
A Computational Approach to a Model for HIV and the Immune System Interaction

 , , and
, , and
Abstract
:1. Introduction
2. Main Objectives
3. Mathematical Formulation of HIV Model
4. Modified Formulation of HIV Model
4.1. Uninfected Steady State
4.2. Infected Steady State
4.3. Reproduction Number
4.4. Jacobian Matrix
4.5. Stability Analysis
- (a)
- ,
- (b)
5. The Numerical Methods
5.1. The Continuous Galerkin–Petrov Method
The cGP(2) Method
5.2. The Classical Explicit Runge–Kutta Method
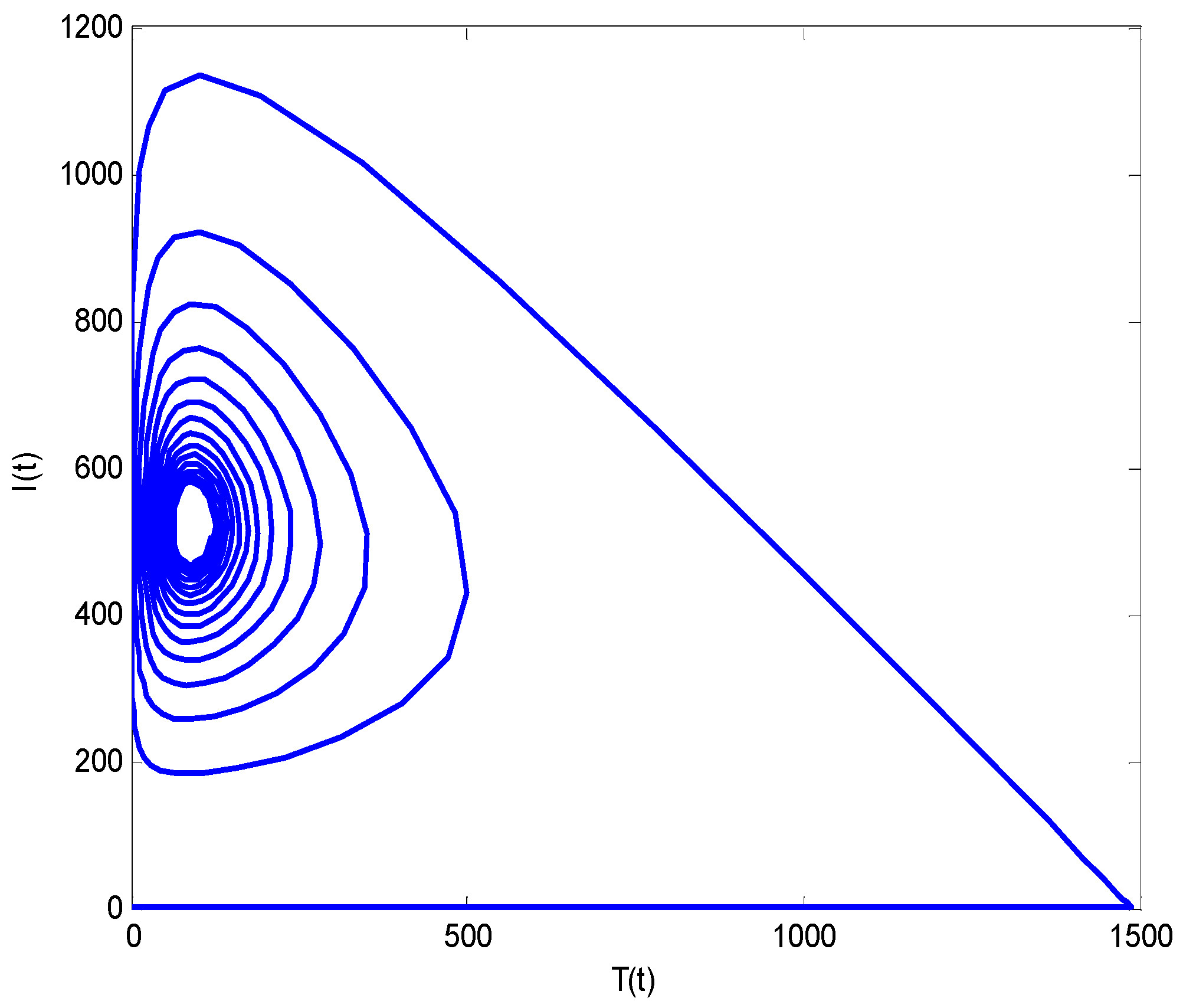
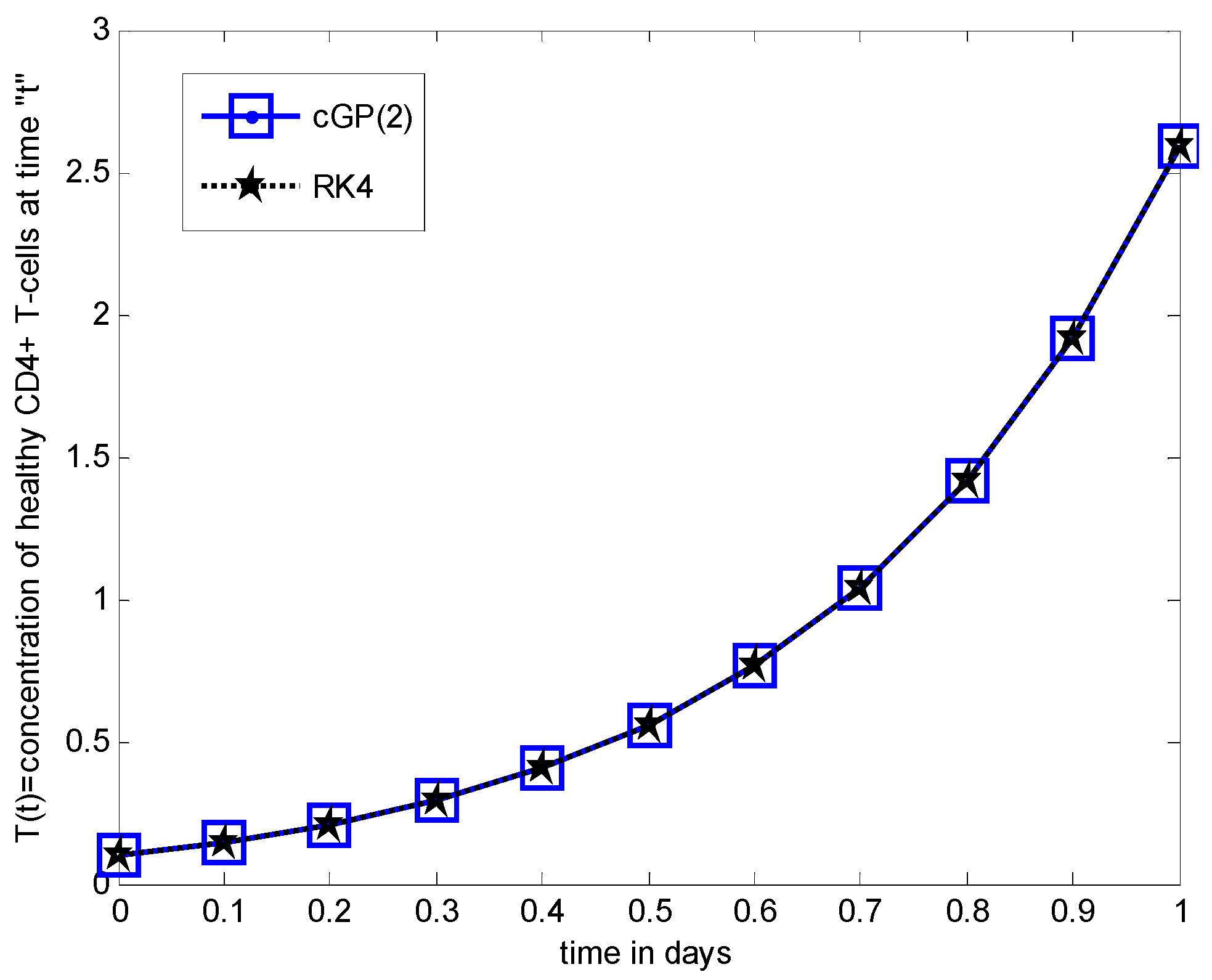
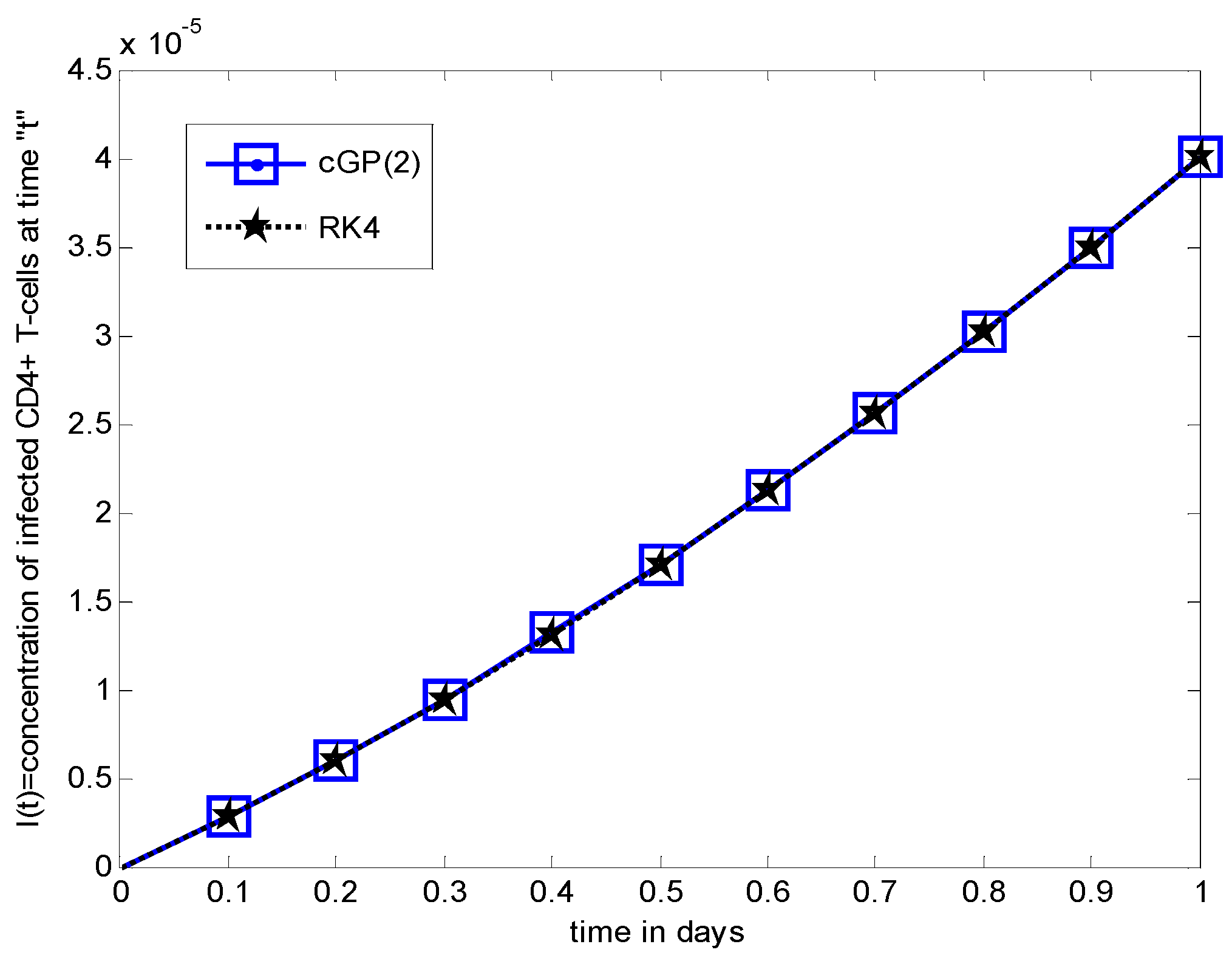
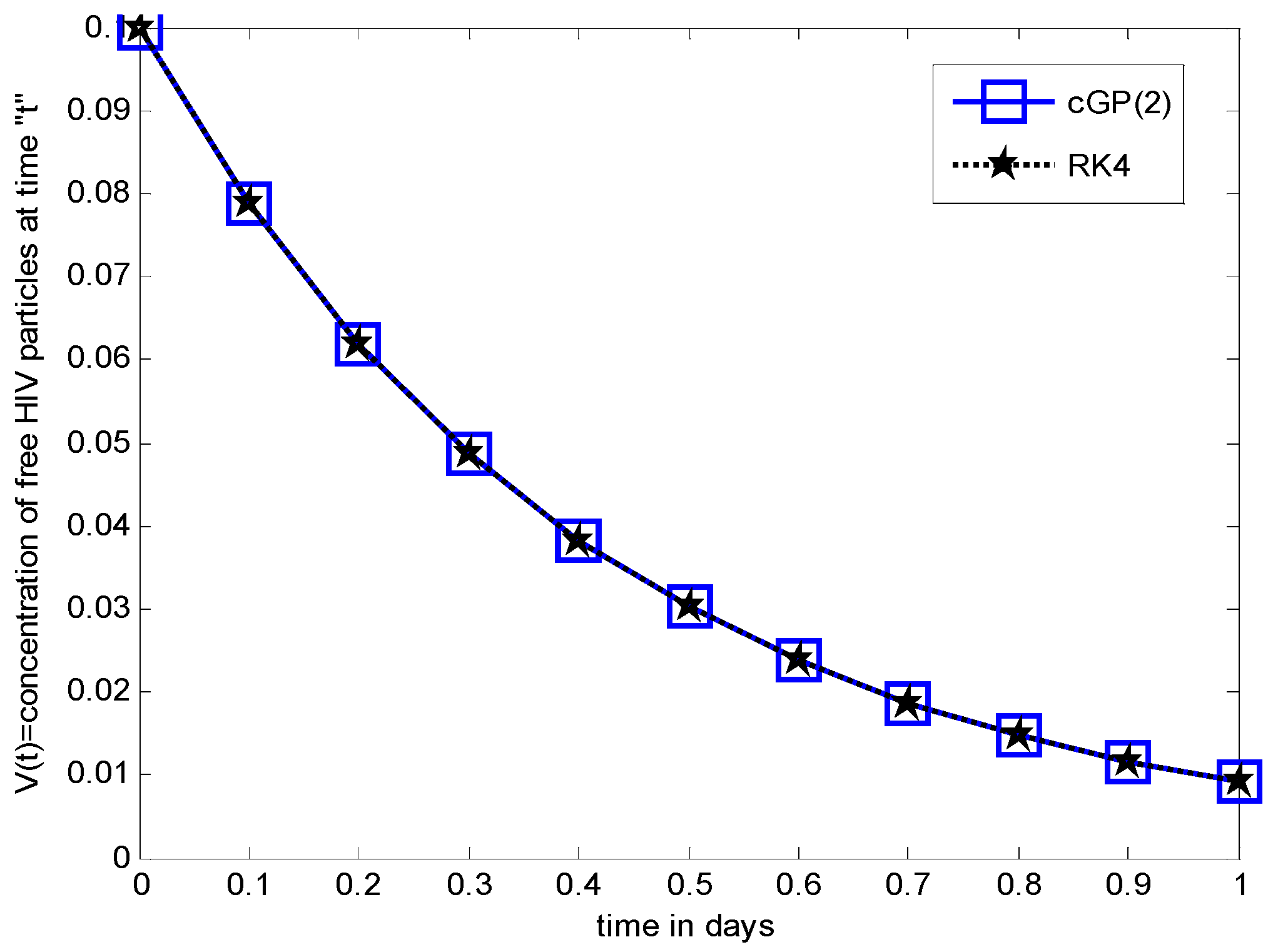
Numerical Results and Discussions
6. Comparison between the Results of Proposed Method and other Classical Methods
7. Conclusions
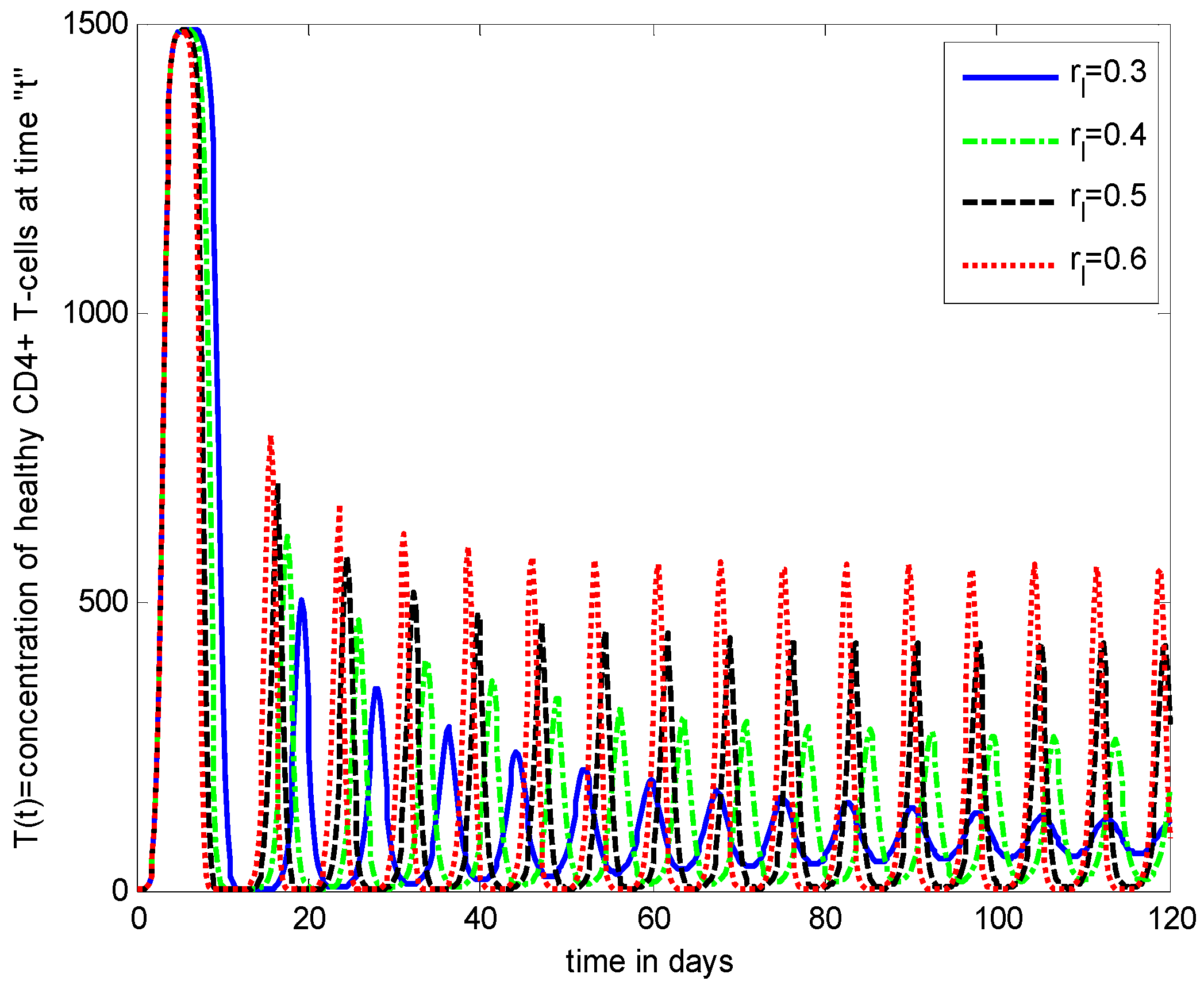
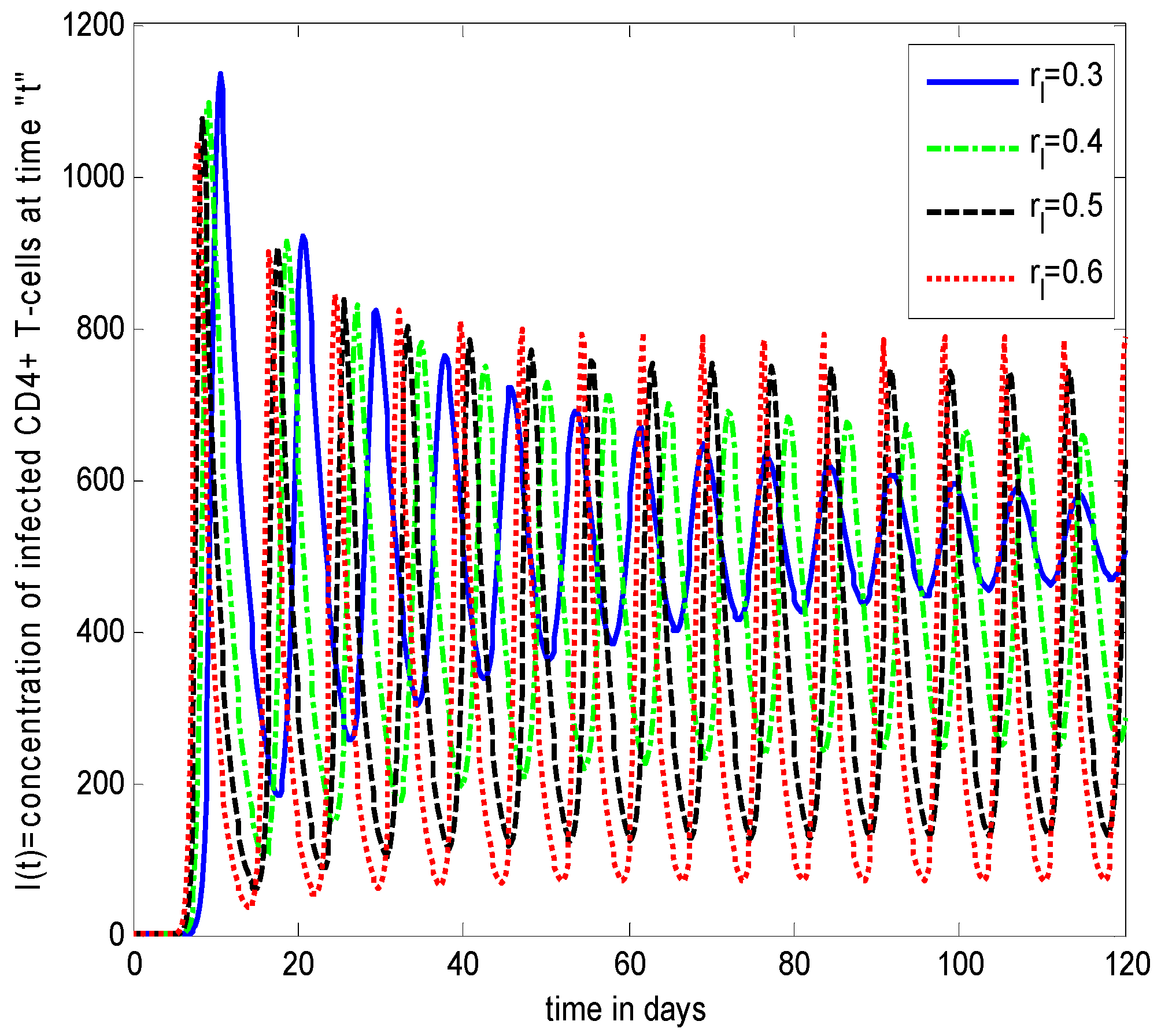
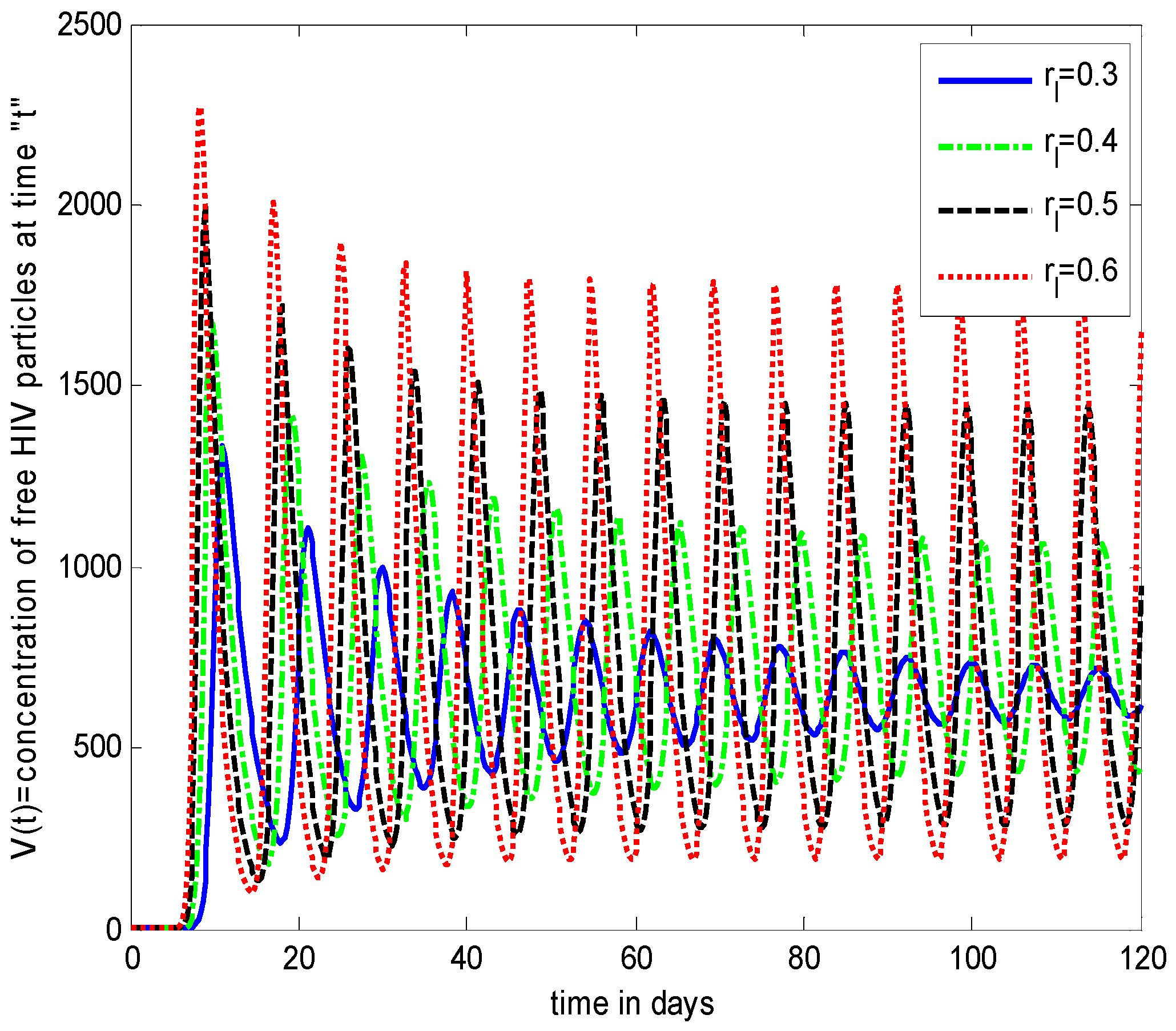
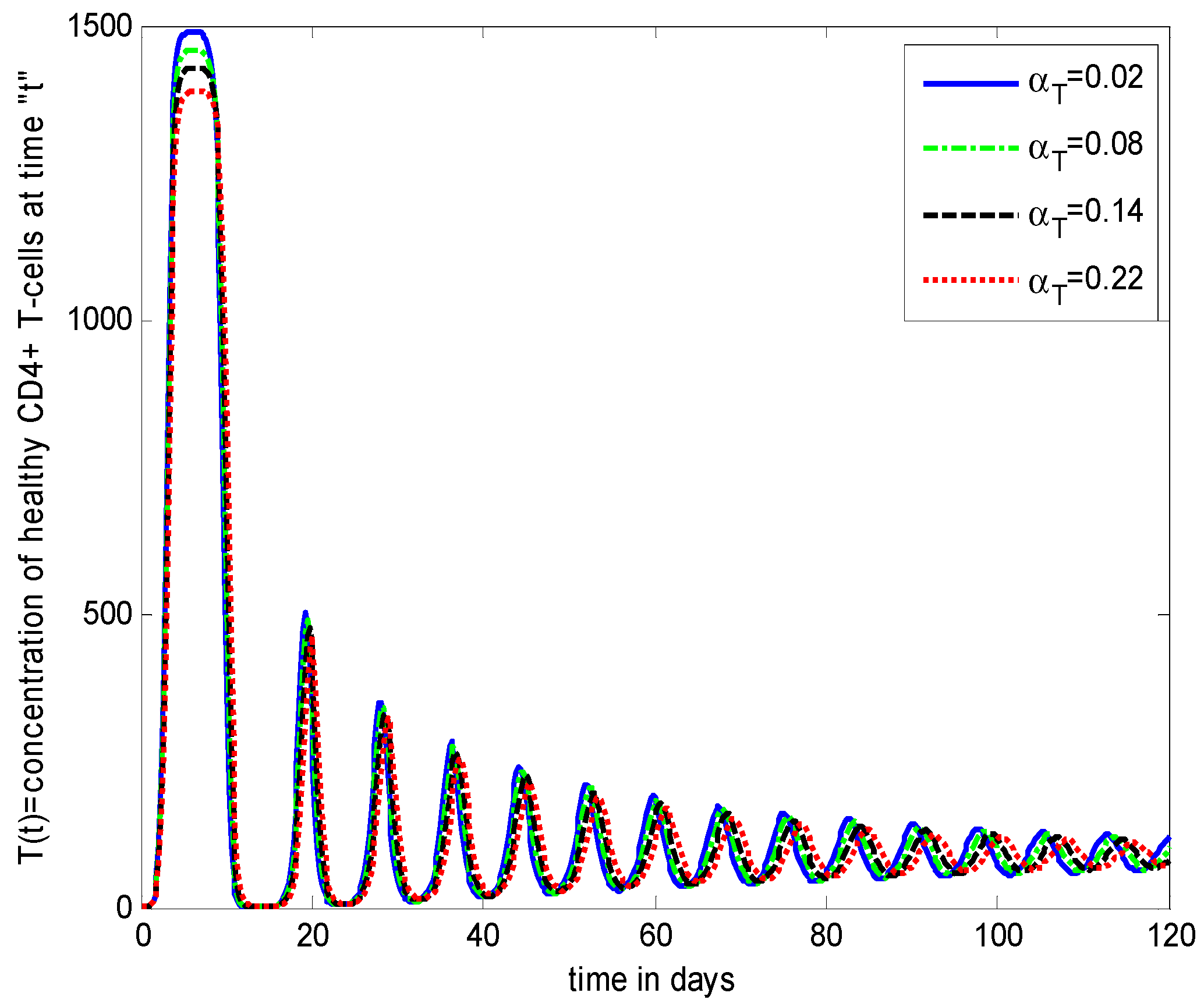
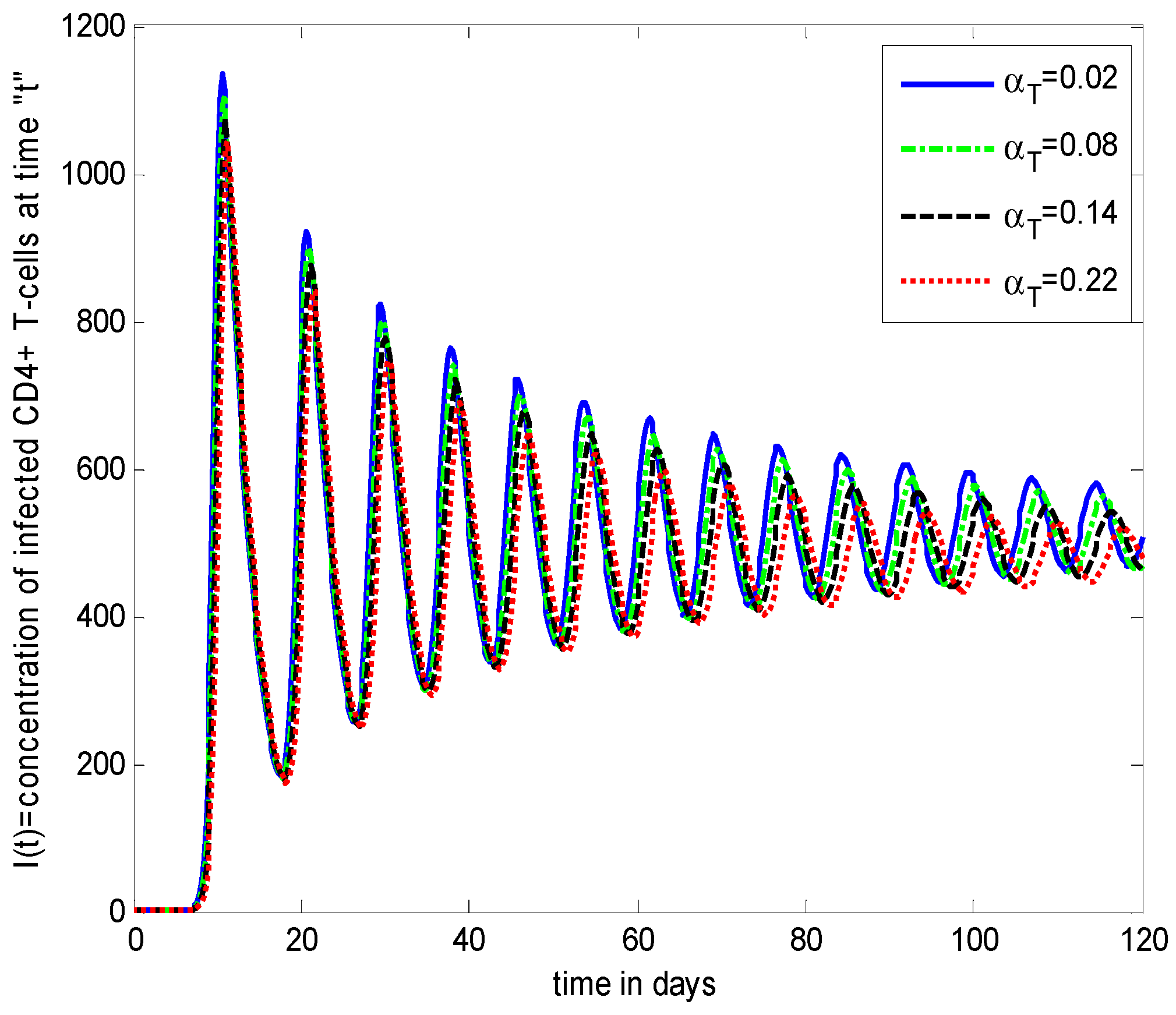
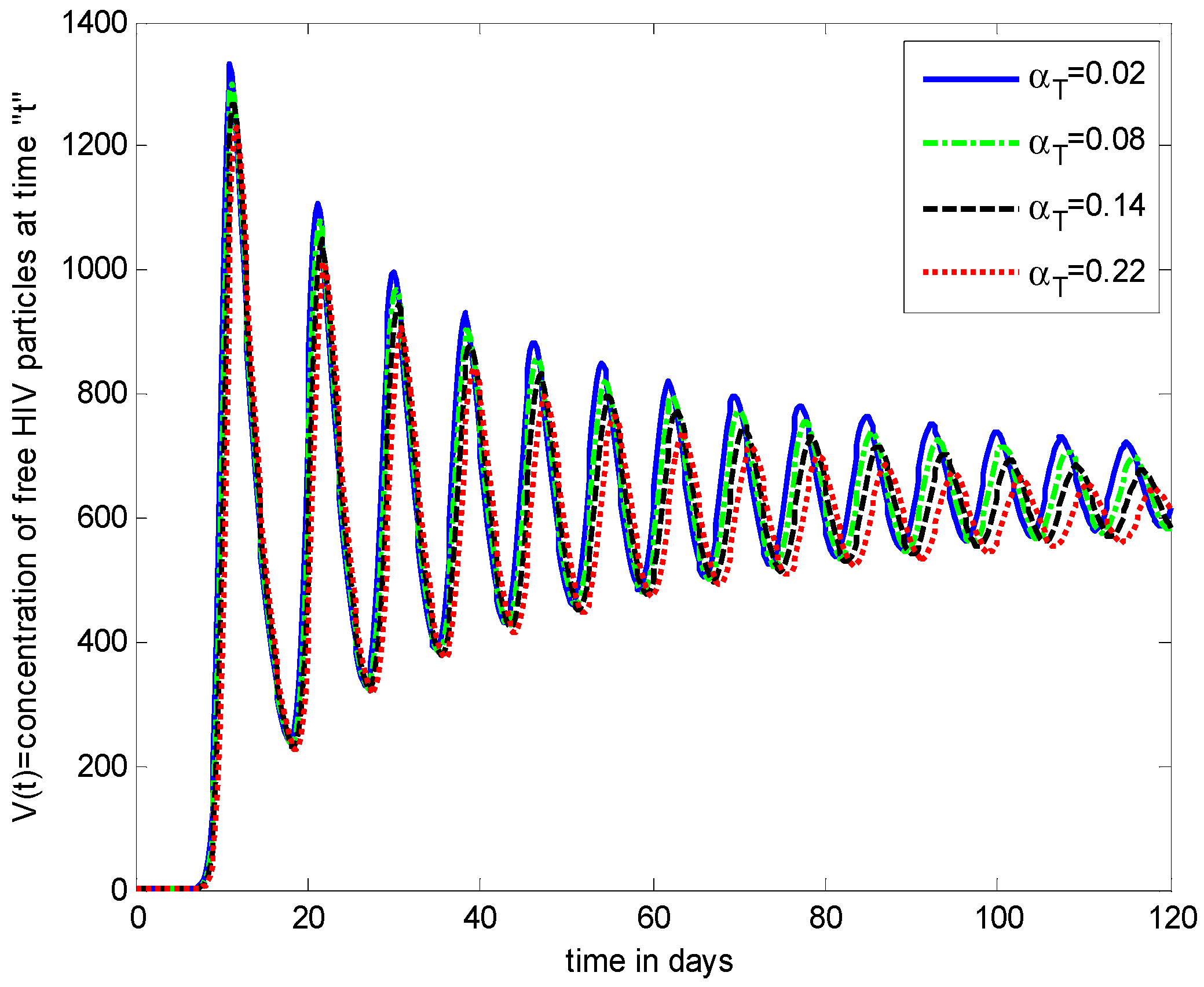
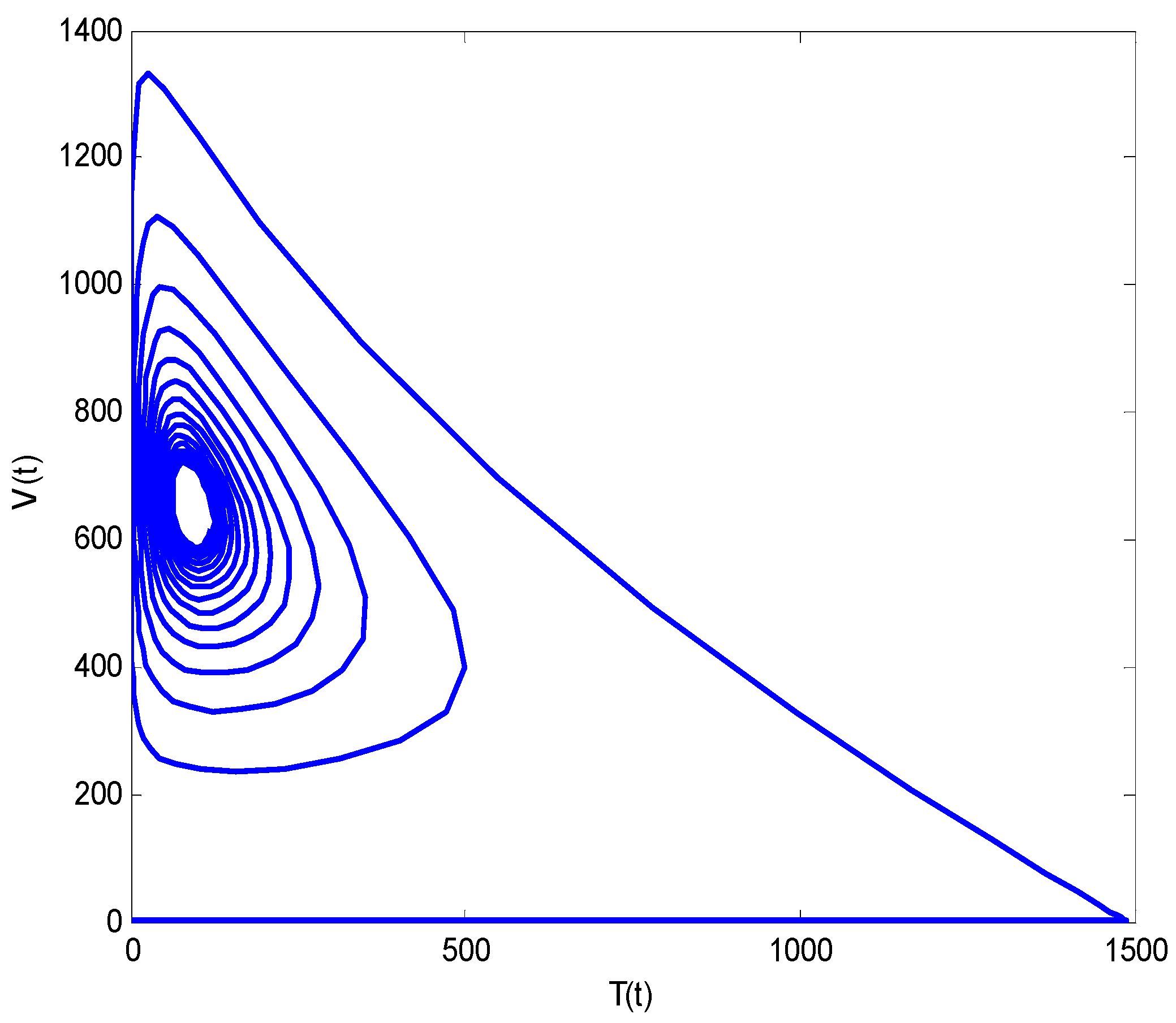
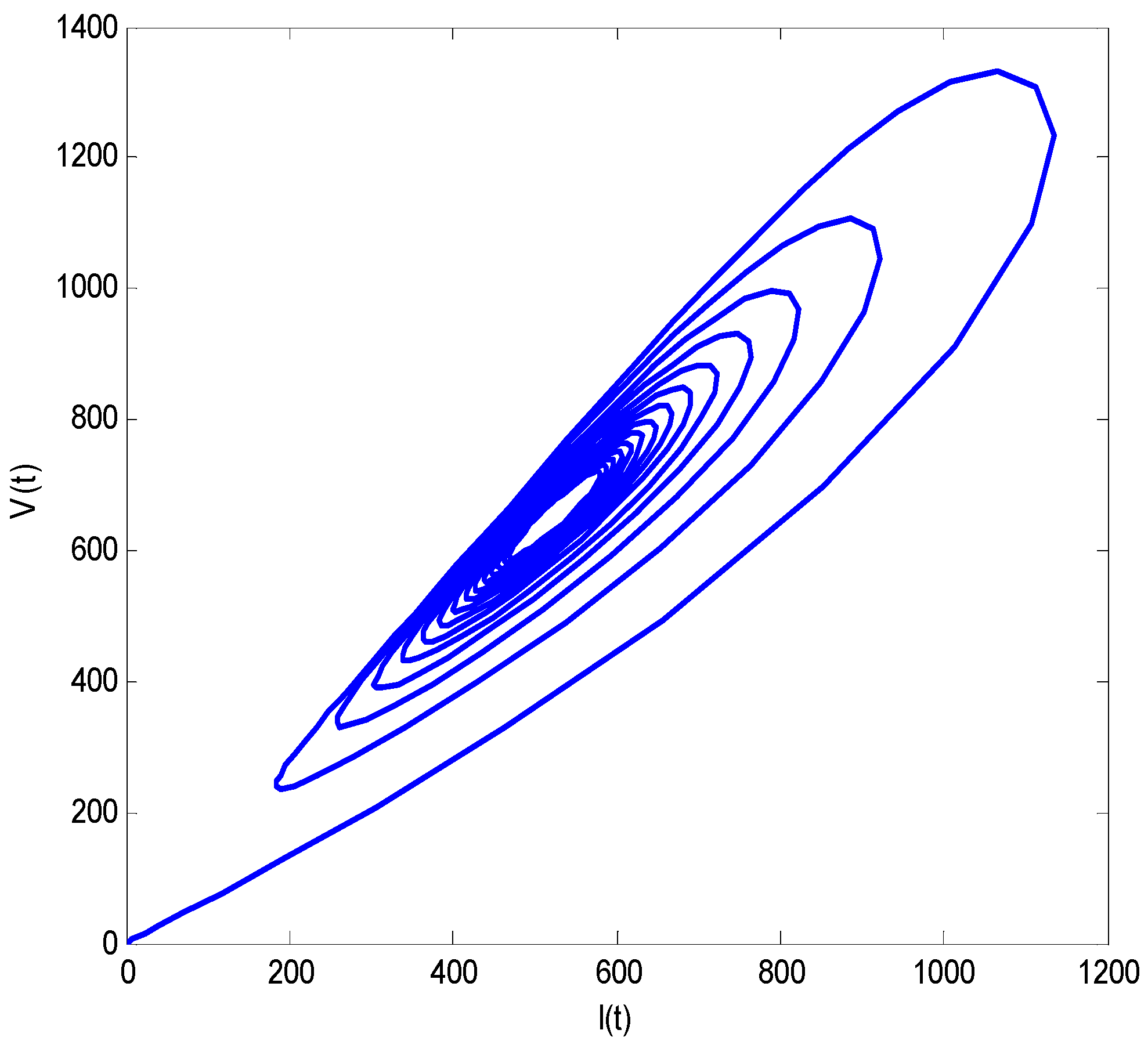
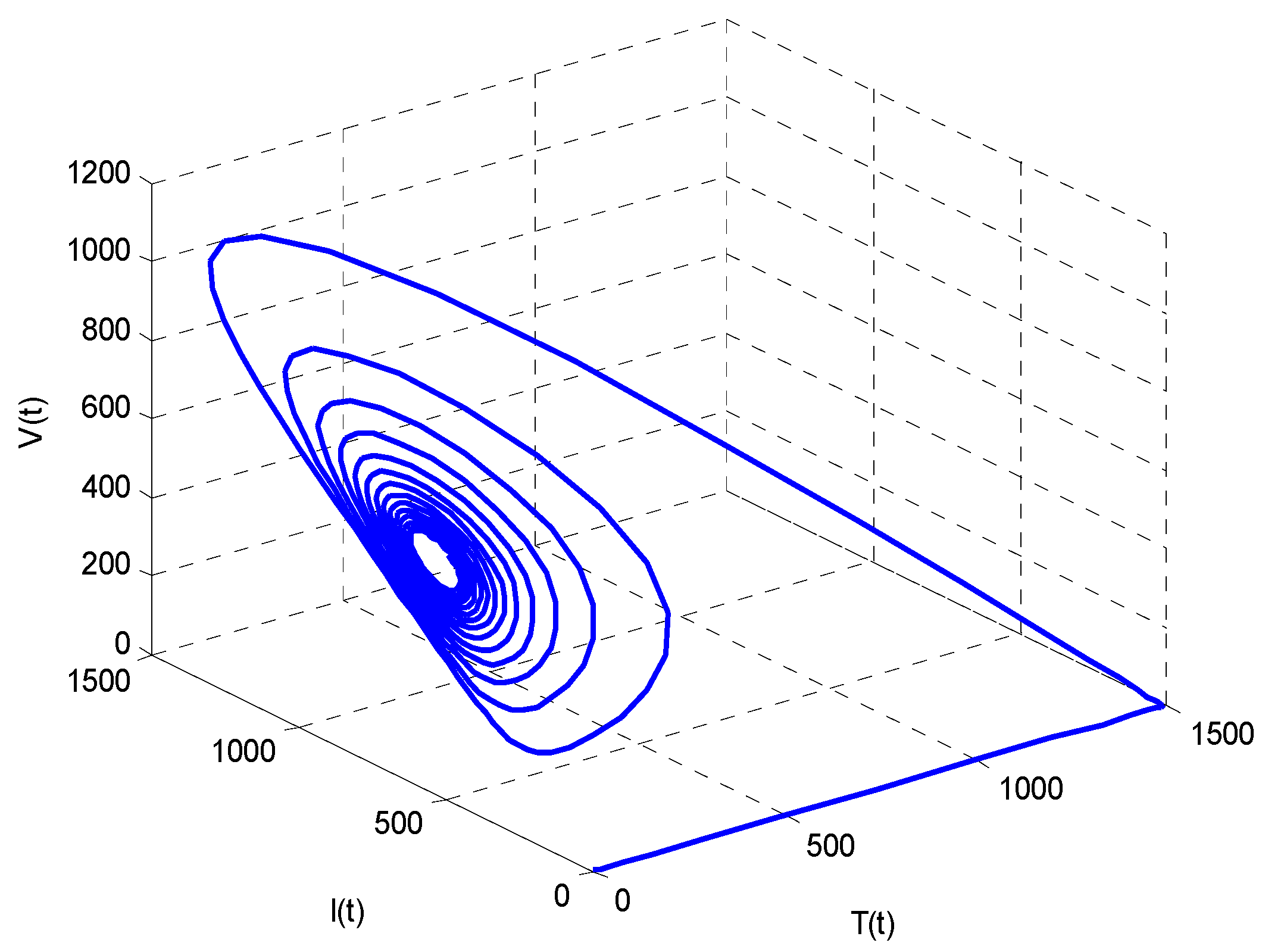
- Increasing growth rate of healthy cells, (), shows a decreasing effect in the population dynamics of healthy cells, while showing an increasing effect in the population dynamics of infected cells and HIV particles. All the profiles showed a decaying oscillatory behavior.
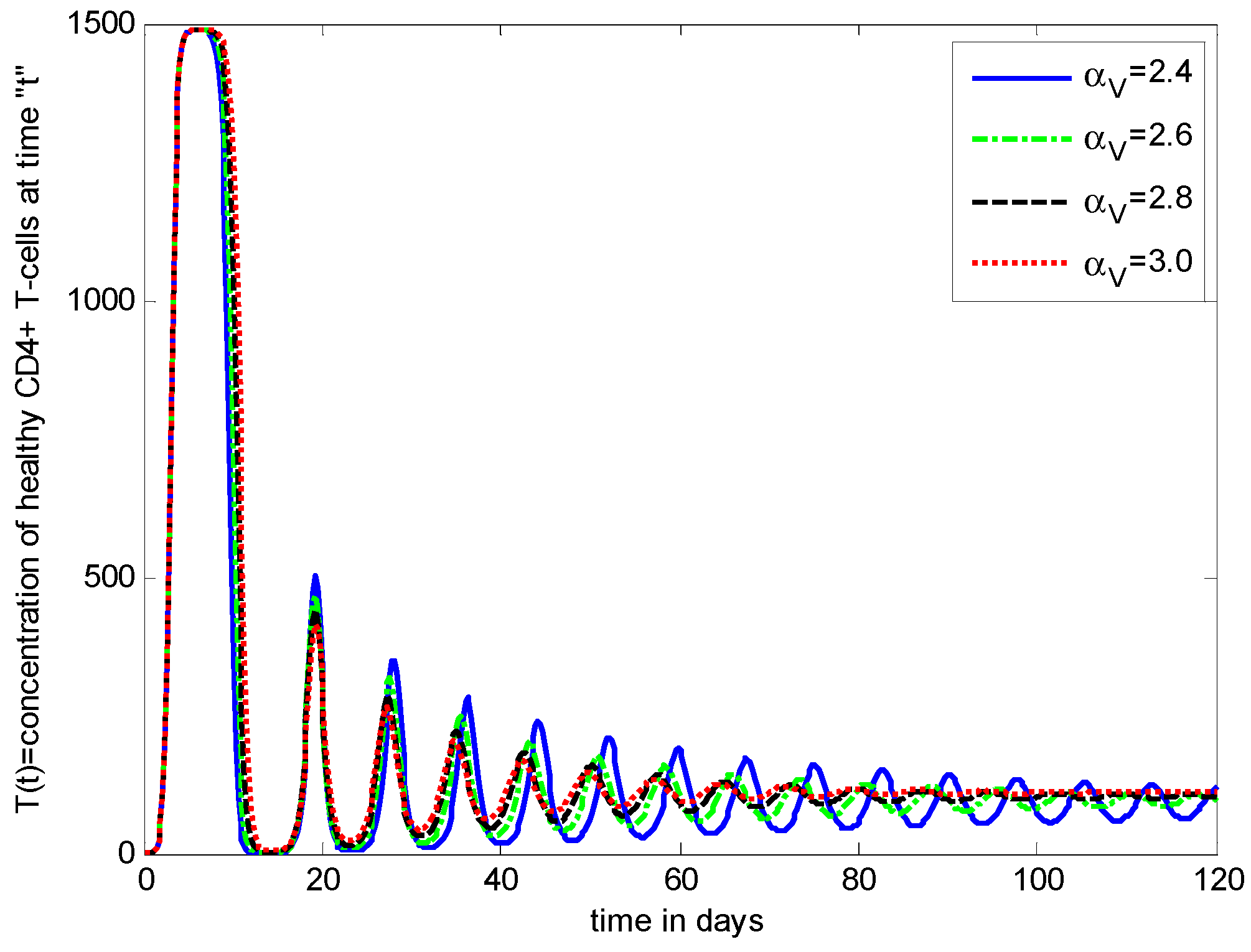
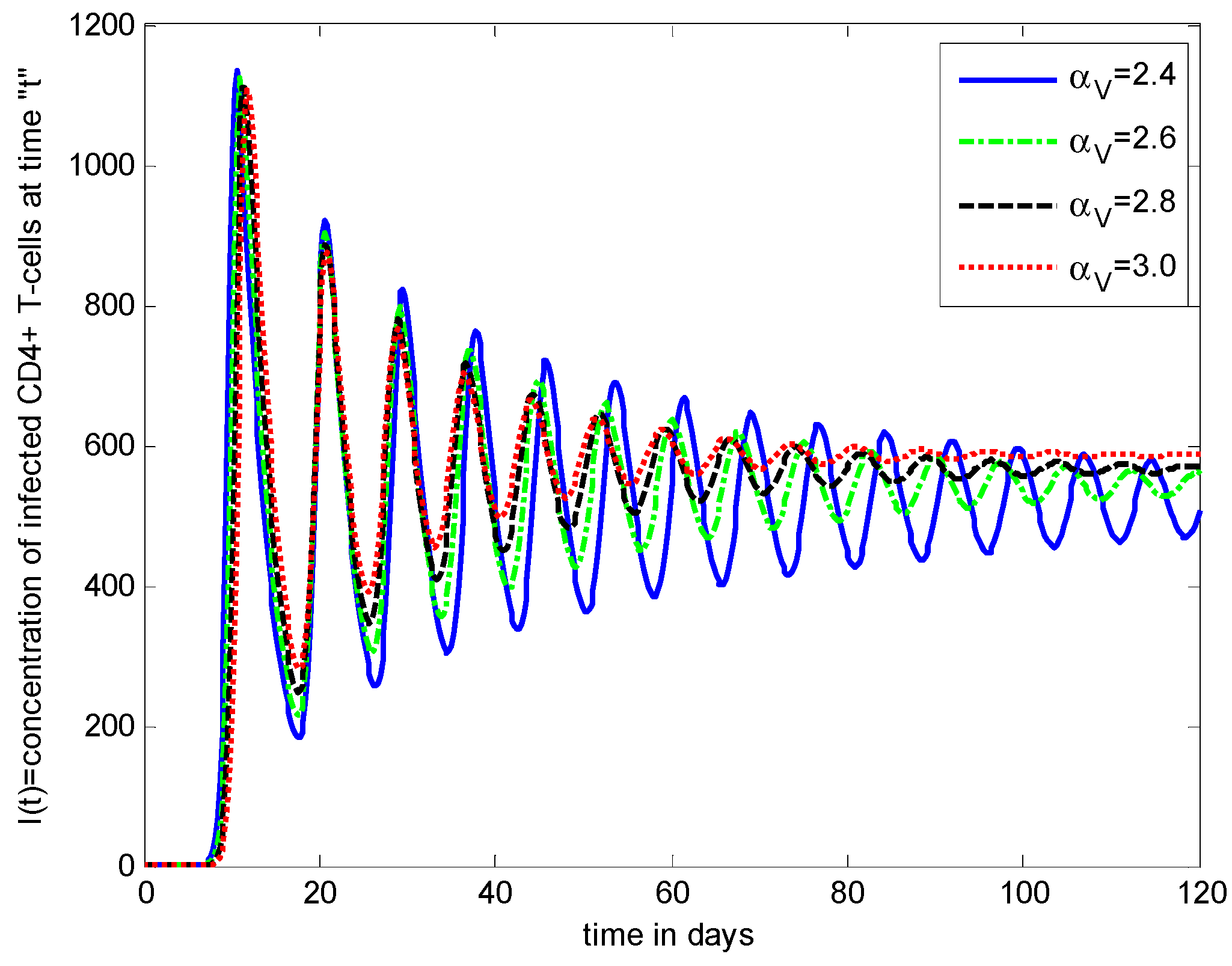
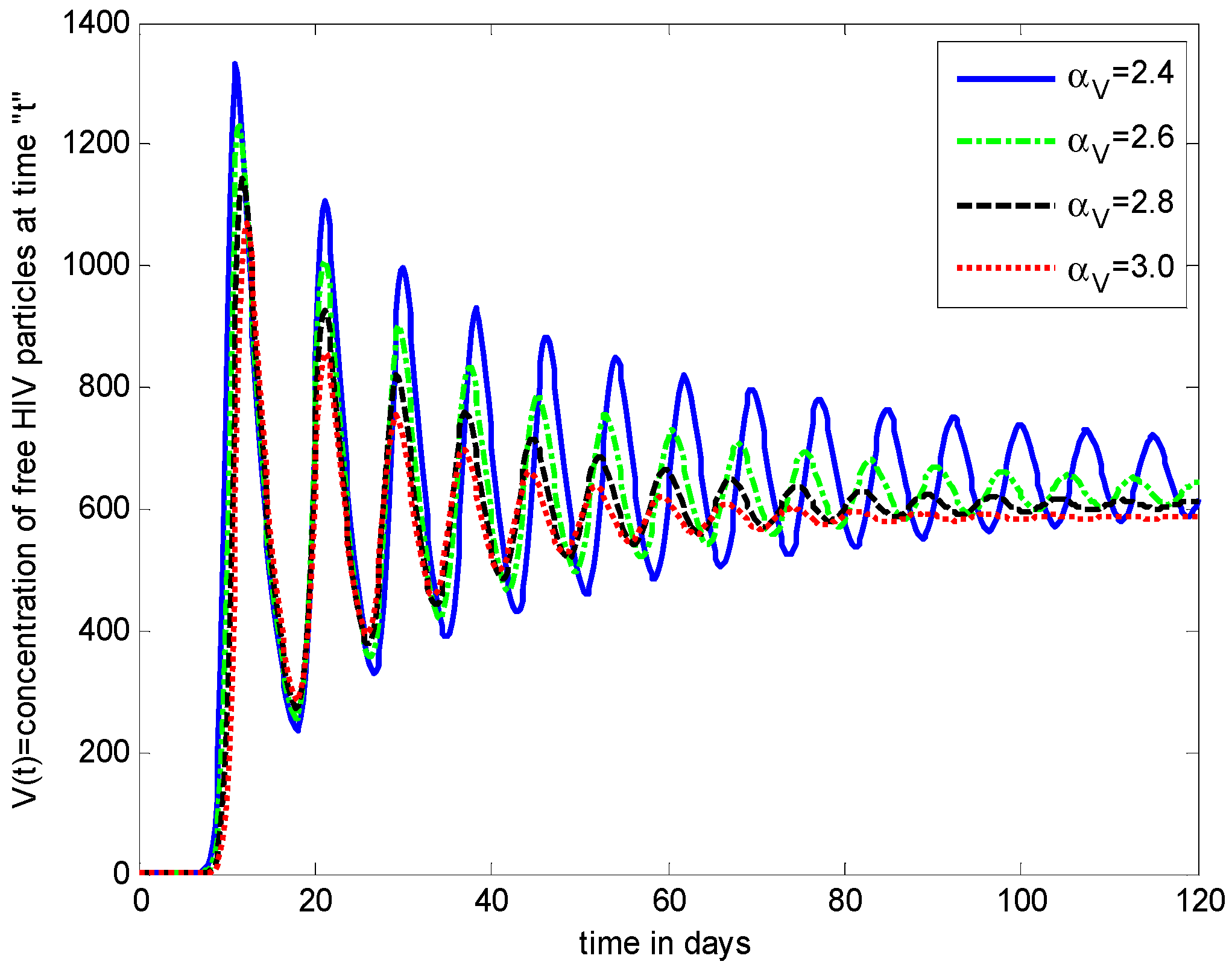
- The healthy cells and infected cells show an increasing effect, while free virus distribution shows a decreasing behavior with an increase in the values of the virus death rate ().
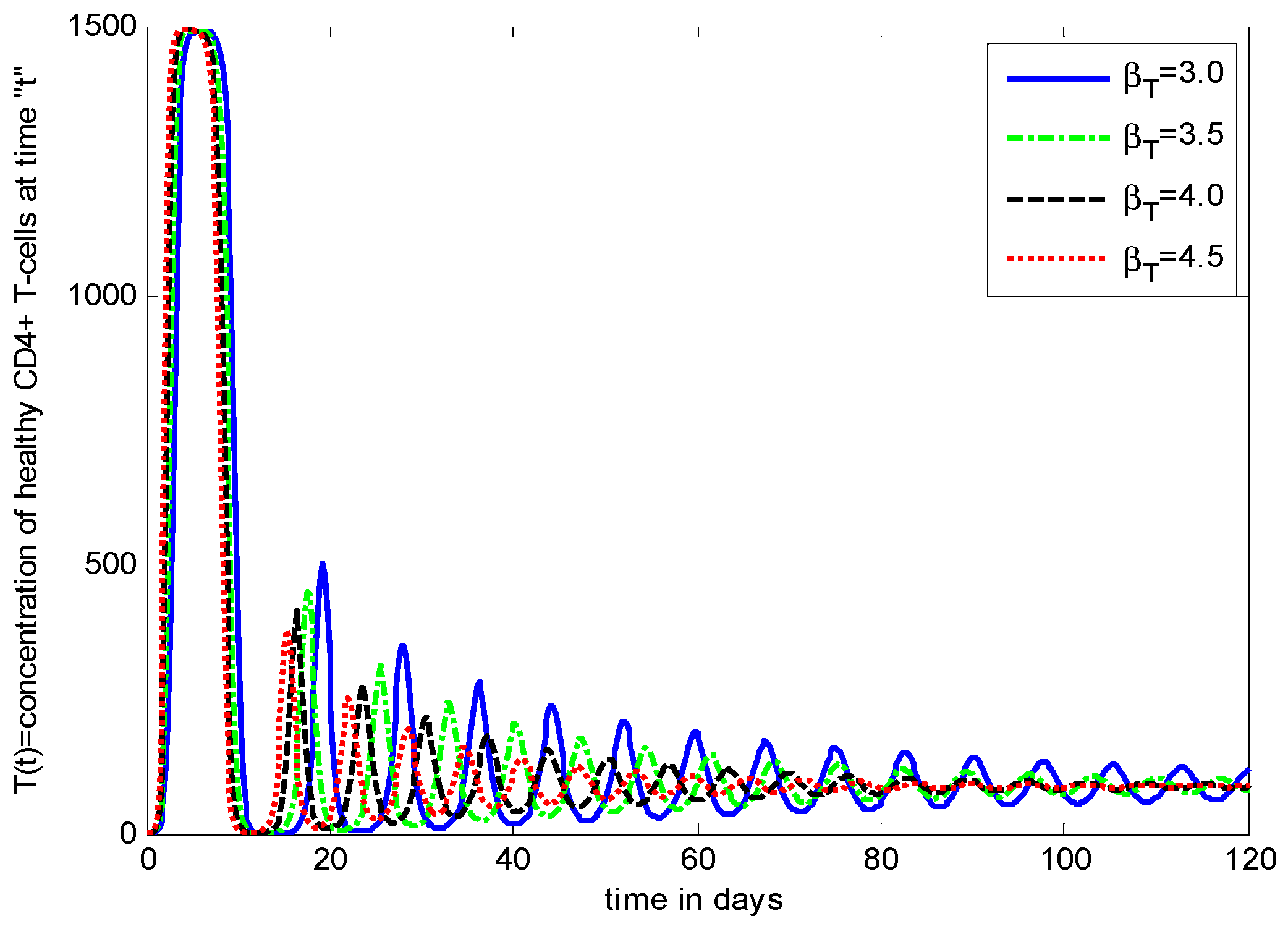
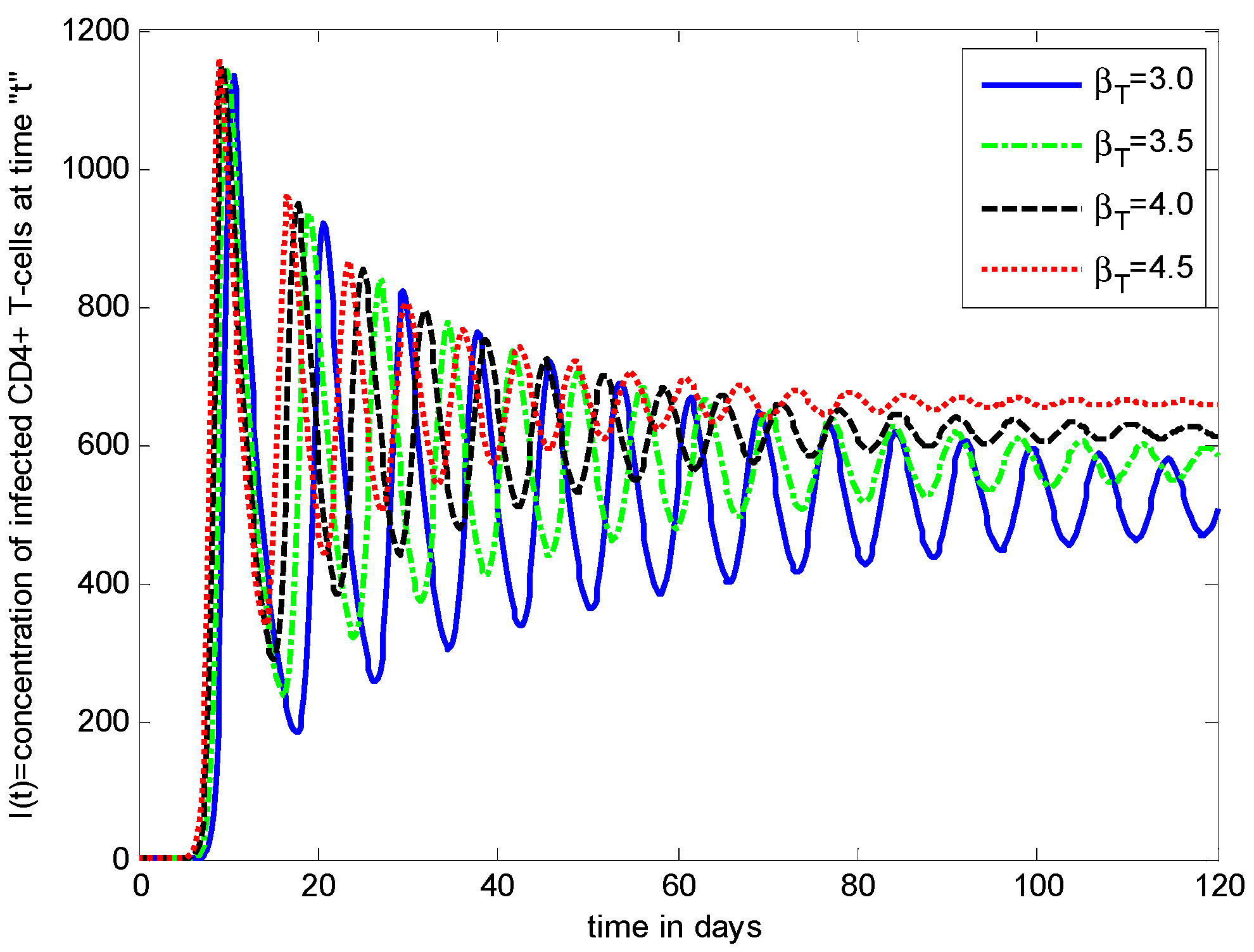
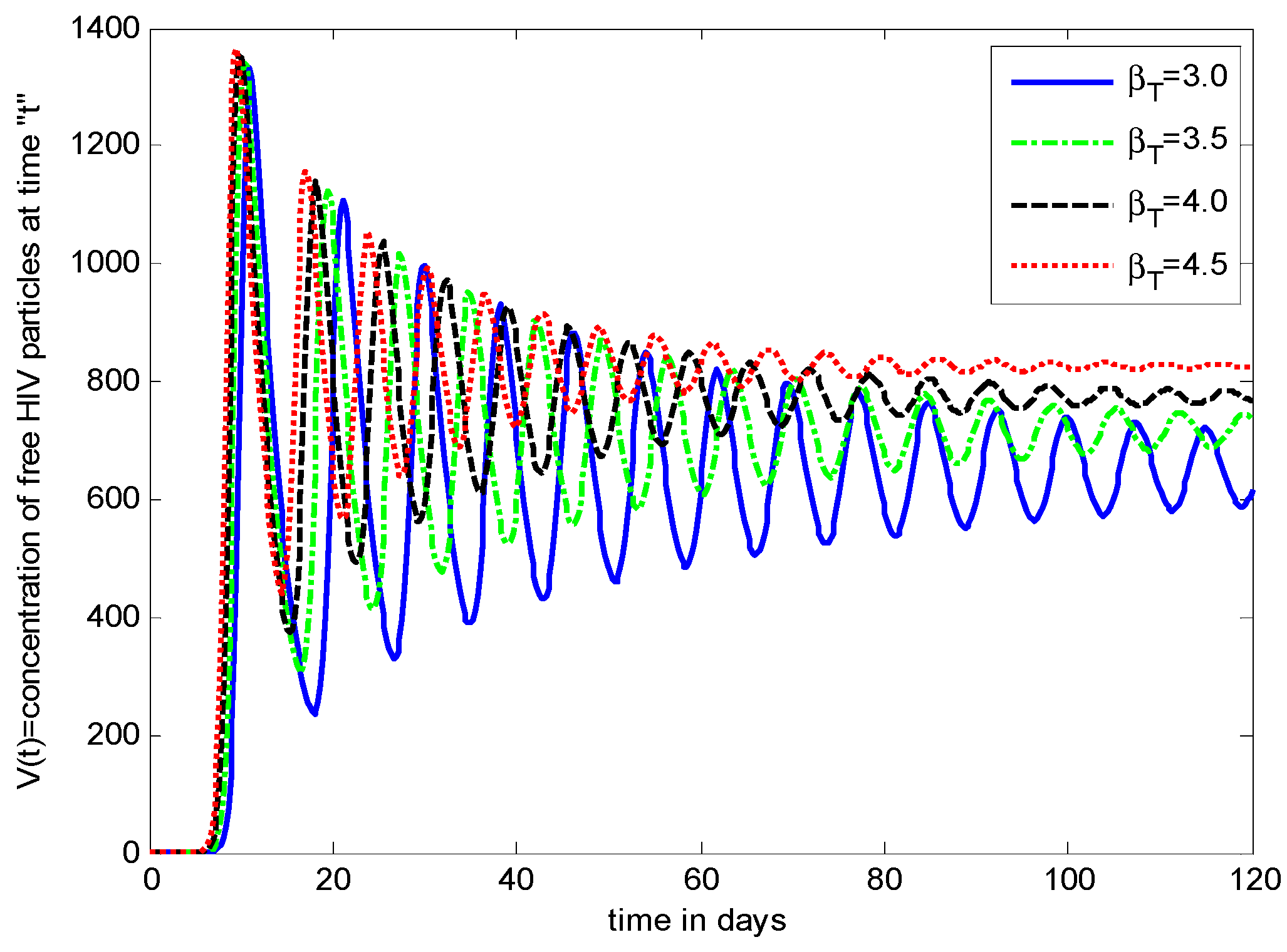
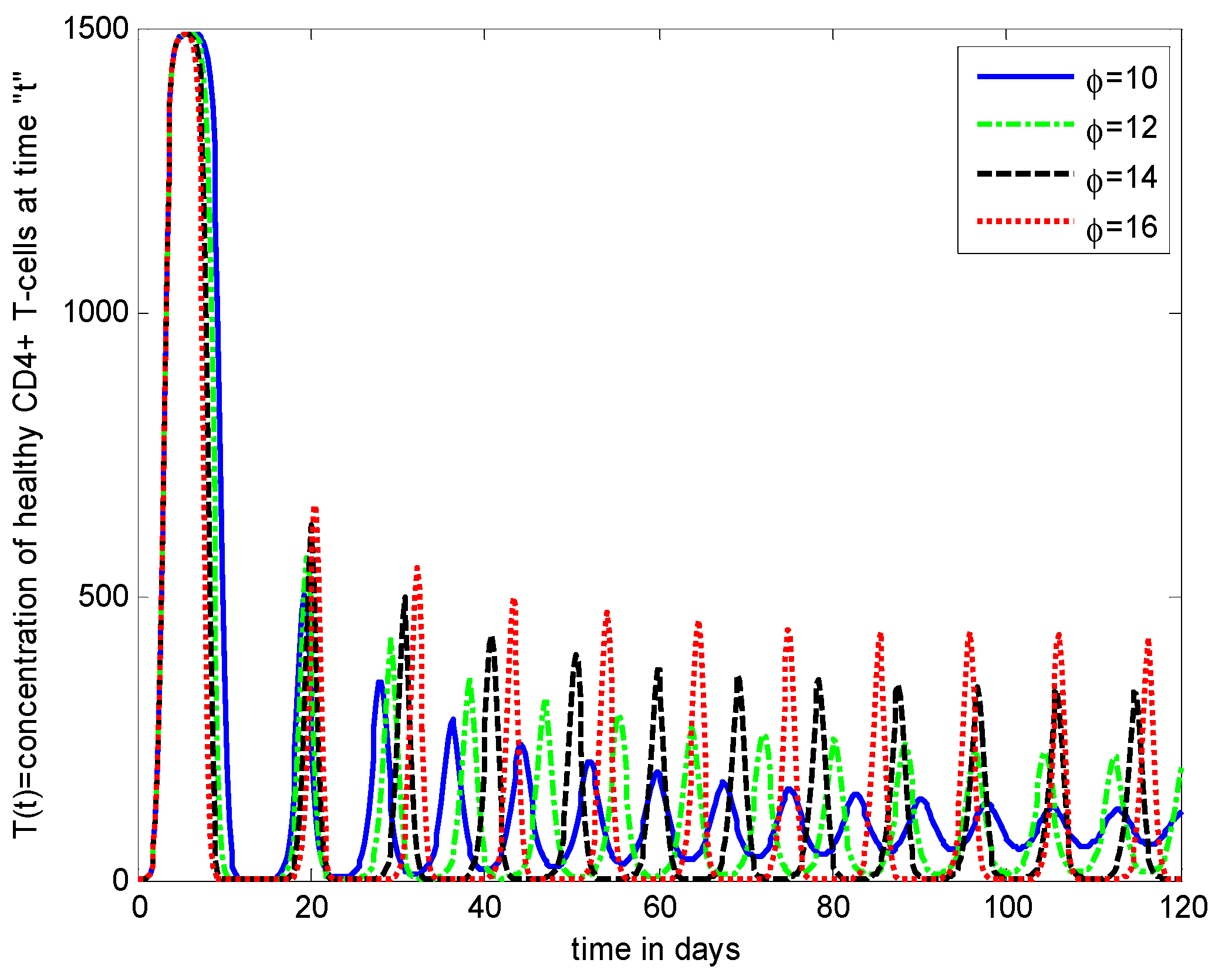
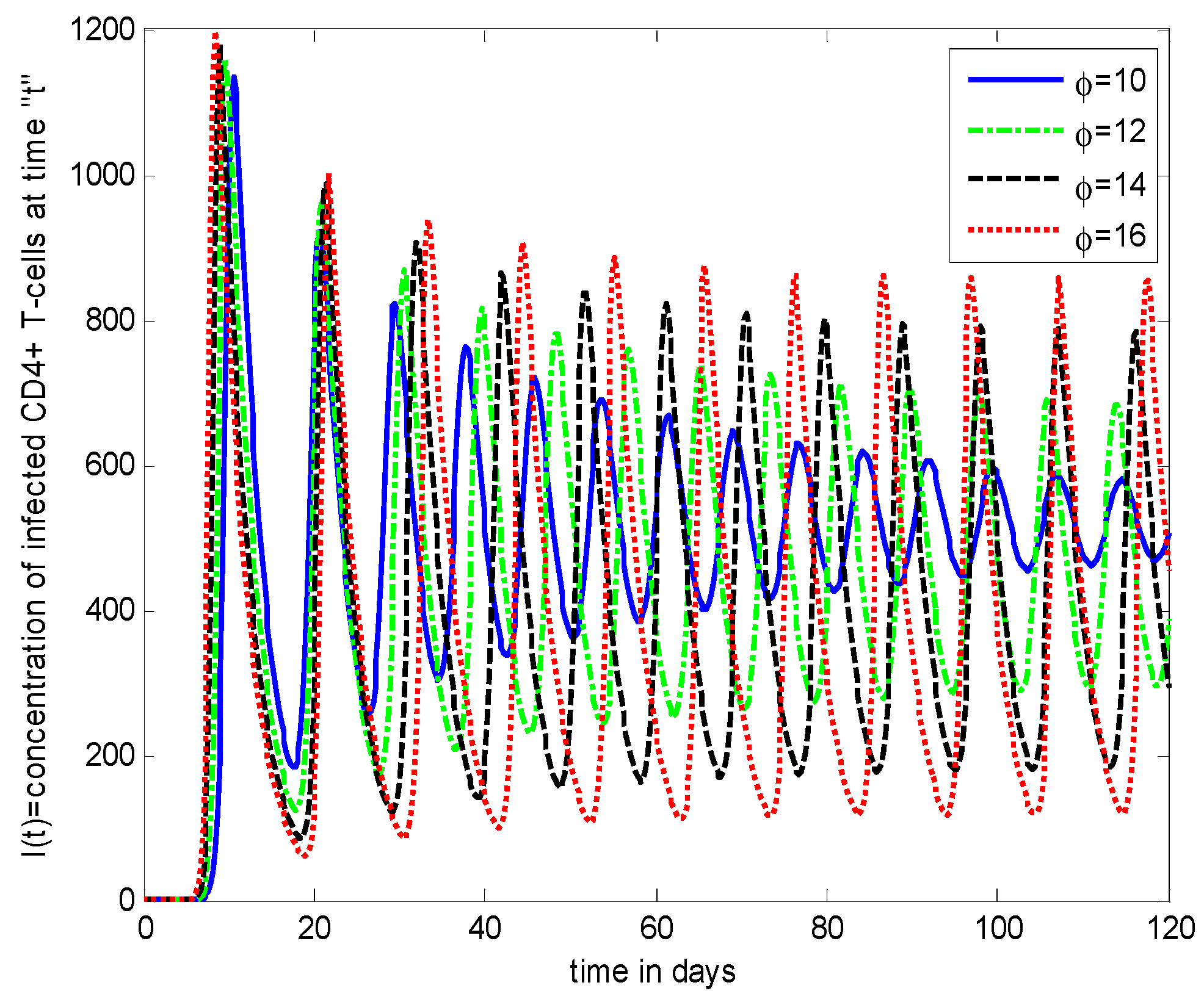
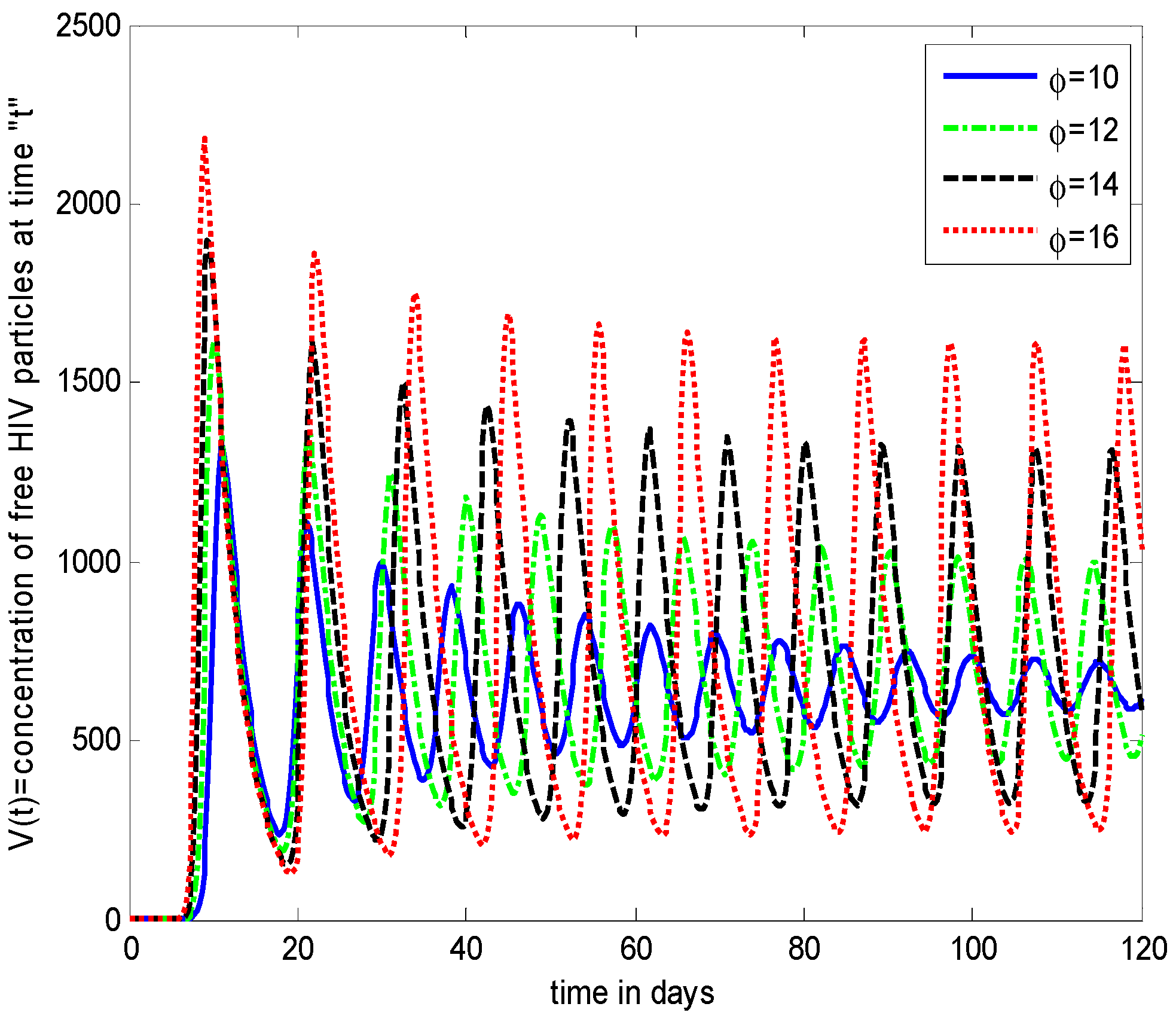
- It is noticed that the virus particles released by infected cells () show significant variations in the population distributions of healthy cells, infected cells, and the virus. By increasing the value of “”, the healthy cells, infected cells, and the virus increases.
- The graphical trends illustrate increased decay in distributions of all dependent variables with an increase in the death rate of infected cells ().
- The decrease in the density of healthy cells, infected cells, and free HIV particles is observed by increasing “”.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ding, Y.; Ye, H. A Fractional Order Differential Equation Model of HIV Infection of CD4+ T-cells. Math. Comput. Model. 2009, 50, 386–392. [Google Scholar] [CrossRef]
- Burton, D.R.; Mascola, J.R. Anti Body Responses to Envelope Glycoprotein’s in HIV-1 Infection. J. Nat. Immunol. 2015, 16, 571–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samanta, G.P. Permanence and Extinction of a Non Autonomous HIV/AIDS Epidemic Model with Distributed Time Delay. J. Non. Linea. Anal. Real World Appl. 2011, 12, 1163–1177. [Google Scholar] [CrossRef]
- Kirschner, D.; Lenhart, S.; Serbin, S. Optimal Control of the Chemotherapy of HIV. J. Math. Biol. 1997, 35, 775–792. [Google Scholar] [CrossRef] [Green Version]
- Chun, H.M.; Carpanter, R.J.; Macalino, G.E.; Cianflono, N.C. The Rloe of Sexually Transmitted Infections in HIV-1 Progression. J. Sex. Trans. Dis. 2013, 2013, 15. [Google Scholar]
- Sum, Q.; Min, L. Dynamic Analysis and Simulation of a Modified HIV Infection Model with a Saturated Infection Rate. J. Com. Math. Meth. 2014, 2014, 14. [Google Scholar]
- Arafa, A.A.M.; Rida, S.Z.; Khalil, M. A Fractional Order Model of HIV Dynamics of HIV Infection with Drug Therapy Effect. J. Non. Linea. Biol. Phys. 2014, 22, 538–543. [Google Scholar]
- Liu, X.; Wang, H.; Hu, Z.; Ma, W. Global Stability of an HIV Pathogenesis Model with Care Rate. J. Non. Linea. Anal. 2011, 12, 2947–2961. [Google Scholar]
- Osman, Z.; Abdurahman, X. Stability Analysis of Delayed HIV/AIDS Epidemic Model with Treatment and Vertical Transmission. J. App. Math. 2015, 6, 1781–1789. [Google Scholar] [CrossRef] [Green Version]
- Tuckwell, H.C.; Wan, F.Y.M. On the Behavior of Solutions in Viral Dynamical Models. J. Biol. Syst. 2004, 73, 157–161. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Li, M.Y. Mathematical Analysis of the Global Dynamics of a Model for HIV Infection of CD4+ T-cells. J. Math. Biosci. 2006, 200, 44–57. [Google Scholar] [CrossRef]
- Srivastava, P.K.; Banerjee, M.; Chandra, P. Modeling the Drug Therapy for HIV Infection. J. Bio. Syst. 2009, 17, 213–223. [Google Scholar] [CrossRef]
- Culshaw, R.V.; Ruan, S. A Delay-Differential Equation Model of HIV Infection of CD4+ T-cells. J. Math. Biosci. 2000, 165, 27–39. [Google Scholar] [CrossRef]
- World Health Organization. Global Health Observatory (GHO) Data. Available online: http://www.who.int/gho/tb/en (accessed on 1 May 2015).
- Nelson, P.W.; Perelson, A.S. Mathematical Analysis of Delay Differential Equation Models of HIV-1 Infection. Math. Biosci. 2002, 179, 73–94. [Google Scholar] [CrossRef]
- Perelson, A.S.; Kirschner, D.E.; de Boer, R. Dynamics of HIV Infection of CD4+ T-cells. Math. Biosci. 1993, 114, 81–125. [Google Scholar] [CrossRef] [Green Version]
- Ronga, L.; Fenga, Z.; Perelson, A. Emergence of HIV-1 Drug Resistance During Anti Retroviral Treatment. Bull. Math. Biol. 2007, 69, 2027–2060. [Google Scholar] [CrossRef] [PubMed]
- Duffin, R.P.; Tullis, R.H. Mathematical Models of the Complete Course of HIV Infection and AIDS. J. Theo. Med. 2002, 4, 215–221. [Google Scholar] [CrossRef] [Green Version]
- Song, X.; Cheng, S. A Delay-Differential Equation Model of HIV Infection of CD4+ T-cells. J. Korean Math. Soc. 2005, 42, 1071–1086. [Google Scholar] [CrossRef] [Green Version]
- Mechee, M.S.; Haitham, N. Application of Lie Symmetry for Mathematical Model of HIV Infection of CD4+ T-cells. J. Appl. Eng. Res. 2018, 13, 5069–5074. [Google Scholar]
- Zhou, X.; Song, X.; Shi, X. A Differential Equation Model of HIV Infection of CD4+ T-cells with Cure Rate. J. Math. Anal. Appl. 2008, 342, 1342–1355. [Google Scholar] [CrossRef] [Green Version]
- Leenheer, P.D.; Smith, H.L. Virus Dynamics: A Global Analysis. J. Appl. Math. 2003, 4, 1313–1327. [Google Scholar]
- Srivastava, P.K.; Chandra, P. Modeling the Dynamics of HIVand CD4+ T-cells during Primary Infection. J. Nonlinear Anal. 2010, 11, 612–618. [Google Scholar] [CrossRef]
- Liu, H.; Li, L. A Class Age-Structured HIV/AIDS Model with Impulsive Drug Treatment Strategy. J. Disc. Dyna. Nat. Soc. 2010, 2010, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Ho, D.D.; Neumann, A.U.; Perelson, A.S.; Chen, W.; Leonard, J.M.; Markowitz, M. Rapid Turnover of Plasma Virion and CD4 Lymphocytes in HIV-1 Infection. Nature 1995, 373, 123–126. [Google Scholar] [CrossRef] [PubMed]
- Perelson, A.S.; Essunger, P.; Cao, Y.; Vesanen, M.; Hurely, A.; Saksela, K.; Makowiz, M. Decay Characteristics of HIV-1 Infected Compartments During Combination Therapy. Nature 1997, 387, 188–191. [Google Scholar] [CrossRef]
- Schieweck, F. A Stable Discontinuous Galerkin-Petrov Time Discretization of Higher Order. J. Numer. Math. 2010, 18, 25–57. [Google Scholar] [CrossRef]
- Kuang, Y. Delay Differential Equation with Applications in Population Dynamics; Academic Press: London, UK, 2004. [Google Scholar]
- Ongun, M.Y. The Laplace Adomian Decomposition Method for Solving a Model for HIV Infection of CD4+ T-cells. Math. Comput. Model. 2011, 53, 597–603. [Google Scholar] [CrossRef]
- Yuzbasi, S. A Numerical Approach to Solve the Model for HIV Infection of CD4 T-cell. J. Appl. Math. Mod. 2012, 36, 5876–5890. [Google Scholar] [CrossRef]
- Khalid, M.; Sultana, M.; Zaidi, M.; Fareeha, S.K. A Numerical Solution of a Model for HIV Infection of CD4 T-Cells. J. Inno. Sci. Res. 2015, 16, 79–85. [Google Scholar]
- Merdan, M.; Gokdogan, A.; Yildirim, A. On the Numerical Solution of the Model for HIV Infection of CD4 T-Cells. J. Comput. Math. Appl. 2011, 62, 118–123. [Google Scholar] [CrossRef] [Green Version]
- Attaullah, A.S.; Yassen, M.F. A study on the transmission and dynamical behavior of an HIV/AIDS epidemic model with a cure rate. AIMS Math. 2022, 7, 17507–17528. [Google Scholar] [CrossRef]
- Ogunlaran, O.M.; Noutchie, S.C.O. Mathematical Model for an Effective Management of HIV Infection. J. Biomed. Res. Int. 2016, 2016, 4217548. [Google Scholar] [CrossRef] [PubMed]
- Boukari, B.E.; Hattaf, K.; Yousfi, N. A Discrete Model for HIV Infection with Distributed Delay. J. Diff. Equa. 2014, 2014, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Xiao, Y. Global Dynamics of a Virus Immune System with Virus Guided Therapy and Saturation Growth of Virus. J. Math. Probl. Eng. 2018, 2018, 1–18. [Google Scholar] [CrossRef]
- Espindola, M.S.; Lima, L.J.G.; Soares, L.S.; Zambuzi, F.A.; Cacemiro, M.; Fontanari, C.; Bollela, V.R.; Frantz, F.G. Classical and Alternative Macrophages have Impaired Function during Acute and Chronic HIV-1 Infection. J. Braz. Infect. Dis. 2017, 21, 42–50. [Google Scholar]
- Kinner, S.A.; Snow, K.; Wirtz, A.L.; Altice, F.L.; Beyrer, C.; Dolan, K. Age-Specific Global Prevalence of Hepatitis B, Hepatitis C, HIV and Tuberculosis Among Incarcerated People: A Systematic Review. J. Adolesc. Health 2018, 62, 18–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angulo, J.M.C.; Cuesta, T.A.C.; Menezes, E.P.; Pedroso, C.; Brites, C. A Systematic Review on the Influence of HLA-B Polymorphisms on HIV-1 Mother to Child Transmission. J. Braz. Infect. Dis. 2019, 23, 53–59. [Google Scholar] [CrossRef]
- Theys, K.; Libin, P.; Pena, A.C.P.; Nowe, A.; Vandamme, A.M.; Abecasis, A.B. The Impact of HIV-1 within Host Evolution on Transmission Dynamics. J. Curr. Opin. Viro. 2018, 28, 92–101. [Google Scholar] [CrossRef]
- Hallberg, D.; Kimario, T.D.; Mtuya, C.; Msuya, M.; Bjorling, G. Factors Affecting HIV Disclosure among Partners in Morongo, Tanzania. J. Inter. J. Afri. Nurs. Sci. 2019, 10, 49–54. [Google Scholar] [CrossRef]
- Ransome, Y.; Thurber, K.A.; Swen, M.; Crawford, N.D.; Germane, D.; Dean, L.T. Social Capital and HIV/AIDS in the United States: Knowledge, Gaps and Future Directions. J. SSM. Popu. Health 2018, 5, 73–85. [Google Scholar] [CrossRef]
- Naidoo, K.; Gengiah, S.; Singh, S.; Stillo, J.; Padayatchi, N. Quality of TB Care among People Living with HIV: Gaps and Solutions. J. Clin. Tube. Myco. Dis. 2019, 17, 100–122. [Google Scholar] [CrossRef]
- Omondi, E.O.; Mbogo, W.R.; Luboobi, L.S. A Mathematical Modeling Study of HIV Infection in two Heterosexual Age Groups in Kenya. J. Infect. Dis. Model. 2019, 4, 83–98. [Google Scholar]
- Duro, R.; Pereira, N.R.; Figueiredo, C.; Pineiro, C.; Caldas, C.; Serrao, R.; Saemento, A. Routine CD4 Monitoring in HIV Patients with Viral Suppression: Is it Really Necessary? A Portuguese Cohort. J. Microbio. Immun. Infect. 2018, 51, 593–597. [Google Scholar] [CrossRef] [PubMed]
- Mbogo, W.R.; Luboobi, L.S.; Odhiambo, J.W. Stochastic Model for In-Host HIV Dynamics with Therapeutic Intervention. Int. Sch. Res. Not. 2013, 2013, 103708. [Google Scholar] [CrossRef]
- Ghoreishi, M.; Ismail, A.I.B.; Alemari, A.K. Application of the Hemotopy Analysis Method for Solving a Model for HIV Infection of CD4+ T-cells. J. Math. Comput. Model. 2011, 54, 3007–3015. [Google Scholar] [CrossRef]
- Elaiw, A.M. Global Dynamics of an HIV Infection Model with two Classes of Target Cells and Distributed Delayes. J. Discret. Dyn. Nat. Soc. 2012, 2012, 13. [Google Scholar]
- Ali, N.; Ahmad, S.; Aziz, S.; Zaman, G. The Adomian Decomposition Method for Solving HIV Infection Model of Latently Infected Cells. J. MSMK 2019, 3, 5–8. [Google Scholar] [CrossRef]
- Yüzbaşı, Ş.; Karaçayır, M. An exponential Galerkin method for solutions of HIV infection model of CD4+ T-cells. Comput. Biol. Chem. 2017, 67, 205–212. [Google Scholar] [CrossRef]
- Kirschner, D. Using Mathematics to Understand HIV Immune Dynamics. J. Math. Biosci. 1996, 43, 191–202. [Google Scholar]
- Webb, G.F.; Kirschner, D. A Model for HIV Treatment Strategy in the Chemotherapy of AIDS. J. Math. Biol. 1996, 58, 367–390. [Google Scholar]
- Attaullah; Sohaib, M. Mathematical modeling and numerical simulation of HIV infection model. Results Appl. Math. 2020, 7, 100118. [Google Scholar] [CrossRef]
- Kutta, W. Beitrag zur naerungsweisen integration totaler differentialgleichungen. Z. Math. Phy. 1901, 46, 435–453. [Google Scholar]
- Butcher, J.C. Numerical Methods for Ordinary Differential Equations; Wiley: Hoboken, NJ, USA, 2008. [Google Scholar]
- Jiwari, R. Local radial basis function-finite difference based algorithms for singularly perturbed Burgers’ model. Math. Comput. Simul. 2022, 198, 106–126. [Google Scholar] [CrossRef]
- Mittal, R.C.; Kumar, S.; Jiwari, R. A cubic B-spline quasi-interpolation algorithm to capture the pattern formation of coupled reaction-diffusion models. Eng. Comput. 2022, 38, 1375–1391. [Google Scholar] [CrossRef]
- Pandit, S. Local radial basis functions and scale-3 Haar wavelets operational matrices based numerical algorithms for generalized regularized long wave model. Wave Motion 2022, 109, 102846. [Google Scholar] [CrossRef]
- Mittal, R.C.; Pandit, S. New scale-3 haar wavelets algorithm for numerical simulation of second order ordinary differential equations. Proc. Natl. Acad. Sci. India Sect. A Phys. Sci. 2019, 89, 799–808. [Google Scholar] [CrossRef]























{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Description | Values |
|---|---|---|
| Concentration of healthy cells | ||
| Population of infected cells | ||
| Dynamics of free viruses | ||
| Supply rate of healthy cells | ||
| Natural death rate for healthy cells | ||
| Maximum density of healthy cells population | ||
| Infection rate of healthy cells | ||
| Virus particles released by infected cells | ||
| Virus death rate | ||
| Death rate of infected cells | ||
| Growth rate of healthy cells |
| t | Runge–Kutta | LADM-Pade [29] | Bessel Coll. N = 8 [30] | PIA(1,1) [31] | MVIM [32] |
|---|---|---|---|---|---|
| 0.2 | 0.2088006789 | 0.2088072731 | 0.2038616561 | 0.2087295073 | 0.2088080868 |
| 0.4 | 0.4062136749 | 0.4061052625 | 0.3803309335 | 0.4059404993 | 0.4062407949 |
| 0.6 | 0.7643508145 | 0.7611467713 | 0.6954623767 | 0.7635790156 | 0.7644287245 |
| 0.8 | 1.4138702489 | 1.3773198590 | 1.2759624442 | 1.4119543417 | 1.4140941730 |
| 1.0 | 2.5911951903 | 2.3291697610 | 2.3832277428 | 2.5867690583 | 2.5919210760 |
| t | DTM N = 6 [33] | EGM N = 3 [50] | EGM N = 4 [50] | EGM N = 5 [50] | cGP(2)-Method |
| 0.2 | 0.2116480000 | 0.2722229510 | 0.2345157340 | 0.1982953765 | 0.2088064964 |
| 0.4 | 0.4226850000 | 0.3065308713 | 0.4201803666 | 0.4183153468 | 0.4062347843 |
| 0.6 | 0.8179400000 | 0.7075440591 | 0.7255920466 | 0.7603331972 | 0.7644082444 |
| 0.8 | 1.5462110000 | 1.5297610198 | 1.4170402360 | 1.4077147917 | 1.4140090611 |
| 1.0 | 2.8540530000 | 2.6678673734 | 2.5916251711 | 2.5915947135 | 2.5915094589 |
| t | Runge–Kutta | LADM-Pade [29] | Bessel coll. N=8 [30] | PIA(1,1) [31] | MVIM [32] |
|---|---|---|---|---|---|
| 0.2 | 0.0000060318 | 0.0000060327 | 0.0000062478 | 0.0000060315 | 0.0000060327 |
| 0.4 | 0.0000131564 | 0.0000131591 | 0.0000129355 | 0.0000131530 | 0.0000131583 |
| 0.6 | 0.0000212206 | 0.0000212683 | 0.0000203526 | 0.0000212101 | 0.0000212233 |
| 0.8 | 0.0000301728 | 0.0000300691 | 0.0000283730 | 0.0000301480 | 0.0000301745 |
| 1.0 | 0.0000400314 | 0.0000398736 | 0.0000369084 | 0.0000399785 | 0.0000400254 |
| t | DTM N = 6 [33] | EGM N = 3 [50] | EGM N = 4 [50] | EGM N = 5 [50] | cGP(2)-Method |
| 0.2 | 0.0000063666 | 0.0000091673 | 0.0000058251 | 0.0000059641 | 0.0000060325 |
| 0.4 | 0.0000139924 | 0.0000155229 | 0.0000134051 | 0.0000131340 | 0.0000131579 |
| 0.6 | 0.0000226514 | 0.0000228459 | 0.0000213405 | 0.0000212682 | 0.0000212231 |
| 0.8 | 0.0000332836 | 0.0000318486 | 0.0000301313 | 0.0000301754 | 0.0000301764 |
| 1.0 | 0.0000485399 | 0.0000421057 | 0.0000400369 | 0.0000400377 | 0.0000400364 |
| t | Runge–Kutta | LADM-Pade [29] | Bessel Coll. N = 8 [30] | PIA(1,1) [31] | MVIM [32] |
|---|---|---|---|---|---|
| 0.2 | 0.0618808474 | 0.0618799602 | 0.0618799185 | 0.0618796999 | 0.0618799087 |
| 0.4 | 0.0382961304 | 0.0383132488 | 0.0382949349 | 0.0382939096 | 0.0382959576 |
| 0.6 | 0.0237057031 | 0.0243917434 | 0.0237043186 | 0.0237016917 | 0.0237102948 |
| 0.8 | 0.0146813143 | 0.0099672189 | 0.0146795698 | 0.0146744145 | 0.0147004190 |
| 1.0 | 0.0091015791 | 0.0033050764 | 0.0090993030 | 0.0090905052 | 0.0091572387 |
| t | DTM N = 6 [33] | EGM N = 3 [50] | EGM N = 4 [50] | EGM N = 5 [50] | cGP(2)-Method |
| 0.2 | 0.0618800000 | 0.0618823466 | 0.0618790041 | 0.0618799035 | 0.0618799805 |
| 0.4 | 0.0383090000 | 0.0383077329 | 0.0382950148 | 0.0382947890 | 0.0382950575 |
| 0.6 | 0.0239200000 | 0.0237055266 | 0.0237053683 | 0.0237046061 | 0.0237047074 |
| 0.8 | 0.0162120000 | 0.0146708169 | 0.0146798882 | 0.0146803810 | 0.0146804932 |
| 1.0 | 0.0160500000 | 0.0091056907 | 0.0091009339 | 0.0091008486 | 0.0091009447 |
| t | LADM-Pade [29] | Bessel Coll. N = 8 [30] | PIA(1,1) [31] | MVIM [32] | DTM N = 6 [33] |
|---|---|---|---|---|---|
| 0.2 | 0.000006594223280 | 0.004939022776720 | 0.000071171576720 | 7.40792327999 × 10−6 | 0.002847321123280 |
| 0.4 | 0.000108412464641 | 0.025882741464641 | 0.000273175664641 | 2.71199353589 × 10−5 | 0.016471325035359 |
| 0.6 | 0.003204043237096 | 0.068888437837096 | 0.000771798937096 | 7.79099629040 × 10−5 | 0.053589185462904 |
| 0.8 | 0.036550389916353 | 0.137907804716353 | 0.001915907216353 | 2.23924083647 × 10−3 | 0.132340751083647 |
| 1.0 | 0.262025429366243 | 0.207967447566243 | 0.004426132066243 | 7.25885633757 × 10−3 | 0.262857809633757 |
| t | EGM N = 3 [50] | EGM N = 4 [50] | EGM N = 5 [50] | cGP(2)-Method | |
| 0.2 | 0.063422272123280 | 0.025715055123280 | 0.010505302376720 | 5.81760745974 × 10−6 | |
| 0.4 | 0.099682803664641 | 0.013966691635359 | 0.012101671835359 | 2.11093478030 × 10−5 | |
| 0.6 | 0.056806755437096 | 0.038758767937096 | 0.004017617337096 | 5.74299020841 × 10−5 | |
| 0.8 | 0.115890770883647 | 0.003169987083647 | 0.006155457216353 | 1.38812210279 × 10−4 | |
| 1.0 | 0.076672183033757 | 0.000429980733757 | 0.000399523133757 | 3.14268571135 × 10−4 | |
| t | LADM-Pade [29] | Bessel Coll. N = 8 [30] | PIA(1,1) [31] | MVIM [32] | DTM N= 6 [33] |
|---|---|---|---|---|---|
| 0.2 | 8.21028439 × 10−10 | 2.15921028439 × 10−7 | 3.7897156100 × 10−10 | 8.2102843899 × 10−10 | 3.3472103 × 10−7 |
| 0.4 | 2.61393801 × 10−9 | 2.20986061988 × 10−7 | 3.4860619888 × 10−9 | 1.8139380112 × 10−9 | 8.3591393 × 10−7 |
| 0.6 | 4.76223967 × 10−8 | 8.68077603328 × 10−7 | 1.0577603329 × 10−8 | 2.6223966711 × 10−9 | 1.4307224 × 10−6 |
| 0.8 | 1.03710297 × 10−7 | 1.79981029736 × 10−6 | 2.4810297362 × 10−8 | 1.6897026384 × 10−9 | 3.1107898 × 10−6 |
| 1.0 | 1.57815845 × 10−7 | 3.12301584505 × 10−6 | 5.2915845057 × 10−8 | 6.0158450573 × 10−9 | 8.5084842 × 10−6 |
| t | EGM N = 3 [50] | EGM N = 4 [50] | EGM N = 5 [50] | cGP(2)-Method | |
| 0.2 | 3.1354210284 × 10−6 | 2.0677897156 × 10−7 | 6.77789715609 × 10−8 | 6.6752196826462 × 10−10 | |
| 0.4 | 2.3664139380 × 10−6 | 2.4861393801 × 10−7 | 2.24860619888 × 10−8 | 1.4924805624754 × 10−9 | |
| 0.6 | 1.6252223967 × 10−6 | 1.1982239667 × 10−7 | 4.75223966710 × 10−8 | 2.4821350652576 × 10−9 | |
| 0.8 | 0.1.67578970 × 10−6 | 4.1510297361 × 10−8 | 2.58970263849 × 10−9 | 3.6558718785258 × 10−9 | |
| 1.0 | 2.0742841549 × 10−6 | 5.4841549427 × 10−9 | 6.28415494269 × 10−9 | 5.0422736406875 × 10−9 | |
| t | LADM-Pade [29] | Bessel Coll. N = 8 [30] | PIA(1,1) [31] | MVIM [32] | DTM N = 6 [33] |
|---|---|---|---|---|---|
| 0.2 | 8.87201671 × 10−7 | 9.28901670999 × 10−7 | 1.14750167099 × 10−6 | 9.3870167099 × 10−7 | 8.47401671 × 10−7 |
| 0.4 | 1.71183628 × 10−5 | 1.19553719699 × 10−6 | 2.22083719699 × 10−6 | 1.7283719699 × 10−7 | 1.28695628 × 10−5 |
| 0.6 | 6.86040272 × 10−4 | 1.38452847400 × 10−6 | 4.01142847400 × 10−6 | 4.5916715259 × 10−6 | 2.14296871 × 10−4 |
| 0.8 | 4.71409543 × 10−3 | 1.74452298399 × 10−6 | 6.89982298399 × 10−6 | 1.9104677016 × 10−5 | 1.53068567 × 10−3 |
| 1.0 | 5.79650268 × 10−3 | 2.27607680900 × 10−6 | 1.10738768090 × 10−5 | 5.5659623191 × 10−5 | 6.94842092 × 10−3 |
| t | EGM N = 3 [50] | EGM N = 4 [50] | EGM N = 5 [50] | cGP(2)-Method | |
| 0.2 | 1.499198329 × 10−6 | 1.8433016709 × 10−6 | 9.43901670998 × 10−7 | 8.6688500106069 × 10−7 | |
| 0.4 | 1.160246280 × 10−5 | 1.1156371969 × 10−6 | 1.34143719699 × 10−6 | 1.0728542867849 × 10−7 | |
| 0.6 | 1.765284740 × 10−7 | 3.3482847399 × 10−7 | 1.09702847399 × 10−6 | 9.9566078337957 × 10−7 | |
| 0.8 | 1.049742298 × 10−5 | 1.4261229840 × 10−6 | 9.33322984000 × 10−7 | 8.2106806348695 × 10−7 | |
| 1.0 | 4.111623191 × 10−6 | 6.4517680900 × 10−7 | 7.30476809001 × 10−7 | 6.3432370331871 × 10−7 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Attaullah; Zeeshan; Tufail Khan, M.; Alyobi, S.; Yassen, M.F.; Prathumwan, D. A Computational Approach to a Model for HIV and the Immune System Interaction. Axioms 2022, 11, 578. https://doi.org/10.3390/axioms11100578
Attaullah, Zeeshan, Tufail Khan M, Alyobi S, Yassen MF, Prathumwan D. A Computational Approach to a Model for HIV and the Immune System Interaction. Axioms. 2022; 11(10):578. https://doi.org/10.3390/axioms11100578
Chicago/Turabian StyleAttaullah, Zeeshan, Muhammad Tufail Khan, Sultan Alyobi, Mansour F. Yassen, and Din Prathumwan. 2022. "A Computational Approach to a Model for HIV and the Immune System Interaction" Axioms 11, no. 10: 578. https://doi.org/10.3390/axioms11100578
APA StyleAttaullah, Zeeshan, Tufail Khan, M., Alyobi, S., Yassen, M. F., & Prathumwan, D. (2022). A Computational Approach to a Model for HIV and the Immune System Interaction. Axioms, 11(10), 578. https://doi.org/10.3390/axioms11100578