
Integrated Information on the Structure and Composition of the Ostrich Eggshell (Struthio camelus)

 ,
, 
 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. CT Scan-X-ray Computed Tomography
2.3. Scanning Electron Microscope (SEM)
2.4. Atomic Force Microscopy
2.5. Fourier Transform Infrared Spectroscopy (FTIR)
2.6. Raman Spectroscopy
2.7. Electron Backscatter Diffraction (EBSD)
2.8. Electron Microprobe Analyses
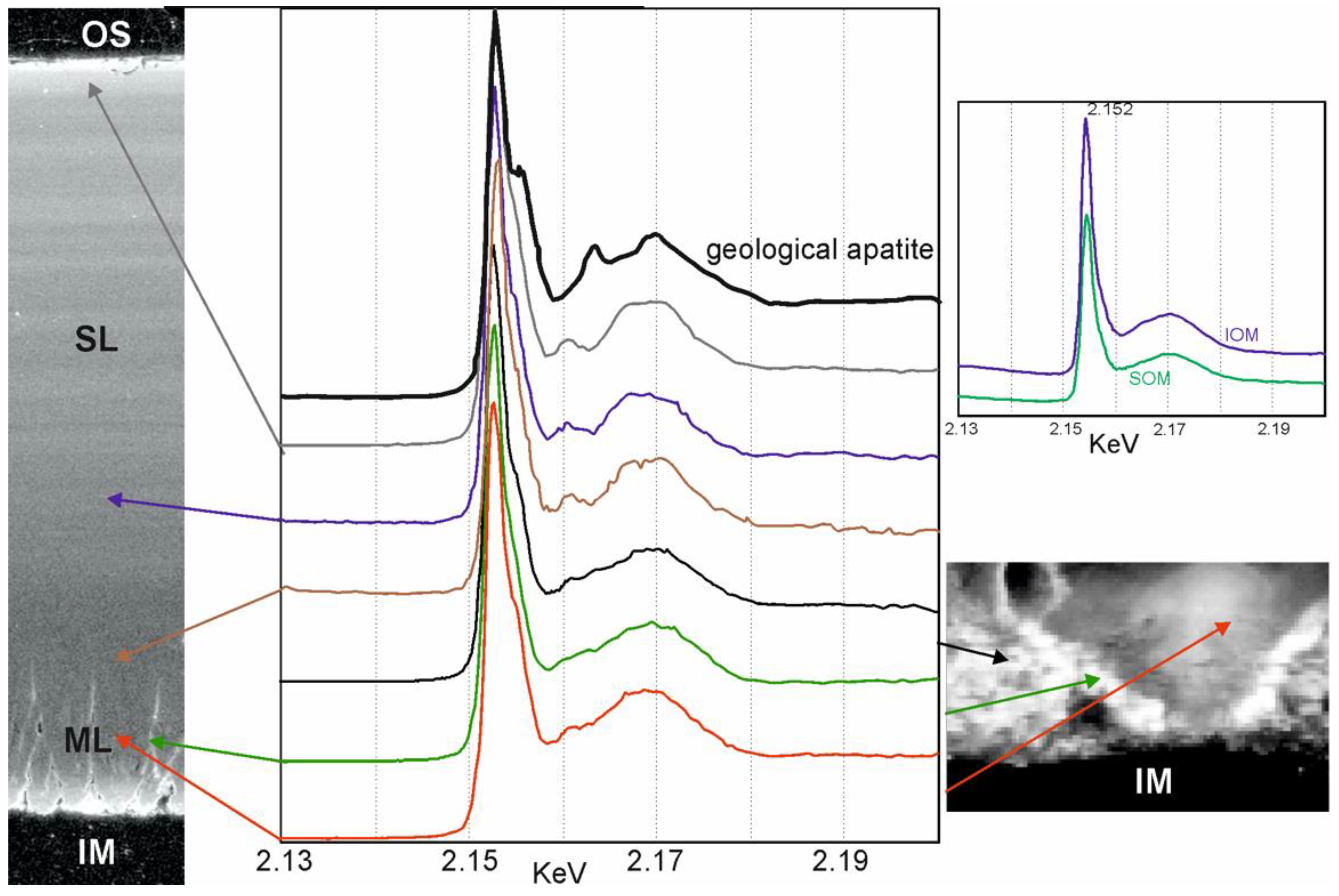
2.9. Micro-XANES Spectroscopy Analyses (X-ray Absorption near Edge Structure)
2.10. Extraction of the Organic Matrices
3. Results
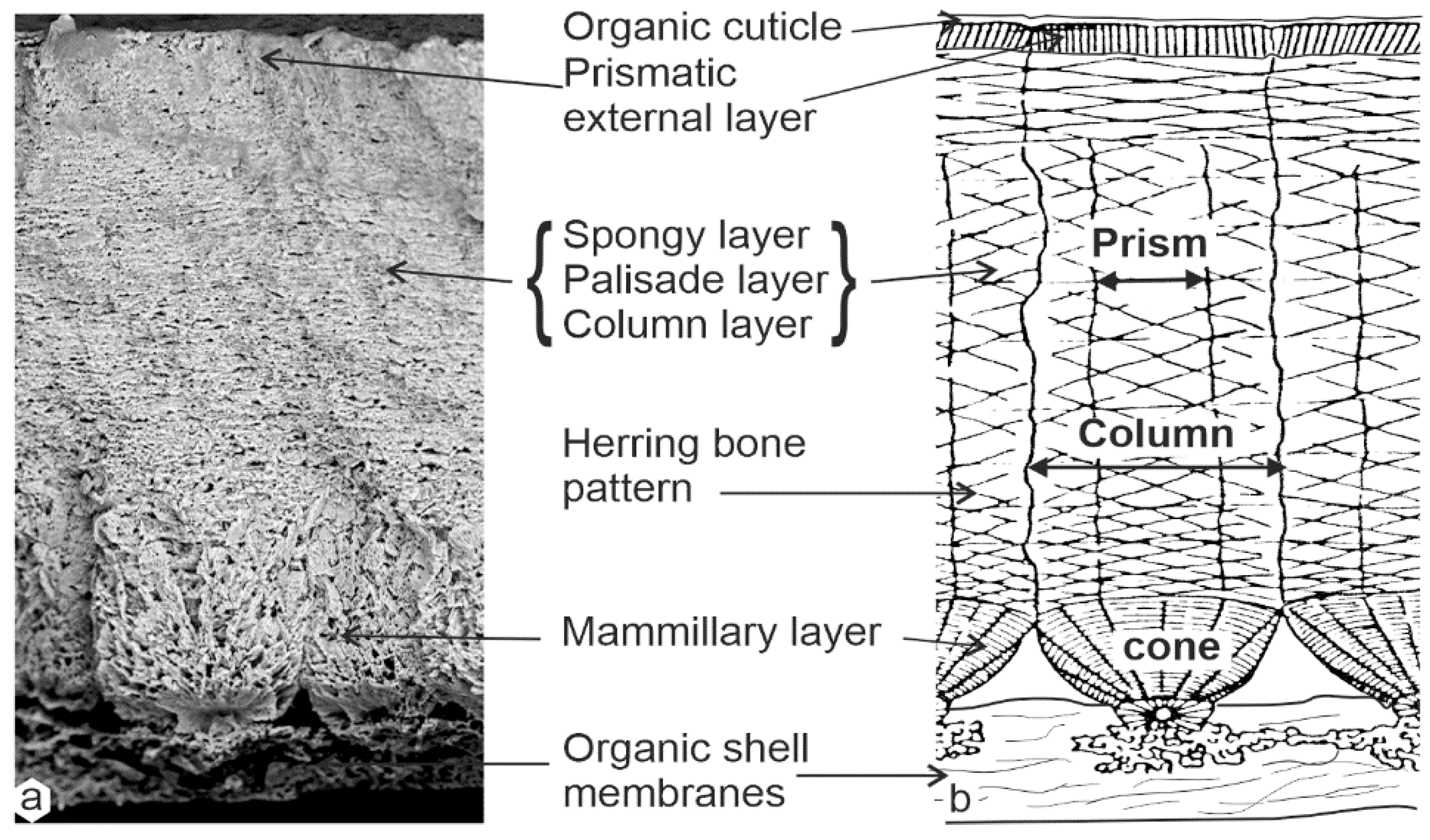
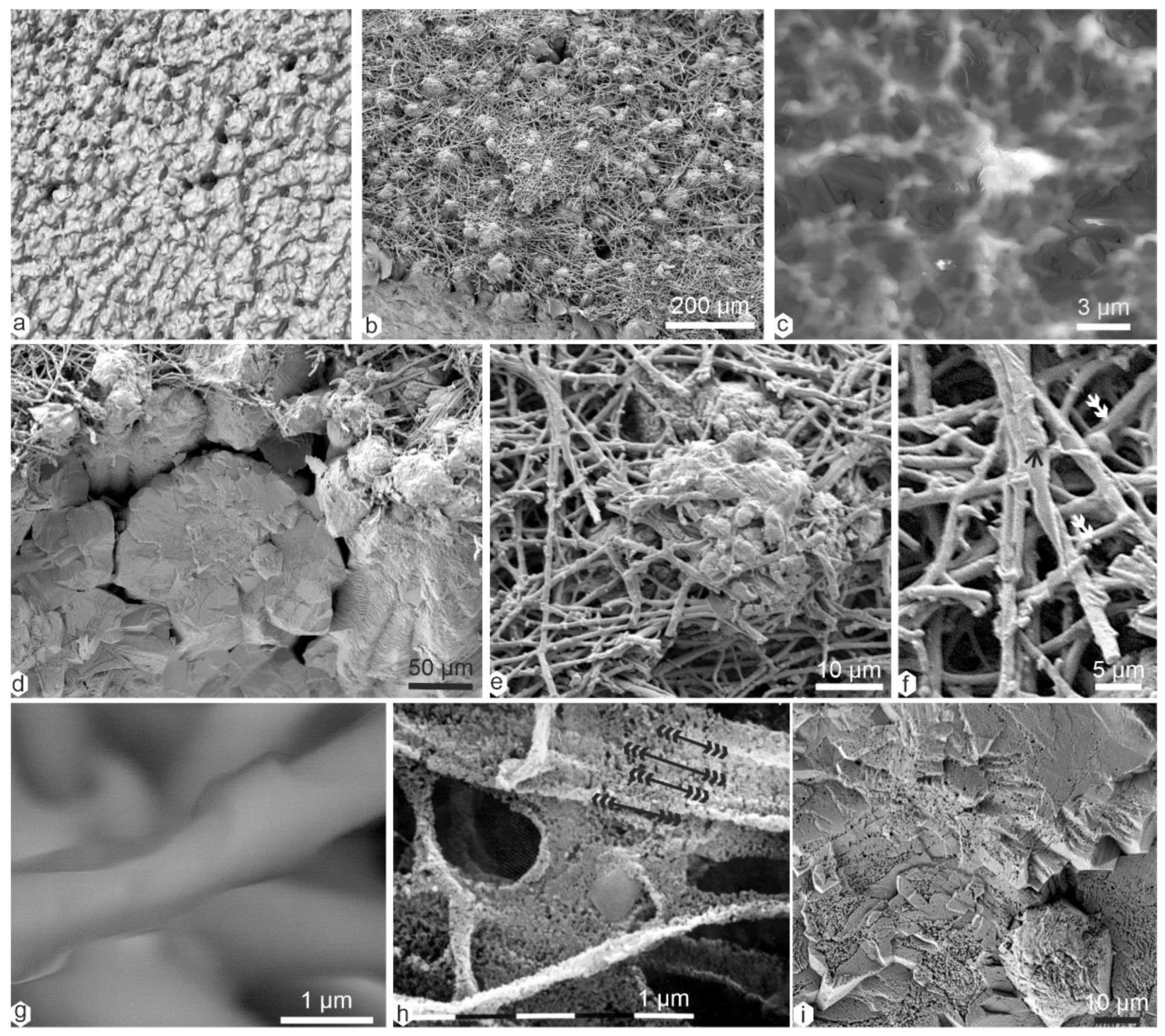
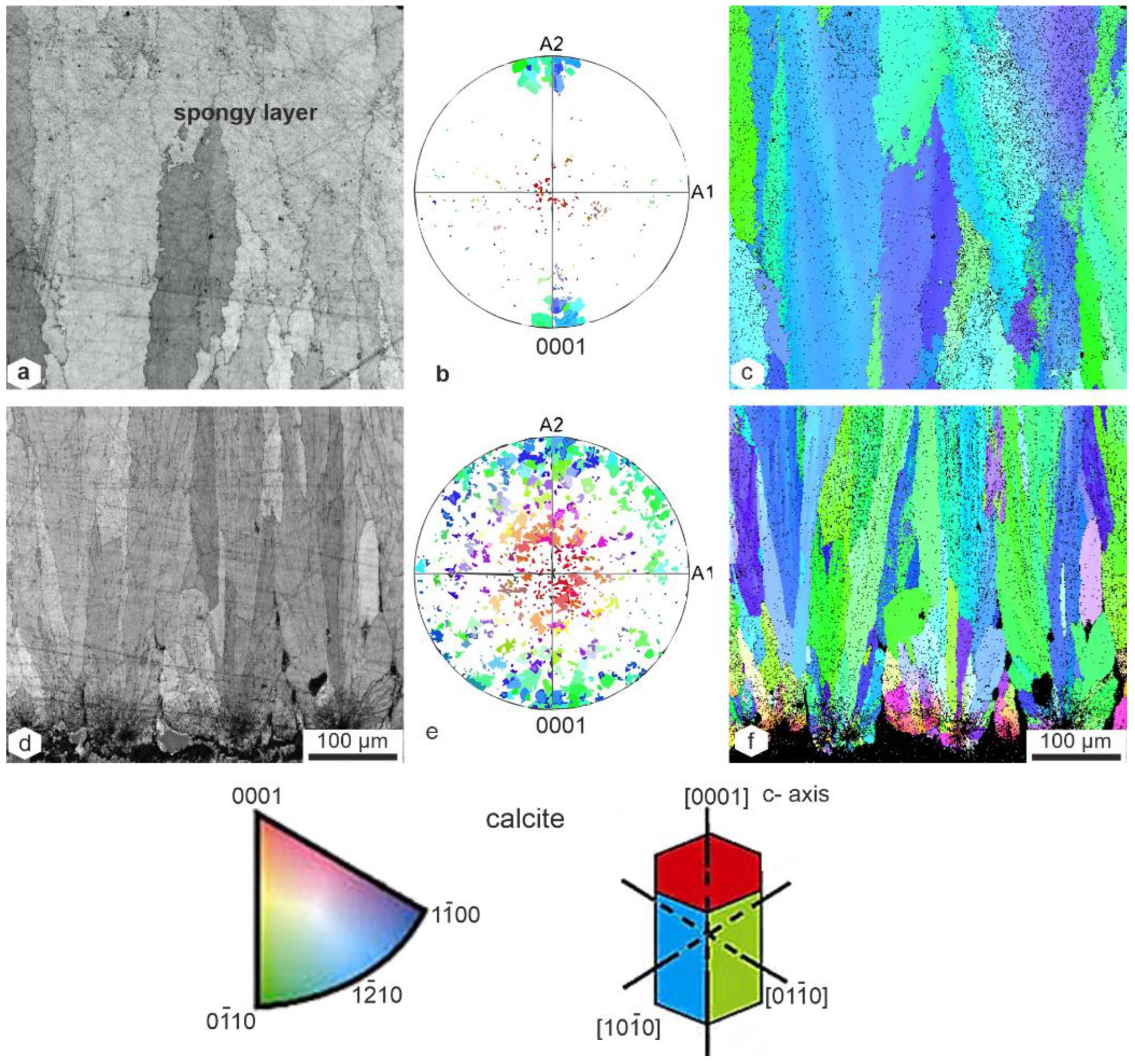
3.1. Structures
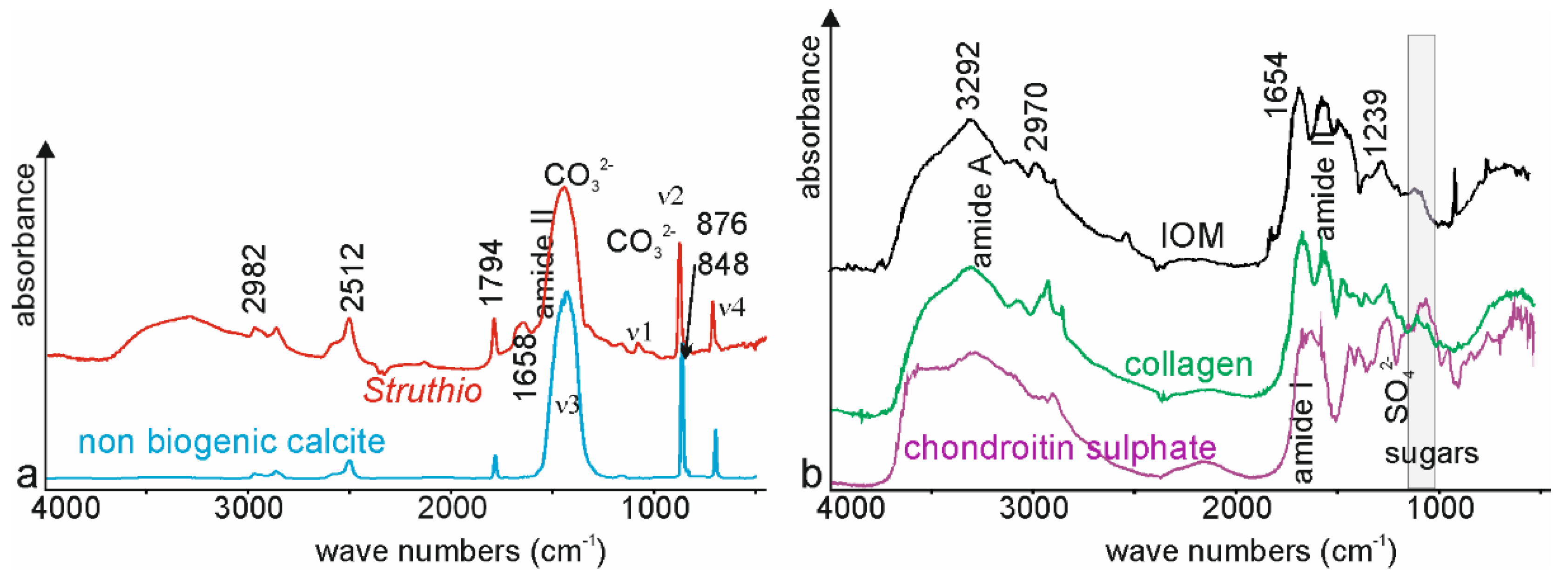
3.2. Bulk Composition–Mineralogy
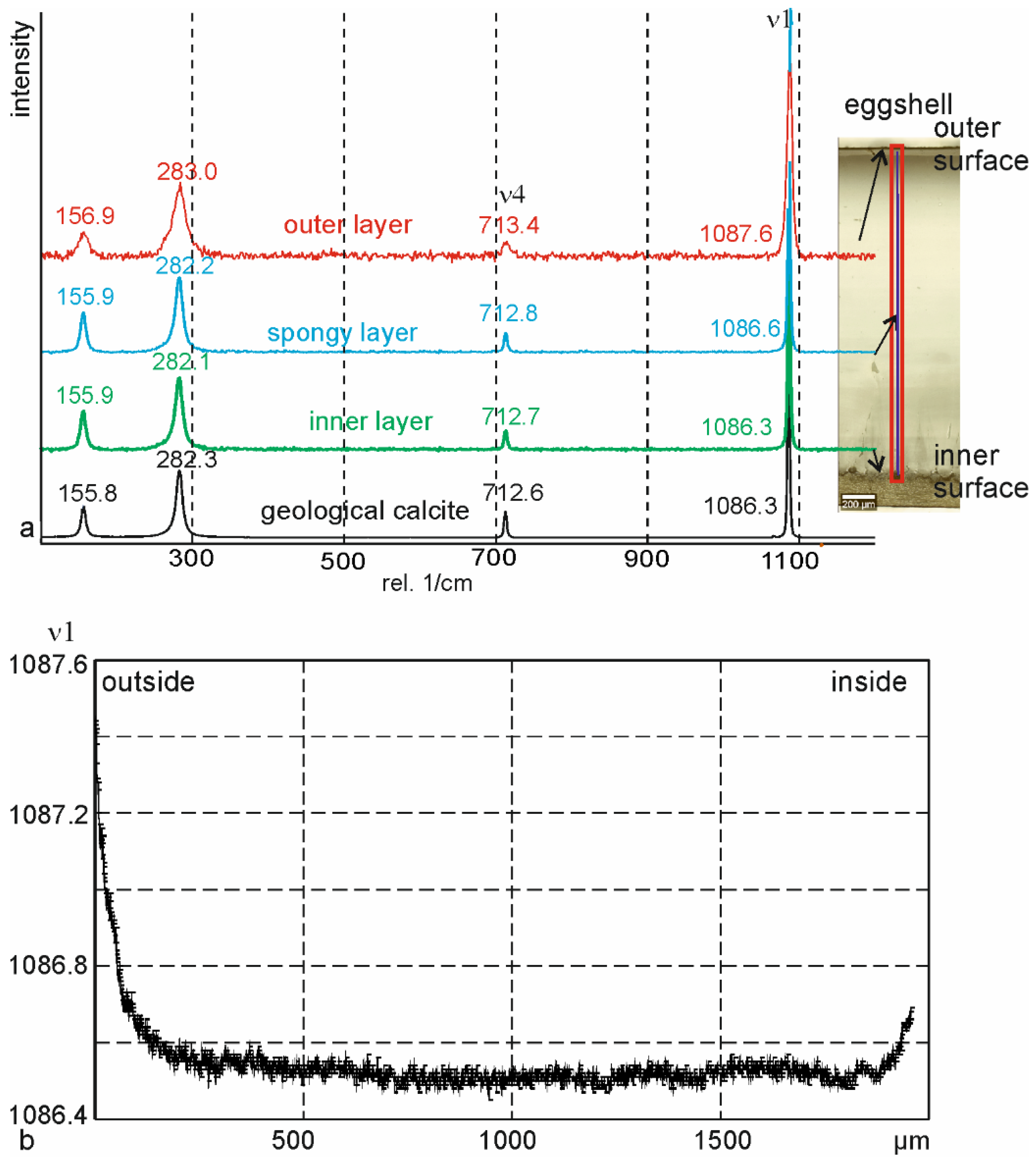
3.3. Crystallography
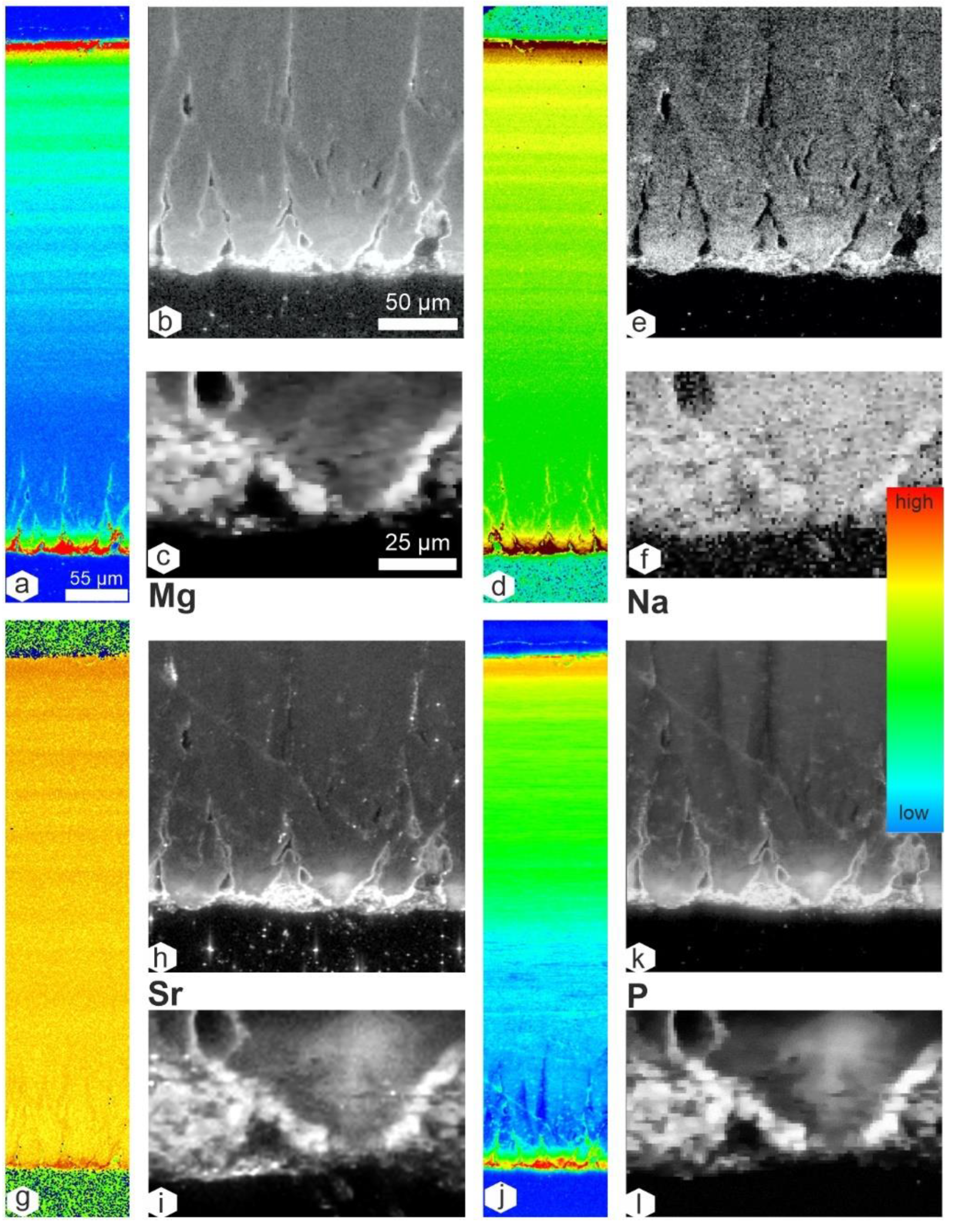
3.4. Chemical Composition
4. Discussion and Concluding Remarks
4.1. Structure
4.2. Mineralogy–Crystallography
4.3. Composition
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rovensky, J.; Stancikova, M.; Masaryk, P.; Svik, K.; Istok, R. Eggshell calcium in the prevention and treatment of osteoporosis. Int. J. Clin. Pharmacol. Res. 2003, 23, 83–92. [Google Scholar] [PubMed]
- Arias, J.L.; Arias, J.I.; Fernandez, M.S. Avian eggshell as a template for biomimetic synthesis of new materials. In Handbook of Biomineralization: Biological Aspects and Structure Formation; Wiley: Hoboken, NJ, USA, 2007; pp. 109–118. [Google Scholar]
- Chen, X.; Zhu, L.; Wen, W.; Lu, L.; Luo, B.; Zhou, C. Biomimetic mineralisation of eggshell membrane featuring natural nanofiber network structure for improving its osteogenic activity. Colloids Surf. B Biointerfaces 2019, 179, 299–308. [Google Scholar] [CrossRef] [PubMed]
- Elizondo-Villarreal, N.; Martinez-de-la-Cruz, A.; Guerra, R.O.; Gomez-Ortega, J.L.; Torres-Martinez, L.M.; Casrano, V.M. Biomaterials from agricultural waste: Eggshell-based hydroxyapatite. Water Air Soil Pollut. 2012, 223, 3643–3646. [Google Scholar] [CrossRef]
- Abdulrahman, I.; Tijani, H.I.; Mohammed, B.A.; Saidu, H.; Yusuf, H.; Jibrin, M.N.; Mohammed, S. From garbage to biomaterials: An overview on eggshell based hydroxyapatite. J. Materials 2014, 2014, 802467. [Google Scholar] [CrossRef]
- Macha, I.J.; Ozyegin, L.S.; Oktar, F.N.; Ben-Nissan, B. Conversion of ostrich eggshells (Struthio camelus) to calcium phosphates. J. Austr. Ceramic. Soc. 2015, 51, 125–133. [Google Scholar]
- Nys, Y.; Gautron, J.; Garcia-Ruiz, M.; Hincke, M.E. Avian eggshell mineralization: Biochemical and functional characterization of matrix proteins. Comptes Rendus Palevol. 2004, 3, 549–562. [Google Scholar] [CrossRef]
- Erben, H.K. Ultrastrukturen und mineralisation rezenter und fossiler eischalen bei vögeln und reptilien. Biomineralization 1970, 1, 1–66. [Google Scholar]
- Picard, J.; Paul-Gardais, A.; Vedel, M. Glycoprotéines sulfates des membranes de l’oeuf de poule et de l’oviducte. Biochim. Biophys. Acta 1973, 320, 427–441. [Google Scholar] [CrossRef] [PubMed]
- Pines, M.; Knopov, V.; Bar, A. Involvement of osteopontin in eggshell formation in the laying chicken. Matrix Biol. 1994, 14, 765–771. [Google Scholar] [CrossRef]
- Hincke, M.T. Ovalbumin is a component of the chicken eggshell matrix. Connect. Tiss. Res. 1995, 31, 227–233. [Google Scholar] [CrossRef]
- Carrino, D.A.; Rodriguez, J.P.; Caplan, A.L. Dermatan sulfate proteoglycans from the mineralized matrix of the avian eggshell. Connect. Tissue Res. 1997, 36, 175–193. [Google Scholar] [CrossRef] [PubMed]
- Mann, K. Isolation of a glycosylated form of the chicken eggshell protein ovocleidin and determination of the glycosylation site. Alternative glycosylation/phosphorylation at an N-glycosylation sequon. FEBS Lett. 1999, 463, 12–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hincke, M.T.; St Maurice, M. Phosphorylation-dependent modulation of calcium carbonate precipitation by chicken eggshell matrix proteins. In Chemistry and Biology of Mineralized Tissues; Goldberg, M., Boskey, A., Robinson, C., Eds.; American Academy of Orthopaedic Surgeons: Rosemont, IL, USA, 2000; pp. 13–17. [Google Scholar]
- Gautron, J.; Hincke, M.T.; Panheleux, M.; Garcia-Ruiz, J.M.; Boldicke, T.; Nys, Y.Y. Ovotransferrin is a matrix protein of the hen eggshell membranes and basal calcified layer. Connect. Tissue Res. 2001, 42, 255–267. [Google Scholar] [CrossRef] [PubMed]
- Hincke, M.T.; Chien, Y.C.; Gerstenfeld, L.C.; McKee, M.D. Colloidal-gold immunocytochemical localization of osteopontin in Avian eggshell gland and eggshell. J. Histochem. Cytochem. 2008, 56, 467–476. [Google Scholar] [CrossRef] [Green Version]
- Pickford, M.; Mein, P.; Senut, B. Fossiliferous neogene karst fillings in Angola, Botswana and Namibia. S. Afr. J. Sci. 1994, 90, 227–230. [Google Scholar]
- Sahni, A.; Kumar, G.; Bajpai, S.; Srinivasan, S. Ultrastructure and taxonomy of ostrich eggshells from Upper Palaeolithic sites of India. J. Paleontol. Soc. India 1989, 34, 91–98. [Google Scholar]
- Jain, S.; Bajpai, S.; Kumar, G.; Pruthi, V. Microstructure, crystallography and diagenetic alteration in fossil ostrich eggshells from Upper Palaeolithic sites of Indian peninsular region. Micron 2016, 84, 72–78. [Google Scholar] [CrossRef]
- Roberts, P.; Henshilwood, C.S.; van Niekerk, K.L.; Keene, P.; Gledhill, A.; Reynard, J.; Badenhorst, S.; Lee-Thorp, J. Climate, environment and early human innovation: Stable isotope and faunal proxy evidence from archaeological sites (98-59ka) in the Southern Cape, South Africa. PLoS ONE 2016, 11, e0157408. [Google Scholar] [CrossRef] [Green Version]
- Miller, J.M.; Yiming, V.; Wang, Y.V. Ostrich eggshell beads reveal 50,000–year-old social network in Africa. Nature 2022, 601, 234–239. [Google Scholar] [CrossRef]
- Diehl, R.J.; Keller, H.M.; Hodgkins, J. Towards an interpretive framework for heated ostrich eggshell: An actualistic study. J. Archaeolog. Sci. Reports 2022, 43, 103465. [Google Scholar] [CrossRef]
- Mikhailov, K.E.; Zelenkov, N. The late Cenozoic history of the ostriches (Aves: Struthionidae), as revealed by fossil eggshell and bone remains. Earth-Sci. Rev. 2020, 208, 103270. [Google Scholar] [CrossRef]
- von Nathusius, W. Über die hüllen, welche den dotter des vogeleies umgeben. Z. Wissensch. Zool. 1868, 18, 225–270. [Google Scholar]
- Demarchi, B.; Hall, S.; Roncal-Herrero, T.; Freeman, C.L.; Woodley, J.; Crisp, M.K.; Collins, M.J. Protein sequences bound to mineral surfaces persist into deep time. eLife 2016, 5, e17092. [Google Scholar] [CrossRef]
- Richards, P.D.G.; Richards, P.A.; Lee, M.E. Ultrastructural characteristics of ostrich eggshell: Outer shell membrane and the calcified layers. J. S. Afr. Vet. Ass. 2000, 71, 97–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richards, P.D.G.; Botha, A.A.; Richards, P.A. Morphological and histochemical observations of the organic components of ostrich eggshell. J. S. Afri. Vet. Ass. 2002, 73, 13–22. [Google Scholar] [CrossRef] [Green Version]
- Szczerbinska, D.; Wiercinska, M. Ultrastructure of the eggshell of selected Palaeognathae species–a comparative analysis. Ann. Anim. Sci. 2014, 14, 167–178. [Google Scholar] [CrossRef] [Green Version]
- Kriesten, K.; Egge, H.; Faust, R. Lipids in the eggshell of the ostrich (Struthio camelus). Experientia 1979, 35, 1032–1033. [Google Scholar] [CrossRef]
- Mann, K.; Siedler, F. Ostrich (Struthio camelus) eggshell matrix contains two different C-type lectin-like proteins. Isolation, amino acid sequence, and posttranslational modifications. Biochim. Biophys. Acta 2004, 1696, 41–50. [Google Scholar] [CrossRef]
- Reyes-Grajeda, J.P.; Marin-Garcia, L.; Stojanoff, V.; Moreno, A. Purification, crystallization and preliminary X-ray analysis of struthiocalcin 1 from ostrich (Struthio camelus) eggshell. Acta Cryst. 2007, F63, 987–989. [Google Scholar] [CrossRef] [Green Version]
- Tyler, C.; Simkiss, K. A study of the eggshells of ratite birds. Zoology 1959, 133, 201–243. [Google Scholar]
- Arias, J.L.; Carrino, D.A.; Fernandez, M.S.; Rodriguez, J.P.; Dennis, J.E.; Caplan, A.I. Partial biochemical and immunochemical characterization of avian eggshell extracellular matrices. Arch. Biochem. Biophys. 1992, 298, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Hincke, M.T.; Nys, Y.; Gautron, J.; Mann, K.; Rodriguez-Navarro, A.B.; McKee, M. The eggshell: Structure, composition and mineralization. Frontiers Biosci. 2012, 17, 1266–1280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simkiss, K.; Tyler, C. A histochemical study of the organic matrix of hen eggshells. Quater. J. Micro. Sci. 1987, 98, 19–28. [Google Scholar]
- Perez-Huerta, A.; Cusack, M. Optimising electron backscatter diffraction (EBSD) of carbonate biominerals—Resin type and carbon thickness. Microsc. Microanal. 2009, 15, 197–203. [Google Scholar] [CrossRef] [Green Version]
- Perez-Huerta, A.; Dauphin, Y.; Cuif, J.P.; Cusack, M. High resolution electron backscatter diffraction (EBSD) data from calcite biominerals in recent gastropod shells. Micron 2011, 42, 246–251. [Google Scholar] [CrossRef]
- Solé, V.A.; Papillon, E.; Cotte, M.; Walter, P.; Susini, J. A multiplatform code for the analysis of energy-dispersive X-ray fluorescence spectra. Spectrochim. Acta Part B 2007, B62, 63–68. [Google Scholar] [CrossRef]
- Kim, B.; Gautier, M.; Rivard, C.; Sanglar, C.; Michel, P.; Gourdon, R. Effect of aging on phosphorus speciation in surface deposit of a vertical flow constructed wetland. Environ. Sci. Technol. 2015, 49, 4903–4910. [Google Scholar] [CrossRef] [Green Version]
- Ravel, B.; Newville, M. Athena, Artemis, Hephaestus: Data analysis for X-ray absorption spectroscopy using IFEFFIT. J. Synchr. Rad. 2005, 12, 537–541. [Google Scholar] [CrossRef] [Green Version]
- Stock, S.R.; Veis, A.; Xiao, X.; Almer, J.; Dorvee, J.R. Sea urchin tooth mineralization: Calcite present early in the aboral plumula. J. Struct. Biol. 2012, 180, 280–289. [Google Scholar] [CrossRef] [Green Version]
- Brandes, J.A.; Ingall, E.; Paterson, D. Characterization of minerals and organic phosphorus species in marine sediments using soft X-ray fluorescence spectromicroscopy. Mar. Chem. 2007, 103, 250–265. [Google Scholar] [CrossRef]
- Ingall, E.D.; Brandes, J.A.; Diaz, J.M.; de Jonge, M.D.; Paterson, D.; McNulty, I.; Elliott, W.C.; Northrup, P. Phosphorus K-edge XANES spectroscopy of mineral standards. J. Synchr. Rad. 2011, 18, 189–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willoughby, W.; Steyn, L.; Bam, L.; Oliver, A.J.; Devey, R.; Maina, J.N. Micro-focus X-Ray tomography study of the microstructure and morphometry of the eggshell of Ostriches (Struthio Camerus). Anat. Rec. 2016, 299, 1015–1026. [Google Scholar] [CrossRef] [Green Version]
- Chien, Y.C.; Hincke, M.T.; Vali, H.; McKee, M.D. Ultrastructural matrix mineral relationships in avian eggshell, and effects of osteopontin on calcite growth in vitro. J. Struct. Biol. 2008, 163, 84–99. [Google Scholar] [CrossRef]
- Becking, J.H. The ultrastructure of the avian eggshell. Ibis 1975, 117, 143–151. [Google Scholar] [CrossRef]
- Athanasiadou, D.; Jiang, W.; Goldbaum, D.; Saleem, A.; Basu, K.; Pacella, M.S.; Böhm, C.F.; Chromik, R.E.; Hincke, M.T.; Rodríguez-Navarro, A.B.; et al. Nanostructure, osteopontin, and mechanical properties of calcitic avian eggshell. Sci. Adv. 2018, 4, eaar3219. [Google Scholar] [CrossRef] [Green Version]
- Mikhailov, K.E. The principal structure of the avian eggshell: Data of SEM studies. Acta Zool. Cracov. 1987, 30, 53–70. [Google Scholar]
- Chien, Y.C.; Hincke, M.T.; McKee, M.D. Avian Eggshell Structure and Osteopontin. Cells Tissues Organs 2009, 189, 38–43. [Google Scholar] [CrossRef]
- Haugstad, G. Atomic Force Microscopy: Understanding Basic Modes and Advanced Applications; John Wiley and Sons: Hoboken, NJ, USA, 2012. [Google Scholar]
- Mittal, V.; Matsko, N.B. Analytical Imaging Techniques for Soft Matter Characterization; Springer Science and Business Media: New York, NY, USA, 2012. [Google Scholar]
- Pérez-Huerta, A.; Dauphin, Y. Comparison of the structure, crystallography and composition of eggshells of the guinea fowl and graylag goose. Zoology 2016, 119, 52–63. [Google Scholar] [CrossRef] [PubMed]
- Masshoff, W.; Stolpmann, H.J. Licht-und Elektronenmikroscopische Untersuchungen and der Schalenaut und Kalschale des Hühnereies. Z. Zellforsch. Mikrosk. Anat. 1961, 55, 818–832. [Google Scholar] [CrossRef]
- Simons, P.C.M.; Wiertz, G. Notes on the structure of membranes and shell in the hen’egg–an electron microscopical study. Z. Zellforsch. 1963, 59, 555–557. [Google Scholar] [CrossRef]
- Tan, C.K.; Chen, T.W.; Chan, H.L.; Ng, L.S. A scanning and transmission electron microscopic study of the membranes of chicken egg. Histol. Histopath. 1992, 7, 339–345. [Google Scholar]
- Böttcher, M.E.; Gehlken, P.L.; Steele, D.F. Characterization of inorganic and biogenic magnesian calcites by Fourier transform infrared spectroscopy. Solid State Ionics 1997, 101, 1379–1385. [Google Scholar] [CrossRef]
- Dauphin, Y. Infrared spectra and elemental composition in recent biogenic calcites: Relationships between the n4 band wavelength and Sr and Mg concentrations. Appl. Spectrosc. 1999, 53, 184–190. [Google Scholar] [CrossRef]
- Beniash, E.; Aizenberg, J.; Addadi, L.; Weiner, S. Amorphous calcium carbonate transforms into calcite during sea urchin larval spicule growth. Proc. R. Soc. Lond. 1997, 264, 461–465. [Google Scholar] [CrossRef]
- Rodriguez-Navarro, A.B.; Marie, P.; Nys, Y.; Hincke, M.T.; Gautron, J. Amorphous calcium carbonate controls avian eggshell mineralization: A new paradigm for understanding rapid eggshell calcification. J. Struct. Biol. 2015, 190, 291–303. [Google Scholar] [CrossRef]
- Jones, G.C.; Jackson, B. Infrared Transmission Spectra of Carbonate Minerals; Chapman & Hall: London, UK, 1993; p. 233. [Google Scholar]
- Mann, K.; Mann, M. The proteome of the calcified layer organic matrix of turkey (Meleagris gallopavo) eggshell. Proteome Sci. 2013, 11, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalbeck, P.; Cusack, M. Crystallography (Electron Backscatter Diffraction) and Chemistry (Electron Probe Microanalysis) of the Avian Eggshell. Cryst. Growth Design. 2006, 6, 2558–2562. [Google Scholar] [CrossRef]
- Cain, C.J.; Heyn, A.N.J. X-ray Diffraction Studies of the Crystalline Structure of the Avian Egg Shell. Biophys. J. 1964, 4, 23–39. [Google Scholar] [CrossRef] [Green Version]
- Silyn-Roberts, H.; Sharp, R.M. Crystal growth and the role of the organic network in eggshells. Proc. R. Soc. Lond. 1986, 227, 303–324. [Google Scholar]
- Heredia, A.; Rodriguez-Hernandez, A.G.; Lozano, L.F.; Pena-Rico, M.A.; Velazquez, R.; Basiuk, V.A.; Bucio, L. Microstructure and thermal change of texture of calcite crystals in ostrich eggshell Struthio camelus. Mater. Sci. Eng. 2005, C25, 1–9. [Google Scholar] [CrossRef]
- Eagle, R.A.; Enriquez, M.; Grellet-Tinner, G.; Pérez-Huerta, A.; Hu, D.; Tütken, T.; Eiler, J.M. Isotopic ordering in eggshells reflects body temperatures and suggest differing thermophysiology in two Cretaceous dinosaurs. Nat. Commun. 2015, 6, 8296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bischoff, W.D.; Sharma, S.K.; MacKenzie, F.T. Carbonate ion disorder in synthetic and biogenic magnesian calcites: A Raman spectral study. Am. Mineral. 1985, 70, 581–589. [Google Scholar]
- Masic, A.; Weaver, J.C. Large area sub-micron chemical imaging of magnesium in sea urchin teeth. J. Struct. Biol. 2015, 189, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Thomas, D.B.; Hauber, M.E.; Hanley, D.; Waterhouse, F.I.N.; Fraser, S.; Gordon, K.C. Analysing avian eggshell pigments with Raman spectroscopy. J. Exper. Biol. 2015, 218, 2670–2674. [Google Scholar] [CrossRef] [Green Version]
- Wedral, E.M.; Vadehra, D.V.; Baker, R.C. Chemical composition of the cuticle, and the inner and outer shell membranes from eggs of Gallus gallus. Comp. Biochem. Physiol. 1974, B47, 361–640. [Google Scholar] [CrossRef]
- Cusack, M.; Fraser, A.C.; Stachel, T. Magnesium and phosphorus distribution in the avian eggshell. Comp. Biochem. Physiol. 2003, B134, 63–69. [Google Scholar] [CrossRef]
- Hahn, E.N.; Sherman, V.R.; Pissarenko, A.; Rohrbach, S.D.; Fernandes, D.J.; Meyers, M.A. Nature’s technical ceramic: The avian eggshell. J. R. Soc. Interface 2017, 14, 20160804. [Google Scholar] [CrossRef] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mg/Na | Mg/Sr | Mg/P | Sr/Na | Sr/P | Na/P | |||
|---|---|---|---|---|---|---|---|---|
| Person-R-values | 0.82 | 0.81 | 0.95 | 0.8 | 0.82 | 0.78 | ||
| Spearman | 0.55 | 0.64 | 0.93 | 0.45 | 0.68 | 0.51 | ||
| Manders M1 | 1 | 0.96 | 0.98 | 1 | 1 | 1 | ||
| Manders M2 | 0.999 | 1 | 1 | 0.935 | 0.985 | 0.96 | ||
| Kendall | 0.4 | 0.48 | 0.79 | 0.33 | 0.51 | 0.37 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez-Huerta, A.; Brugal, J.-P.; Salomé, M.; Schmitt, C.N.Z.; Dauphin, Y. Integrated Information on the Structure and Composition of the Ostrich Eggshell (Struthio camelus). Minerals 2023, 13, 481. https://doi.org/10.3390/min13040481
Pérez-Huerta A, Brugal J-P, Salomé M, Schmitt CNZ, Dauphin Y. Integrated Information on the Structure and Composition of the Ostrich Eggshell (Struthio camelus). Minerals. 2023; 13(4):481. https://doi.org/10.3390/min13040481
Chicago/Turabian StylePérez-Huerta, Alberto, Jean-Philip Brugal, Murielle Salomé, Clemens N. Z. Schmitt, and Yannicke Dauphin. 2023. "Integrated Information on the Structure and Composition of the Ostrich Eggshell (Struthio camelus)" Minerals 13, no. 4: 481. https://doi.org/10.3390/min13040481
APA StylePérez-Huerta, A., Brugal, J. -P., Salomé, M., Schmitt, C. N. Z., & Dauphin, Y. (2023). Integrated Information on the Structure and Composition of the Ostrich Eggshell (Struthio camelus). Minerals, 13(4), 481. https://doi.org/10.3390/min13040481