

Complex Dynamic Behaviors of a Modified Discrete Leslie–Gower Predator–Prey System with Fear Effect on Prey Species



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
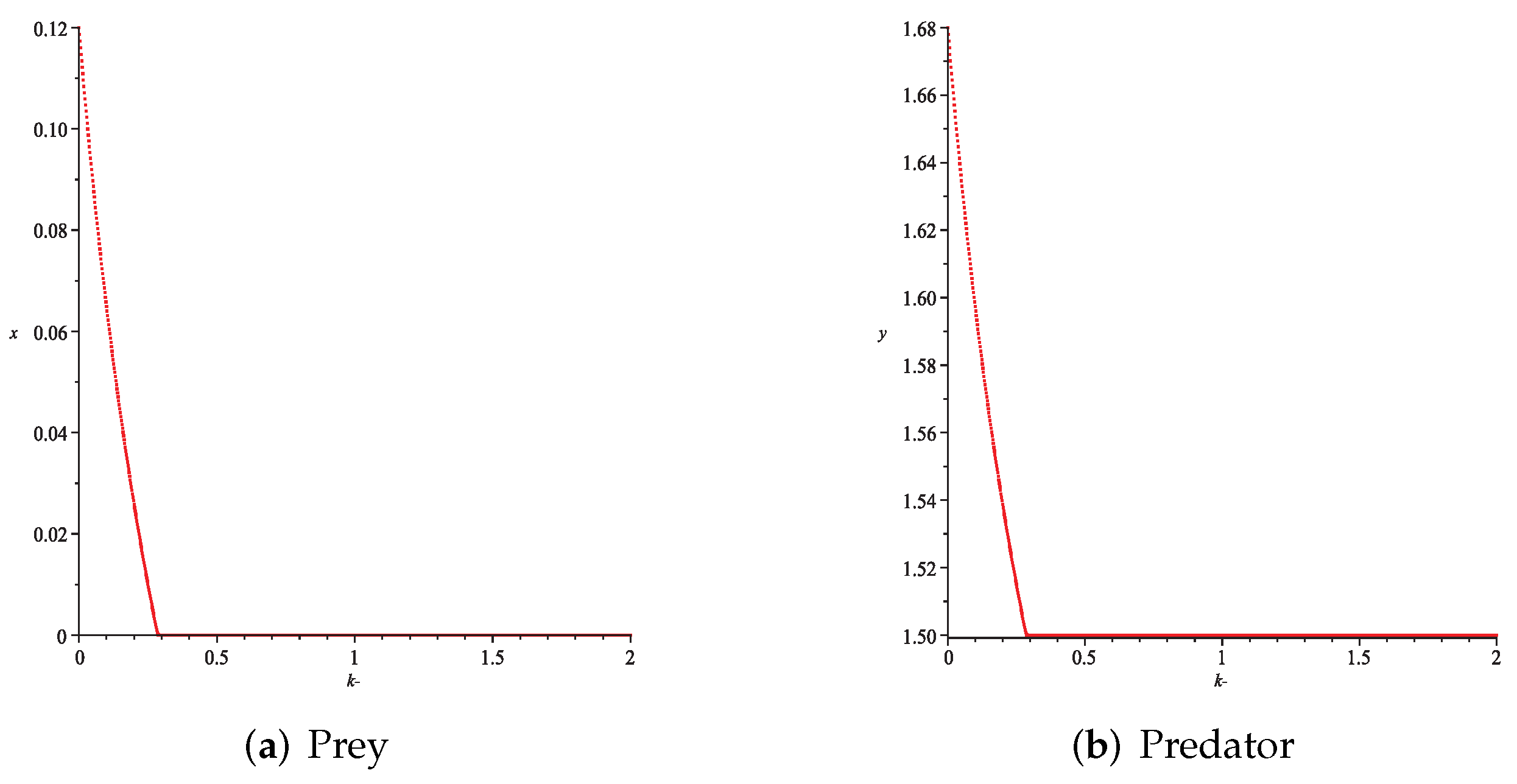{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. The Existence of Equilibria
- (1)
- , , if ;
- (2)
- , , and , if ;
- (3)
- , , , and , if .
3. The Local Stability of Equilibria
3.1. The Local Stability of Boundary Equilibria
- (1)
- A source if ;
- (2)
- A saddle if ;
- (3)
- Non-hyperbolic if .
- (1)
- A sink if and ;
- (2)
- A source if and ;
- (3)
- A saddle if one of the following conditions holds:
- (i)
- and ;
- (ii)
- and ;
- (4)
- Non-hyperbolic if or .
- (1)
- A source if ;
- (2)
- A saddle if ;
- (3)
- Non-hyperbolic if .
3.2. The Local Stability of the Positive Equilibrium
- (1)
- A sink if or ;
- (2)
- A source if one of the following conditions holds:
- (i)
- and ;
- (ii)
- and ;
- (3)
- A saddle if and ;
- (4)
- Non-hyperbolic if , where
4. The Global Attractivity of Equilibria
4.1. The Global Attractivity of the Positive Equilibrium
- (I)
- Through the first iteration, we can prove that there exist , such that and .
- (i)
- Given by the first equation of system (11), we getConsider the auxiliary equationsince , we obtain for all , where is any solution of system (23) with according to Lemma 3 in [35]. From Lemma 4 in [35], we have is nondecreasing for . Hence, according to Lemma 5 in [35], we have that for all , where is any solution of (23) satisfying the initial condition . Furthermore, because , we have according to Lemma 3 in [35]. Therefore, we haveHence, for sufficiently small there exists an integer such that if , then
- (ii)
- Given by the second equation of system (11), we getConsider the auxiliary equationSince , we obtain that for all from Lemma 3 in [35]. According to Lemma 4 in [35], we have that is nondecreasing for . Hence, according to Lemma 5 in [35], we have that for all . Therefore, as with (I) (i), we getHence, for sufficiently small there exists an integer such that if , then
- (II)
- Through the second iteration, we can prove that there exist , such that and .
- (i)
- Given by the first equation of system (11), we obtain thatConsider the auxiliary equationSince , we obtainHence for all according to Lemma 3 in [35]. From Lemma 4 in [35], we know that is nondecreasing for . Hence, according to Lemma 5 in [35], we have that for all . According to Lemma 3 in [35], we haveHence, for sufficiently small , there exists an integer such that if , then
- (ii)
- Given by the second equation of system (11), we obtainConsider the auxiliary equationSince , with a similar argument as above, we can obtainHence, for sufficiently small there exists an integer such that if , then
- (III)
- Through the third iteration, we can prove that there exist , such that and .
- (i)
- Given by the first equation of system (11), we obtain thatSince , we obtain , according to Lemma 5 in [35], we have thatHence, for sufficiently small , there exists an integer such that if , then
- (ii)
- Given by the second equation of system (11), we obtainSince , with a similar argument as above, we can obtainHence, for sufficiently small , there exists an integer such that if , then
- (IV)
- Through the fourth iteration, we can prove that there exist , such that and .
- (i)
- Similarly, from the first equation of system (11), we obtainSince , we can obtainAccording to Lemma 5 in [35], we haveHence, for sufficiently small there exists an integer such that if , then
- (ii)
- Similarly, from the second equation of system (11), we obtainSince , with a similar argument as above, we can obtainHence, for sufficiently small there exists an integer such that if , then
4.2. The Global Attractivity of the Boundary Equilibrium
5. Bifurcation Analysis
5.1. Flip Bifurcation
5.2. Transcritical Bifurcation
6. Numerical Simulations
- (1)
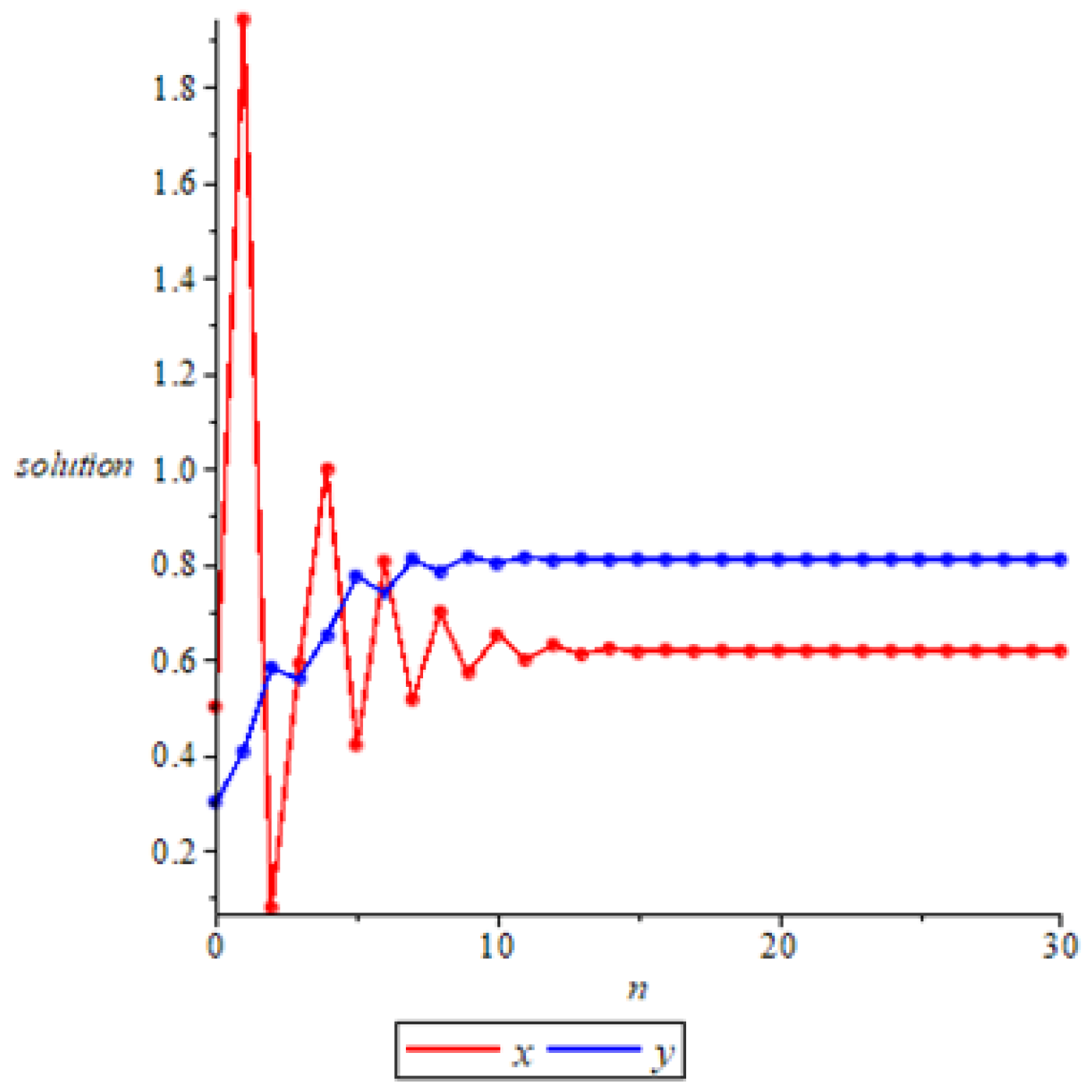
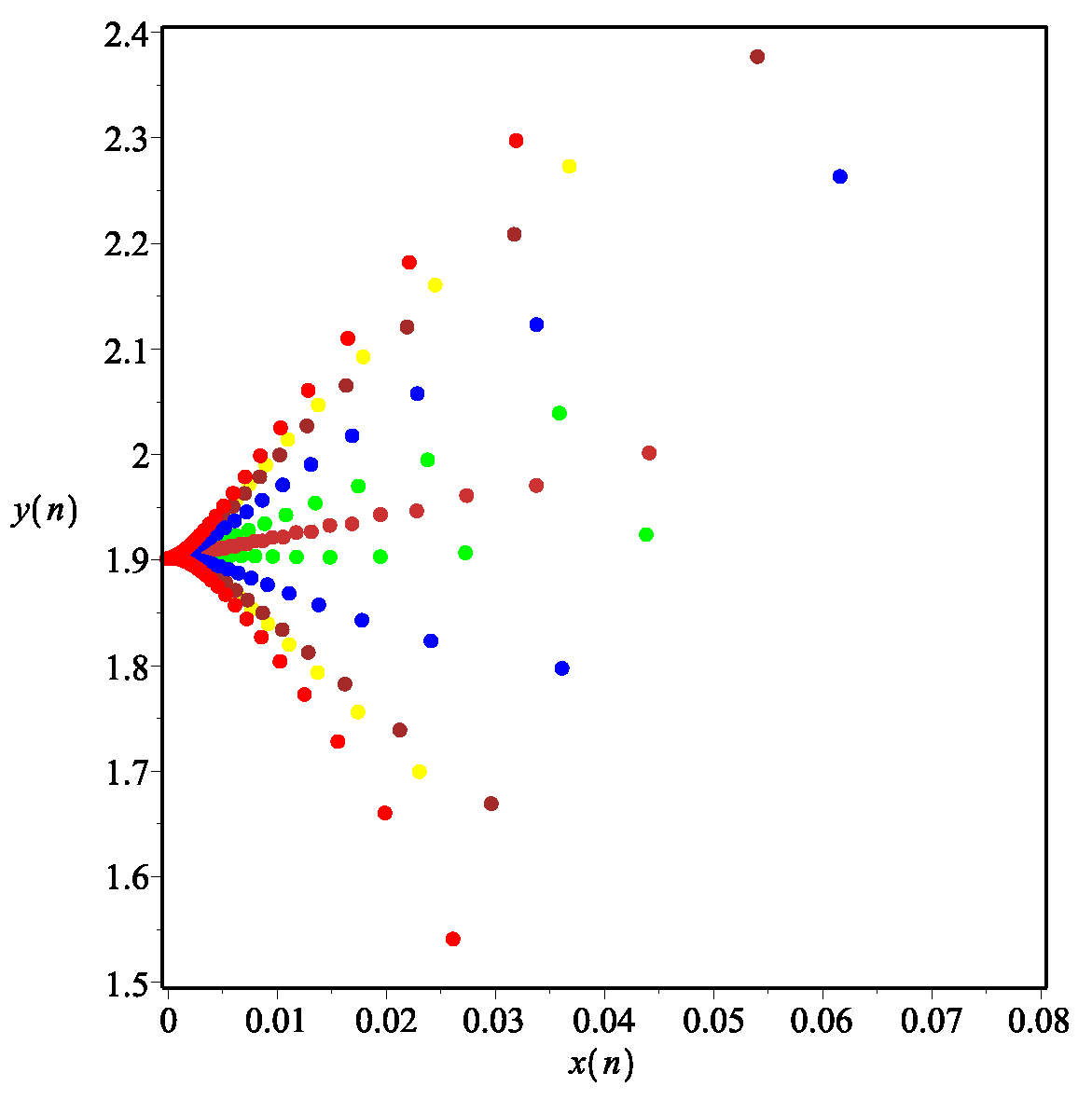
- Whenwe take the initial values of system (11) as , we have , then the system (11) admits a unique positive equilibrium according to Theorem 2. At this point, we have , then is locally asymptotically stable according to Theorem 6, as shown in Figure 2. In fact, if we take the initial values of system (11) as , we can see that is also globally stable in Figure 3, but at this point we have . It shows that it is possible for to be globally attractive even if the conditions of Theorem 7 are not satisfied. This means that sufficient conditions to ensure the globally asymptotically stable equilibrium is too strict, which is probably due to the contraction and expansion of the proof.
- (2)
- When
- (1)
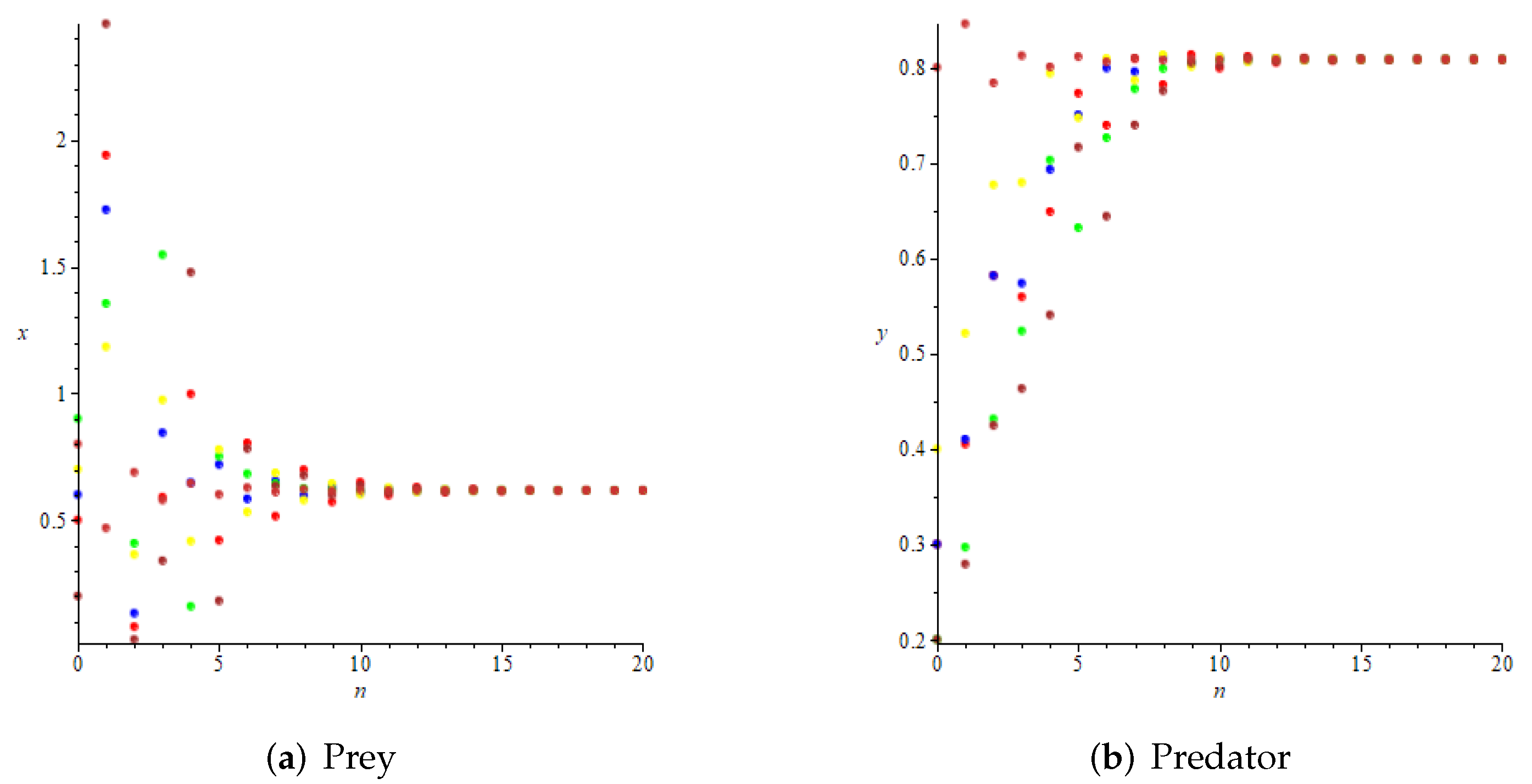
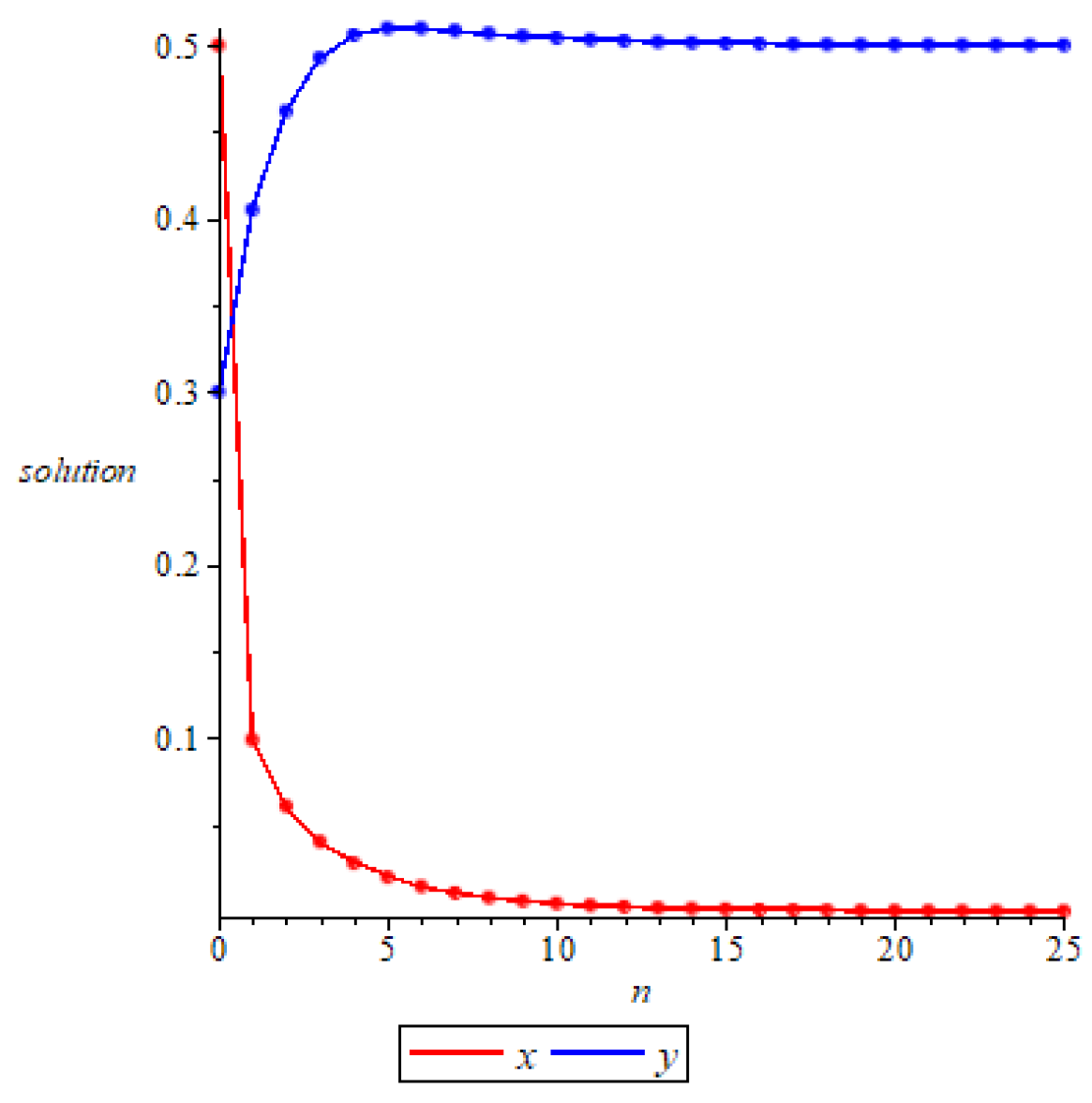
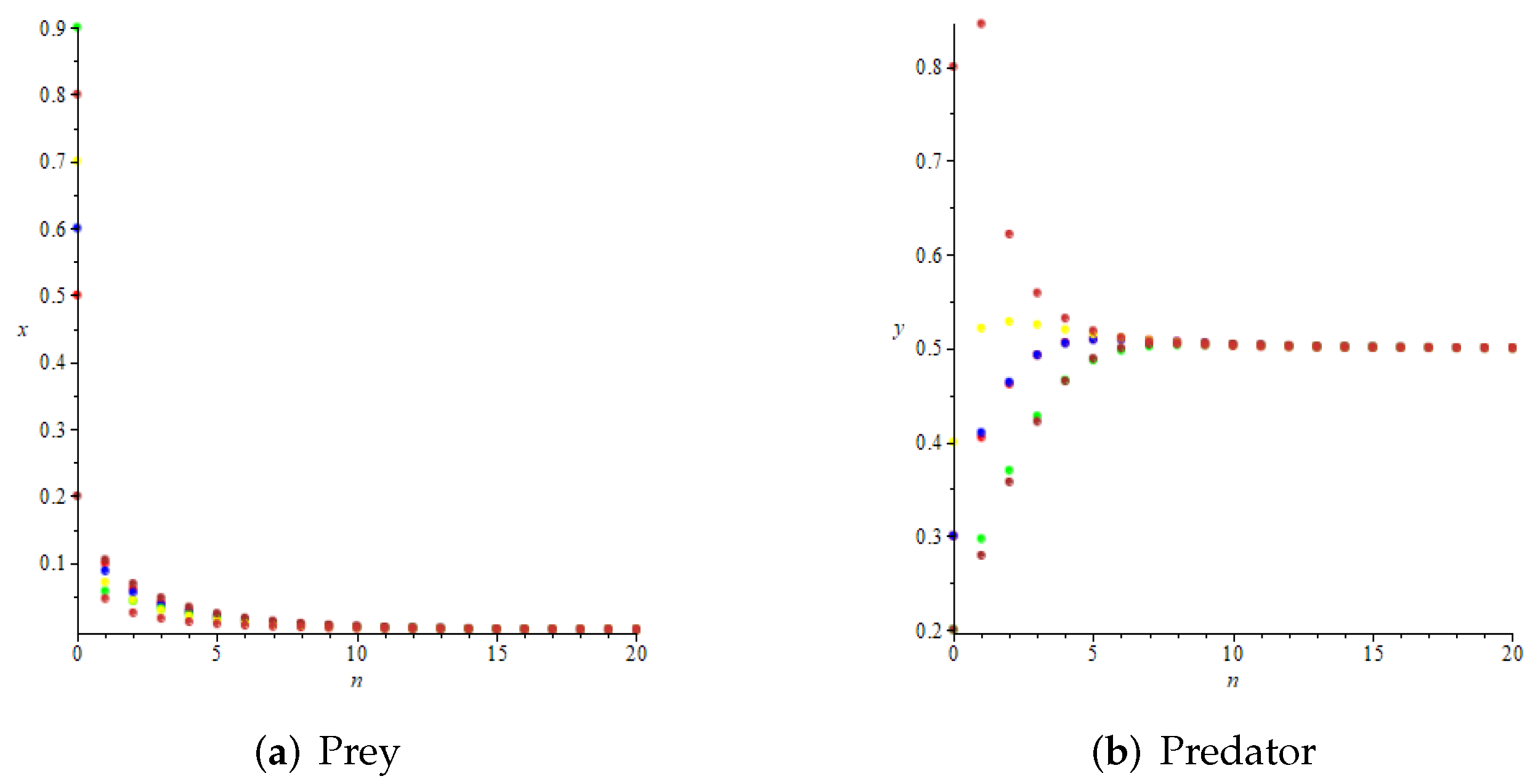
- Whenand , we have and , then is locally asymptotically stable according to Theorem 4, as shown in Figure 5. Further, we can see that and , which means that is globally attractive according to Theorem 9. We take the initial values of the system (11) asrespectively, for numerical simulation, and the results verify the accuracy of the conclusion of Theorem 9, as shown in Figure 6.
- (2)
- Whenwe haveand , which satisfy the conditions of Theorem 4, but do not meet the requirements of Theorem 9. However, we take the initial values of the system aswe can see that is also globally attractive at this time from Figure 7.
- (1)
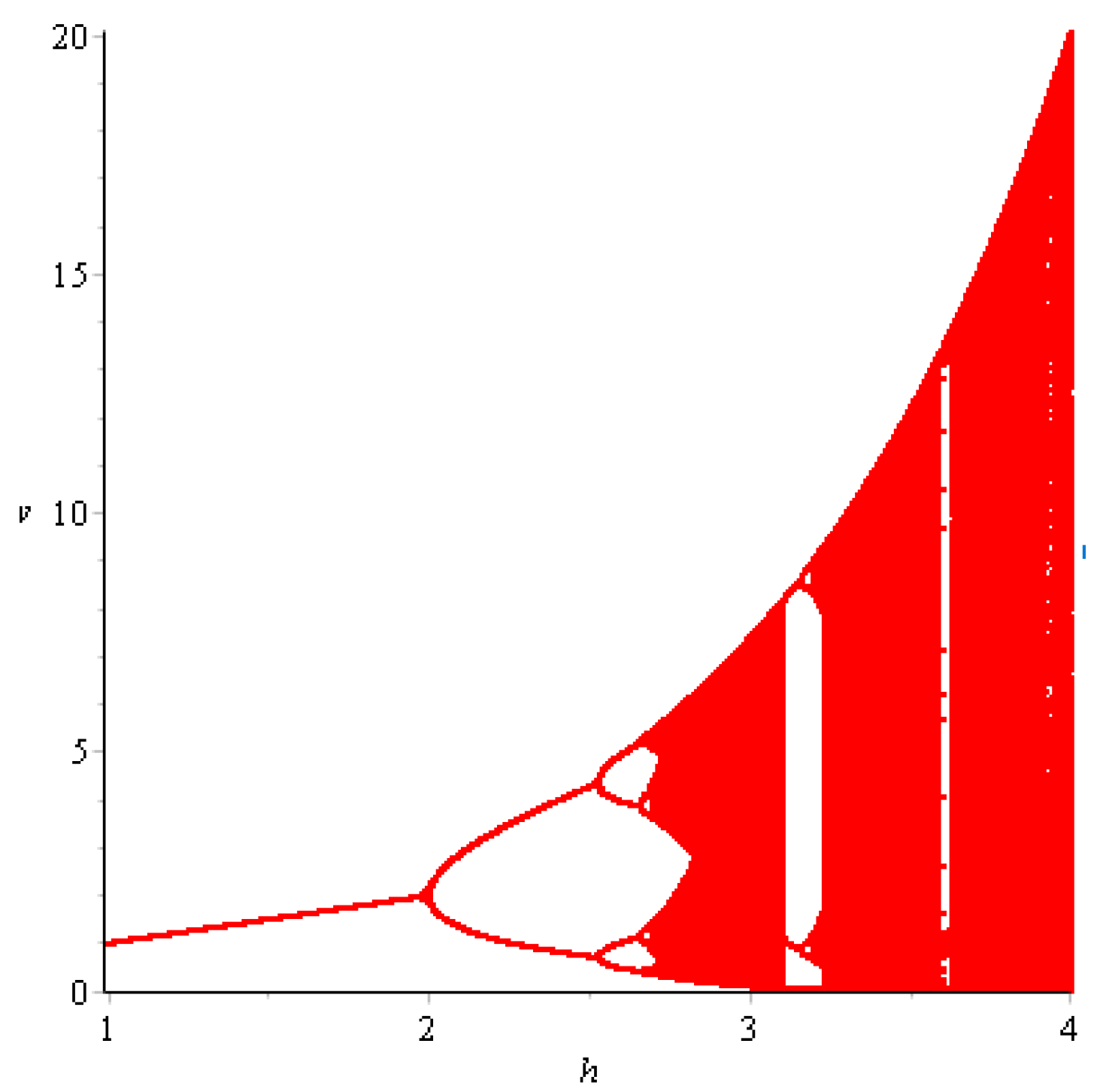
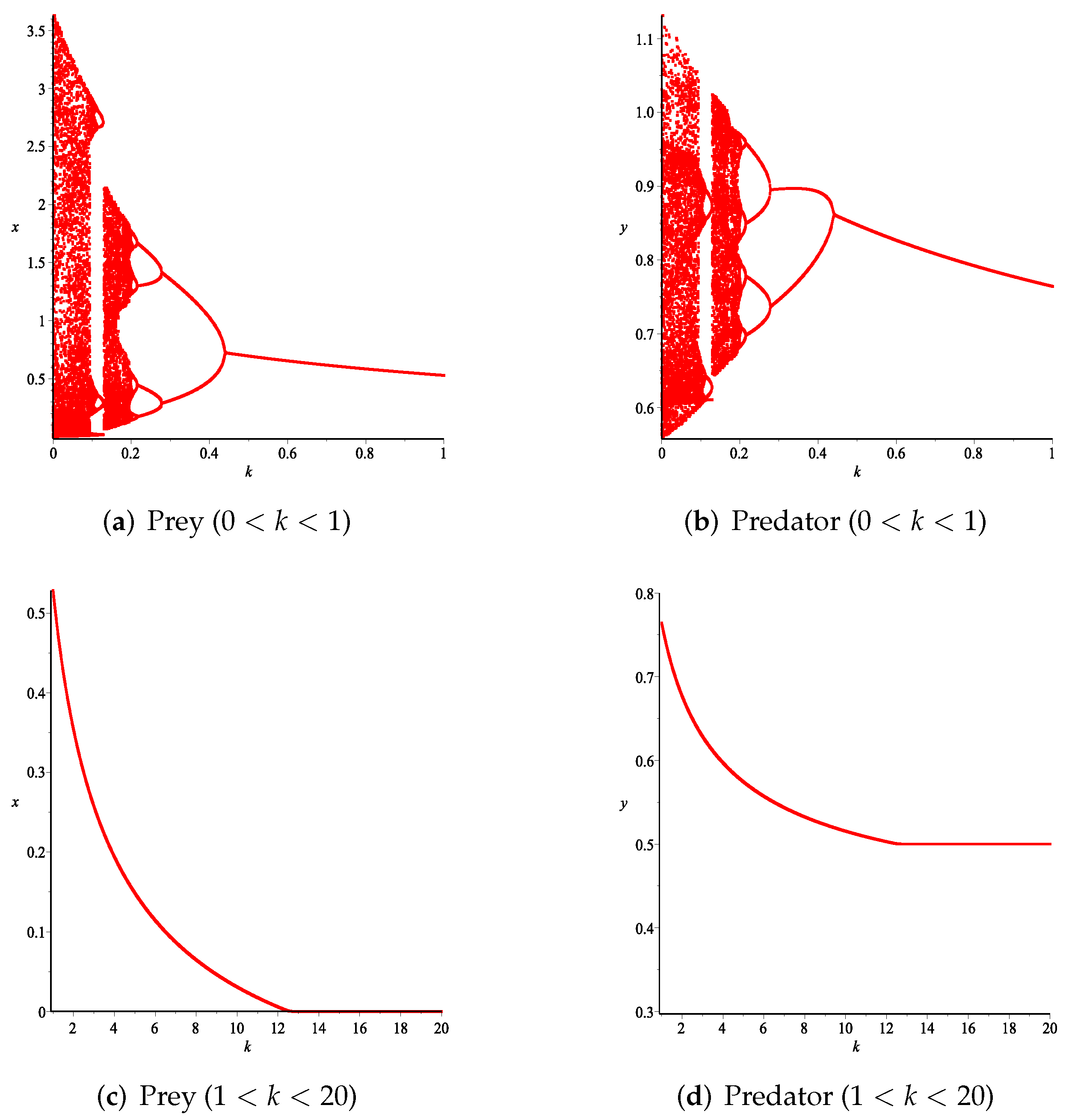
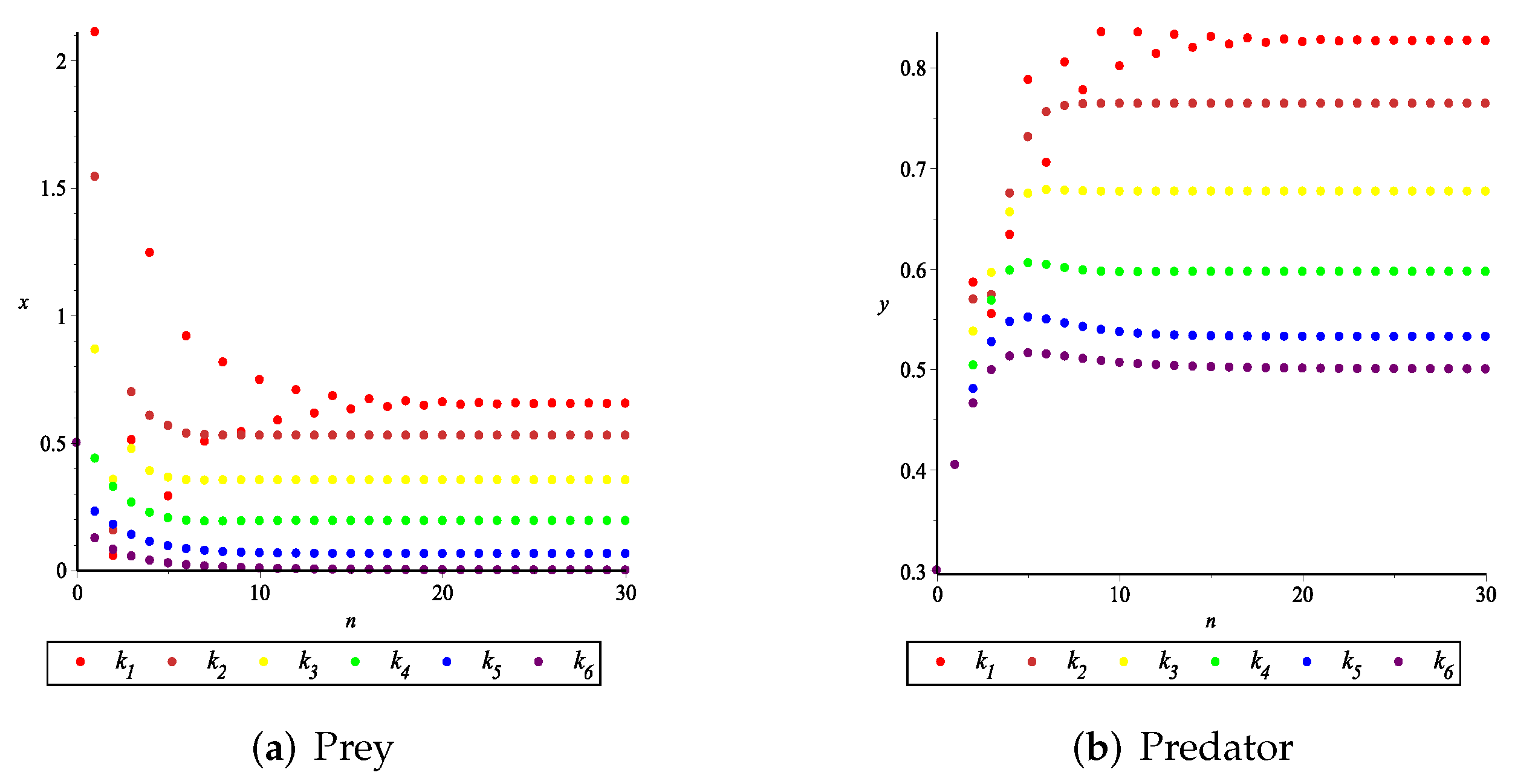
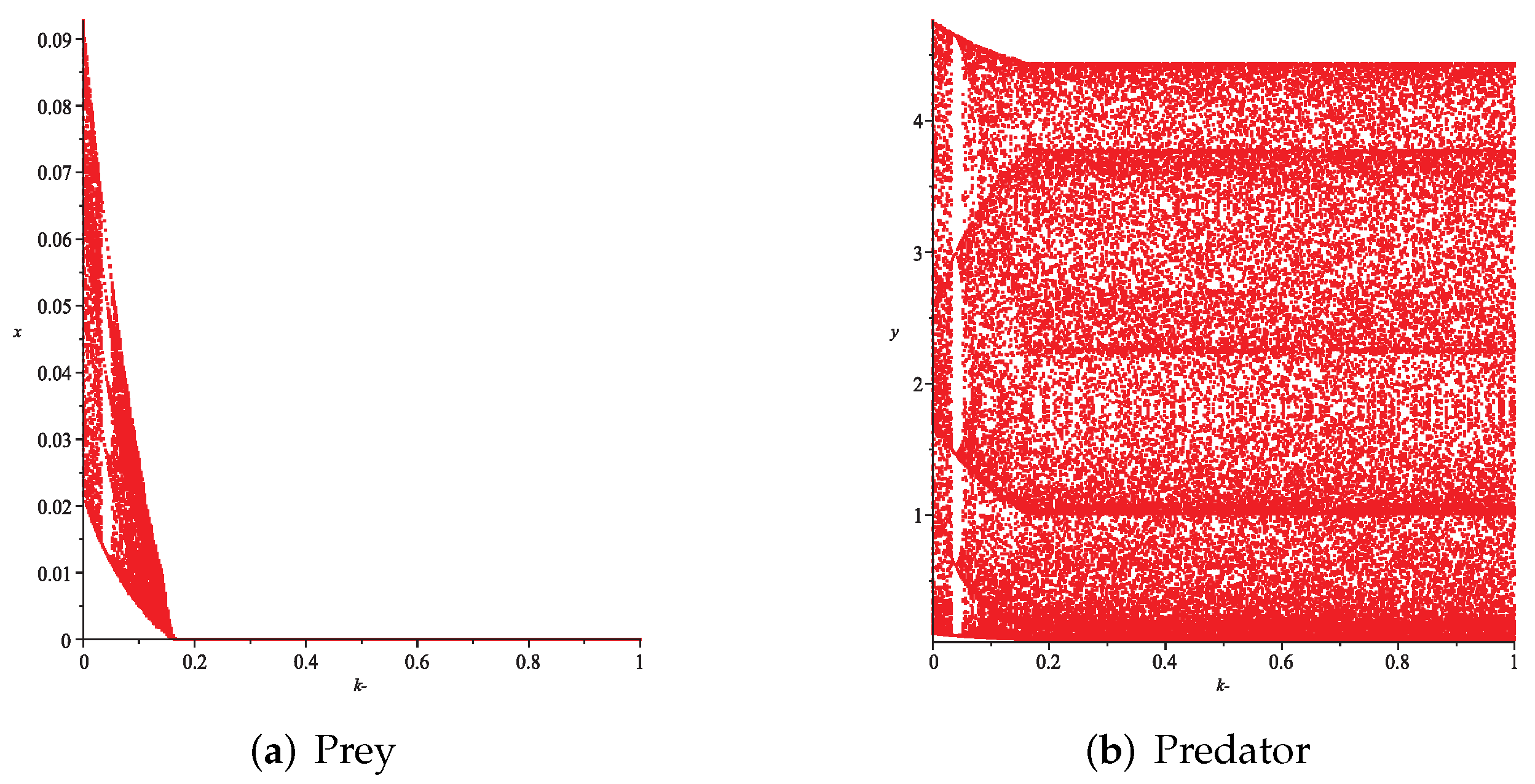
- First, we consider the case where there is no fear effect. Set each parameter value of system (11) as and set . Through numerical simulation, we find that when , system (11) has a stable equilibrium point. When continues to increase, this equilibrium point first bifurcates into a two-period orbit (), then bifurcates into a four-period orbit (), and finally chaos occurs (), as shown in Figure 8. We can see that with the increasing of grow rate of prey species, the dynamic behaviors of both predator and prey becomes complicated.
- (2)
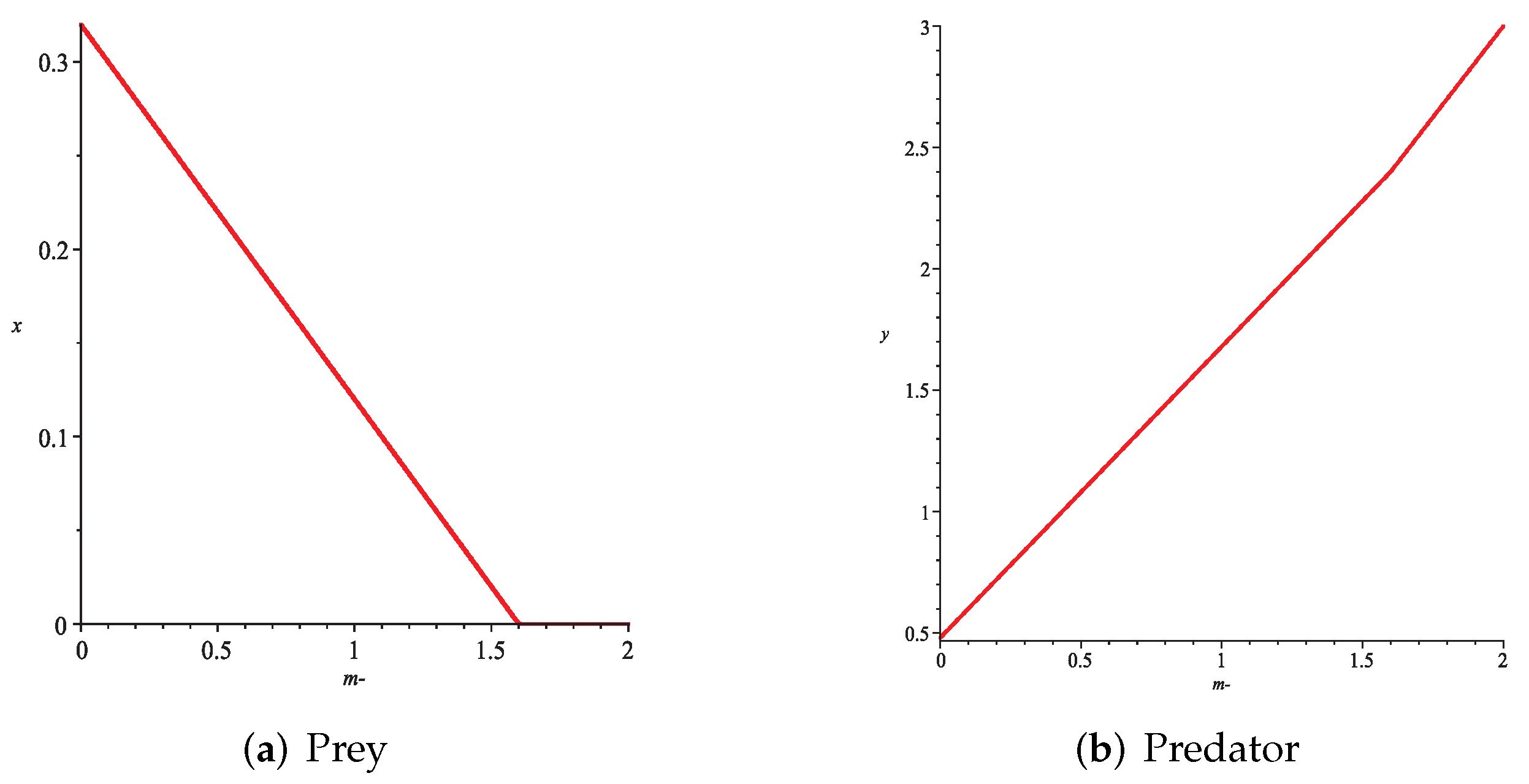
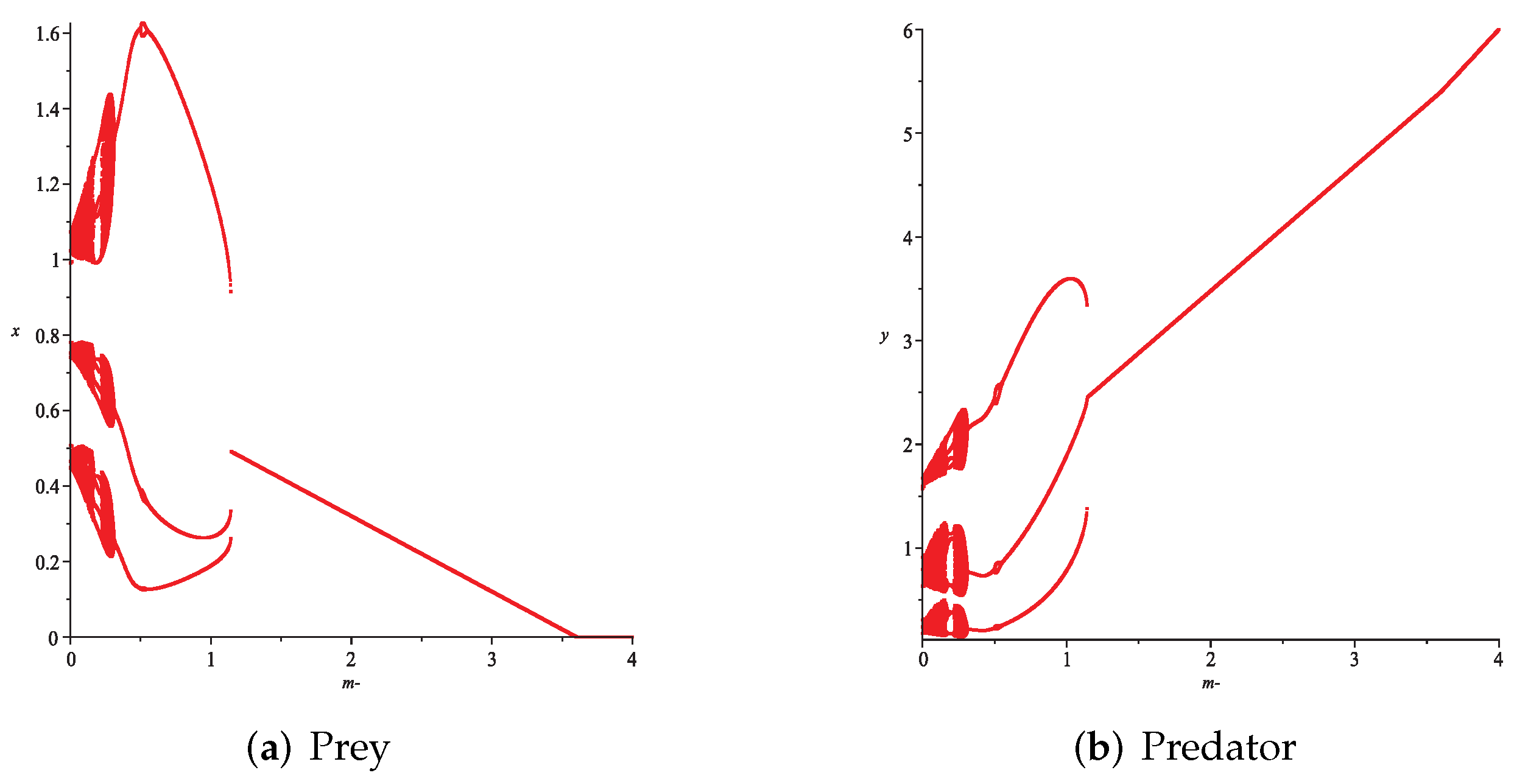
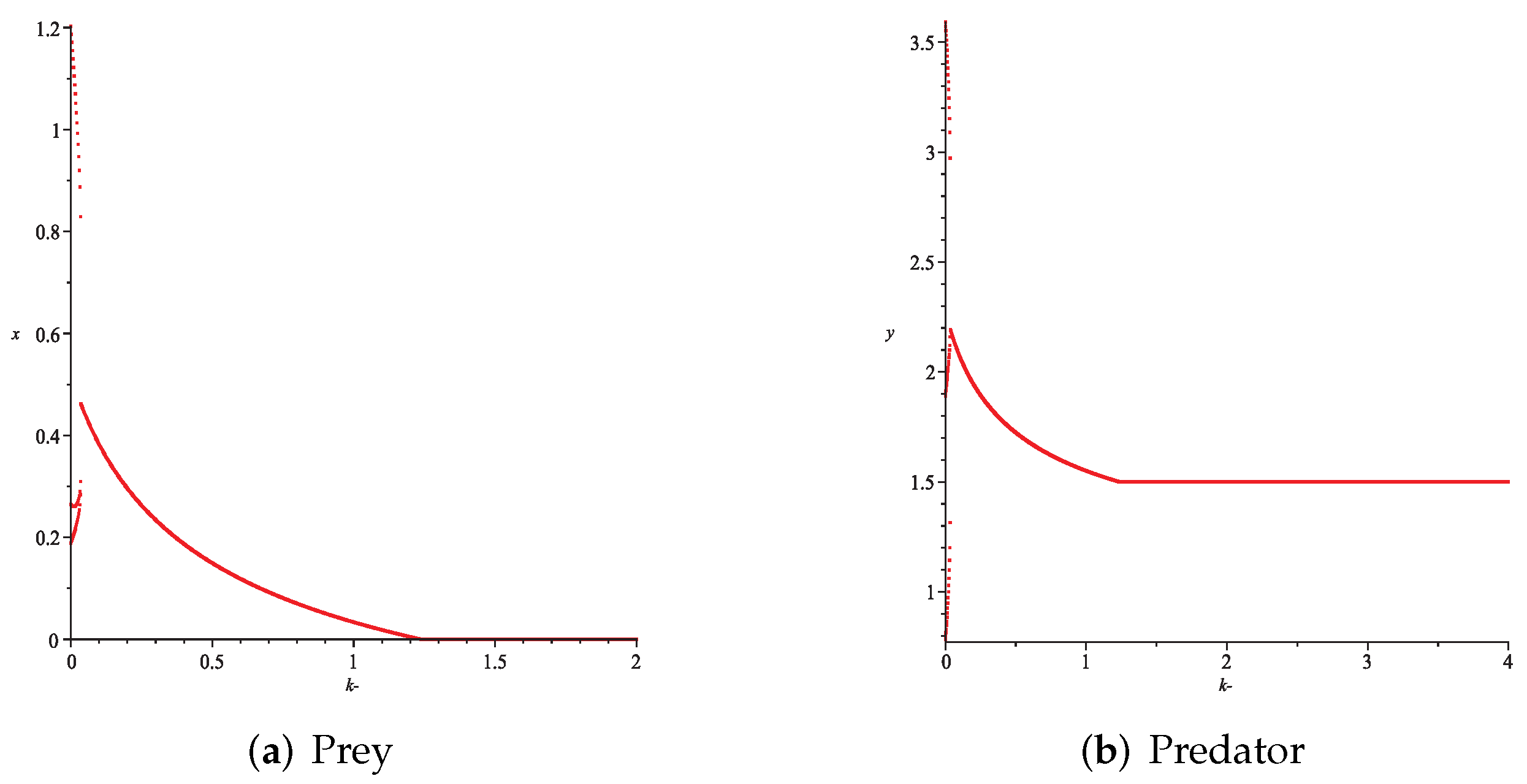
- Then, we consider the fear effect. Set each parameter value of system (11) as:set , and plot with k as the abscissa. We can see from Figure 9 that the system (11) changes from chaotic state () to eight-period orbit (), four-period orbit (), then to two-period orbit (), to stable state (), and finally to be the state that the prey is driven to extinction while the predator survive in a stable state (). This means that the stability of system (11) increases as the fear effect of prey increases within a certain range, which is similar to the result in [35]. In addition, it can be seen from Figure 9 (c)-(d) that the positive equilibrium solution of the system (11) will decrease with the increasing of k, which is consistent with the conclusion of Remark 2. If k is sufficiently large, the prey will become extinct, while the predator will not become extinct due to the presence of other food sources.
- (1)
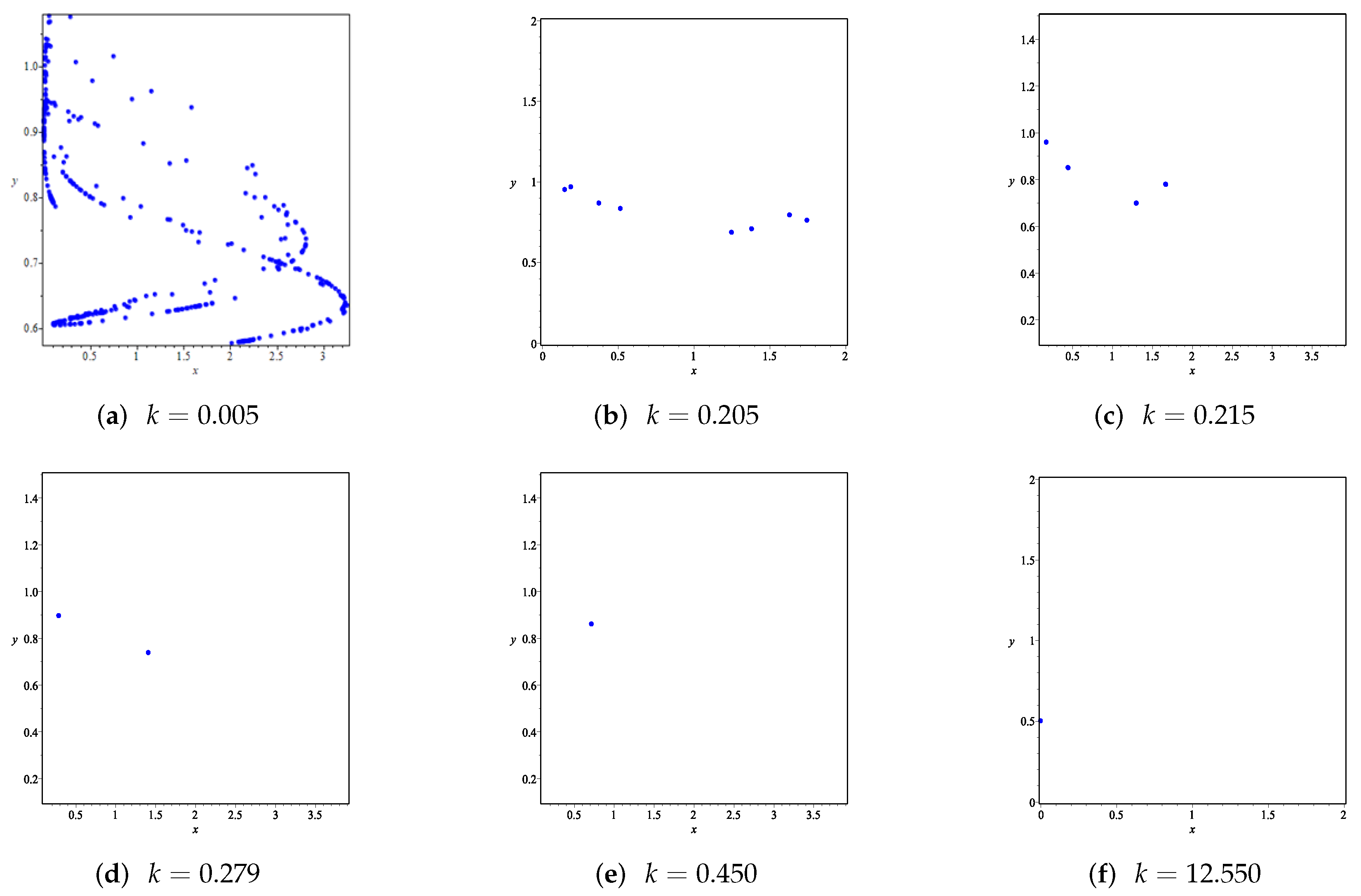
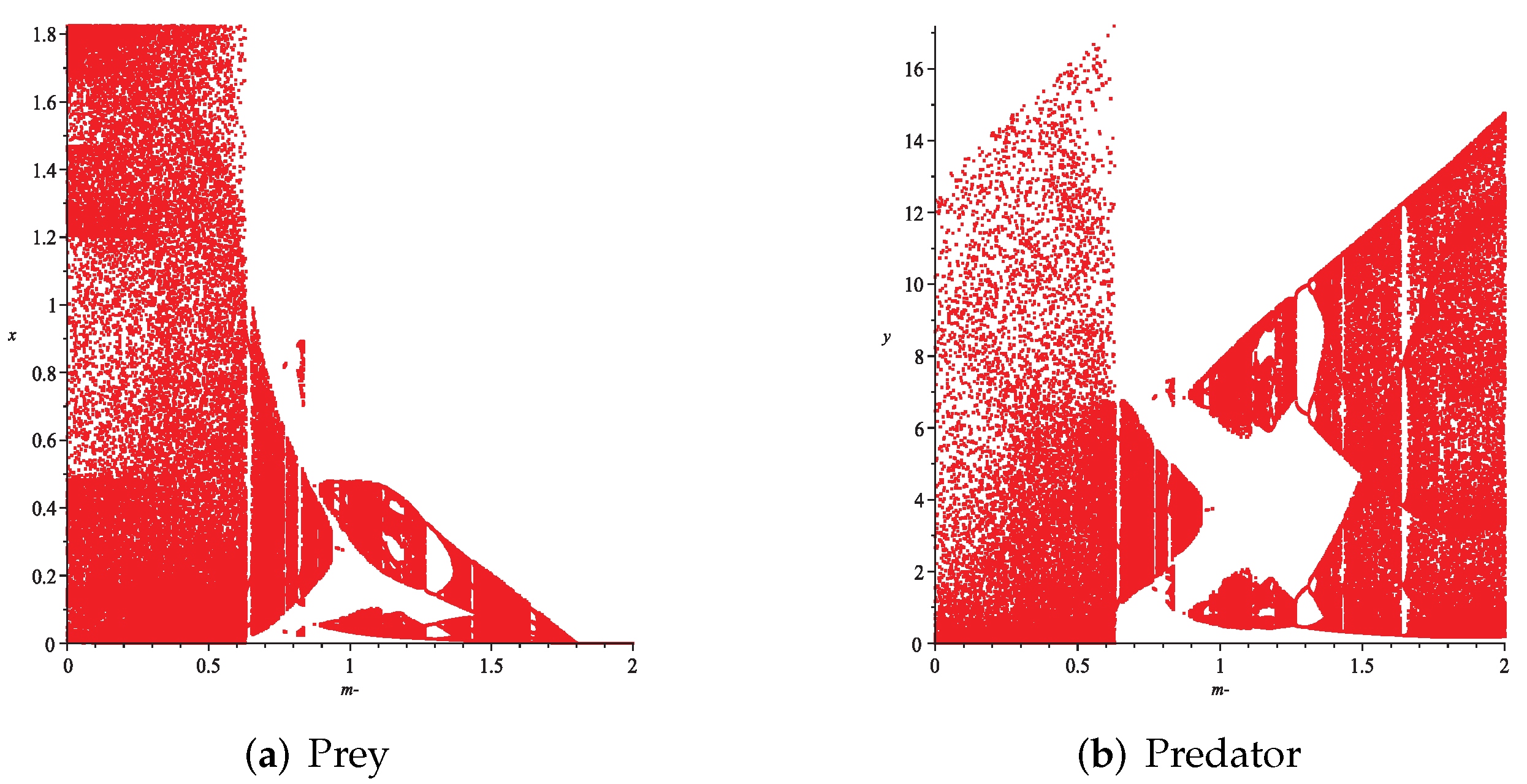
- First, we consider the case where there is no fear effect and has the variable other food resource. Set each parameter value of system (11) as and set . Depending on , , and , we give the corresponding numerical simulations (Figure 12, Figure 13, Figure 14 and Figure 15). We found that in any cases, too many other food sources can accelerate the demise of prey populations. The possible reason is that with the increasing of other food sources, the number of predator populations has increased. Therefore, although the influence of a single predator on the prey is reduced, the overall predator population will have an excessive impact on the prey population, which will lead to the decreasing of the prey population, and finally the prey species will be driven to extinction.
- (2)
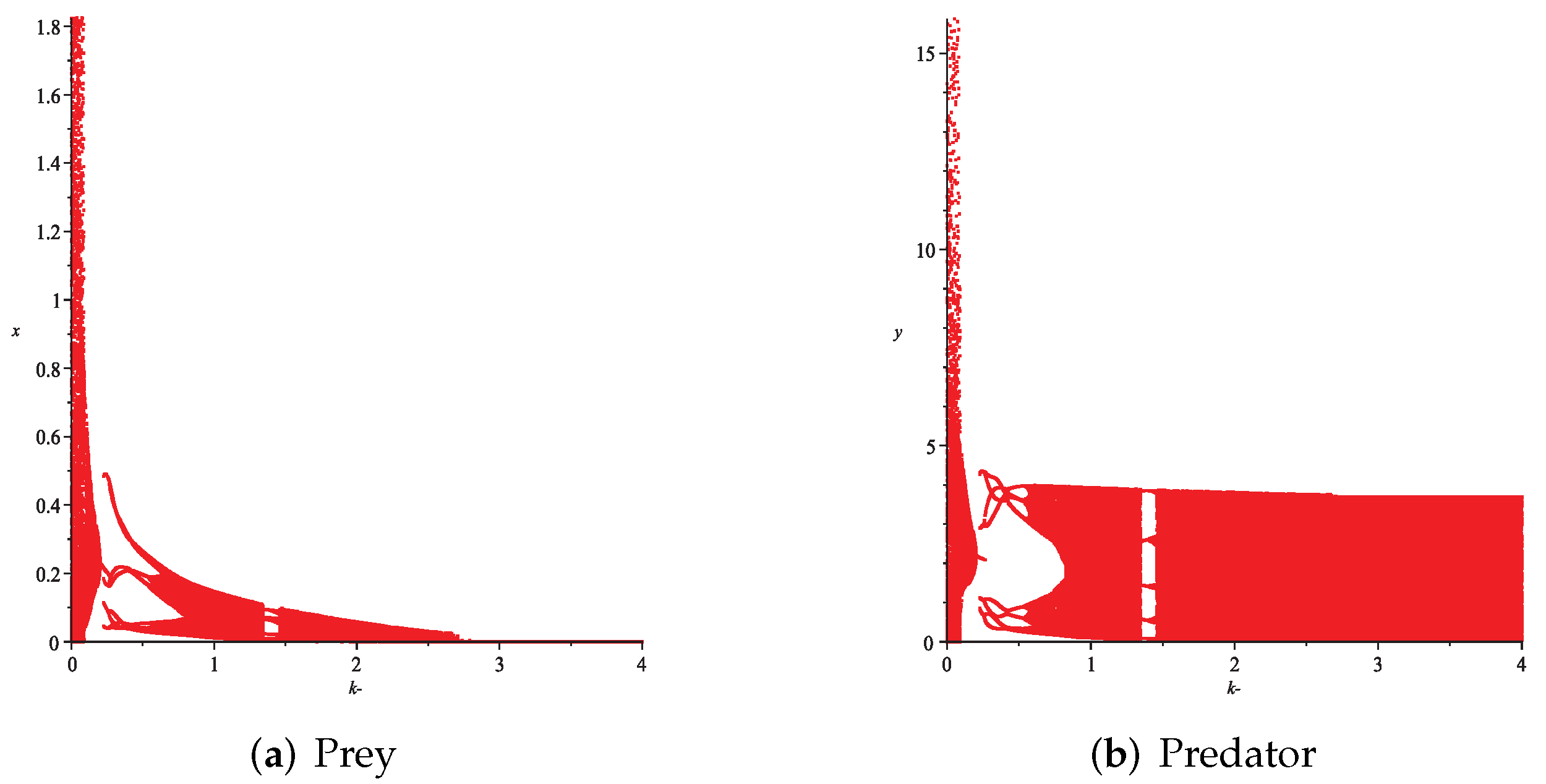
- Then, we consider the influence of fear effect. Set each parameter value of system (11) as:set , and plot with k as the abscissa. Corresponding to the four cases discussed above, we choose suitable m, such that two species could be coexist, no matter in the stable state or chaos state. Figure 16, Figure 17, Figure 18 and Figure 19 show that in any cases, if the fear effect is too large, the prey species will be driven to extinction, while depending on the intrinsic growth rate of predator species, the predator species will state in stable state for the case , and in chaos state for the case .
7. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chen, L.S.; Song, X.Y.; Lu, Z.Y. Mathematical Models and Methods in Ecology; Sichuan Science and Technology Press: Chengdu, China, 2003. (In Chinese) [Google Scholar]
- Pal, S.; Majhi, S.; Mandal, S.; Pal, N. Role of fear in a predator-prey model with Beddington-DeAngelis functional response. Z. Naturforschung A 2019, 74, 581–595. [Google Scholar] [CrossRef]
- Huang, Y.; Li, Z. The stability of a predator-prey model with fear effect in prey and square root functional response. Ann. Appl. Math. 2020, 36, 186–194. [Google Scholar]
- Leslie, P.H. A stochastic model for studying the properties of certain biological systems by numerical methods. Biometrika 1958, 45, 16–31. [Google Scholar] [CrossRef]
- Korobeinikov, A. A Lyapunov function for Leslie-Gower predator-prey models. Appl. Math. Lett. 2001, 14, 697–699. [Google Scholar] [CrossRef] [Green Version]
- Chen, F.; Chen, L.; Xie, X. On a Leslie-Gower predator-prey model incorporating a prey refuge. Nonlinear Anal. Real World Appl. 2009, 10, 2905–2908. [Google Scholar] [CrossRef]
- Chen, L.; Chen, F. Global stability of a Leslie-Gower predator-prey model with feedback controls. Appl. Math. Lett. 2009, 22, 1330–1334. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Han, M.; Chen, F. Global stability of a stage-structured predator-prey model with modified Leslie-Gower and Holling-type II schemes. Int. J. Biomath. 2012, 5, 1250057. [Google Scholar] [CrossRef]
- Yin, W.; Li, Z.; Chen, F.; He, M. Modeling Allee effect in the Leslie-Gower predator-prey system incorporating a prey refuge. Int. J. Bifurc. Chaos 2022, 32, 2250086. [Google Scholar] [CrossRef]
- Liu, T.; Chen, L.; Chen, F.; Li, Z. Stability analysis of a Leslie-Gower model with strong Allee effect on prey and fear effect on predator. Int. J. Bifurc. Chaos 2022, 32, 2250082. [Google Scholar] [CrossRef]
- Zhu, Z.; Chen, Y.; Li, Z.; Chen, F. Stability and bifurcation in a Leslie-Gower predator-prey model with Allee effect. Int. J. Bifurc. Chaos 2022, 32, 2250040. [Google Scholar] [CrossRef]
- Fang, K.; Zhu, Z.; Chen, F.; Li, Z. Qualitative and bifurcation analysis in a Leslie-Gower model with Allee effect. Qual. Theory Dyn. Syst. 2022, 21, 86. [Google Scholar] [CrossRef]
- Yu, S.; Chen, F. Almost periodic solution of a modified Leslie-Gower predator-prey model with Holling-type II schemes and mutual interference. Int. J. Biomath. 2014, 7, 1450028. [Google Scholar] [CrossRef]
- Yu, S. Effect of predator mutual interference on an autonomous Leslie-Gower predator-prey model. IAENG Int. J. Appl. Math. 2019, 49, 229–233. [Google Scholar]
- Zhu, C.; Kong, L. Bifurcations analysis of Leslie-Gower predator-prey models with nonlinear predator-harvesting. Discret. Contin. Dyn. Syst.-S 2017, 10, 1187–1206. [Google Scholar] [CrossRef] [Green Version]
- Zou, R.; Guo, S. Dynamics of a diffusive Leslie-Gower predator-prey model in spatially heterogeneous environment. Discret. Contin. Dyn. Syst.-B 2020, 25, 4189–4210. [Google Scholar] [CrossRef] [Green Version]
- Mondal, A.; Pal, A.; Samanta, G. On the dynamics of evolutionary Leslie-Gower predator-prey eco-epidemiological model with disease in predator. Ecol. Genet. Genom. 2018, 10, 100034. [Google Scholar] [CrossRef]
- Liang, Z.; Zeng, X.; Pang, G.; Liang, Y. Periodic solution of a Leslie predator-prey system with ratio-dependent and state impulsive feedback control. Nonlinear Dyn. 2017, 89, 2941–2955. [Google Scholar] [CrossRef]
- Aziz-Alaoui, M.; Okiye, M. Boundedness and global stability for a predator-prey model with modified Leslie-Gower and Holling-type II schemes. Appl. Math. Lett. 2003, 16, 1069–1075. [Google Scholar] [CrossRef] [Green Version]
- Wu, H. On a predator prey model with Leslie-Gower and prey refuge. J. Fuzhou Univ. 2010, 38, 342–346. [Google Scholar]
- Zanette, L.Y.; White, A.F.; Allen, M.C.; Clinchy, M. Perceived predation risk reduces the number of offspring songbirds produce per year. Science 2011, 334, 1398–1401. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zanette, L.; Zou, X. Modelling the fear effect in predator Cprey interactions. J. Math. Biol. 2016, 73, 1179–1204. [Google Scholar] [CrossRef]
- Zhu, Z.; Wu R., X.; Lai L., Y.; Yu X., Q. The influence of fear effect to the Lotka-Volterra predator-prey system with predator has other food resource. Adv. Differ. Equ. 2020, 2020, 237. [Google Scholar] [CrossRef]
- Firdiansyah, A.L. Effect of fear in Leslie-Gower predator-prey model with Beddington-DeAngelis functional response incorporating prey refuge. (IJCSAM) Int. J. Comput. Sci. Appl. Math. 2021, 7, 56–62. [Google Scholar] [CrossRef]
- Pal, S.; Pal, N.; Samanta, S.; Chattopadhyay, J. Fear effect in prey and hunting cooperation among predators in a Leslie-Gower model. Math. Biosci. Eng. 2019, 16, 5146–5179. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Tan, Y.; Cai, Y.; Wang, W. Impact of the fear effect on the stability and bifurcation of a Leslie-Gower predator-prey model. Int. J. Bifurc. Chaos 2020, 30, 2050210. [Google Scholar] [CrossRef]
- Sasmal, S. Population dynamics with multiple Allee effects induced by fear factors-a mathematical study on prey-predator interactions. Appl. Math. Model. 2018, 64, 1–14. [Google Scholar] [CrossRef]
- Pal, S.; Pal, N.; Samanta, S.; Chattopadhyay, J. Effect of hunting cooperation and fear in a predator-prey model. Ecol. Complex. 2019, 39, 100770. [Google Scholar] [CrossRef]
- Wang, J.; Cai, Y.; Fu, S.; Wang, W. The effect of the fear factor on the dynamics of a predator-prey model incorporating the prey refuge. Chaos 2019, 29, 083109. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Z.; Li, Z. Stability analysis of a mutual interference predator-prey model with the fear effect. J. Appl. Sci. Eng. 2019, 22, 205–211. [Google Scholar]
- Li, X.; Zhang, M. Integrability and multiple limit cycles in a predator-prey system with fear effect. J. Funct. Spaces 2019, 2019, 3948621. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Cai, Y.; Fu, S.; Wang, W. Impact of the fear effect in a prey-predator model incorporating a prey refuge. Appl. Math. Comput. 2019, 356, 328–337. [Google Scholar] [CrossRef]
- Wang, X.; Zou, X. Modeling the fear effect in predator-prey interactions with adaptive avoidance of predators. Bull. Math. Biol. 2017, 79, 1325–1359. [Google Scholar] [CrossRef] [PubMed]
- Kundu, K.; Pal, S.; Samanta, S.; Sen, A.; Pal, N. Impact of fear effect in a discrete-time predator-prey system. Bull. Calcutta Math. Soc. 2018, 110, 245–264. [Google Scholar]
- Chen, J.; He, X.; Chen, F. The influence of fear effect to a discrete-time predator-prey system with predator has other food resource. Mathematics 2021, 9, 865. [Google Scholar] [CrossRef]
- Liu, C. Precision algorithms in second-order fractional differential equations. Appl. Math. Nonlinear Sci. 2022, 7, 155–164. [Google Scholar] [CrossRef]
- Guckenheimer, J.; Holmes, P. Nonlinear Oscillations, Dynamical Systems, and Bifurcations of Vector Fields; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- Robinson, C. Dynamical Systems: Stability, Symbolic Dynamics, and Chaos; CRC Press: Boca Raton, FL, USA, 1998. [Google Scholar]



















Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, S.; Chen, F.; Li, Z.; Chen, L. Complex Dynamic Behaviors of a Modified Discrete Leslie–Gower Predator–Prey System with Fear Effect on Prey Species. Axioms 2022, 11, 520. https://doi.org/10.3390/axioms11100520
Lin S, Chen F, Li Z, Chen L. Complex Dynamic Behaviors of a Modified Discrete Leslie–Gower Predator–Prey System with Fear Effect on Prey Species. Axioms. 2022; 11(10):520. https://doi.org/10.3390/axioms11100520
Chicago/Turabian StyleLin, Sijia, Fengde Chen, Zhong Li, and Lijuan Chen. 2022. "Complex Dynamic Behaviors of a Modified Discrete Leslie–Gower Predator–Prey System with Fear Effect on Prey Species" Axioms 11, no. 10: 520. https://doi.org/10.3390/axioms11100520
APA StyleLin, S., Chen, F., Li, Z., & Chen, L. (2022). Complex Dynamic Behaviors of a Modified Discrete Leslie–Gower Predator–Prey System with Fear Effect on Prey Species. Axioms, 11(10), 520. https://doi.org/10.3390/axioms11100520