Responses of Acidithiobacillus thiooxidans A01 to Individual and Joint Nickel (Ni2+) and Ferric (Fe3+)

Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strain, Medium and Growth Condition
2.2. Nickel Resistance Experiment with Different Ferric Iron Concentration
2.3. Shock Treatment
2.4. Cell Collection and RNA Extraction
2.5. Primers and Real-Time Polymerase Chain Reaction (PCR)
3. Results
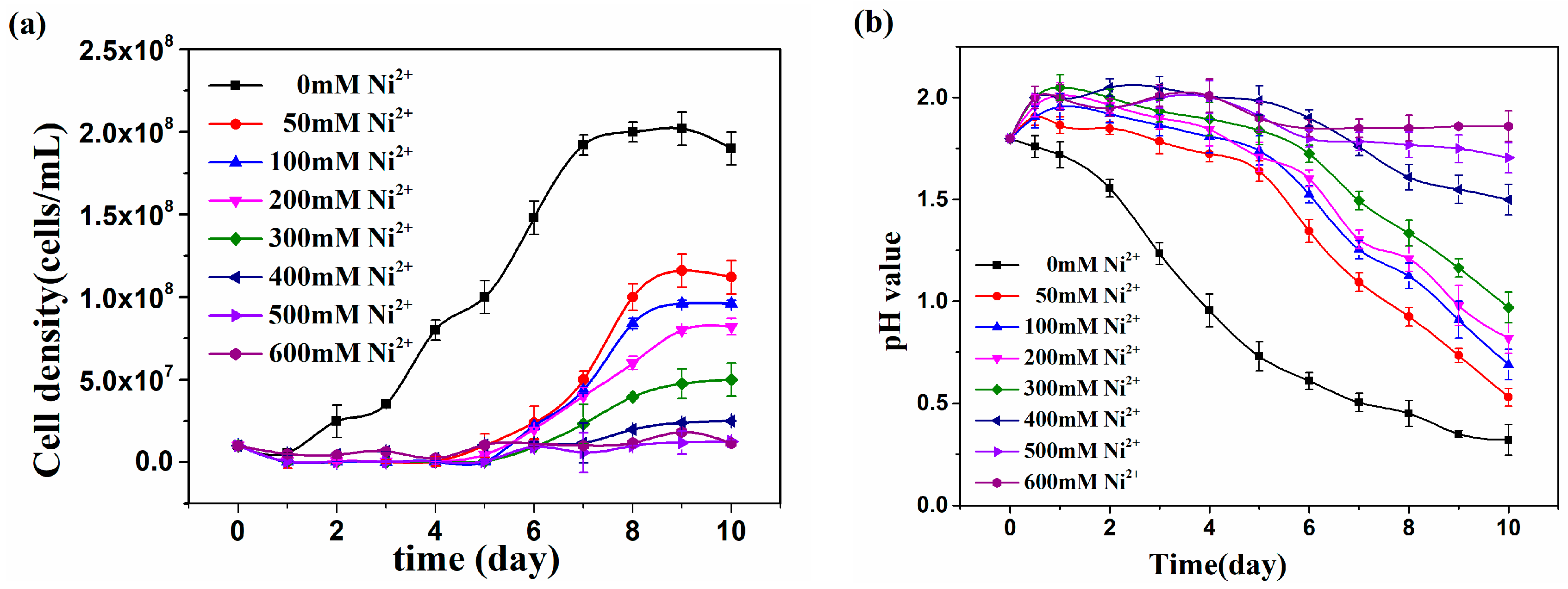
3.1. Growth of At. thiooxidans A01 under Different Ni2+ Concentrations
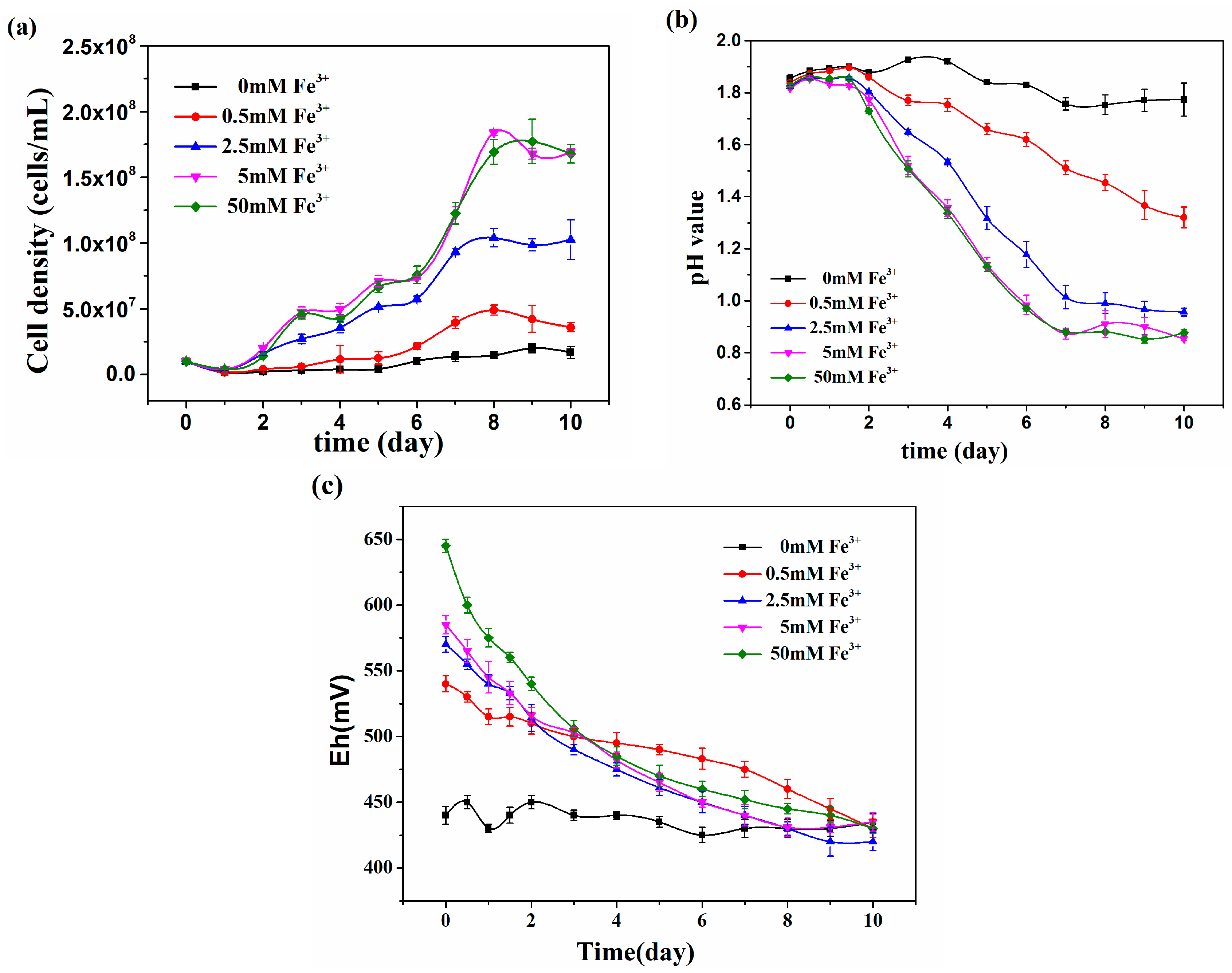
3.2. The effect of Ferric Iron on Growth of At. thiooxidans A01 under Ni2+
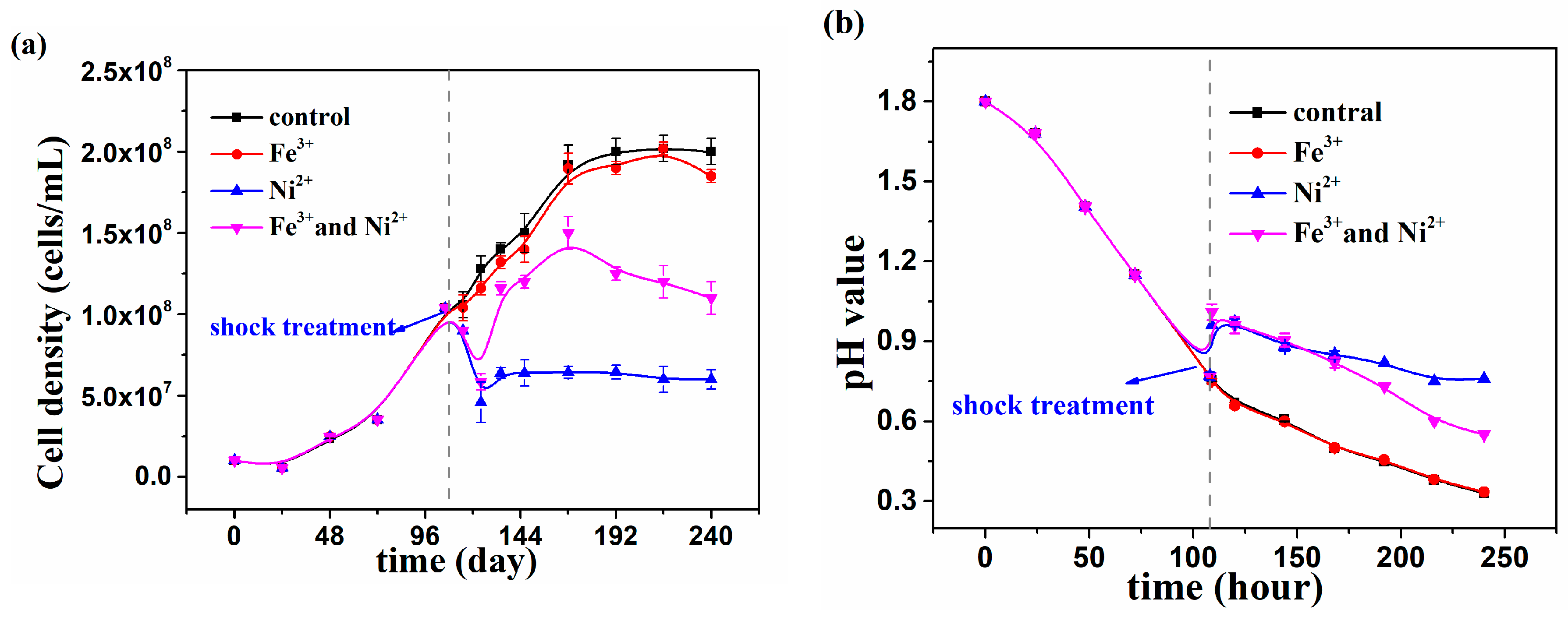
3.3. Growth of At. thiooxidans A01 under Shock Treatment of Individual and Joint Ni2+ and Fe3+
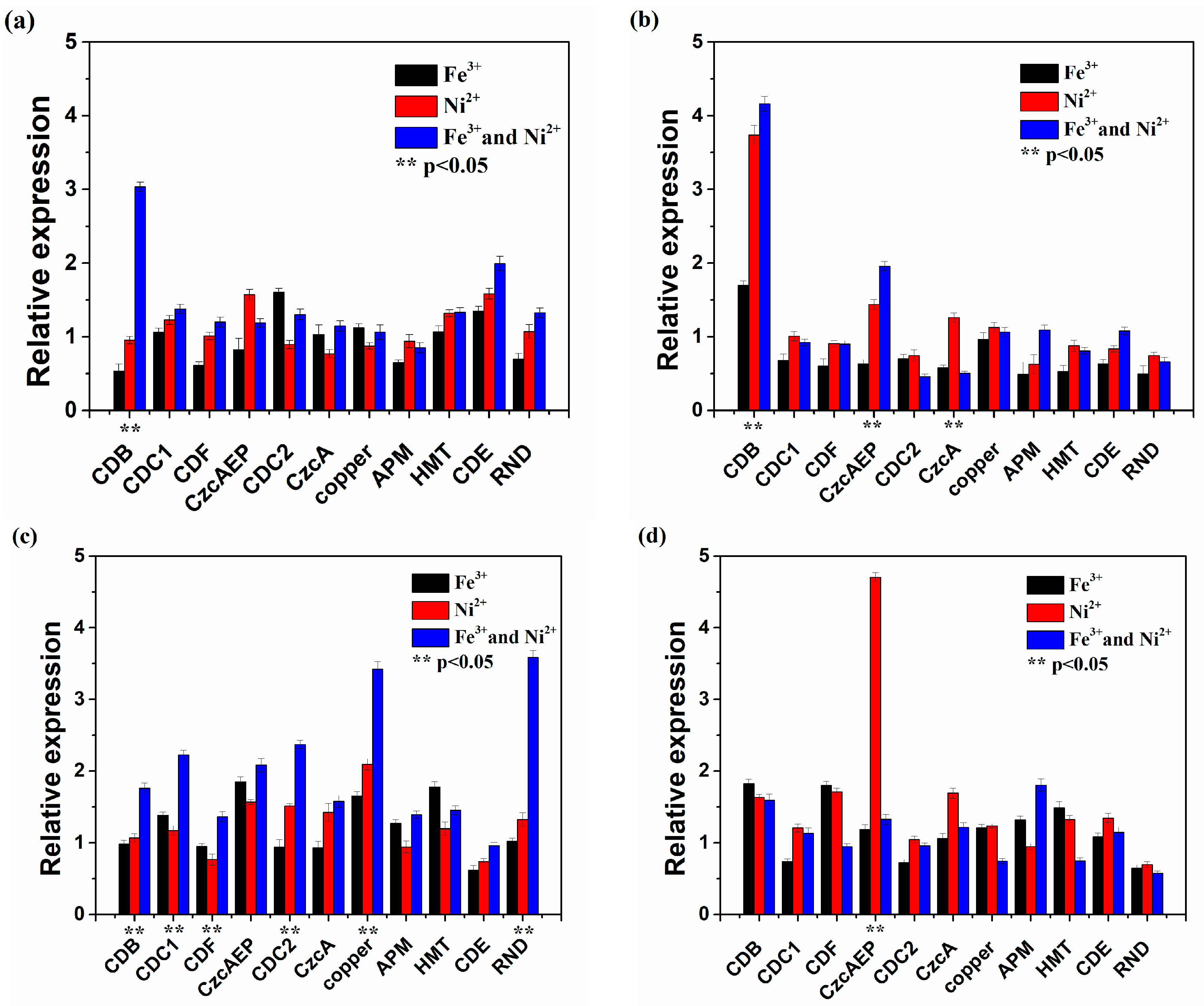
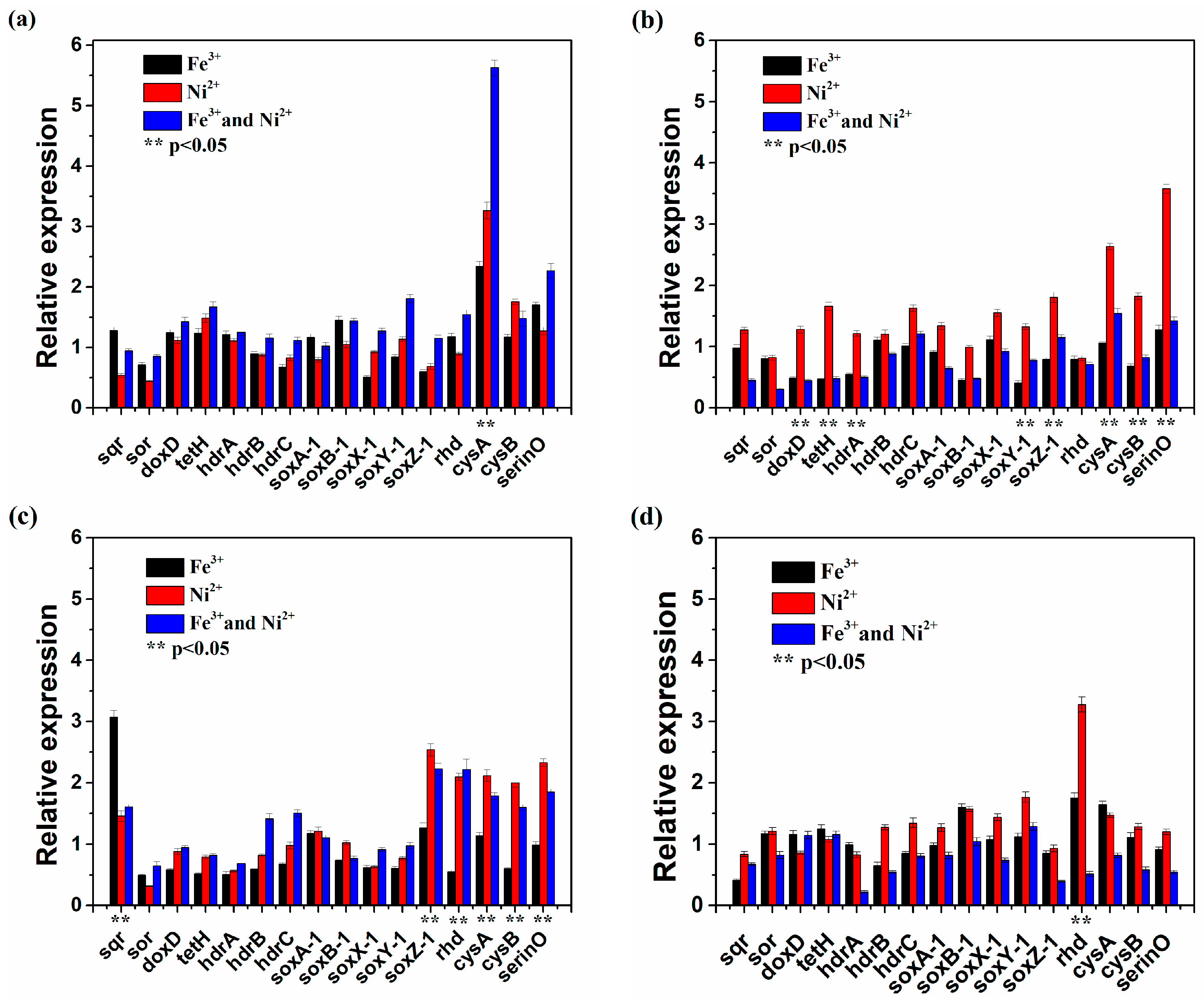
3.4. Functional Gene Expression of At. thiooxidans A01 by Single or Joint Ni2+ and Fe3+ Shock
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Schippers, A.; Hedrich, S.; Vasters, J.; Drobe, M.; Sand, W.; Willscher, S. Biomining: Metal Recovery from Ores with Microorganisms. Geobiotechnol. I Met.-Relat. Issues 2014, 141, 1–47. [Google Scholar]
- Gikas, P. Kinetic responses of activated sludge to individual and joint nickel (Ni(II)) and cobalt (Co(II)): An isobolographic approach. J. Hazard. Mater. 2007, 143, 246–256. [Google Scholar] [CrossRef] [PubMed]
- Stohs, S.J.; Bagchi, D. Oxidative mechanisms in the toxicity of metal ions. Free Radic. Biol. Med. 1995, 18, 321–336. [Google Scholar] [CrossRef]
- Macomber, L.; Hausinger, R.P. Mechanisms of nickel toxicity in microorganisms. Metallomics 2011, 3, 1153–1162. [Google Scholar] [CrossRef] [PubMed]
- Rouch, D.A.; Lee, B.T.; Morby, A.P. Understanding cellular responses to toxic agents: A model for mechanism-choice in bacterial metal resistance. J. Ind. Microbiol. 1995, 14, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Nies, D.H. Microbial heavy-metal resistance. Appl. Microbiol. Biotechnol. 1999, 51, 730–750. [Google Scholar] [CrossRef] [PubMed]
- Nies, D.H. Efflux-mediated heavy metal resistance in prokaryotes. FEMS Microbiol. Rev. 2003, 27, 313–339. [Google Scholar] [CrossRef]
- Wang, W.; Xiao, S.; Chao, J.; Chen, Q.; Qiu, G.; Liu, X. Regulation of CO2 Fixation Gene Expression in Acidithiobacillus ferrooxidans ATCC 23270 by Lix984n Shock. J. Microbiol. Biotechnol. 2008, 18, 1747–1754. [Google Scholar]
- Kim, E.H.; Nies, D.H.; McEvoy, M.M.; Rensing, C. Switch or Funnel: How RND-Type Transport Systems Control Periplasmic Metal Homeostasis. J. Bacteriol. 2011, 193, 2381–2387. [Google Scholar] [CrossRef]
- Rensing, C.; Pribyl, T.; Nies, D.H. New functions for the three subunits of the CzcCBA cation-proton antiporter. J. Bacteriol. 1997, 179, 6871–6879. [Google Scholar] [CrossRef]
- Nies, D.H. RND Efflux Pumps for Metal Cations. In Microbial Efflux Pumps: Current Research; Yu, E.W., Zhang, Q., Brown, M.H., Eds.; Caister Academic Press: Wymondham, UK, 2013; pp. 79–121. [Google Scholar]
- Mirete, S.; de Figueras, C.G.; Gonzalez-Pastor, J.E. Novel nickel resistance genes from the rhizosphere metagenome of plants adapted to acid mine drainage. Appl. Environ. Microbiol. 2007, 73, 6001–6011. [Google Scholar] [CrossRef] [PubMed]
- San Martin-Uriz, P.; Mirete, S.; Alcolea, P.J.; Gomez, M.J.; Amils, R.; Gonzalez-Pastor, J.E. Nickel-Resistance Determinants in Acidiphilium sp PM Identified by Genome-Wide Functional Screening. PLoS ONE 2014, 9, e95041. [Google Scholar] [CrossRef] [PubMed]
- Mykytczuk, N.C.S.; Trevors, J.T.; Ferroni, G.D.; Leduc, L.G. Cytoplasmic membrane response to copper and nickel in Acidithiobacillus ferrooxidans. Microbiol. Res. 2011, 166, 186–206. [Google Scholar] [CrossRef] [PubMed]
- Zhan, G.; Li, D.; Zhang, L. Aerobic bioreduction of nickel(II) to elemental nickel with concomitant biomineralization. Appl. Microbiol. Biotechnol. 2012, 96, 273–281. [Google Scholar] [CrossRef] [PubMed]
- Hallberg, K.B.; Grail, B.M.; du Plessis, C.A.; Johnson, D.B. Reductive dissolution of ferric iron minerals: A new approach for bio-processing nickel laterites. Miner. Eng. 2011, 24, 620–624. [Google Scholar] [CrossRef]
- Aston, J.E. Effects of Organic Acids and Heavy Metals on the Biomining Bacterium: Acidithiobacillus Caldus Strain BC13. Ph.D. Thesis, Montana State University (MSU), Bozeman, MT, USA, 2010. [Google Scholar]
- Xu, Y.; Yin, H.; Jiang, H.; Liang, Y.; Guo, X.; Ma, L.; Xiao, Y.; Liu, X. Comparative study of nickel resistance of pure culture and co-culture of Acidithiobacillus thiooxidans and Leptospirillum ferriphilum. Arch. Microbiol. 2013, 195, 637–646. [Google Scholar] [CrossRef] [PubMed]
- Dunbar, K.L.; Scharf, D.H.; Litomska, A.; Hertweck, C. Enzymatic Carbon-Sulfur Bond Formation in Natural Product Biosynthesis. Chem. Rev. 2017, 117, 5521–5577. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Liang, Y.; Yin, H.; Xiao, Y.; Guo, X.; Xu, Y.; Hu, Q.; Liu, H.; Liu, X. Effects of Arsenite Resistance on the Growth and Functional Gene Expression of Leptospirillum ferriphilum and Acidithiobacillus thiooxidans in Pure Culture and Coculture. BioMed Res. Int. 2015. [Google Scholar] [CrossRef]
- Yin, H.; Zhang, X.; Li, X.; He, Z.; Liang, Y.; Guo, X.; Hu, Q.; Xiao, Y.; Cong, J.; Ma, L.; et al. Whole-genome sequencing reveals novel insights into sulfur oxidation in the extremophile Acidithiobacillus thiooxidans. BMC Microbiol. 2014, 14, 179. [Google Scholar] [CrossRef]
- Gikas, P. Single and combined effects of nickel (Ni(II)) and cobalt (Co(II)) ions on activated sludge and on other aerobic microorganisms: A review. J. Hazard. Mater. 2008, 159, 187–203. [Google Scholar] [CrossRef]
- Sugio, T.; Taha, T.M.; Kanao, T.; Takeuchi, F. Increase in Fe2+-producing activity during growth of Acidithiobacillus ferrooxidans ATCC23270 on sulfur. Biosci. Biotechnol. Biochem. 2007, 71, 2663–2669. [Google Scholar] [CrossRef] [PubMed]
- Macomber, L.; Imlay, J.A. The iron-sulfur clusters of dehydratases are primary intracellular targets of copper toxicity. Proc. Natl. Acad. Sci. USA 2009, 106, 8344–8349. [Google Scholar] [CrossRef] [PubMed]
- Schalk, I.J.; Melissa, H.; Braud, A. New roles for bacterial siderophores in metal transport and tolerance. Environ. Microbiol. 2011, 13, 2844–2854. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Wu, Y.; Outten, F.W. Fur and the Novel Regulator YqjI Control Transcription of the Ferric Reductase Gene yqjH in Escherichia coli. J. Bacteriol. 2011, 193, 563–574. [Google Scholar] [CrossRef] [PubMed]
- Begg, S.L.; Eijkelkamp, B.A.; Luo, Z.; Counage, R.M.; Morey, J.R.; Maher, M.J.; Ong, C.L.; Mcewan, A.G.; Kobe, B.; Omara, M.L.; et al. Dysregulation of transition metal ion homeostasis is the molecular basis for cadmium toxicity in Streptococcus pneumoniae. Nat. Commun. 2015, 6, 6418. [Google Scholar] [CrossRef] [PubMed]
- Fisher, B.; Yarmolinsky, D.; Abdel-Ghany, S.; Pilon, M.; Pilon-Smits, E.A.; Sagi, M.; Van Hoewyk, D. Superoxide generated from the glutathione-mediated reduction of selenite damages the iron-sulfur cluster of chloroplastic ferredoxin. Plant Physiol. Biochem. 2016, 106, 228–235. [Google Scholar] [CrossRef]
- Xu, F.F.; Imlay, J.A. Silver(I), Mercury(II), Cadmium(II), and Zinc(II) Target Exposed Enzymic Iron-Sulfur Clusters when They Toxify Escherichia coli. Appl. Environ. Microbiol. 2012, 78, 3614–3621. [Google Scholar] [CrossRef]
- Aston, J.E.; Peyton, B.M.; Lee, B.D.; Apel, W.A. effects of ferrous sulfate, inoculum history, and anionic from on lead, zinc, and copper toxicity to acidithiobacillus caldus strain BC13. Environ. Toxicol. Chem. 2010, 29, 2669–2675. [Google Scholar] [CrossRef]
- Zheng, C.; Chen, M.; Tao, Z.; Zhang, L.; Zhang, X.F.; Wang, J.-Y.; Liu, J. Differential expression of sulfur assimilation pathway genes in Acidithiobacillus ferrooxidans under Cd2+ stress: Evidence from transcriptional, enzymatic, and metabolic profiles. Extremophiles 2015, 19, 429–436. [Google Scholar] [CrossRef]
- Nocito, F.F.; Lancilli, C.; Crema, B.; Fourcroy, P.; Davidian, J.-C.; Sacchi, G.A. Heavy metal stress and sulfate uptake in maize roots. Plant Physiol. 2006, 141, 1138–1148. [Google Scholar] [CrossRef]
- Sugio, T.; Fujii, M.; Takeuchi, F.; Negishi, A.; Maeda, T.; Kamimura, K. Volatilization of mercury by an iron oxidation enzyme system in a highly mercury-resistant Acidithiobacillus ferrooxidans strain MON-1. Biosci. Biotechnol. Biochem. 2003, 67, 1537–1544. [Google Scholar] [CrossRef] [PubMed]
- Pereira, C.T.; Moutran, A.; Fessel, M.; Balan, A. The sulfur/sulfonates transport systems in Xanthomonas citri pv. citri. Bmc Genom. 2015, 16, 525. [Google Scholar] [CrossRef] [PubMed]
- Denk, D.; Bock, A. L-cysteine biosynthesis in Escherichia coli: Nucleotide sequence and expression of the serine acetyltransferase (cysE) gene from the wild-type and a cysteine-excreting mutant. J. Gen. Microbiol. 1987, 133, 515–525. [Google Scholar] [CrossRef] [PubMed]
- Kawano, Y.; Suzuki, K.; Ohtsu, I. Current understanding of sulfur assimilation metabolism to biosynthesize L-cysteine and recent progress of its fermentative overproduction in microorganisms. Appl. Microbiol. Biotechnol. 2018, 102, 8203–8211. [Google Scholar] [CrossRef] [PubMed]
- Clemens, S. Evolution and function of phytochelatin synthases. J. Plant Physiol. 2006, 163, 319–332. [Google Scholar] [CrossRef] [PubMed]
- Li, S.Z.; Zhong, H.; Hu, Y.H.; Zhao, J.C.; He, Z.G.; Gu, G.H. Bioleaching of a low-grade nickel–copper sulfide by mixture of four thermophiles. Bioresour. Technol. 2014, 153, 300–306. [Google Scholar] [CrossRef]
- Rojas, C.J.A.; Tributsch, H. Bioleaching of pyrite accelerated by cysteine. Process Biochem. 2000, 35, 815–824. [Google Scholar] [CrossRef]
- Goldberg, M.; Pribyl, T.; Juhnke, S.; Nies, D.H. Energetics and topology of CzcA, a cation/proton antiporter of the resistance-nodulation-cell division protein family. J. Biol. Chem. 1999, 274, 26065–26070. [Google Scholar] [CrossRef]
- Diels, L.; Dong, Q.; Vanderlelie, D.; Baeyens, W.; Mergeay, M. The czc operon of Alcaligenes eutrophus CH34: from resistance mechanism to the removal of heavy metals. J. Ind. Microbiol. 1995, 14, 142–153. [Google Scholar] [CrossRef]
- Intorne, A.C.; Deoliveira, W.V.V.; Pereira, L.D.; Desouza, G.A. Essential role of the czc determinant for cadmium, cobalt and zinc resistance in Gluconacetobacter diazotrophicus PAI 5. Int. Microbiol. 2012, 15, 69–78. [Google Scholar]
- Eijkelkamp, B.A.; Morey, J.R.; Ween, M.P. Extracellular Zinc Competitively Inhibits Manganese Uptake and Compromises Oxidative Stress Management in Streptococcus pneumoniae. PLoS ONE 2014, 9, 89427. [Google Scholar] [CrossRef] [PubMed]
- Hassan, K.A.; Pederick, V.G.; Elbourne, L.D.H.; Ian, T.P.; James, C.P.; Christopher, A.M.; Bart, A.E. Zinc stress induces copper depletion in Acinetobacter baumannii. BMC Microbiol. 2017, 17, 59. [Google Scholar] [CrossRef] [PubMed]
- Schneider, E.; Hunke, S. ATP-binding-cassette (ABC) transport systems: Functional and structural aspects of the ATP-hydrolyzing subunits/domains. FEMS Microbiol. Rev. 1998, 22, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Mikolay, A.; Nies, D.H. The ABC-transporter AtmA is involved in nickel and cobalt resistance of Cupriavidus metallidurans strain CH34. Antonie Van Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2009, 96, 183–191. [Google Scholar] [CrossRef]
- Braud, A.; Geoffroy, V.; Hoegy, F.; Mislin, G.L.; Schalk, I.J. Presence of the siderophores pyoverdine and pyochelin in the extracellular medium reduces toxic metal accumulation in Pseudomonas aeruginosa and increases bacterial metal tolerance. Environ. Microbiol. Rep. 2010, 2, 419–425. [Google Scholar] [CrossRef] [PubMed]
- Braud, A.; Hoegy, F.; Jezequel, K.; Lebeau, T.; Schalk, I.J. New insights into the metal specificity of the Pseudomonas aeruginosa pyoverdine-iron uptake pathway. Environ. Microbiol. 2009, 11, 1079–1091. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Target Gene | Sequences (5′–3′) | Amplicon Length (bp) |
|---|---|---|---|
| Sqr-F | Sulfide quinone reductase | GCTCGGCAGCCTCAATAC | 136 |
| Sqr-R | GGTCGGACGGTGGTTACTG | ||
| Sor-F | Sulfur oxygenase reductase | AAGCCCGTGCCTAAAGTG | 266 |
| Sor-R | CTGCCATAGTTGGTGTTGT | ||
| DoxD-F | Thiosulfate:quinone oxidoreductase | CATCCCAGGACTCCACAA | 223 |
| DoxD-R | GTCGCCACCTATTCTTACTATC | ||
| TetH-F | Tetrathionate hydrolase | TGAAAGACACGCTACCCG | 270 |
| TetH-R | GGCCGCTCAATGATAACC | ||
| HdrA-F | Heterodisulfide reductase A | CCGATTTGAAGGTGAAGC | 185 |
| HdrA-R | CGGTTGCGACCATCTGTT | ||
| HdrB-F | Heterodisulfide reductase B | GTGGACCAGCGGGAAGAA | 126 |
| HdrB-R | TACCACGGCTCTGGCATCG | ||
| HdrC-F | Heterodisulfide reductase C | TATTGAGTTTGGTCGCATTG | 114 |
| HdrC-R | CCCTTGGACAGACGCTTT | ||
| soxA-I-F | Sox system related protein A | GCTCAGTCAGGGTAAGGC | 161 |
| soxA-I-R | GACAACTATTCAAACGCATC | ||
| soxB-I-F | Sox system related protein B | GCGTATTACCGATTTGCG | 198 |
| soxB-I-R | GGATTACCGGCCATGTTT | ||
| soxX-I-F | Sox system related protein X | GCAGGGTAATTGTTTGGC | 163 |
| soxX-I-R | CATATTGATGTGCGGGAT | ||
| soxY-I-F | Sox system related protein Y | GGAATGTCAGCAGTGGGTAT | 203 |
| soxY-I-R | TTCTCCGCTATGGTTGGT | ||
| soxZ-I-F | Sox system related protein Z | AAGCGGGCAAGTTGATTC | 173 |
| soxZ-I-R | CGTATTGTCTTTCCAGGTC | ||
| Rhd-F | Rhodanese | GTGGTCCTGCTTACCCTCAA | 130 |
| Rhd-R | GCCCGATAATATCCTGCTACTG | ||
| CysA-F | sulfate/thiosulfate import ATP-binding protein | GCCCCATGCAATTCAGTAGT | 208 |
| CysA-R | GCTGAAGGAGCGTTGTAAGC | ||
| CysB-F | cys regulon genes regulator | AGGCTTCATGCTTGACCAGT | 197 |
| CysB-R | GTTGTACGCCGACAATCTGA | ||
| serine O-F | serine O-acetyltransferase | GTGTGCATGCCCTGTTTATG | 197 |
| serine O-R | GTCACCAATTTCTGCCGTCT | ||
| HMT-F | heavy metal transport/detoxif -ication protein | GGCACTTCGGGTCCTCTATT | 119 |
| HMT-R | GACGATGTGATGTTCGGTTG | ||
| CzcA-F | Cobalt-zinc-cadmium resistance protein CzcA gene | GCAGATTCCCCTCGCACAGT | 123 |
| CzcA-R | CCAATACTCGTCCCCGGTTT | ||
| APM-F | arsenical pump membrane protein gene | GTTGGGTGCTTGTATTGCTG | 112 |
| APM-R | AAAAGTCGCTGTGGGTGAAA | ||
| Copper-F | copper resistance gene | GCAAGGACTTACAGGGCACG | 152 |
| Copper-R | TGACCATACGATTGATTAGACGAT | ||
| CDF-F | cation diffusion facilitator family transporter gene | TCCGCTGCTCAGTGTCTCC | 129 |
| CDF-R | GCACCACCCTCTTCGTCA | ||
| RND-F | RND family efflux transporter membrane fusion protein subunit gene | AAAGTGTCGCAACCAGTCG | 129 |
| RND-R | CAGCGGGAACCAGATAGTGT | ||
| CzcAEP-F | CzcA family heavy metal efflux pump | ATCGTGCGTTGGTGTATGGA | 117 |
| CzcAEP-R | CGAGATGTCGGGGAGTGCTT | ||
| CDB-F | Adenosine triphosphate(ATP)-binding-cassette (ABC) transporter ATP-binding protein | TTCCTCTGGGCATCAAACAA | 184 |
| CDB-R | TCATCCGTGAAATGGGTGGT | ||
| CDC1-F | ABC transporter permease, putative | TAGCCATTGCTTTTGTCCTG | 118 |
| CDC1-R | CTGGTTTCTGCGGTGGGTTG | ||
| CDC2-F | ABC transporter permease, putative | TGTTCTGGTTTCAGGTGCCC | 127 |
| CDC2-R | CCTTGCTGTTGAGTGACCGA | ||
| CDE-F | MerR family transcriptional regulator | CAAGTCTGCTCAGCACCTCA | 201 |
| CDE-R | CTGACTGCAGGAACGAATG |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, A.; Hao, X.; Xiao, Y.; Zou, K.; Liu, H.; Liu, X.; Yin, H.; Qiu, G.; Liang, Y. Responses of Acidithiobacillus thiooxidans A01 to Individual and Joint Nickel (Ni2+) and Ferric (Fe3+). Minerals 2019, 9, 82. https://doi.org/10.3390/min9020082
Chen A, Hao X, Xiao Y, Zou K, Liu H, Liu X, Yin H, Qiu G, Liang Y. Responses of Acidithiobacillus thiooxidans A01 to Individual and Joint Nickel (Ni2+) and Ferric (Fe3+). Minerals. 2019; 9(2):82. https://doi.org/10.3390/min9020082
Chicago/Turabian StyleChen, Aijia, Xiaodong Hao, Yunhua Xiao, Kai Zou, Hongwei Liu, Xueduan Liu, Huaqun Yin, Guanzhou Qiu, and Yili Liang. 2019. "Responses of Acidithiobacillus thiooxidans A01 to Individual and Joint Nickel (Ni2+) and Ferric (Fe3+)" Minerals 9, no. 2: 82. https://doi.org/10.3390/min9020082
APA StyleChen, A., Hao, X., Xiao, Y., Zou, K., Liu, H., Liu, X., Yin, H., Qiu, G., & Liang, Y. (2019). Responses of Acidithiobacillus thiooxidans A01 to Individual and Joint Nickel (Ni2+) and Ferric (Fe3+). Minerals, 9(2), 82. https://doi.org/10.3390/min9020082