Experimental Deployment of Microbial Mineral Carbonation at an Asbestos Mine: Potential Applications to Carbon Storage and Tailings Stabilization

 , , and
, , and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Regional Geology of Woodsreef Mine
2.2. Experiment Design
2.3. Mineralogical Characterization of the Tailings
3. Results
3.1. Field Observations
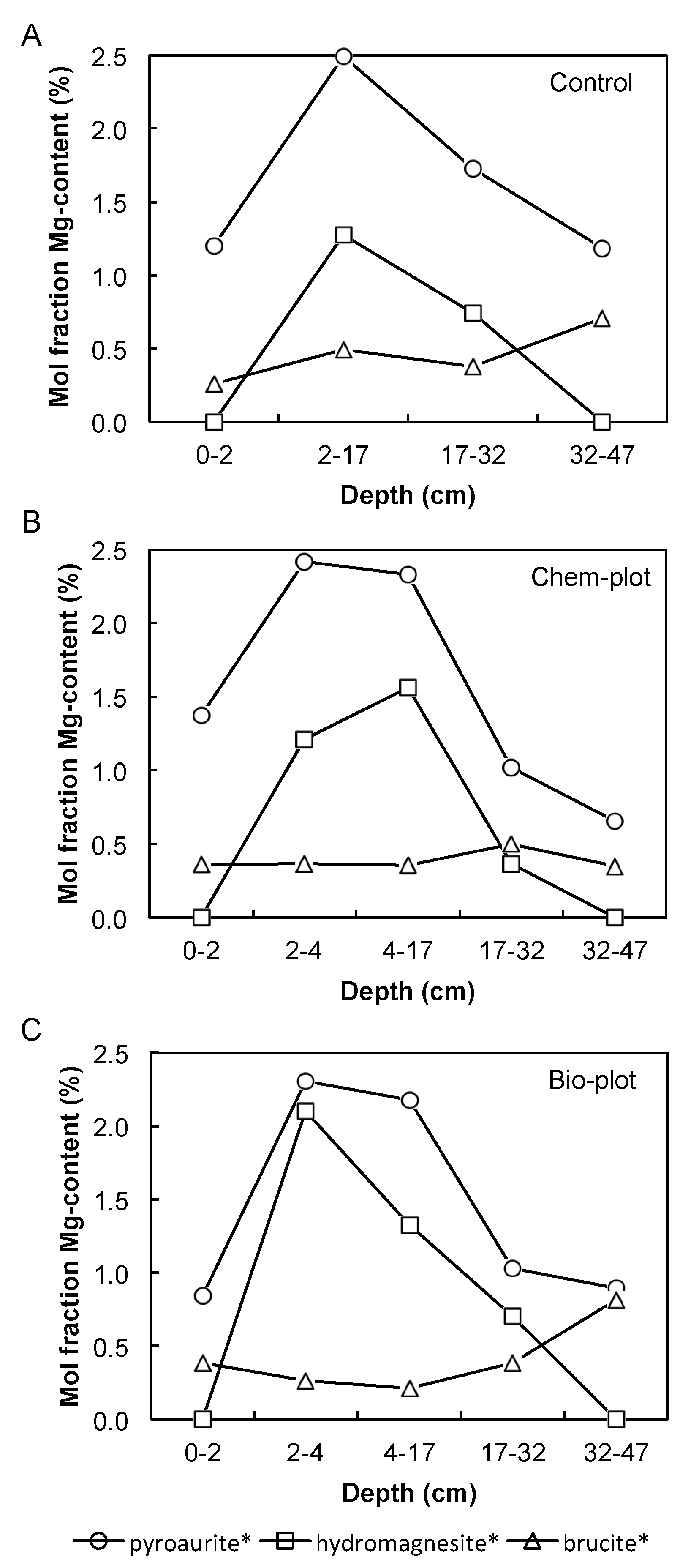
3.2. Rietveld Refinement Results
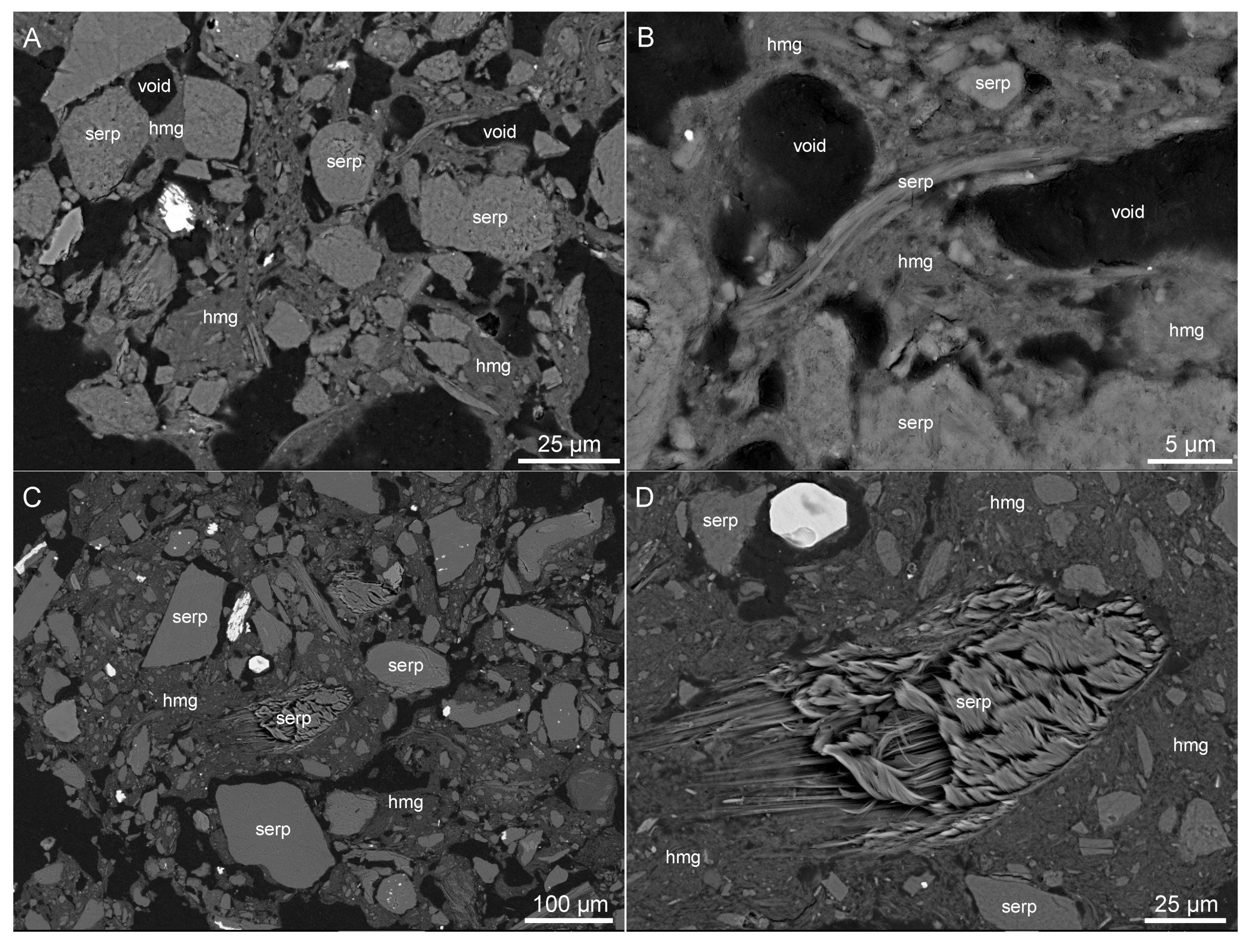
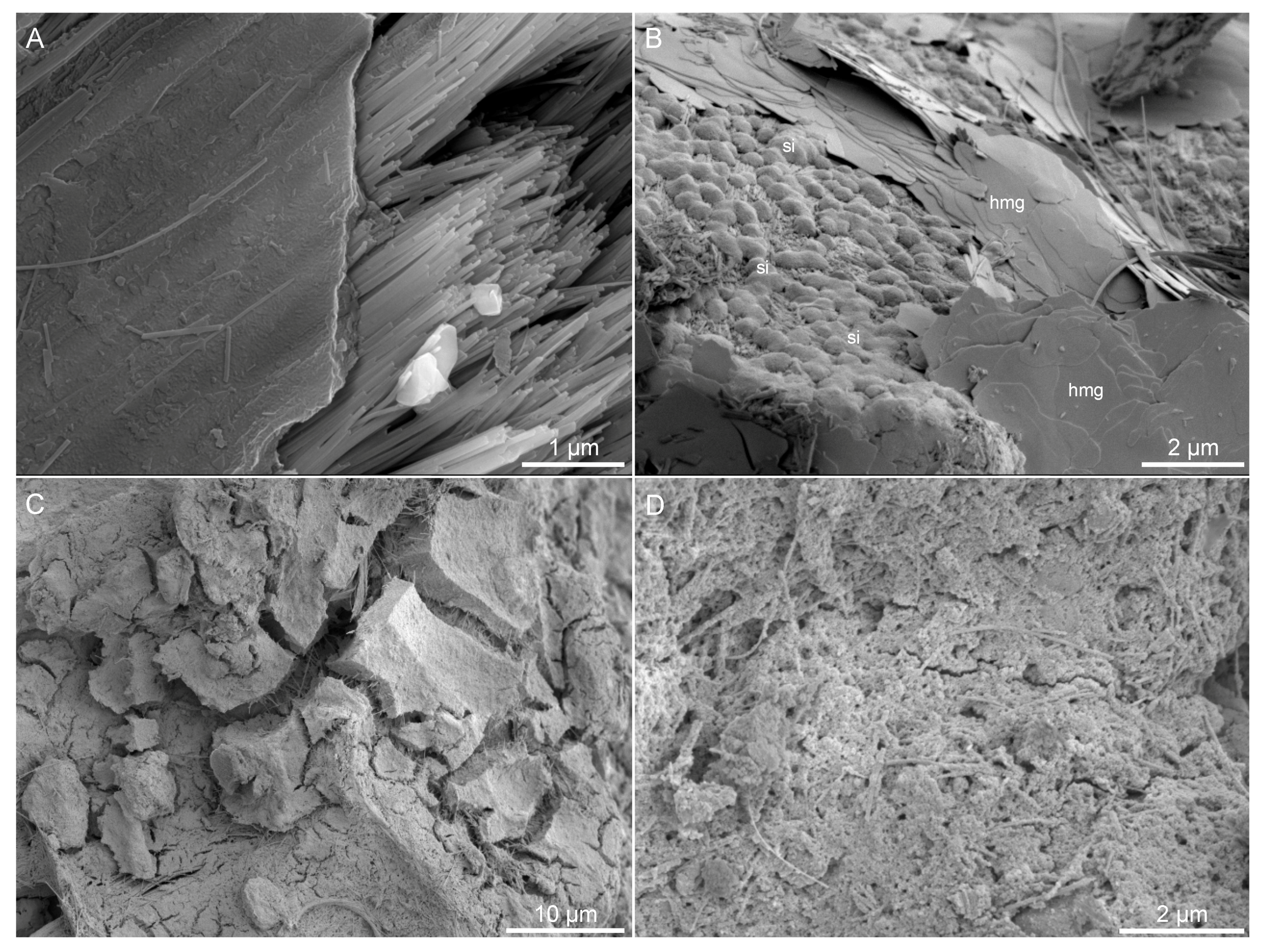
3.3. Electron Microscopy
4. Discussion
4.1. Precipitate Morphologies As an Indication of Chemistry
4.2. Mineral Carbonation Success
4.3. Water as a Limiting Factor
4.4. Carbon as a Limiting Factor
4.5. Carbonation: Maximizing Tailings Stabilization versus Carbon Sequestration
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Lackner, K.S. A guide to CO2 sequestration. Science 2003, 300, 1677–1678. [Google Scholar] [CrossRef] [PubMed]
- Lackner, K.S.; Wendt, C.H.; Butt, D.P.; Joyce, E.L.; Sharp, D.H. Carbon-dioxide disposal in carbonate minerals. Energy 1995, 20, 1153–1170. [Google Scholar] [CrossRef]
- Seifritz, W. CO2 disposal by means of silicates. Nature 1990, 345, 486. [Google Scholar] [CrossRef]
- Power, I.M.; Dipple, G.M.; Southam, G. Bioleaching of ultramafic tailings by Acidithiobacillus spp. For CO2 sequestration. Environ. Sci. Technol. 2010, 44, 456–462. [Google Scholar] [CrossRef] [PubMed]
- Power, I.M.; Wilson, S.A.; Small, D.P.; Dipple, G.M.; Wan, W.; Southam, G. Microbially mediated mineral carbonation: Roles of phototrophy and heterotrophy. Environ. Sci. Technol. 2011, 45, 9061–9068. [Google Scholar] [CrossRef] [PubMed]
- Thom, J.G.M.; Dipple, G.M.; Power, I.M.; Harrison, A.L. Chrysotile dissolution rates: Implications for carbon sequestration. Appl. Geochem. 2013, 35, 244–254. [Google Scholar] [CrossRef]
- Wilson, S.A.; Dipple, G.M.; Power, I.M.; Barker, S.L.L.; Fallon, S.J.; Southam, G. Subarctic weathering of mineral wastes provides a sink for atmospheric CO2. Environ. Sci. Technol. 2011, 45, 7727–7736. [Google Scholar] [CrossRef] [PubMed]
- Wilson, S.A.; Dipple, G.M.; Power, I.M.; Thom, J.M.; Anderson, R.G.; Raudsepp, M.; Gabite, J.E.; Southam, G. Carbon dioxide fixation within mine wastes of ultramafic-hosted ore deposits: Examples from the Clinton Creek and Cassiar chrysotile deposits, Canada. Econ. Geol. 2009, 104, 95–112. [Google Scholar] [CrossRef]
- Wilson, S.A.; Harrison, A.L.; Dipple, G.M.; Power, I.M.; Barker, S.L.L.; Mayer, U.K.; Fallon, S.J.; Raudsepp, M.; Southam, G. Offsetting of CO2 emissions by air capture in mine tailings at the Mount Keith Nickel Mine, Western Australia: Rates, controls and prospects for carbon neutral mining. Int. J. Greenhouse Gas Control 2014, 25, 121–140. [Google Scholar] [CrossRef]
- Kelemen, P.B.; Matter, J. In situ carbonation of peridotite for CO2 storage. Proc. Natl. Acad. Sci. USA 2008, 104, 17295–17300. [Google Scholar] [CrossRef]
- Kelemen, P.B.; Matter, J.; Streit, E.E.; Rudge, J.F.; Curry, W.B.; Blusztajn, J. Rates and mechanisms of mineral carbonation in peridotite- natural processes and recipes for enhanced, in situ CO2 capture and storage. Annu. Rev. Earth Planet. Sci. 2011, 39, 545–576. [Google Scholar] [CrossRef]
- Paukert, A.N.; Matter, J.M.; Kelemen, P.B.; Shock, E.L.; Havig, J.R. Reaction path modeling of enhanced in situ CO2 mineralization for carbon sequestration in the peridotite of the Samail Ophiolite, Sultanate of Oman. Chem. Geol. 2012, 330–331, 86–100. [Google Scholar] [CrossRef]
- Pronost, J.; Beaudoin, G.; Tremblay, J.; Larachi, F.; Duchesne, J.; Hébert, R.; Constantin, M. Carbon sequestration kinetic and storage capacity of ultramafic mining waste. Environ. Sci. Technol. 2011, 45, 9413–9420. [Google Scholar] [CrossRef] [PubMed]
- Harrison, A.L.; Power, I.M.; Dipple, G.M. Accelerated carbonation of brucite in mine tailings for carbon sequestration. Environ. Sci. Technol. 2013, 47, 126–134. [Google Scholar] [CrossRef] [PubMed]
- Pronost, J.; Beaudoin, G.; Lemieux, J.-M.; Hébert, R.; Constantin, M.; Marcouiller, S.; Klein, M.; Duchesne, J.; Molson, J.W.; Larchi, F.; et al. CO2-depleted warm air venting from chrysotile milling waste (Thetford Mines, Canada): Evidence for in-situ carbon capture from the atmosphere. Geology 2012, 40, 275–278. [Google Scholar] [CrossRef]
- Bea, S.A.; Wilson, S.A.; Mayer, K.U.; Dipple, G.M.; Power, I.M.; Gamazo, P. Reactive transport modeling of natural carbon sequestration in ultramafic mine tailings. Vadose Zone J. 2012, 11. [Google Scholar] [CrossRef]
- Oskierski, H.C.; Dlugogorski, B.Z.; Jacobsen, G. Sequestration of atmospheric CO2 in chrysotile mine tailings of the Woodsreef Asbestos Mine, australia: Quantitative mineralogy, isotopic fingerprinting and carbonation rates. Chem. Geol. 2013, 358, 156–169. [Google Scholar] [CrossRef]
- Lechat, K.; Lemieux, J.-M.; Molson, J.; Beaudoin, G.; Hébert, R. Field evidence of CO2 sequestration by mineral carbonation in ultramafic milling wastes, Thetford Mines, Canada. Int. J. Greenh. Gas Control 2016, 47, 110–121. [Google Scholar] [CrossRef]
- Power, I.; McCutcheon, J.; Harrison, A.; Wilson, S.; Dipple, G.; Kelly, S.; Southam, C.; Southam, G. Strategizing carbon-neutral mines: A case for pilot projects. Minerals 2014, 4, 399–436. [Google Scholar] [CrossRef]
- Assima, G.P.; Larachi, F.; Beaudoin, G.; Molson, J. Dynamics of carbon dioxide uptake in chrysotile mining residues—Effect of mineralogy and liquid saturation. Int. J. Greenh. Gas Control 2013, 12, 124–135. [Google Scholar] [CrossRef]
- Wilson, S.A.; Raudsepp, M.; Dipple, G.M. Verifying and quantifying carbon fixation in minerals from serpentine-rich mine tailings using the Rietveld method with X-ray powder diffraction data. Am. Mineral. 2006, 91, 1331–1341. [Google Scholar] [CrossRef]
- Koumantakis, E.; Kalliopi, A.; Dimitrios, K.; Gidarakos, E. Asbestos pollution in an inactive mine: Determination of asbestos fibers in the deposit tailings and water. J. Hazard. Mater. 2009, 167, 1080–1088. [Google Scholar] [CrossRef] [PubMed]
- Assima, G.P.; Larachi, F.; Molson, J.; Beaudoin, G. Comparative study of five Quebec ultramafic mining residues for use in direct ambient carbon dioxide mineral sequestration. Chem. Eng. J. 2014, 245, 56–64. [Google Scholar] [CrossRef]
- Turci, F.; Favero-Longo, S.E.; Gazzano, C.; Tomatis, M.; Gentile-Garofalo, L.; Bergamini, M. Assessment of asbestos exposure during a simulated agricultural activity in the proximity of the former asbestos mine of Balangero, Italy. J. Hazard. Mater. 2016, 308, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Levitan, D.M.; Hammarstrom, J.M.; Gunter, M.E.; Ii, S.; Robert, R.; Chou, I.; Piatak, N.M. Mineralogy of mine waste at the vermont asbestos group mine, Belvidere Mountain, Vermont. Am. Mineral. 2009, 94, 1063–1066. [Google Scholar] [CrossRef]
- Vignaroli, G.; Ballirano, P.; Belardi, G.; Rossetti, F. Asbestos fibre identification vs. Evaluation of asbestos hazard in ophiolitic rock mélanges, a case study from the Ligurian Alps (Italy). Environ. Earth Sci. 2014, 72, 3679–3698. [Google Scholar] [CrossRef]
- Vignaroli, G.; Rossetti, F.; Belardi, G.; Billi, A. Linking rock fabric to fibrous mineralisation: A basic tool for the asbestos hazard. Nat. Hazards Earth Syst. Sci. 2011, 11, 1267–1280. [Google Scholar] [CrossRef]
- McCutcheon, J.; Wilson, S.A.; Southam, G. Microbially accelerated carbonate mineral precipitation as a strategy for in situ carbon sequestration and rehabilitation of asbestos mine sites. Environ. Sci. Technol. 2016, 50, 1419–1427. [Google Scholar] [CrossRef] [PubMed]
- Pasquier, L.-C.; Mercier, G.; Blais, J.-F.; Cecchi, E.; Kentish, S. Parameters optimization for direct flue gas CO2 capture and sequestration by aqueous mineral carbonation using activated serpentinite based mining residue. Appl. Geochem. 2014, 50, 66–73. [Google Scholar] [CrossRef]
- Kemache, N.; Pasquier, L.-C.; Mouedhen, I.; Cecchi, E.; Blais, J.-F.; Mercier, G. Aqueous mineral carbonation of serpentinite on a pilot scale: The effect of liquid recirculation on CO2 sequestration and carbonate precipitation. Appl. Geochem. 2016, 67, 21–29. [Google Scholar] [CrossRef]
- Drew, G.H. On the precipitation of calcium carbonate in the sea by marine bacteria, and on the action of denitrifying bacteria in tropical and temperate seas. J. Mar. Biol. Assoc. U. K. 1913, 9, 479–524. [Google Scholar] [CrossRef]
- Power, I.M.; Wilson, S.A.; Thom, J.M.; Dipple, G.M.; Southam, G. Biologically induced mineralization of dypingite by cyanobacteria from an alkaline wetland near Atlin, British Columbia, Canada. Geochem. Trans. 2007, 8, 13. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.B.; Ferris, F.G. Cyanobacterial precipitation of gypsum, calcite, and magnesite from natural alkaline water. Geology 1990, 18, 995–998. [Google Scholar] [CrossRef]
- Riding, R. Microbial carbonates: The geological record of calcified bacterial-algal mats and biofilms. Sedimentology 2000, 47, 179–214. [Google Scholar] [CrossRef]
- Riding, R. Cyanobacterial calcification, carbon dioxide concentrating mechanisms, and Proterozoic-Cambrian changes in atmospheric composition. Geobiology 2006, 4, 299–316. [Google Scholar] [CrossRef]
- Aloisi, G. The calcium carbonate saturation state in cyanobacterial mats throughout Earth’s history. Geochim. Cosmochim. Acta 2008, 72, 6037–6060. [Google Scholar] [CrossRef]
- Ferris, F.G.; Phoenix, V.; Fujita, Y.; Smith, R.W. Kinetics of calcite precipitation induced by ureolytic bacteria at 10 to 20 °C in artificial groundwater. Geochim. Cosmochim. Acta 2004, 68, 1701–1710. [Google Scholar] [CrossRef]
- Mitchell, A.C.; Ferris, F.G. The influence of Bacillus pasteurii on the nucleation and growth of calcium carbonate. Geomicrobiol. J. 2006, 23, 213–226. [Google Scholar] [CrossRef]
- Dupraz, C.; Reid, R.P.; Braissant, O.; Decho, A.W.; Norman, R.S.; Visscher, P.T. Processes of carbonate precipitation in modern microbial mats. Earth-Sci. Rev. 2009, 96, 141–162. [Google Scholar] [CrossRef]
- Visscher, P.T.; Stolz, J.F. Microbial mats as bioreactors: Populations, processes, and products. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2005, 219, 87–100. [Google Scholar] [CrossRef]
- Braissant, O.; Decho, A.W.; Przekop, K.M.; Gallagher, K.L.; Glunk, C.; Dupraz, C.; Visscher, P.T. Characteristics and turnover of exopolymeric substances in a hypersaline microbial mat. FEMS Microbiol. Ecol. 2009, 67, 293–307. [Google Scholar] [CrossRef] [PubMed]
- Decho, A.W.; Visscher, P.T.; Reid, R.P. Production and cycling of natural microbial exopolymers (eps) within a marine stromatolite. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2005, 219, 71–86. [Google Scholar] [CrossRef]
- McCutcheon, J.; Power, I.M.; Harrison, A.L.; Dipple, G.M.; Southam, G. A greenhouse-scale photosynthetic microbial bioreactor for carbon sequestration in magnesium carbonate minerals. Environ. Sci. Technol. 2014, 48, 9142–9151. [Google Scholar] [CrossRef] [PubMed]
- Glen, R.A.; Butt, B.C. Chrysotile asbestos at Woodsreef, New South Wales. Econ. Geol. 1981, 76, 1153–1169. [Google Scholar] [CrossRef]
- Cawood, P.A.; Pisarevsky, S.A.; Leitch, E.C. Unraveling the New England orocline, east Gondwana accretionary margin. Tectonics 2011, 30. [Google Scholar] [CrossRef]
- NSW Government Trade & Investment. Woodsreef Mine Major Rehabilitation Project. Available online: http://www.resourcesandenergy.nsw.gov.au/miners-and-explorers/programs-and-initiatives/derelict/woodsreef-mine-major-rehabilitation-project (accessed on 30 July 2014).
- Brown, R.E.; Brownlow, J.W.; Krynen, J.P. Manilla–Narrabri 1:250 000 Metallogenic Map sh/56–9, sh/55–12: Metallogenic Study and Mineral Deposit Data Sheets; Geological Survey of New South Wales, Department of Mineral Resources: Sydney, Australia, 1992.
- Turvey, C.C.; Wilson, S.A.; Hamilton, J.L.; Southam, G. Field-based accounting of CO2 sequestration in ultramafic mine wastes using portable X-ray diffraction. Am. Mineral. 2017, 102, 1302–1310. [Google Scholar] [CrossRef]
- Vonshak, A. Laboratory techniques for the cultivation of microalgae. In CRC Handbook of Microalgae Mass Culture; Richmond, A., Ed.; CRC Press Inc.: Bocca Raton, FL, USA, 1986; p. 117. [Google Scholar]
- Bureau of Meteorology. Barraba, New South Wales 2016 Daily Weather Observations. Available online: http://www.bom.gov.au/climate/dwo/IDCJDW2010.latest.shtml (accessed on 10 February 2016).
- Lahav, O.; Morgan, B.E.; Loewenthal, R.E. Measurement of pH, alkalinity and acidity in ultra-soft waters. Water SA 2001, 27, 423–431. [Google Scholar] [CrossRef]
- Bruker AXS. Topas v. 3.0: General Profile and Structure Analysis Software for Powder Diffraction Data; Bruker AXS: Karlsruhe, Germany, 2004. [Google Scholar]
- Rietveld, H.M. A profile refinement method for nuclear and magnetic structures. J. Appl. Crystallogr. 1969, 2, 65–71. [Google Scholar] [CrossRef]
- Bish, D.L.; Howard, S.A. Quantitative phase analysis using the Rietveld method. J. Appl. Crystallogr. 1988, 21, 86–91. [Google Scholar] [CrossRef]
- Hill, R.; Howard, C. Quantitative phase analysis from neutron powder diffraction data using the Rietveld method. J. Appl. Crystallogr. 1987, 20, 467–474. [Google Scholar] [CrossRef]
- Cheary, R.W.; Coelho, A. A fundamental parameters approach to X-ray line-profile fitting. J. Appl. Crystallogr. 1992, 25, 109–121. [Google Scholar] [CrossRef]
- Pawley, G. Unit-cell refinement from powder diffraction scans. J. Appl. Crystallogr. 1981, 14, 357–361. [Google Scholar] [CrossRef]
- Falini, G.; Foresti, E.; Gazzano, M.; Gualtieri, A.F.; Leoni, M.; Lesci, I.G.; Roveri, N. Tubular-shaped stoichiometric chrysotile nanocrystals. Chemistry 2004, 10, 3043–3049. [Google Scholar] [CrossRef] [PubMed]
- Wilson, S.A.; Raudsepp, M.; Dipple, G.M. Quantifying carbon fixation in trace minerals from processed kimberlite: A comparative study of quantitative methods using X-ray powder diffraction data with applications to the Diavik Diamond Mine, Northwest Territories, Canada. Appl. Geochem. 2009, 24, 2312–2331. [Google Scholar] [CrossRef]
- Gronow, J.R. The dissolution of asbestos fibres in water. Clay Miner. 1987, 22, 21–35. [Google Scholar] [CrossRef]
- Morgan, A. Acid leaching studies of chrysotile asbestos from mines in the Coalinga region of California and from Quebec and British Columbia. Ann. Occup. Hyg. 1997, 41, 249–268. [Google Scholar] [CrossRef]
- Wang, L.; Lu, A.; Wang, C.; Zheng, X.; Zhao, D.; Liu, R. Nano-fibriform production of silica from natural chrysotile. J. Colloid Interface Sci. 2006, 295, 436–439. [Google Scholar] [CrossRef] [PubMed]
- Rozalen, M.; Huertas, F.J. Comparative effect of chrysotile leaching in nitric, sulfuric and oxalic acids at room temperature. Chem. Geol. 2013, 352, 134–142. [Google Scholar] [CrossRef]
- Park, A.-H.A.; Fan, L.-S. CO2 mineral sequestration: Physically activated dissolution of serpentine and pH swing process. Chem. Eng. Sci. 2004, 59, 5241–5247. [Google Scholar] [CrossRef]
- McCutcheon, J.; Dipple, G.M.; Wilson, S.A.; Southam, G. Production of magnesium-rich solutions by acid leaching of chrysotile: A precursor to field-scale deployment of microbially enabled carbonate mineral precipitation. Chem. Geol. 2015, 413, 119–131. [Google Scholar] [CrossRef]
- Taylor, H.M.; Hansen, H.C.B.; Stanger, G.; Bender Koch, C. On the genesis and composition of natural pyroaurite. Clay Miner. 1991, 26, 297–309. [Google Scholar] [CrossRef]
- Schaef, H.T.; Windisch, C.F., Jr.; McGrail, B.P.; Martin, P.F.; Rosso, K.M. Brucite [Mg(OH2)] carbonation in wet supercritical CO2: An in situ high pressure X-ray diffraction study. Geochim. Cosmochim. Acta 2011, 75, 7458–7471. [Google Scholar] [CrossRef]
- Assima, G.P.; Larachi, F.; Beaudoin, G.; Molson, J. CO2 sequestration in chrysotile mining residues—Implication of watering and passivation under environmental conditions. Ind. Eng. Chem. Res. 2012, 51, 8726–8734. [Google Scholar] [CrossRef]
- Harrison, A.L.; Dipple, G.M.; Power, I.M.; Mayer, K.U. Influence of surface passivation and water content on mineral reactions in unsaturated porous media: Implications for brucite carbonation and CO2 sequestration. Geochim. Cosmochim. Acta 2015, 148, 477–495. [Google Scholar] [CrossRef]
- Altermann, W.; Kazmierczak, J.; Oren, A.; Wright, D.T. Cyanobacterial calcification and its rock-building potential during 3.5 billion years of Earth history. Geobiology 2006, 4, 147–166. [Google Scholar] [CrossRef]
- Dupraz, C.; Visscher, P.T. Microbial lithification in marine stromatolites and hypersaline mats. Trends Microbiol. 2005, 13, 429–438. [Google Scholar] [CrossRef] [PubMed]
- Beveridge, T.J. The bacterial surface: General considerations towards design and function. Can. J. Microbiol. 1988, 34, 363–372. [Google Scholar] [CrossRef] [PubMed]
- Kluge, S.; Weston, J. Can a hydroxide ligand trigger a change in the coordination number of magnesium ions in biological systems? Biochemistry 2005, 44, 4877–4885. [Google Scholar] [CrossRef] [PubMed]
- Trichet, J.; Défarge, C. Non-biologically supported organomineralization. Bull. Inst. Oceanogr. 1995, 14, 203–226. [Google Scholar]
- Obst, M.; Dynes, J.J.; Lawrence, J.R.; Swerhone, G.D.W.; Benzerara, K.; Karunakaran, C.; Kaznatcheev, K.; Tyliszczak, T.; Hitchock, A.P. Precipitation of amorphous CaCO3 (aragonite-like) by cyanobacteria: A STXM study of the influence of eps on the nucleation process. Geochim. Cosmochim. Acta 2009, 72, 4180–4198. [Google Scholar] [CrossRef]
- Hamilton, J.L.; Wilson, S.A.; Morgan, B.; Turvey, C.C.; Paterson, D.J.; MacRae, C.; McCutcheon, J.; Southam, G. Nesquehonite sequesters transition metals and CO2 during accelerated carbon mineralisation. Int. J. Greenh. Gas Control 2016, 55, 73–81. [Google Scholar] [CrossRef]
- Šourková, M.; Frouz, J.; Fettweis, U.; Bens, O.; Hüttl, R.F.; Šantrůčková, H. Soil development and properties of microbial biomass succession in reclaimed post mining sites near Sokolov (Czech Republic) and near Cottbus (Germany). Geoderma 2005, 129, 73–80. [Google Scholar] [CrossRef]
- Frouz, J.; Keplin, B.; Pižl, V.; Tajovský, K.; Starý, J.; Lukešová, A.; Nováková, A.; Balı́k, V.R.; Háněl, L.; Materna, J.; et al. Soil biota and upper soil layer development in two contrasting post-mining chronosequences. Ecol. Eng. 2001, 17, 275–284. [Google Scholar] [CrossRef]
- Meyer, D.R. Nutritional problems associated with the establishment of vegetation on tailings from an asbestos mine. Environ. Pollut. Ser. A Ecol. Biol. 1980, 23, 287–298. [Google Scholar] [CrossRef]
- Moore, T.R.; Zimmermann, R.C. Establishment of vegetation on serpentine asbestos mine wastes, southeastern Quebec, Canada. J. Appl. Ecol. 1977, 14, 589–599. [Google Scholar] [CrossRef]
- Oskierski, H.C.; Dlugogorski, B.Z.; Oliver, T.K.; Jacobsen, G. Chemical and isotopic signatures of waters associated with the carbonation of ultramafic mine tailings, Woodsreef Asbestos Mine, Australia. Chem. Geol. 2016, 436, 11–23. [Google Scholar] [CrossRef]
- Mata, T.M.; Martins, A.A.; Caetano, N.S. Microalgae for biodiesel production and other applications: A review. Renew. Sustain. Energy Rev. 2010, 14, 217–232. [Google Scholar] [CrossRef]
- Ramanan, R.; Kannan, K.; Deshkar, A.; Yadav, R.; Chakrabarti, T. Enhanced algal CO2 sequestration through calcite deposition by Chlorella sp. and Spirulina platensis in a mini-raceway pond. Bioresour. Technol. 2010, 101, 2616–2622. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Depth Profile | Depth (cm) | Mineral Phase (wt %) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Serpentine | Pyroaurite | Magnetite | Hydromagnesite | Brucite | Calcite | Forsterite | Enstatite | Quartz | Total | ||
| Unreacted tailings | 0–2 | 91.2 | 1.3 | 3.5 | 0.0 | 0.1 | 0.2 | 2.3 | 0.8 | 0.5 | 100.0 |
| 2–17 | 88.8 | 2.6 | 2.2 | 1.1 | 0.3 | 0.4 | 2.7 | 1.5 | 0.2 | 100.0 | |
| 17–32 | 90.2 | 1.9 | 2.1 | 0.7 | 0.2 | 0.7 | 2.4 | 1.7 | 0.1 | 100.0 | |
| 32–47 | 90.8 | 1.3 | 2.1 | 0.0 | 0.4 | 0.5 | 2.9 | 2.0 | 0.0 | 100.0 | |
| Leached tailings (2 weeks) | 0–2 | 92.7 | 1.3 | 2.5 | 0.2 | 0.2 | 0.1 | 1.7 | 0.8 | 0.5 | 100.0 |
| 2–17 | 88.6 | 3.4 | 2.1 | 1.3 | 0.2 | 0.4 | 2.3 | 1.4 | 0.2 | 100.0 | |
| 17–32 | 89.1 | 2.3 | 2.5 | 1.2 | 0.2 | 0.6 | 2.8 | 1.2 | 0.1 | 100.0 | |
| 32–47 | 93.2 | 1.5 | 1.9 | 0.3 | 0.1 | 0.5 | 1.7 | 0.8 | 0.0 | 100.0 | |
| Leached tailings (11 weeks) | 0–2 | 90.2 | 1.5 | 2.8 | 0.0 | 0.2 | 0.2 | 2.7 | 1.9 | 0.5 | 100.0 |
| 2–4 | 88.6 | 2.6 | 2.2 | 1.1 | 0.2 | 0.3 | 3.3 | 1.5 | 0.3 | 100.0 | |
| 4–17 | 89.9 | 2.5 | 2.4 | 1.4 | 0.2 | 0.4 | 1.9 | 1.2 | 0.1 | 100.0 | |
| 17–32 | 91.9 | 1.1 | 2.2 | 0.3 | 0.3 | 0.5 | 2.1 | 1.5 | 0.0 | 100.0 | |
| 32–47 | 95.0 | 0.7 | 1.9 | 0.0 | 0.2 | 0.5 | 1.3 | 0.5 | 0.0 | 100.0 | |
| Leached and inoculated tailings (11 weeks) | 0–2 | 90.6 | 0.9 | 3.2 | 0.0 | 0.2 | 0.2 | 3.0 | 1.3 | 0.6 | 100.0 |
| 2–4 | 89.6 | 2.5 | 2.1 | 1.9 | 0.1 | 0.4 | 2.2 | 0.8 | 0.4 | 100.0 | |
| 4–17 | 90.4 | 2.4 | 2.0 | 1.2 | 0.1 | 0.7 | 2.2 | 1.0 | 0.0 | 100.0 | |
| 17–32 | 92.0 | 1.1 | 2.1 | 0.7 | 0.2 | 0.5 | 2.5 | 0.9 | 0.0 | 100.0 | |
| 32–47 | 92.4 | 1.0 | 2.3 | 0.0 | 0.5 | 0.6 | 2.2 | 1.1 | 0.0 | 100.0 | |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
McCutcheon, J.; Turvey, C.C.; Wilson, S.; Hamilton, J.L.; Southam, G. Experimental Deployment of Microbial Mineral Carbonation at an Asbestos Mine: Potential Applications to Carbon Storage and Tailings Stabilization. Minerals 2017, 7, 191. https://doi.org/10.3390/min7100191
McCutcheon J, Turvey CC, Wilson S, Hamilton JL, Southam G. Experimental Deployment of Microbial Mineral Carbonation at an Asbestos Mine: Potential Applications to Carbon Storage and Tailings Stabilization. Minerals. 2017; 7(10):191. https://doi.org/10.3390/min7100191
Chicago/Turabian StyleMcCutcheon, Jenine, Connor C. Turvey, Sasha Wilson, Jessica L. Hamilton, and Gordon Southam. 2017. "Experimental Deployment of Microbial Mineral Carbonation at an Asbestos Mine: Potential Applications to Carbon Storage and Tailings Stabilization" Minerals 7, no. 10: 191. https://doi.org/10.3390/min7100191
APA StyleMcCutcheon, J., Turvey, C. C., Wilson, S., Hamilton, J. L., & Southam, G. (2017). Experimental Deployment of Microbial Mineral Carbonation at an Asbestos Mine: Potential Applications to Carbon Storage and Tailings Stabilization. Minerals, 7(10), 191. https://doi.org/10.3390/min7100191