

Deciphering Alterations of Rodent Bones through In Vitro Digestion: An Avenue to Understand Pre-Diagenetic Agents?



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
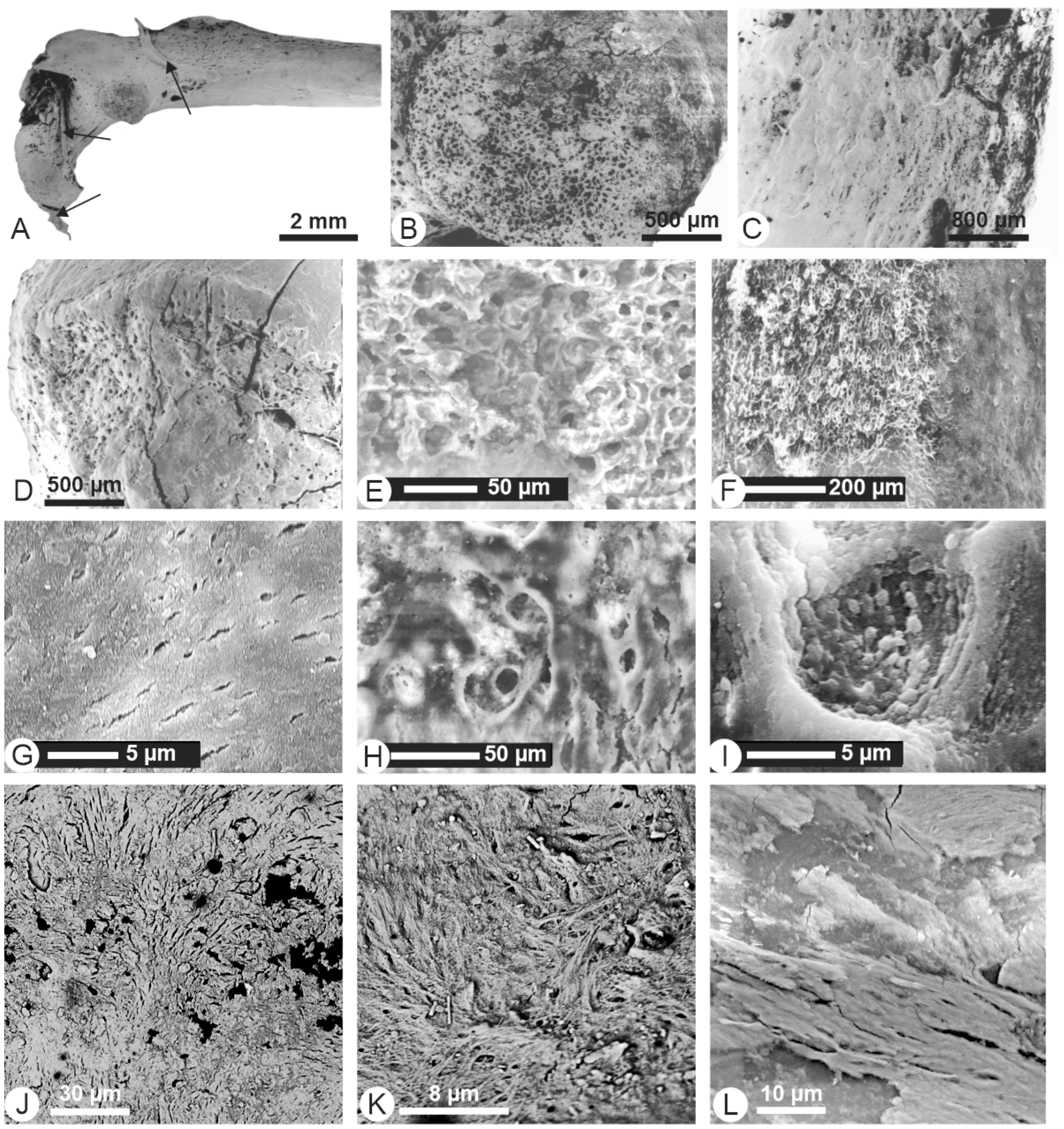{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Material
2.1.1. Fresh Specimens
2.1.2. Pellet Specimens
2.1.3. Enzymes Buffers
2.2. Methods
2.2.1. Bone Preparation
2.2.2. Observations
2.2.3. Experimental Set Up of Acidic and Enzymatic Hydrolyses
3. Results
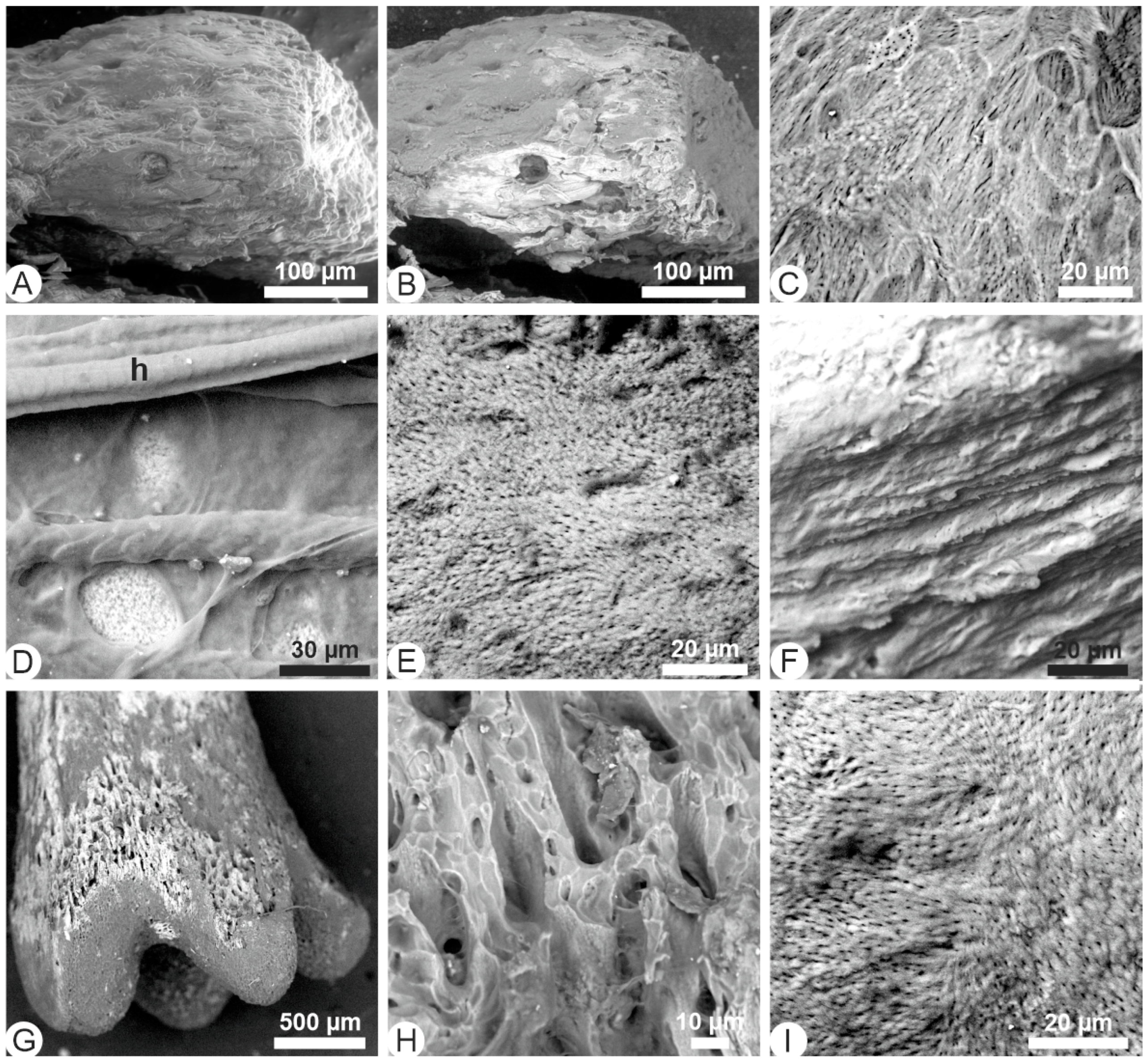
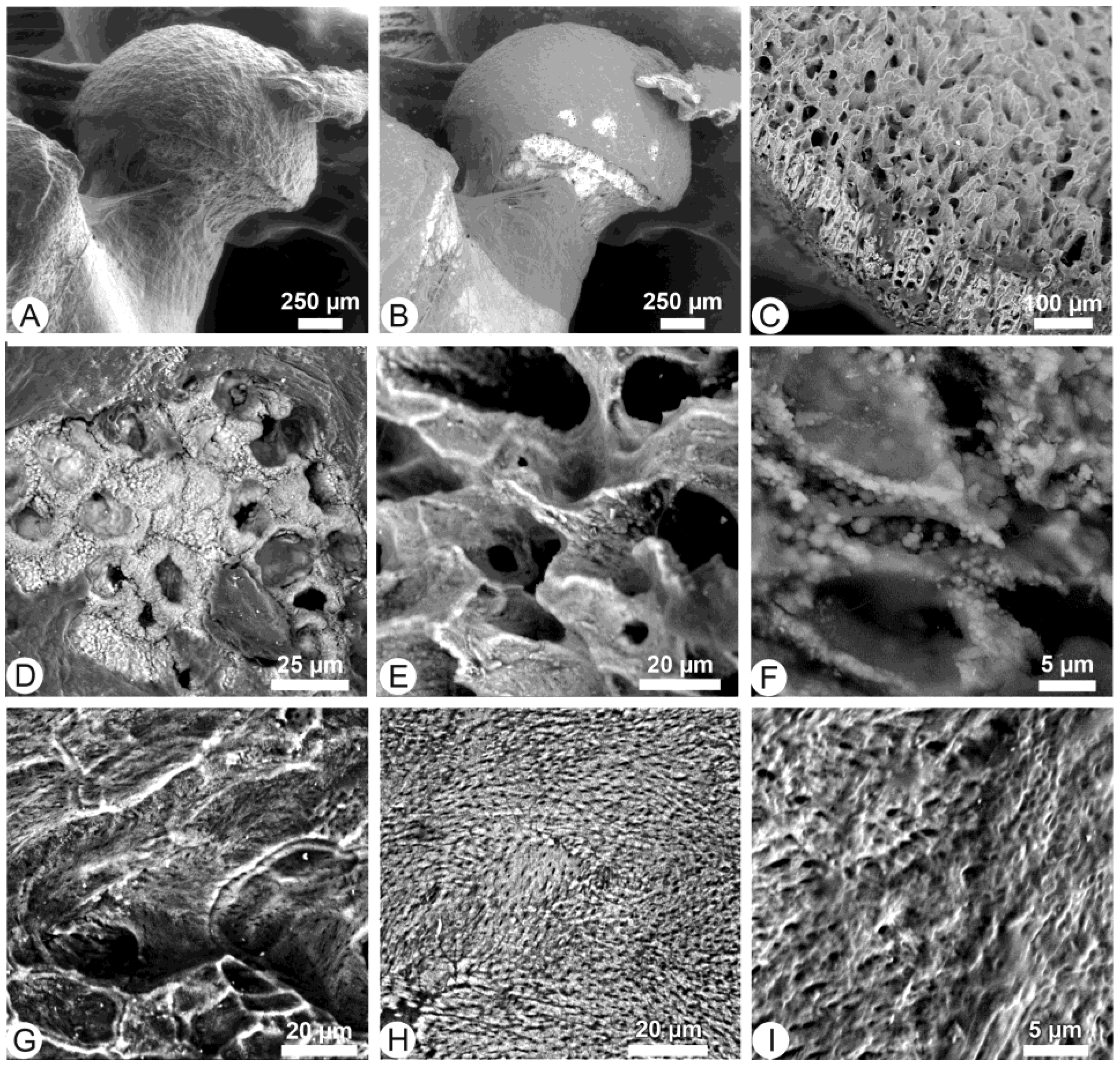
3.1. Unetched Bones
3.2. Acidic Hydrolyses
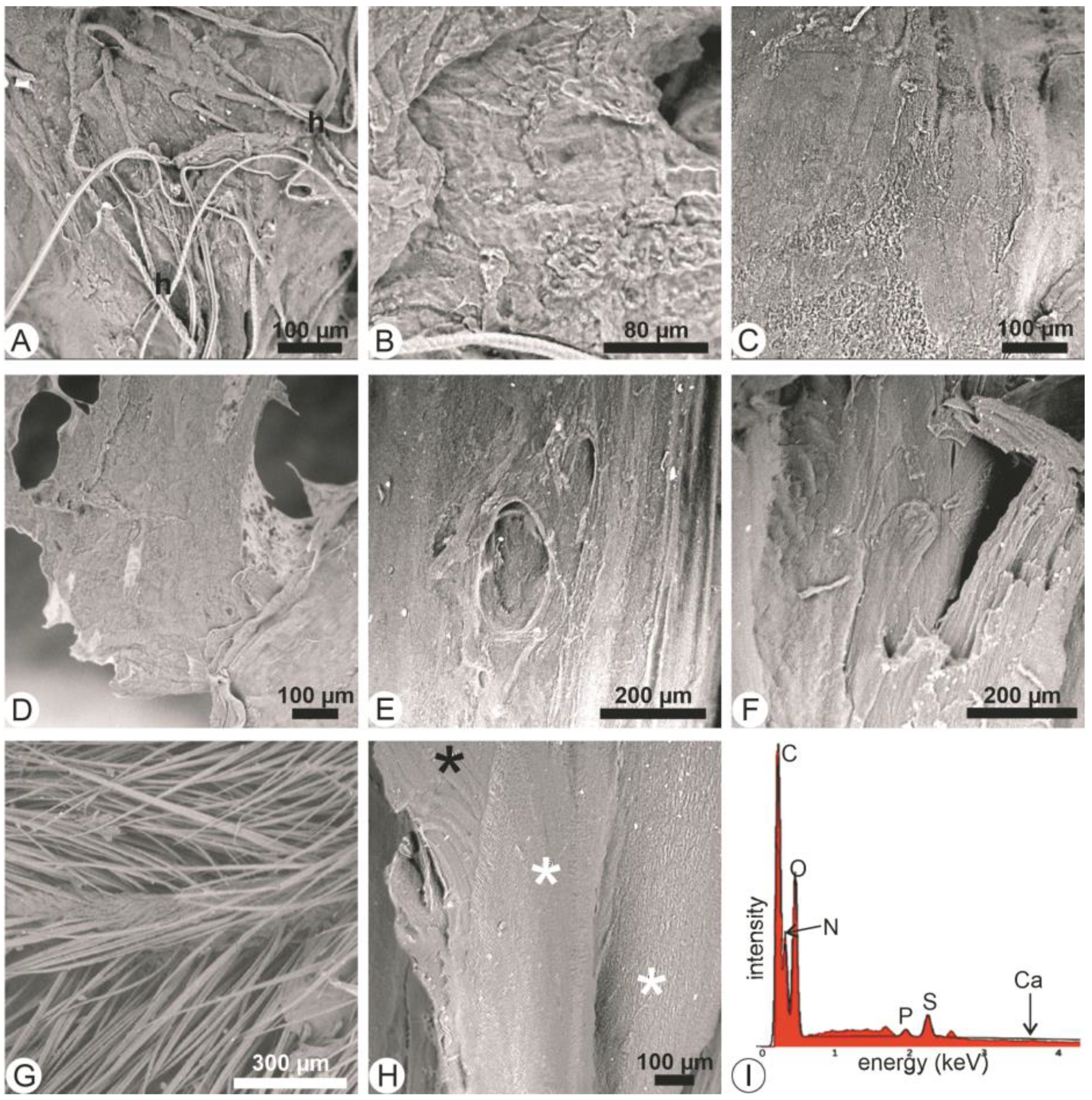
3.3. Trypsin Hydrolyses
3.4. Lipase Hydrolyses
3.5. Trypsin + Lipase Hydrolyses
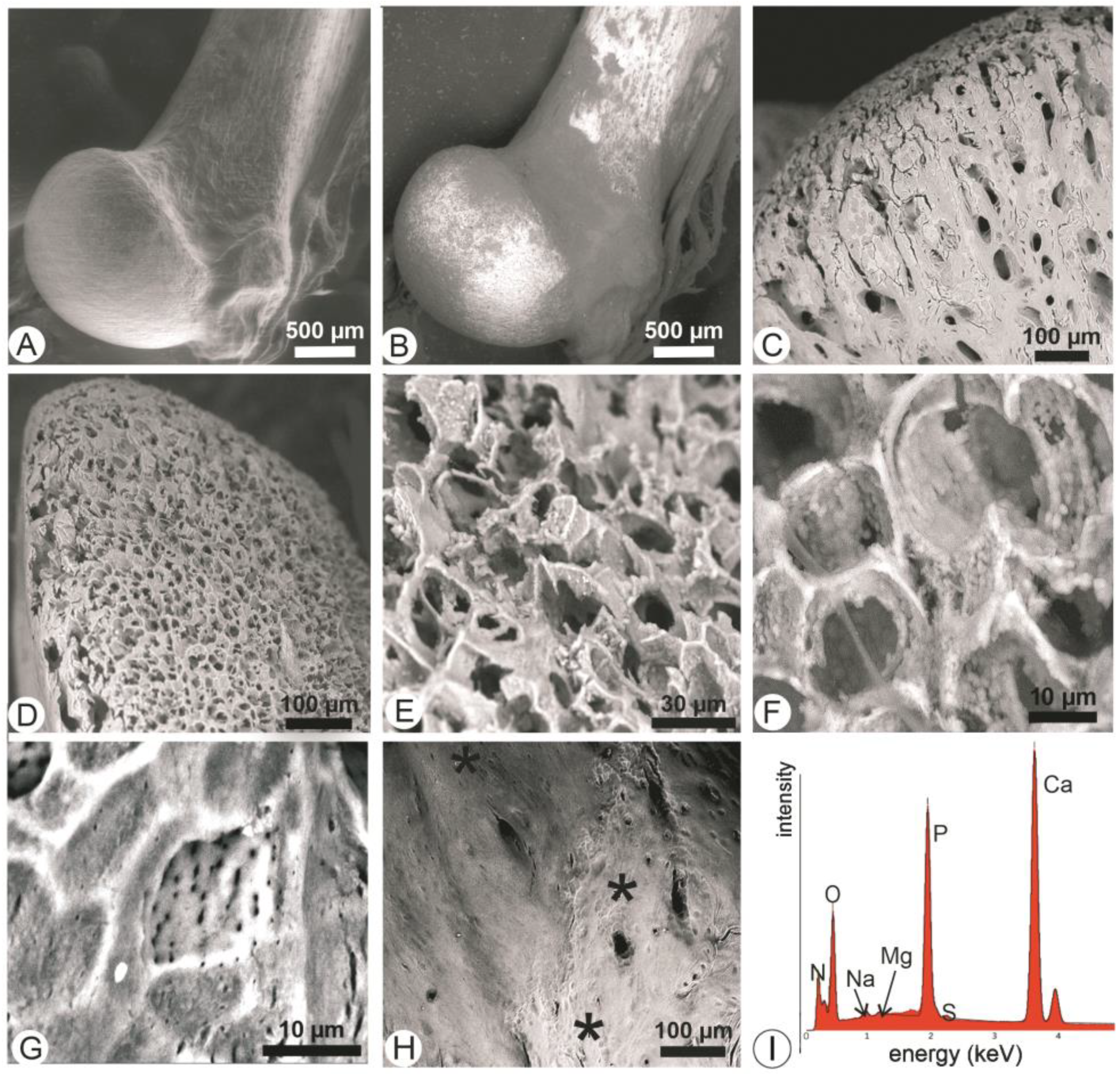
3.6. Bones Extracted from Tyto alba Pellets
3.7. Bones Extracted from Bubo bubo Pellets
4. Discussion
4.1. Samples
4.2. Choice of Experimental Conditions
4.3. Physical Parameters
4.4. In Vitro Digestion Effects
4.5. Comparison of In Vitro and In Vivo Alterations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Efremov, I.A. Taphonomy: A new branch of paleontology. Pan-Amer. Geol. 1940, 74, 81–93. [Google Scholar]
- Fernandez-Jalvo, Y.; Andrews, P.; Denys, C.; Sese, C.; Stoetzel, E.; Marin Monfort, D.; Pequero, D. Taphonomy for taxonomists, implications in small mammal studies. Quarter. Sci. Rev. 2016, 139, 138–157. [Google Scholar] [CrossRef]
- Klaphake, E.; Clancy, J. Raptor gastroenterology. Vet. Clin. Exotic Animal 2005, 8, 307–327. [Google Scholar] [CrossRef] [PubMed]
- Weissbrod, L.; Dayan, T.; Kaufman, D.; Weinstein-Evron, M. Micromammal taphonomy of el-Wad Terrace, Mount Carmel, Israel: Distinguishing cultural from natural depositional agents in the Late Natufian. J. Archaeol. Sci. 2005, 32, 1–17. [Google Scholar] [CrossRef]
- Reed, D.N. Taphonomic implications of roosting behavior and trophic habits in two species of African owl. J. Archaeol. Sci. 2005, 32, 1669–1676. [Google Scholar] [CrossRef]
- Fernandez-Jalvo, Y.; Avery, D.M. Pleistocene micromammals and their predators at Wondevewerk Cave, South Africa. Afr. Arch. Rev. 2015, 32, 751–791. [Google Scholar] [CrossRef]
- Fernandez-Jalvo, Y.; Andrews, P. Atlas of Taphonomic Identifications: 1001+ Images of Fossil and Recent Mammal Bone Modification; Springer: Dordrecht, The Netherlands, 2016; 359p. [Google Scholar]
- Andrews, P. Owls, Caves and Fossils; Natural History Museum Publications: London, UK, 1990. [Google Scholar]
- Barbosa, B.B.; Barauna, A.C.G.; Domingues, S.F.S. Birds of prey pellet analysis: Morphometry and consumption interval of captive Accipitridae species. Cienca Animal 2021, 31, 30–37. [Google Scholar]
- Bochenski, Z.M.; Tomek, T. Preservation of bird bones: Erosion versus digestion by owls. Intern. J. Osteoarch. 1997, 7, 372–387. [Google Scholar] [CrossRef]
- Fernandez-Jalvo, Y.; Andrews, P. Small Mammals taphonomy of Grand Dolina, Atapuerca (Burgos), Spain. J. Archaeol. Sci. 1992, 19, 407–428. [Google Scholar] [CrossRef]
- Rensberger, J.M.; Krentz, H.B. Microscopic effects of predator digestion on the surfaces of bones and teeth. Scanning Microsc. 1988, 2, 30. [Google Scholar]
- Comay, O.; Dayan, T. Taphonomic signatures of owls: New insights into micromammal assemblages. Palaeogeogr. Palaeoclim. Palaeoecol. 2018, 492, 81–91. [Google Scholar] [CrossRef]
- Simmons, R.E.; Avery, D.M.; Avery, G. Biases in diets determined from pellets and remains: Correction factors for a mammal and bird-eating raptor. J. Raptor Res. 1991, 25, 63–67. [Google Scholar]
- Marchesi, L.; Pedrini, P.; Sergio, F. Biases associated with diet study methods in the European eagle—Owl. J. Raptor Res. 2002, 36, 11–16. [Google Scholar]
- Denys, C. Des référentiels en taphonomie des petits vertébrés: Bilan et perspectives. In Taphonomie des Petits Vertébrés: Référentiels et Transferts Aux Fossiles; Laroulandie, V., Mallye, J.B., Denys, C., Eds.; Series 2269; British Archaeological Reports Oxford Ltd.: Oxford, UK, 2011; pp. 7–22. [Google Scholar]
- Fernandez-Jalvo, Y.; Andrews, P.; Sevilla, P.; Requejo, V. Digestion versus abrasion features in rodent bones. Lethaia 2014, 47, 323–336. [Google Scholar] [CrossRef]
- Weber, K.; Winkler, D.E.; Schulz-Kornas, E.; Kaiser, Y.M.; Tütken, T. The good, the bad and the ugly—A visual guide for common post-mortem wear patterns in vertebrate teeth. Palaeogeogr. Palaeoclim. Palaeoecol. 2021, 578, 110577. [Google Scholar] [CrossRef]
- Kusmer, K.D. Taphonomy of owl pellet deposition. J. Paleont. 1990, 64, 629–637. [Google Scholar] [CrossRef]
- Yalden, D.W.; Yalden, P.E. An experimental investigation of examining Kestrel diet by pellet analysis. Bird Study 1985, 32, 50–55. [Google Scholar] [CrossRef]
- Denys, C. Taphonomie des sites à petits vertébrés, importance de la digestion et des altérations post-prédation. In Manuel de Taphonomie; Denys, C., Pathou-Mathis, M., Eds.; Errance Editions: Paris, France, 2014; pp. 101–122. [Google Scholar]
- Denys, C. Accumulations de rapaces nocturnes et diurnes. TaphonomieS. Paris: Éditions Des Arch. Contemp. 2017, 347–367. [Google Scholar]
- Shipman, P. Applications of scanning electron microscopy to taphonomic problems. Ann. N. Y. Acad. Sci. 1981, 376, 357–385. [Google Scholar] [CrossRef]
- Cummings, J.H.; Duke, G.E.; Jegers, A.A. Corrosion of bone by solutions simulating raptor gastric juices. Raptor Res. 1976, 10, 55–57. [Google Scholar]
- Terry, R.C.; Laney, J.A.; Hay-Roe, S.H. Quantifying the digestive fingerprints of predators on the bones of their preys using scanning electron microscopy. Palaios 2018, 33, 487–497. [Google Scholar] [CrossRef]
- Bruderer, C.; Denys, C. Inventaire taxonomique et taphonomique d’un assemblage de pelotes d’un site de nidification de T. alba de Mauritanie. Bonner Zool. Beit. 1999, 48, 245–257. [Google Scholar]
- Dodson, P.; Wexlar, D. Taphonomic investigations of owl pellets. Paleobiology 1979, 5, 275–284. [Google Scholar] [CrossRef]
- Denys, C.; Dauphin, Y.; Rzebik-Kowalska, B.; Kowalski, K. Taphonomic study of Algerian owl pellet assemblages and differential preservation of some rodents: Palaeontological implications. Acta Zool. Crac. 1996, 39, 103–116. [Google Scholar]
- Lopez, J.M. Actualistic taphonomy of barn owl pellet derived small mammal bone accumulations in arid environment of South America. J. Quater. Sci. 2020, 35, 1057–1069. [Google Scholar] [CrossRef]
- Boyde, A.; Jones, S.J. Back-scattered electron imaging of skeletal tissues. Metab. Bone Dis. Rel. Res. 1983, 5, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Denys, C.; Fernandez-Jalvo, Y.; Dauphin, Y. Experimental taphonomy: Preliminary results of the digestion of micromammal bones in the laboratory. C. R Acad. Sci. Paris 1995, 321, 803–809. [Google Scholar]
- Chen, Y.-H.; Zhao, H. Evolution of digestive enzymes and dietary diversification in birds. PeerJ 2019, 46, 73–82. [Google Scholar] [CrossRef] [Green Version]
- Duke, G.E.; Jegers, A.A.; Loff, G.; Evanson, O.A. Gastric digestion in some raptors. Comp. Biochem. Physiol. 1975, 50A, 649–656. [Google Scholar] [CrossRef]
- Ben Bacha, A.; Frikha, F.; Djemal, I.; Fendri, A.; Miled, N.; Gargouri, Y.; Medjoub, H. Biochemical and structural comparative study between bird and mammal pancreatic colipases. J. Lipid Res. 2006, 47, 2701–2711. [Google Scholar] [CrossRef]
- Daneel, A. Dietary Modulation of Intestinal Fermentation in Raptors. Master’s Thesis, Faculty of Veterinary Medicine, Ghent University, Ghent, Belgium, 2018. [Google Scholar]
- Leprince, P.; Dandrifosse, G.; Schoffeniels, E. The digestive enzymes and acidity of the pellets regurgitated by raptors. Biochem. System. Ecol. 1979, 7, 223–227. [Google Scholar] [CrossRef]
- Reed, C.I.; Reed, B.P. The mechanism of pellet formation in the Great Horned Owl (Bubo virginianus). Science 1928, 68, 359–360. [Google Scholar] [CrossRef]
- Boyde, A.; Hobdell, M.H. Scanning electron microscopy of lamellar bone. Z. Zellforsch. 1969, 93, 213–231. [Google Scholar] [CrossRef] [PubMed]
- Chambers, T.J.; Revell, P.A.; Fuller, K.; Athanasou, N.A. Resorption of bone by isolated rabbit osteoclasts. J. Cell Sci. 1984, 66, 383–399. [Google Scholar] [CrossRef] [PubMed]
- Gentzsch, C.; Delling, G.; Kaiser, E. Microstructural classification of resorption lacunae and perforations in human proximal femora. Calcif. Tissue Int. 2003, 72, 698–709. [Google Scholar] [CrossRef]
- Hefti, T.; Frischherz, M.; Spencer, N.D.; Hall, H.; Schlottig, F. A comparison of osteoclast resorption pits on bone with titanium and zirconia surfaces. Biomaterials 2010, 31, 7321–7331. [Google Scholar] [CrossRef]
- Dauphin, Y.; Denys, C.; Denis, A. Les mécanismes de formation des gisements de microvertébrés: Modifications de la composition chimique des os et dents de rongeurs issus de pelotes de régurgitation de rapaces. C. R. Acad. Sci. Paris 1988, 307, 603–608. [Google Scholar]
- Dauphin, Y.; Denys, C.; Denis, A. 1989. Les mécanismes de formation des gisements de microvertébrés. 2- Composition chimique élémentaire des os et dents de rongeurs provenant de pelotes de régurgitation. Bull. Mus. Natl. Hist. Nat. 1989, 11, 253–269. [Google Scholar]
- High, K.; Milner, N.; Panter, I.; Penkman, K.E.H. Apatite for destruction; investigating bone degradation due to high acidity at Star Carr. J. Arch. Sci. 2015, 50, 159–168. [Google Scholar] [CrossRef]
- Duke, G.E.; Evanson, O.A.; Jegers, A. Meal to pellet intervals in 14 species of captive raptors. Comp. Biochem Physiol. 1976, 53A, 1–6. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Denys, C.; Reed, D.N.; Dauphin, Y. Deciphering Alterations of Rodent Bones through In Vitro Digestion: An Avenue to Understand Pre-Diagenetic Agents? Minerals 2023, 13, 124. https://doi.org/10.3390/min13010124
Denys C, Reed DN, Dauphin Y. Deciphering Alterations of Rodent Bones through In Vitro Digestion: An Avenue to Understand Pre-Diagenetic Agents? Minerals. 2023; 13(1):124. https://doi.org/10.3390/min13010124
Chicago/Turabian StyleDenys, Christiane, Denné N. Reed, and Yannicke Dauphin. 2023. "Deciphering Alterations of Rodent Bones through In Vitro Digestion: An Avenue to Understand Pre-Diagenetic Agents?" Minerals 13, no. 1: 124. https://doi.org/10.3390/min13010124
APA StyleDenys, C., Reed, D. N., & Dauphin, Y. (2023). Deciphering Alterations of Rodent Bones through In Vitro Digestion: An Avenue to Understand Pre-Diagenetic Agents? Minerals, 13(1), 124. https://doi.org/10.3390/min13010124