

Imaging of Ancient Microbial Biomarkers within Miocene Dolomite (Kuwait) Using Time-of-Flight Secondary Ion Mass Spectrometry
Abstract
:1. Introduction
2. Materials and Methods
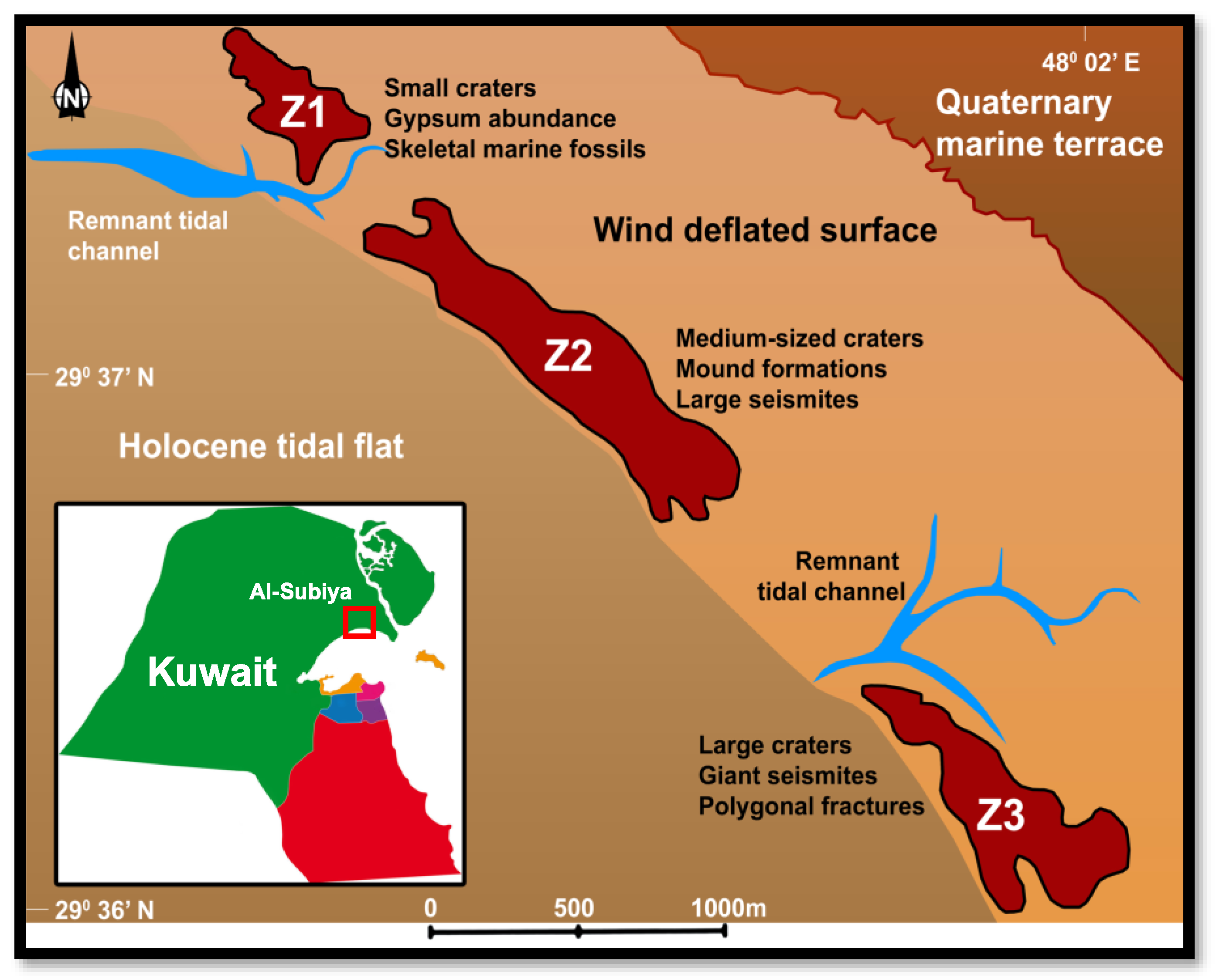
2.1. Sample Materials
2.2. Time-of-Flight–Secondary Ion Mass Spectrometry (ToF-SIMS) Analysis
3. Results
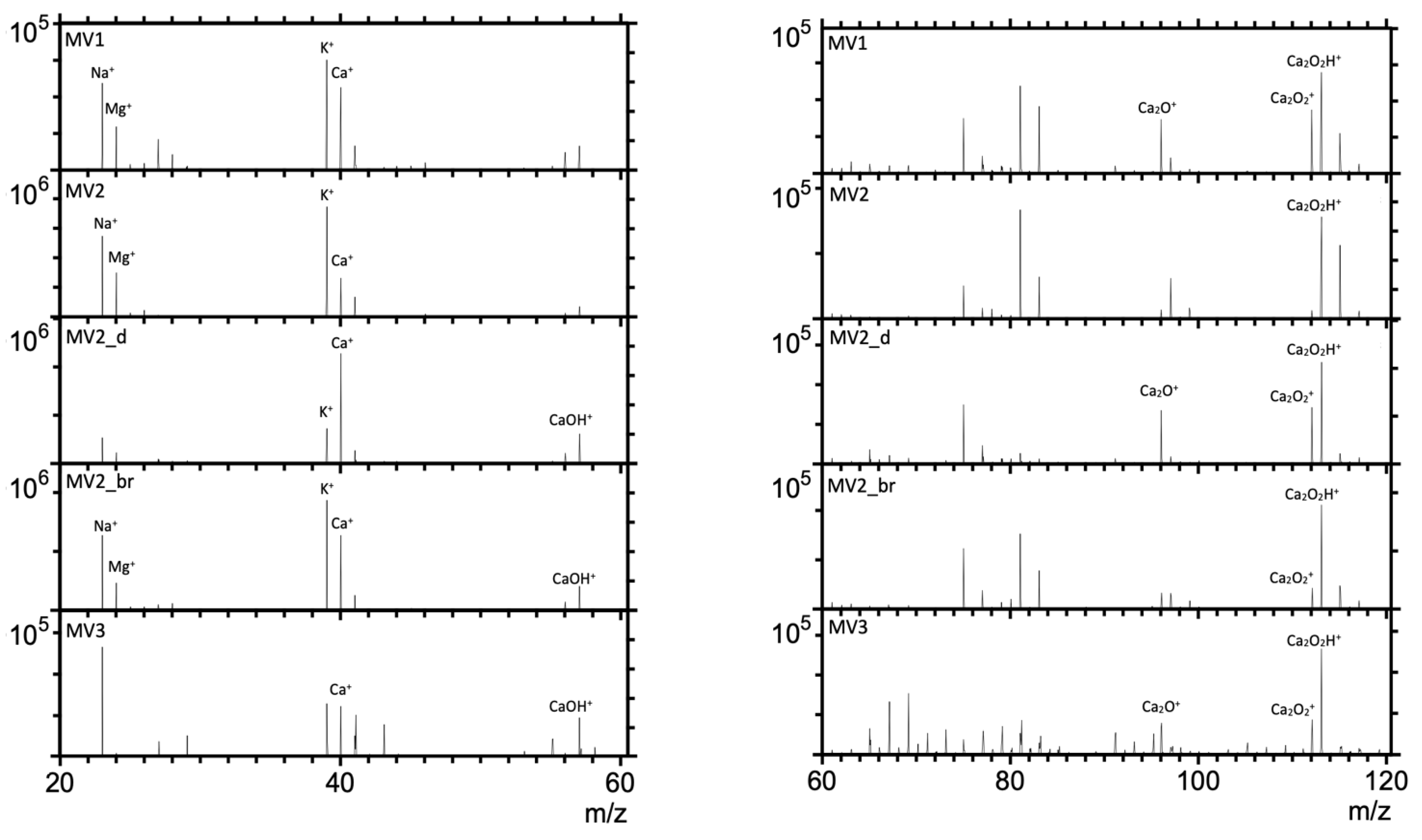
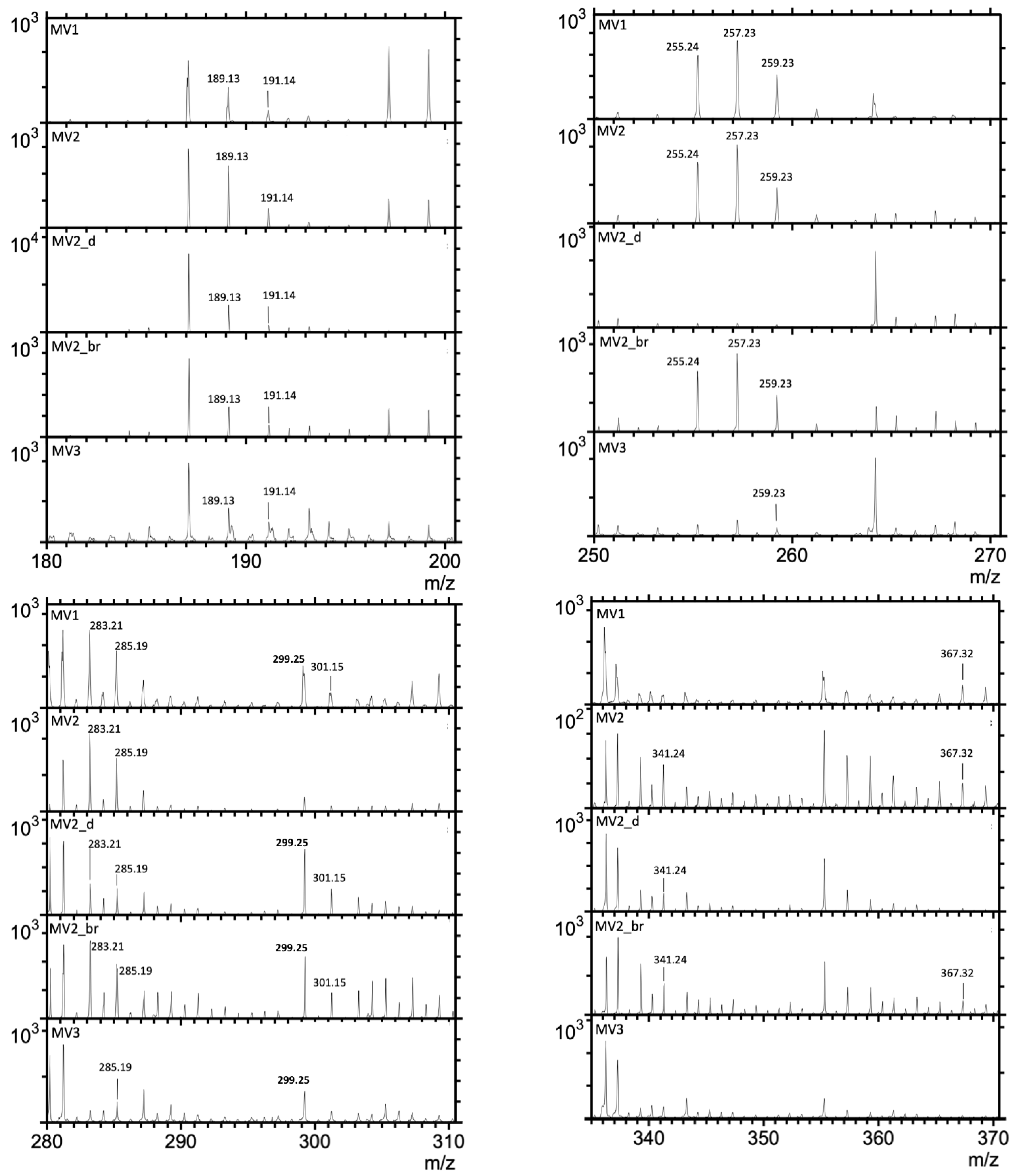
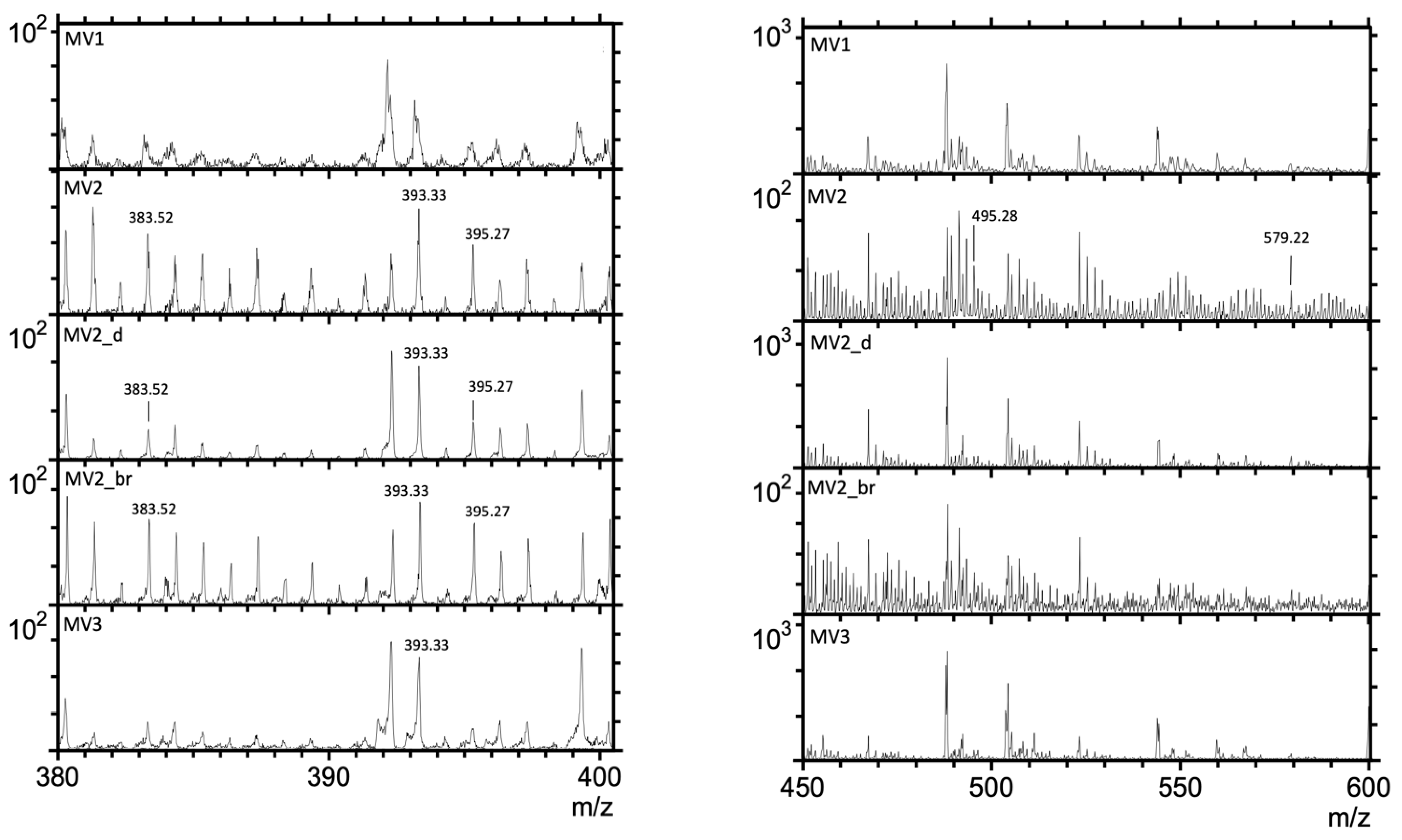
3.1. Time-of-Flight–Secondary Ion Mass Spectrometry (ToF-SIMS) Spectra
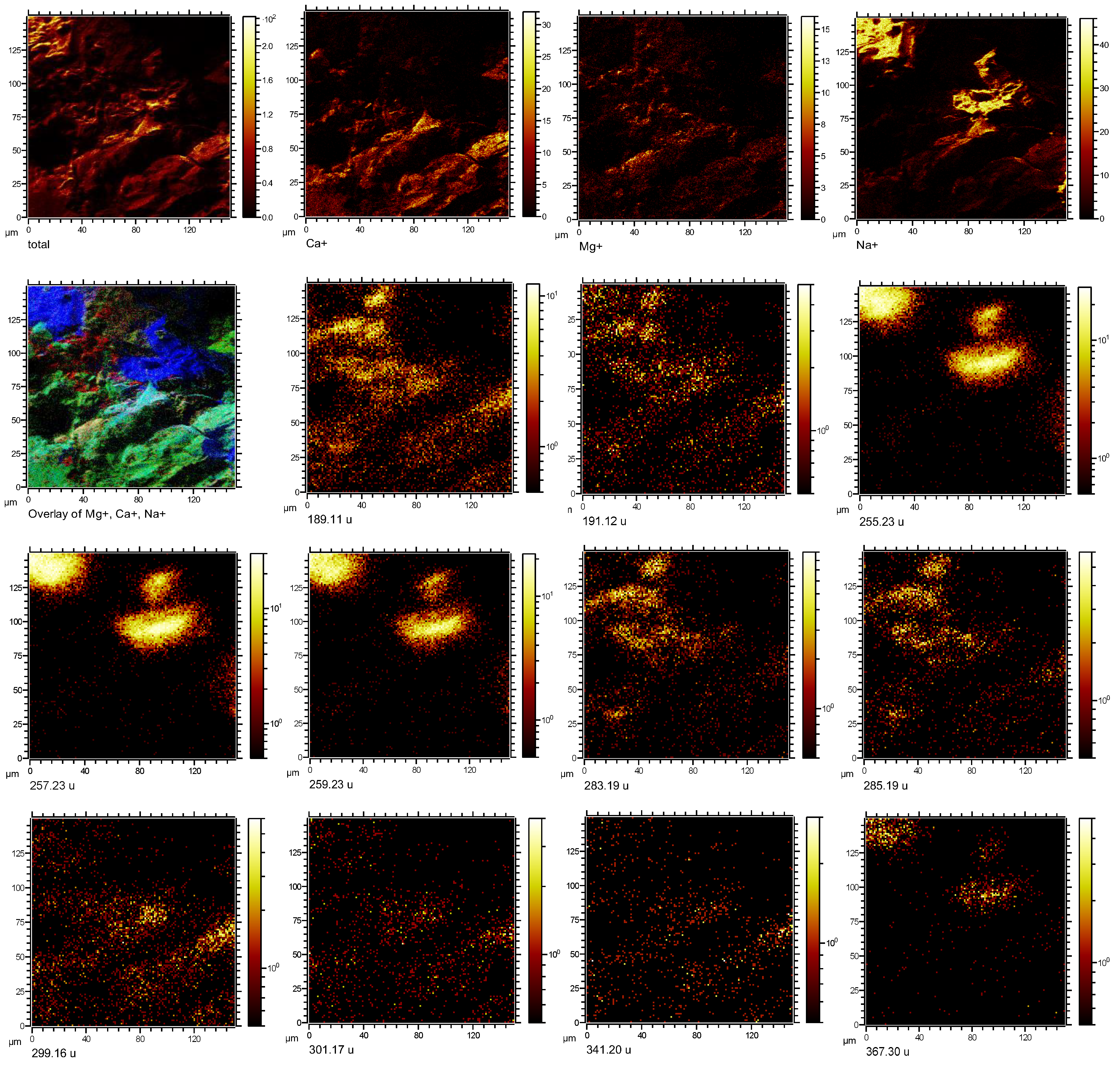
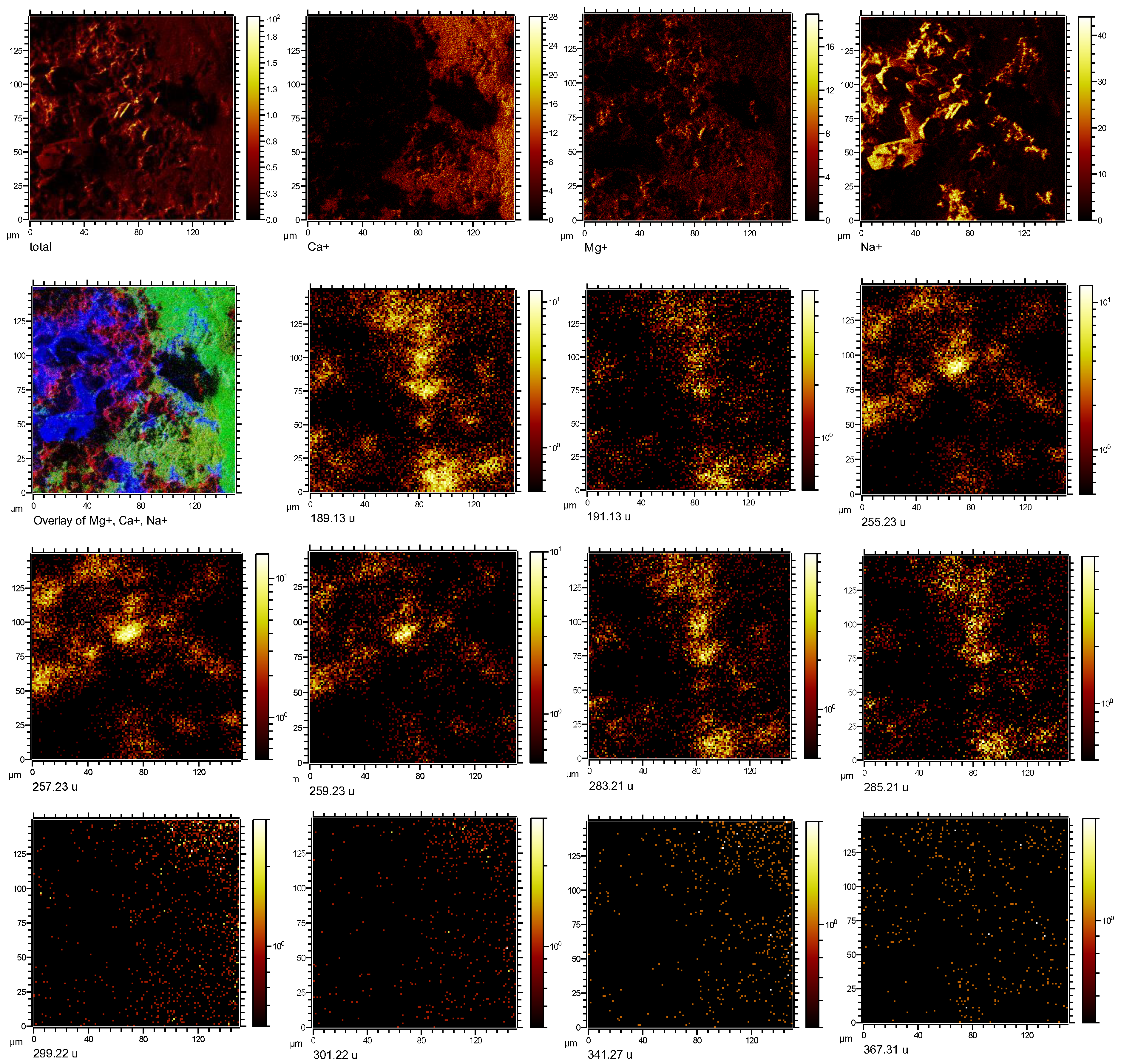
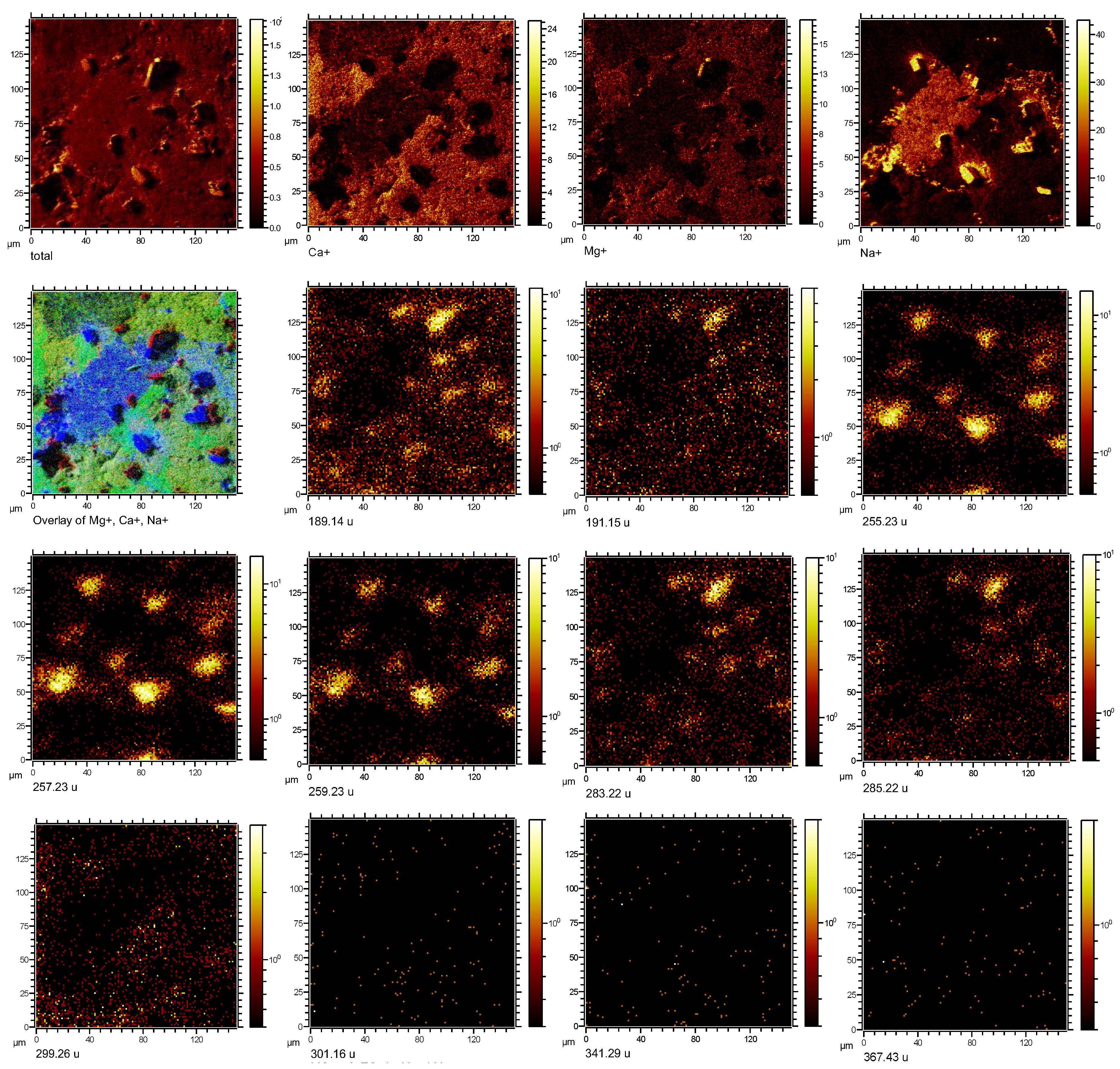
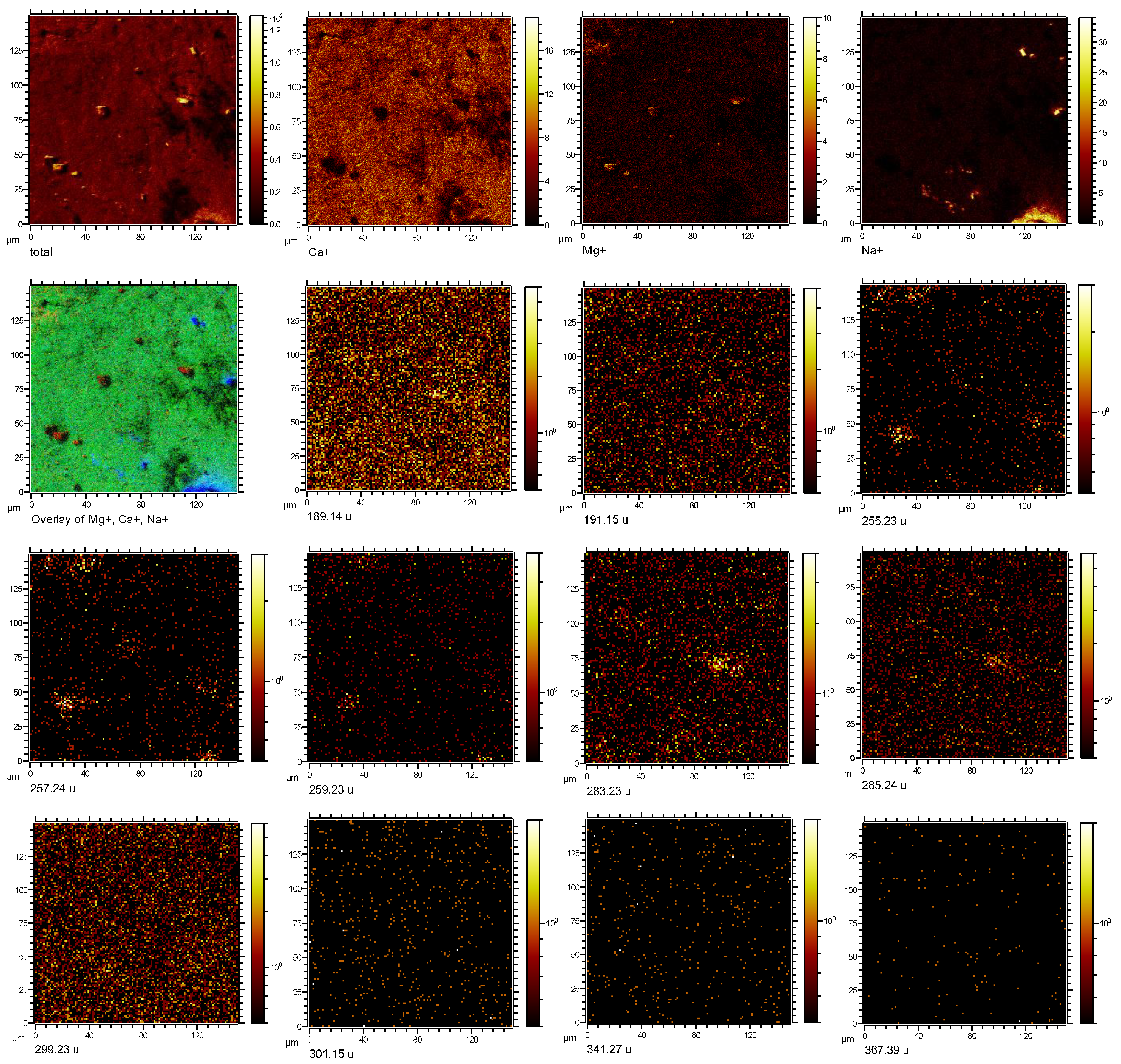
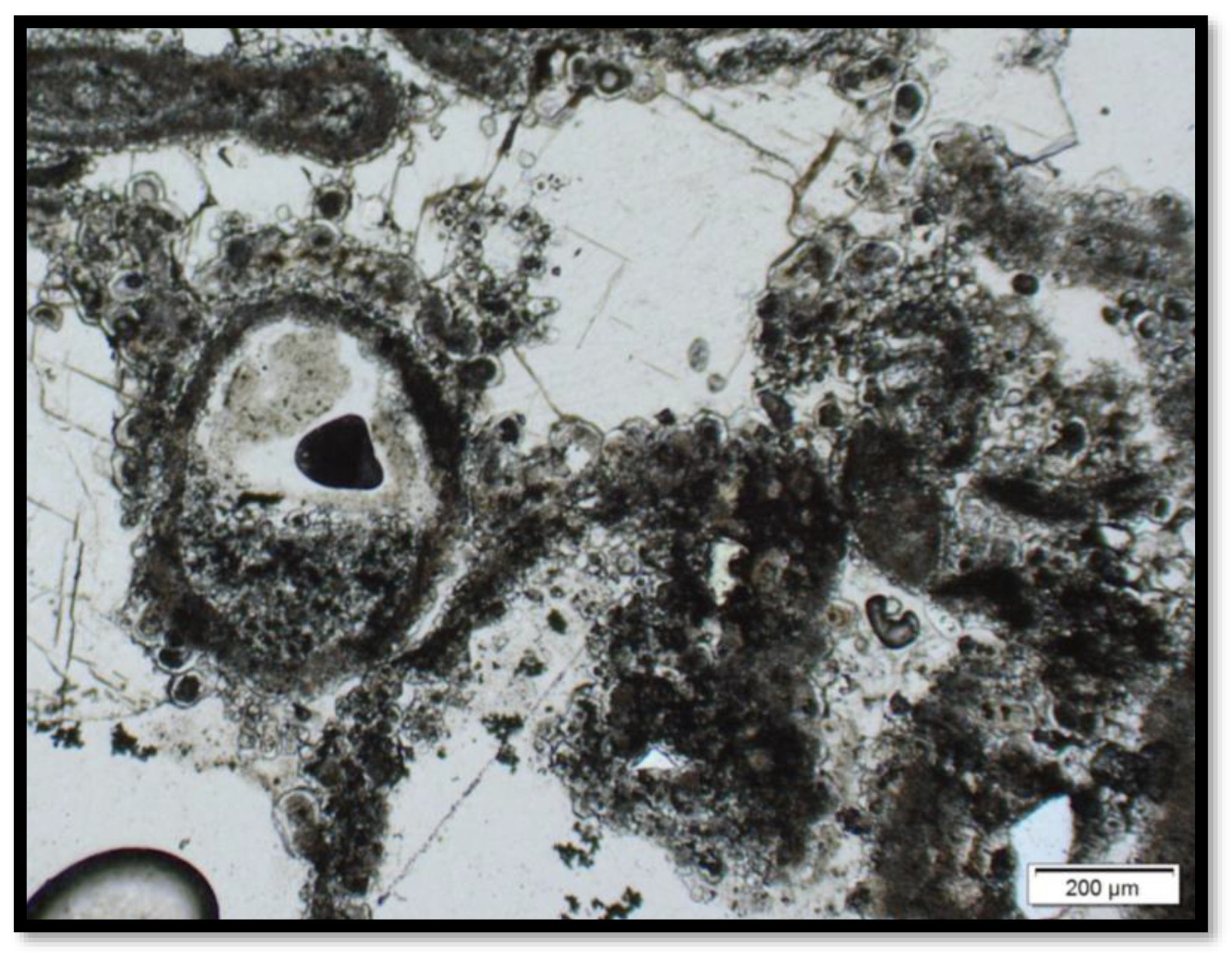
3.2. Imaging Microbial Biomarkers
4. Discussion
4.1. Microbial Biomarkers Related to Hydrocarbon Seepage
4.2. Co-Localization of Microbial Biomarkers within Dolomite
4.3. Beyond ToF-SIMS Microscale View
4.4. Significance and Implication
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Milkov, A.V. Worldwide distribution of submarine mud volcanoes and associated gas hydrates. Mar. Geol. 2000, 167, 29–42. [Google Scholar] [CrossRef]
- Mellors, R.; Kilb, D.; Aliyev, A.; Gasanov, A.; Yetirmishli, G. Correlations between earthquakes and large mud volcano eruptions. J. Geophys. Res. Solid Earth 2007, 112, 1–11. [Google Scholar] [CrossRef]
- Kopf, A.J. Significance of mud volcanism. Rev. Geophys. 2002, 40, 1–52. [Google Scholar] [CrossRef] [Green Version]
- Elvert, M.; Suess, E.; Greinert, J.; Whiticar, M.J. Archaea mediating anaerobic methane oxidation in deep-sea sediments at cold seeps of the eastern Aleutian subduction zone. Org. Geochem. 2000, 31, 1175–1187. [Google Scholar] [CrossRef]
- Boetius, A.; Ravenschlag, K.; Schubert, C.J.; Rickert, D.; Widdel, F.; Gieseke, A.; Amann, R.; Jùrgensen, B.B.; Witte, U.; Pfannkuche, O. A marine microbial consortium apparently mediating anaerobic oxidation of methane. Nature 2000, 407, 623–626. [Google Scholar] [CrossRef]
- Reeburgh, W.S. Oceanic Methane Biogeochemistry. Chem. Rev. 2007, 107, 486–513. [Google Scholar] [CrossRef] [Green Version]
- Ussler, W.; Paull, C.K. Rates of anaerobic oxidation of methane and authigenic carbonate mineralization in methane-rich deep-sea sediments inferred from models and geochemical profiles. Earth Planet. Sci. Lett. 2008, 266, 271–287. [Google Scholar] [CrossRef]
- Aloisi, G.; Pierrea, C.; Rouchyb, J.-M.; Faugeres, J.-C. Isotopic evidence of methane-related diagenesis in the mud volcanic sediments of the Barbados Accretionary Prism. Cont. Shelf Res. 2002, 22, 2355–2372. [Google Scholar] [CrossRef]
- Chen, H.; Wang, S.; Chen, Z.; Yan, W.; Li, G. Geochemical and magnetic signals for the mud volcano-induced methane seepage in the core sediments of Shenhu area, northern South China Sea. Environ. Earth Sci. 2014, 73, 6365–6378. [Google Scholar] [CrossRef]
- Hu, Y.; Feng, D.; Peckmann, J.; Roberts, H.H.; Chen, D. New insights into cerium anomalies and mechanisms of trace metal enrichment in authigenic carbonate from hydrocarbon seeps. Chem. Geol. 2014, 381, 55–66. [Google Scholar] [CrossRef]
- Kioka, A.; Tsuji, T.; Otsuka, H.; Ashi, J. Methane Concentration in Mud Conduits of Submarine Mud Volcanoes: A Coupled Geochemical and Geophysical Approach. Geochem. Geophys. Geosyst. 2019, 20, 792–813. [Google Scholar] [CrossRef] [Green Version]
- Omrani, H.; Raghimi, M. Origin of the mud volcanoes in the south east Caspian Basin, Iran. Mar. Pet. Geol. 2018, 96, 615–626. [Google Scholar] [CrossRef]
- Duane, M.J.; Reinink-Smith, L.; Eastoe, C.; Al-Mishwat, A.T. Mud volcanoes and evaporite seismites in a tidal flat of northern Kuwait—Implications for fluid flow in sabkhas of the Persian (Arabian) Gulf. Geo-Mar. Lett. 2015, 35, 237–246. [Google Scholar] [CrossRef]
- Warren, J. Dolomite: Occurrence, evolution and economically important associations. Earth-Sci. Rev. 2000, 52, 1–81. [Google Scholar] [CrossRef]
- Al-Awadi, M.; Clark, W.J.; Moore, W.R.; Herron, M.; Zhang, T.; Zhao, W.; Hurley, N.; Kho, D.; Montaron, B.; Sadooni, F. Dolomite: Perspectives on a Perplexing Mineral. Oilfield Rev. 2009, 21, 32–45. [Google Scholar]
- Kaczmarek, S.E.; Gregg, J.M.; Bish, D.L.; Machel, H.G.; Fouke, B.W. Dolomite, very high-magnesium calcite, and microbes- implications for the microbial model of dolomitization. In Characterization and Modeling of Carbonates–Mountjoy Symposium 1; Macneil, A.J., Lonnee, J., Wood, R., Eds.; SEPM Special Publication No. 109; SEPM Society for Sedimentary Geology: Broken Arrow, OK, USA, 2017; pp. 7–20. [Google Scholar]
- van Lith, Y.; Warthmann, R.; Vasconcelos, C.; Mckenzie, J.A. Microbial fossilization in carbonate sediments: A result of the bacterial surface involvement in dolomite precipitation. Sedimentology 2003, 50, 237–245. [Google Scholar] [CrossRef]
- Qiu, X.; Wang, H.; Yao, Y.; Duan, Y. High salinity facilitates dolomite precipitation mediated by Haloferax volcanii DS52. Earth Planet. Sci. Lett. 2017, 472, 197–205. [Google Scholar] [CrossRef]
- Kenward, P.A.; Fowle, D.A.; Goldstein, R.H.; Ueshima, M.; González, L.A.; Roberts, J.A. Ordered low-temperature dolomite mediated by carboxyl-group density of microbial cell walls. AAPG Bull. 2013, 97, 2113–2125. [Google Scholar] [CrossRef]
- Sánchez-Román, M.; Vasconcelos, C.; Schmid, T.; Dittrich, M.; McKenzie, J.A.; Zenobi, R.; Rivadeneyra, M.A. Aerobic microbial dolomite at the nanometer scale: Implications for the geologic record. Geology 2008, 36, 879–882. [Google Scholar] [CrossRef]
- Bontognali, T.R.R.; McKenzie, J.A.; Warthmann, R.J.; Vasconcelos, C. Microbially influenced formation of Mg-calcite and Ca-dolomite in the presence of exopolymeric substances produced by sulphate-reducing bacteria. Terra Nova 2014, 26, 72–77. [Google Scholar] [CrossRef]
- Alibrahim, A.; Al-Gharabally, D.; Mahmoud, H.; Dittrich, M. Proto-dolomite formation in microbial consortia dominated by Halomonas strains. Extremophiles 2019, 23, 765–781. [Google Scholar] [CrossRef] [PubMed]
- Paulo, C.; Dittrich, M. 2D Raman spectroscopy study of dolomite and cyanobacterial extracellular polymeric substances from Khor Al-Adaid sabkha (Qatar). J. Raman Spectrosc. 2013, 44, 1563–1569. [Google Scholar] [CrossRef]
- Diloreto, Z.A.; Garg, S.; Bontognali, T.R.R.; Dittrich, M. Modern dolomite formation caused by seasonal cycling of oxygenic phototrophs and anoxygenic phototrophs in a hypersaline sabkha. Sci. Rep. 2021, 11, 4170. [Google Scholar] [CrossRef]
- Gunatilaka, A. Dolomite formation in coastal Al-Khiran, Kuwait Arabian Gulf—A re-examination of the sabkha model. Sediment. Geol. 1991, 72, 35–53. [Google Scholar] [CrossRef]
- Mauger, C.L.; Compton, J.S. Formation of modern dolomite in hypersaline pans of the Western Cape, South Africa. Sedimentology 2011, 58, 1678–1692. [Google Scholar] [CrossRef]
- Sánchez-Román, M.; Vasconcelos, C.; Warthmann, R.; Rivadeneyra, M.; McKenzie, J.A. Microbial Dolomite Precipitation under Aerobic Conditions: Results from Brejo do Espinho Lagoon (Brazil) and Culture Experiments; International Association of Sedimentologists Special Publications; Blackwell Publishing Ltd.: Oxford, UK, 2009; Volume 41, pp. 167–178. [Google Scholar]
- Diver, W.L. Precambrian microfossils of Carpentarian age from Bungle Bungle dolomite of western Australia. Nature 1974, 247, 361–363. [Google Scholar] [CrossRef]
- Vasconcelos, C.; McKenzie, J.A. Dolomite as a Biomineral and Possible Implications. Rev. Soc. Españ. Mineral. 2008, 9, 21–22. [Google Scholar]
- Li, J.; Benzerara, K.; Bernard, S.; Beyssac, O. The link between biomineralization and fossilization of bacteria: Insights from field and experimental studies. Chem. Geol. 2013, 359, 49–69. [Google Scholar] [CrossRef]
- Aquilina, A.; Knab, N.J.; Knittel, K.; Kaur, G.; Geissler, A.; Kelly, S.P.; Fossing, H.; Boot, C.S.; Parkes, R.J.; Mills, R.A.; et al. Biomarker indicators for anaerobic oxidizers of methane in brackish-marine sediments with diffusive methane fluxes. Org. Geochem. 2010, 41, 414–426. [Google Scholar] [CrossRef]
- Bouloubassi, I.; Nabais, E.; Pancost, R.D.; Lorre, A.; Taphanel, M.-H. First biomarker evidence for methane oxidation at cold seeps in the Southeast Atlantic (REGAB pockmark). Deep. Sea Res. Part II Top. Stud. Oceanogr. 2009, 56, 2239–2247. [Google Scholar] [CrossRef]
- Chevalier, N.; Bouloubassi, I.; Birgel, D.; Crémière, A.; Taphanel, M.-H.; Pierre, C. Authigenic carbonates at cold seeps in the Marmara Sea (Turkey): A lipid biomarker and stable carbon and oxygen isotope investigation. Mar. Geol. 2011, 288, 112–121. [Google Scholar] [CrossRef]
- Gontharet, S.; Stadnitskaia, A.; Bouloubassi, I.; Pierre, C.; Damsté, J.S.S. Palaeo methane-seepage history traced by biomarker patterns in a carbonate crust, Nile deep-sea fan (Eastern Mediterranean Sea). Mar. Geol. 2009, 261, 105–113. [Google Scholar] [CrossRef]
- Pancost, R.D.; Hopmans, E.C.; Damsté, J.S.S. Archaeal lipids in Mediterranean Cold Seeps- Molecular proxies for anaerobic methane oxidation. Geochim. Cosmochim. Acta. 2001, 65, 1611–1627. [Google Scholar] [CrossRef] [Green Version]
- Stadnitskaia, A.; Nadezhkin, D.; Abbas, B.; Blinova, V.; Ivanov, M.K.; Sinninghe Damsté, J.S. Carbonate formation by anaerobic oxidation of methane: Evidence from lipid biomarker and fossil 16S rDNA. Geochim. Et Cosmochim. Acta 2008, 72, 1824–1836. [Google Scholar] [CrossRef]
- Zhang, Z.-X.; Li, J.; Chen, Z.; Sun, Z.; Yang, H.; Fu, M.; Peng, X. The effect of methane seeps on the bacterial tetraether lipid distributions at the Okinawa Trough. Mar. Chem. 2020, 225, 103845. [Google Scholar] [CrossRef]
- Benninghoven, A. Chemical analysis of inorganic and organic surfaces and thin films by static time-of-flight secondary ion mass spectrometry (TOF-SIMS). Angew. Chem. Int. Ed. 1994, 33, 1023–1043. [Google Scholar] [CrossRef]
- Sodhi, R.N. Time-of-flight secondary ion mass spectrometry (TOF-SIMS):—Versatility in chemical and imaging surface analysis. Analyst 2004, 129, 483–487. [Google Scholar] [CrossRef]
- Thiel, V.; Heim, C.; Arp, G.; Hahmann, U.; SjÖVall, P.; Lausmaa, J. Biomarkers at the microscopic range: ToF-SIMS molecular imaging of Archaea-derived lipids in a microbial mat. Geobiology 2007, 5, 413–421. [Google Scholar] [CrossRef]
- Leefmann, T.; Heim, C.; Kryvenda, A.; Siljeström, S.; Sjövall, P.; Thiel, V. Biomarker imaging of single diatom cells in a microbial mat using time-of-flight secondary ion mass spectrometry (ToF-SIMS). Org. Geochem. 2013, 57, 23–33. [Google Scholar] [CrossRef]
- Steelea, A.; Toporskia, J.K.W.; Avcic, R.; Guidryd, S.; McKayb, D.S. Time of flight secondary ion mass spectrometry (ToFSIMS) of a number of hopanoids. Org. Geochem. 2001, 32, 905–911. [Google Scholar] [CrossRef]
- Leefmann, T.; Heim, C.; Siljestrom, S.; Blumenberg, M.; Sjovall, P.; Thiel, V. Spectral characterization of ten cyclic lipids using time-of-flight secondary ion mass spectrometry. Rapid Commun. Mass Spectrom. 2013, 27, 565–581. [Google Scholar] [CrossRef]
- Sjövall, P.; Thiel, V.; Siljeström, S.; Heim, C.; Hode, T.; Lausmaa, J. Organic Geochemical Microanalysis by Time-of-FlightSecondary Ion Mass Spectrometry (ToF-SIMS). Geostand. Geoanal. Res. 2008, 32, 267–277. [Google Scholar] [CrossRef]
- Gunatilaka, A.; Al-Zamel, A.; Shearman, D.J.; Reda, A. A Spherulitic Fabric in Selectively Dolomitized Siliciclastic Crustacean Burrows, Northern Kuwait. J. Sediment. Petrol. 1987, 57, 922–927. [Google Scholar]
- Gunatilaka, A. Spheroidal dolomites—Origin by hydrocarbon seepage? Sedimentology 1989, 36, 701–710. [Google Scholar] [CrossRef]
- Alibrahim, A.; Duane, M.J.; Dittrich, M. Dolomite genesis in bioturbated marine zones of an early-middle Miocene coastal mud volcano outcrop (Kuwait). Sci. Rep. 2021, 11, 6636. [Google Scholar] [CrossRef]
- Alrashidi, A.F. Field Map and Geochemistry of Miocene Mud Volcanoes (North of Kuwait). Kuwait, Kuwait. 2023. [Google Scholar]
- Behbehani, S.; Hollis, C.; Holland, G.; Singh, P.; Edwards, K. A seismically controlled seal breach in a major hydrocarbon province: A study from the Mauddud Formation in the Bahrah field, Kuwait. Mar. Pet. Geol. 2019, 107, 255–277. [Google Scholar] [CrossRef]
- Drake, H.; Astrom, M.E.; Heim, C.; Broman, C.; Astrom, J.; Whitehouse, M.; Ivarsson, M.; Siljestrom, S.; Sjovall, P. Extreme 13C depletion of carbonates formed during oxidation of biogenic methane in fractured granite. Nat. Commun. 2015, 6, 7020. [Google Scholar] [CrossRef] [Green Version]
- Al-Hajeri, M.M.; Amer, A.; Djawair, D.; Green, D.; Al-Naqi, M. Origin of enigmatic sand injectite outcrops associated with non-tectonic forced-folding structure in Bahrah area, northern Kuwait Bay. Mar. Pet. Geol. 2020, 115, 104268. [Google Scholar] [CrossRef]
- Yadav, L.D.S. Mass Spectroscopy (MS). In Organic Spectroscopy; Springer Science + Business Media, B.V.: Berlin/Heidelberg, Germany, 2005; pp. 250–294. [Google Scholar]
- Stephan, T. TOF-SIMS in cosmochemistry. Planet. Space Sci. 2001, 49, 859–906. [Google Scholar] [CrossRef]
- Siljestrom, S.; Lausmaa, J.; Sjovall, P.; Broman, C.; Thiel, V.; Hode, T. Analysis of hopanes and steranes in single oil-bearing fluid inclusions using time-of-flight secondary ion mass spectrometry (ToF-SIMS). Geobiology 2010, 8, 37–44. [Google Scholar] [CrossRef]
- Venosa, A.; Suidan, M.; King, D.; Wrenn, B. Use of hopane as a conservative biomarker for monitoring the bioremediation effectiveness of crude oil contaminating a sandy beach. J. Ind. Microbiol. Biotechnol. 1997, 18, 131–139. [Google Scholar] [CrossRef]
- Hinrichs, K.-U.; Hayes, J.M.; Sylva, S.P.; Brewer, P.G.; DeLong, E.F. Methane-consuming archaebacteria in marine sediments. Nature 1999, 398, 802–805. [Google Scholar] [CrossRef] [PubMed]
- Peckmann, J.; Thiel, V. Carbon cycling at ancient methane–seeps. Chem. Geol. 2004, 205, 443–467. [Google Scholar] [CrossRef]
- Langworthy, T.A.; Holzer, G.; Zeikus, J.G.; Tornabene, T.G. Iso- and Anteiso-Branched Glycerol Diethers of the Thermophilic Anaerobe Thermodesulfotobacterium commune. Syst. Appl. Microbiol. 1983, 4, 1–17. [Google Scholar] [CrossRef]
- Niemann, H.; Elvert, M. Diagnostic lipid biomarker and stable carbon isotope signatures of microbial communities mediating the anaerobic oxidation of methane with sulphate. Org. Geochem. 2008, 39, 1668–1677. [Google Scholar] [CrossRef]
- Pape, T.; Blumenberg, M.; Seifert, R.; Egorov, V.N.; Gulin, S.B.; Michaelis, W. Lipid geochemistry of methane-seep-related Black Sea carbonates. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2005, 227, 31–47. [Google Scholar] [CrossRef]
- Pancost, R.D.; Bouloubassi, I.; Aloisi, G.; Damsté, J.S.S.; Party, T.M.S.S. Three series of non-isoprenoidal dialkyl glycerol diethers in cold-seep carbonate crusts. Org. Geochem. 2001, 32, 695–707. [Google Scholar] [CrossRef]
- Saito, R.; Oba, M.; Kaiho, K.; Maruo, C.; Fujibayashi, M.; Chen, J.; Chen, Z.-Q.; Tong, J. Ether lipids from the Lower and Middle Triassic at Qingyan, Guizhou Province, Southern China. Org. Geochem. 2013, 58, 27–42. [Google Scholar] [CrossRef]
- Grossi, V.; Mollex, D.; Vincon-Laugier, A.; Hakil, F.; Pacton, M.; Cravo-Laureau, C. Mono- and dialkyl glycerol ether lipids in anaerobic bacteria: Biosynthetic insights from the mesophilic sulfate reducer Desulfatibacillum alkenivorans PF2803T. Appl. Environ. Microbiol. 2015, 81, 3157–3168. [Google Scholar] [CrossRef] [Green Version]
- DasSarma, S. Extreme Halophiles Are Models for Astrobiology. Microbe 2006, 1, 120–126. [Google Scholar] [CrossRef] [Green Version]
- Maignien, L.; Parkes, R.J.; Cragg, B.; Niemann, H.; Knittel, K.; Coulon, S.; Akhmetzhanov, A.; Boon, N. Anaerobic oxidation of methane in hypersaline cold seep sediments. FEMS Microbiol. Ecol. 2013, 83, 214–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziegenbalg, S.B.; Birgel, D.; Hoffmann-Sell, L.; Pierre, C.; Rouchy, J.M.; Peckmann, J. Anaerobic oxidation of methane in hypersaline Messinian environments revealed by 13C-depleted molecular fossils. Chem. Geol. 2012, 292–293, 140–148. [Google Scholar] [CrossRef]
- Sanz-Montero, M.E.; Rodriguez-Aranda, J.P.; Calvo, J.P. Mediation of Endoevaporitic Microbial Communities in Early Replacement of Gypsum by Dolomite: A Case Study from Miocene Lake Deposits of the Madrid Basin, Spain. J. Sediment. Res. 2006, 76, 1257–1266. [Google Scholar] [CrossRef]
- Vandeginste, V.; Snell, O.; Hall, M.R.; Steer, E.; Vandeginste, A. Acceleration of dolomitization by zinc in saline waters. Nat. Commun. 2019, 10, 1851. [Google Scholar] [CrossRef] [Green Version]
- Deng, S.; Dong, H.; Lv, G.; Jiang, H.; Yu, B.; Bishop, M.E. Microbial dolomite precipitation using sulfate reducing and halophilic bacteria: Results from Qinghai Lake, Tibetan Plateau, NW China. Chem. Geol. 2010, 278, 151–159. [Google Scholar] [CrossRef]
- Vasconcelos, C.; McKenzie, J.A. Microbial mediation of modern dolomite precipitation and diagenesis under anoxic conditions (Lagoa Vermelha, Rio de Janeiro, Brazil). J. Sediment. Res. 1997, 67, 378–390. [Google Scholar]
- Vasconcelos, C.; McKenzie, J.A.; Bernasconi, S.; Grujic, D.; Tien, A.J. Microbial mediation as a possible mechanism for natural dolomite formation at low temperatures. Nature 1995, 377, 220–222. [Google Scholar] [CrossRef]
- Roberts, J.A.; Bennett, P.C.; González, L.A.; MacPherson, G.L.; Miliken, K.L. Microbial precipitation of dolomite in methanogenic groundwater. Geology 2004, 32, 277–280. [Google Scholar] [CrossRef]
- Ritger, S.; Carson, B.; Suess, E. Methane-derived authigenic carbonates formed by subduction-induced pore-water expulsion along the Oregon:Washington margin. Geol. Soc. Am. Bull. 1987, 98, 147–156. [Google Scholar] [CrossRef]
- Feenstra, E.J.; Birgel, D.; Heindel, K.; Wehrmann, L.M.; Jaramillo-Vogel, D.; Grobety, B.; Frank, N.; Hancock, L.G.; Van Rooij, D.; Peckmann, J.; et al. Constraining the formation of authigenic carbonates in a seepage-affected cold-water coral mound by lipid biomarkers. Geobiology 2020, 18, 185–206. [Google Scholar] [CrossRef]
- Peckmann, J.; Thiel, V.; Michaelis, W.; Clari, P.; Gaillard, C.; Martire, L.; Reitner, J. Cold seep deposits of Beauvoisin (Oxfordian; southeastern France) and Marmorito (Miocene; northern Italy): Microbially induced authigenic carbonates. Int. J. Earth Sci. 1999, 88, 60–75. [Google Scholar] [CrossRef] [Green Version]
- Shiraishi, F.; Akimoto, T.; Tomioka, N.; Takahashi, Y.; Matsumoto, R.; Snyder, G.T. Microbial traces found in microdolomite associated with seep-related shallow gas hydrate. Front. Earth Sci. 2023, 11, 1188142. [Google Scholar] [CrossRef]
- Daly, R.A. The limeless ocean of Pre-cambrian time. Am. J. Sci. 1907, 23, 93–115. [Google Scholar] [CrossRef]
- Steidtmann, E. The evolution of limestone and dolomite. J. Geol. 1911, 19, 323–345. [Google Scholar] [CrossRef]
- Van Tuyl, F.M. The Origin of Dolomite. Iowa Geol. Surv. Annu. Rep. 1916, 25, 251–422. [Google Scholar] [CrossRef] [Green Version]
- Oehler, D.Z.; Allen, C.C. Evidence for pervasive mud volcanism in Acidalia Planitia, Mars. Icarus 2010, 208, 636–657. [Google Scholar] [CrossRef]
- Perl, S.; Baxter, B. Great Salt Lake as an Astrobiology Analogue for Ancient Martian Hypersaline Aqueous Systems. In Great Salt Lake Biology; Baxter, B., Butler, J., Eds.; Springer Nature Switzerland AG: Basel, Switzerland, 2020. [Google Scholar]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Putative Fragmentation of Biomarker Compounds, Formula, and Exact Mass (u) | Standard m/z (+) | Average Measured m/z (+) | Tentative Formula |
|---|---|---|---|
17α(H),21β(H)-Norhopane (norhopane; C29H50; 398.39) | 191.18 | 191.14 (σ = 0.02) | [C14H23] + |
| 259.25 | 259.23 (σ = 0.008) | [C19H31] + | |
| 367.35 | 367.32 (σ = 0.03) | [C27H43] + | |
| 383.39 | 383.52 (σ = 0.02) | [C28H47] + | |
Hop-17(21)-ene or Hop-22(29)-ene (diploptene) (C30H50; 410.39) | 189.17 | 189.13 (σ = 0.008) | [C14H21] + |
| 393.36 | 393.33 (σ = 0.06) | [C29H45] + | |
| 395.37 | 395.27 (σ = 0.04) | [C29 H47] + | |
Dialkyl glycerol diether (DAGE, C35H68O3; 536.52) | 283.26 | 283.21 (σ = 0.01) | [C18H35O2] + |
Dialkyl glycerol diether (DAGE, C33H68O3; 512.52) | 285.27 | 285.19 (σ = 0.04) | [C18H37O2] + |
| 301.26 | 301.15 (σ = 0.04) | [C18H37O3] + | |
| 495.51 | 495.28 (σ = 0.04) | [M-H2O] + | |
Dialkyl glycerol diether (DAGE, C29H58O3; 454.44) | 255.23 | 255.24 (σ = 0.04) | [C16H31O2] + |
| 257.24 | 257.23 (σ = 0.02) | [C16H33O2] + | |
| 455.43 | 455.5 (σ = 0.2) | [M+H] + | |
Diacylglycerolester (DG 16:0/18:0; C37H72O5; 596.54) | 341.32 | 341.24 (σ = 0.03) | [C21H41O3] + |
| 579.54 | 579.22 (σ = 0.1) | [M-H2O] + | |
Diacylglycerolester (DG; 15:0/17:0 C35H68O5; 568.51) | 299.26 | 299.25 (σ = 0.01) | [C18H35O3] + |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alibrahim, A.S.; Sodhi, R.N.S.; Duane, M.J.; Dittrich, M. Imaging of Ancient Microbial Biomarkers within Miocene Dolomite (Kuwait) Using Time-of-Flight Secondary Ion Mass Spectrometry. Minerals 2023, 13, 968. https://doi.org/10.3390/min13070968
Alibrahim AS, Sodhi RNS, Duane MJ, Dittrich M. Imaging of Ancient Microbial Biomarkers within Miocene Dolomite (Kuwait) Using Time-of-Flight Secondary Ion Mass Spectrometry. Minerals. 2023; 13(7):968. https://doi.org/10.3390/min13070968
Chicago/Turabian StyleAlibrahim, Ammar S., Rana N. S. Sodhi, Michael J. Duane, and Maria Dittrich. 2023. "Imaging of Ancient Microbial Biomarkers within Miocene Dolomite (Kuwait) Using Time-of-Flight Secondary Ion Mass Spectrometry" Minerals 13, no. 7: 968. https://doi.org/10.3390/min13070968
APA StyleAlibrahim, A. S., Sodhi, R. N. S., Duane, M. J., & Dittrich, M. (2023). Imaging of Ancient Microbial Biomarkers within Miocene Dolomite (Kuwait) Using Time-of-Flight Secondary Ion Mass Spectrometry. Minerals, 13(7), 968. https://doi.org/10.3390/min13070968