Schaeffer, P.; Pancost, R.D. Bacteriohopanepolyols in hydrothermal vent biogenic silicates. Org. Geochem. 2005, 36, 663–672. [Google Scholar] [CrossRef]
- Shiea, J.; Brassel, S.C.; Ward, D.M. Comparative analysis of extractable lipids in hot spring microbial mats and their component photosynthetic bacteria. Org. Geochem. 1991, 17, 309–319. [Google Scholar] [CrossRef]
- Matsumoto, G.I.; Ohtani, S.; Hirota, K. Biogeochemical feature of hydrocarbons on cyanobacterial mats from the McMurdo Dry Valleys, Antarctica. Proceedings of the National Institute of Polar Research (NIPR). Symp. Polar Biol. 1993, 6, 98–105. [Google Scholar]
- Wada, H.; Murata, N. Membrane Lipids in Cyanobacteria. In Lipids in Photosynthesis: Structure, Function and Genetics; Springer International Publishing: Dordrecht, The Netherlands, 2006; Volume 6, pp. 65–81. [Google Scholar]
- Teece, B.L.; George, S.C.; Djokic, T.; Campbell, K.A.; Ruff, S.W.; Van Kranendonk, M.J. Biomolecules from Fossilized Hot Spring Sinters: Implications for the Search for Life on Mars. Astrobiology 2020, 20, 537–551. [Google Scholar] [CrossRef] [Green Version]
- Havig, J.R.; Raymond, J.; Meyer-Dombard, D.R.; Zolotova, N.; Shock, E.L. Merging isotopes and community genomics in a siliceous sinter-depositing hot spring. J. Geophys. Res. Space Phys. 2011, 116. [Google Scholar] [CrossRef]
- Van Der Meer, M.T.J.; Schouten, S.; De Leeuw, J.W.; Ward, D.M. Autotrophy of green non-sulphur bacteria in hot spring microbial mats: Biological explanations for isotopically heavy organic carbon in the geological record. Environ. Microbiol. 2000, 2, 428–435. [Google Scholar] [CrossRef] [PubMed]
- Kjartansdóttir, R. Carbon Isotopes and Systematics of Icelandic Low-Temperature Geothermal Waters. Master’s Thesis, University of Iceland, Reykjavik, Iceland, 3 October 2014; 39p. [Google Scholar]
- Sveinbjörnsdóttir, A.E.; Arnórsson, S.; Heinemeier, J.; Ármannsson, H.; Kristmannsdóttir, H. Dissolved inorganic carbon isotopes in natural waters in Iceland. In Water-Rock Interaction XIII; Torres-Alvarado, P.A., Ed.; CRC Press: London, UK, 2010; pp. 99–102. [Google Scholar]
- Sveinbjörnsdóttir, A.E.; Heinemeier, J.; Arnórsson, S. Isotopic characteristics (δ18O, δD, δ13C, 14C) of thermal waters in the Mosfellssveit and Reykjavík low-temperature areas, Iceland. In Proceedings of the World Geothermal Congress, Antalya, Turkey, 24–29 April 2005; pp. 1–5. [Google Scholar]
- Skirnisdottir, S.; Hreggvidsson, G.O.; Hjörleifsdottir, S.; Marteinsson, V.T.; Petursdottir, S.K.; Holst, O.; Kristjansson, J.K. Influence of Sulfide and Temperature on Species Composition and Community Structure of Hot Spring Microbial Mats. Appl. Environ. Microbiol. 2000, 66, 2835–2841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez-Turiel, J.; Garcia-Valles, M.; Gimeno-Torrente, D.; Saavedra-Alonso, J.; Martinez-Manent, S. The hot spring and geyser sinters of El Tatio, Northern Chile. Sediment. Geol. 2005, 180, 125–147. [Google Scholar] [CrossRef]
- Farmer, J.D.; Marais, D.J.D. Exploring for a record of ancient Martian life. J. Geophys. Res. Space Phys. 1999, 104, 26977–26995. [Google Scholar] [CrossRef]
- Ruff, S.W.; Campbell, K.A.; Van Kranendonk, M.J.; Rice, M.S.; Farmer, J.D. The Case for Ancient Hot Springs in Gusev Crater, Mars. Astrobiology 2020, 20, 475–499. [Google Scholar] [CrossRef] [Green Version]
- Frydenvang, J.; Gasda, P.J.; Hurowitz, J.A.; Grotzinger, J.P.; Wiens, R.C.; Newsom, H.E.; Edgett, K.S.; Watkins, J.; Bridges, J.C.; Maurice, S.; et al. Diagenetic silica enrichment and late-stage groundwater activity in Gale crater, Mars. Geophys. Res. Lett. 2017, 44, 4716–4724. [Google Scholar] [CrossRef] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Temperature Range (°C) | pH Range | Morphology and Size | ||
|---|---|---|---|---|
| Bacteria | ||||
| Aquificales | Aquifex | 85–95 | 6.8–9.0 | rods, L 2–6 μm, Ø~0.5 μm |
| Sulfurihydrogenibium | 79–83 | <8.8 | rods, L 0.5–2.8 μm, Ø 0.5–1.0 μm | |
| Thermocrinis (ruber) | 82–88 | 7–10 | rods (L < 5 μm) to filaments (L > 30 μm long, Ø ~ 0.5 μm) | |
| FeOB | Acidithiobacillus | 25–30 | <4.0 | rods, L < 3 μm, Ø ~ 1 μm |
| Gallionella | <47 | 7.0–7.4 | helical filaments | |
| Leptothrix | 35–37 | 7.0–7.4 | sheathed filaments, L < 20 μm, Ø ~ 1 μm | |
| Cyanobacteria | ||||
| Anabaena | <30 | 7–10.5 | sheathed filamentous trichomes with constrictions at cross-walls, L < 20 μm | |
| Calothrix | <40 | 7–10 | sheathed filamentous trichomes, potential branching, L < 3 mm, Ø 15 μm | |
| Chloroflexus | 55–70 | 7.5–7.7 | unbranched sheathed filamentous cells (trichome), L 4–7 μm, Ø 0.6 μm | |
| Fischerella | 28–64 | 6.8–9 | barrel-shaped to short filamentous trichomes, potential branching, L < 10 μm, Ø 5 μm | |
| Oscillatoria | 20–30 | 7–7.5 | unbranched filaments with straight trichome | |
| Phormidium | 60–65 | 8.5–8.7 | unbranched, septate filaments with calyptra (thicker cap) at edge, L < 3 μm, Ø 1.5–3 μm, lumen Ø < 1 μm | |
| Spirulina | 30–35 | 7–10 | coiled filamentous trichomes, L 50–300 μm, Ø 10 μm | |
| Synechococcus (vulcanus) | 50–70 | >6.5 | subspherical, Ø < 6 μm | |
| Eukaryotic red algae (Cyanidiales) | ||||
| Cyanidium | 35–55 | 0.5–5 | subspherical, Ø < 10 μm | |
| Galderia | 35–55 | 0.5–5 | subspherical, Ø < 10 μm | |
| Cyanidioschyzon | 35–55 | 0.5–5 | subspherical, Ø 1–2 μm | |
| Fabric | Wavy (in Pond) | Wavy (in Meander) | Bubble | Palisade | Vitreous | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Compound | ||||||||||||
| Dialkyl glycerol diethers | ||||||||||||
| C18/C18 | −26.3 | −25.3 | −25.4 | −24.9 | −24.9 | −24.1 | −26.3 | −26.0 | −26.3 | −25.8 | ||
| C18/cycpropC20 | −25.0 | −24.9 | −24.7 | −24.2 | −23.4 | −24.0 | −25.7 | −25.9 | −25.5 | −24.7 | ||
| C18/C20 | −25.8 | −26.3 | −25.5 | −25.1 | −27.0 | −20.9 | −25.5 | −27.6 | −27.6 | −27.7 | ||
| cyclopropC20/cyclopropC20 | −23.0 | −22.2 | −24.9 | −24.2 | ||||||||
| C20/cyclopropC20 | −25.1 | −23.9 | −25.2 | −25.9 | ||||||||
| Archaeol | −26.7 | −27.2 | −26.8 | −27.8 | −27.1 | −30.8 | ||||||
| Monoalkyl glycerol ethers | ||||||||||||
| C18 | −25.6 | −24.6 | −24.5 | −25.0 | −25.9 | −24.7 | −24.6 | −24.5 | −24.9 | −24.8 | ||
| C20 | −26.5 | −25.3 | −27.0 | −24.8 | −25.6 | −24.5 | −25.9 | −24.0 | −25.3 | −26.0 | ||
| n-Alcohols | ||||||||||||
| C18 | −29.9 | −26.4 | −24.9 | −27.0 | −25.3 | −25.3 | −25.4 | |||||
| C20 | −32.9 | −31.1 | −24.8 | −27.2 | −24.6 | −26.1 | −26.6 | |||||
| n-Fatty acids | ||||||||||||
| C16:1 | −28.4 | −30.6 | ||||||||||
| C16 | −27.8 | −30.0 | −30.6 | −25.4 | −27.5 | |||||||
| C18:1 | −30.2 | −30.1 | −28.8 | −26.7 | ||||||||
| C18 | −30.6 | −29.8 | −22.7 | −24.9 | −26.8 | |||||||
| Pentacyclic terpenoids | ||||||||||||
| Friedolean-4-en-3-one | −31.2 | −31.1 | −30.0 | |||||||||
| Friedelin | −30.0 | −28.2 | −29.5 | |||||||||
| C29 alkane | −30.2 | |||||||||||
| C31 alkane | −29.8 | |||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Álvaro, J.J.; Sánchez-Román, M.; Nierop, K.G.J.; Peterse, F. Multiscale Microbial Preservation and Biogeochemical Signals in a Modern Hot-Spring Siliceous Sinter Rich in CO2 Emissions, Krýsuvík Geothermal Field, Iceland. Minerals 2021, 11, 263. https://doi.org/10.3390/min11030263
Álvaro JJ, Sánchez-Román M, Nierop KGJ, Peterse F. Multiscale Microbial Preservation and Biogeochemical Signals in a Modern Hot-Spring Siliceous Sinter Rich in CO2 Emissions, Krýsuvík Geothermal Field, Iceland. Minerals. 2021; 11(3):263. https://doi.org/10.3390/min11030263
Chicago/Turabian StyleÁlvaro, Jose Javier, Mónica Sánchez-Román, Klaas G.J. Nierop, and Francien Peterse. 2021. "Multiscale Microbial Preservation and Biogeochemical Signals in a Modern Hot-Spring Siliceous Sinter Rich in CO2 Emissions, Krýsuvík Geothermal Field, Iceland" Minerals 11, no. 3: 263. https://doi.org/10.3390/min11030263
APA StyleÁlvaro, J. J., Sánchez-Román, M., Nierop, K. G. J., & Peterse, F. (2021). Multiscale Microbial Preservation and Biogeochemical Signals in a Modern Hot-Spring Siliceous Sinter Rich in CO2 Emissions, Krýsuvík Geothermal Field, Iceland. Minerals, 11(3), 263. https://doi.org/10.3390/min11030263