

Open Problems and Conjectures in the Evolutionary Periodic Ricker Competition Model


Abstract
:1. Introduction
2. Single-Species Evolutionary Ricker Model
3. Multi-Species Ricker Competition Model
4. Open Problems and Conjectures
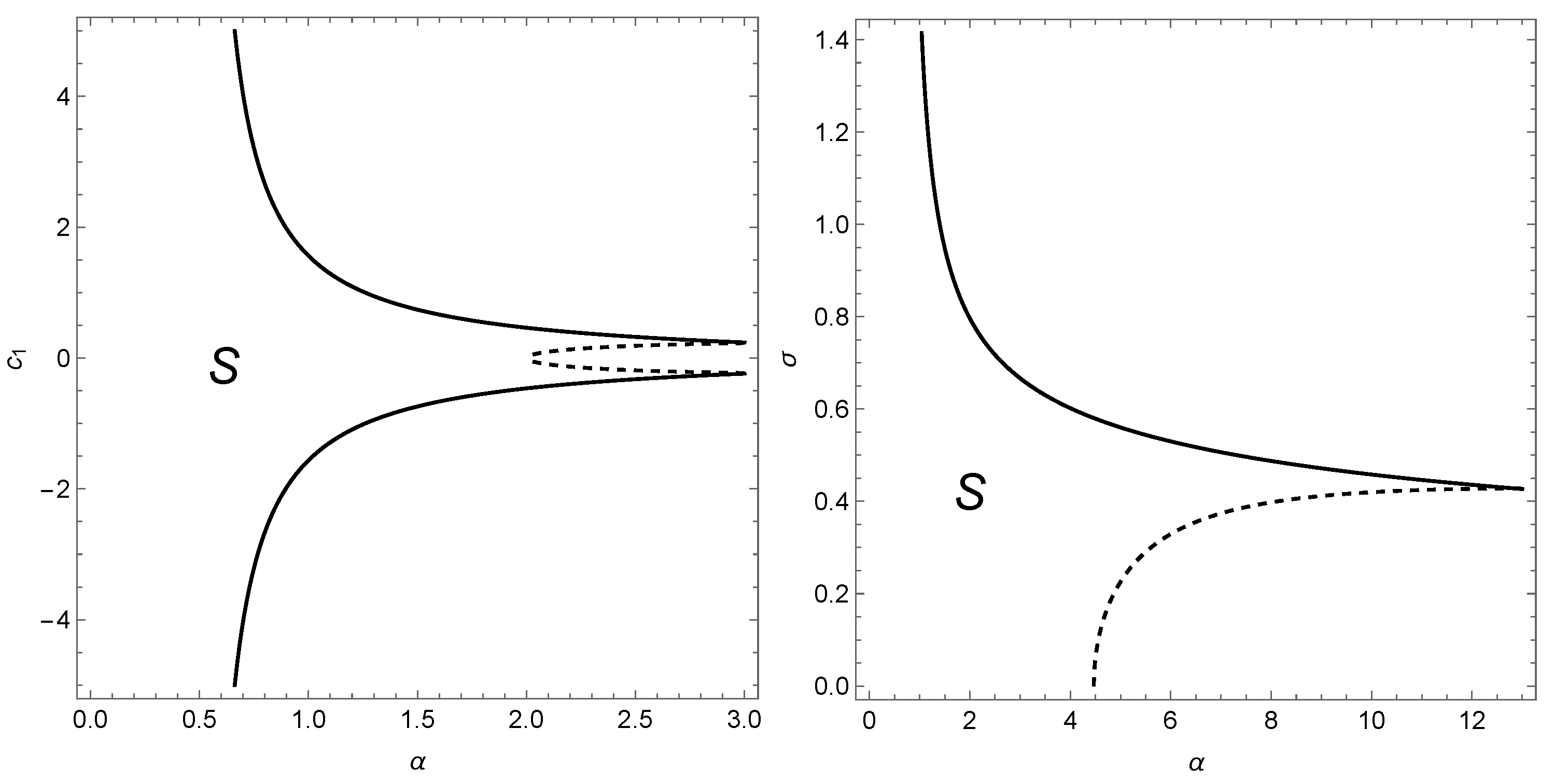
4.1. Single Species
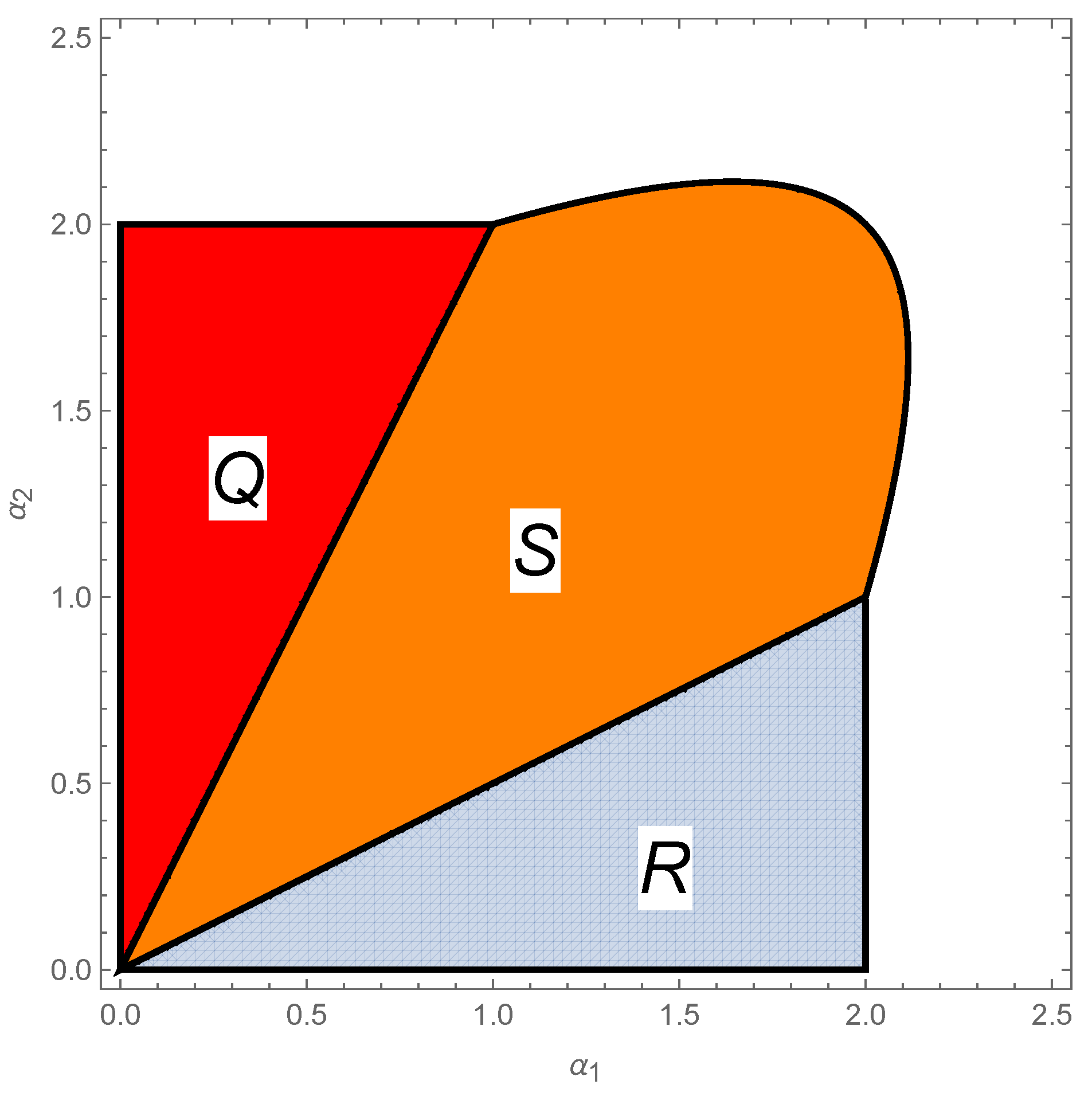
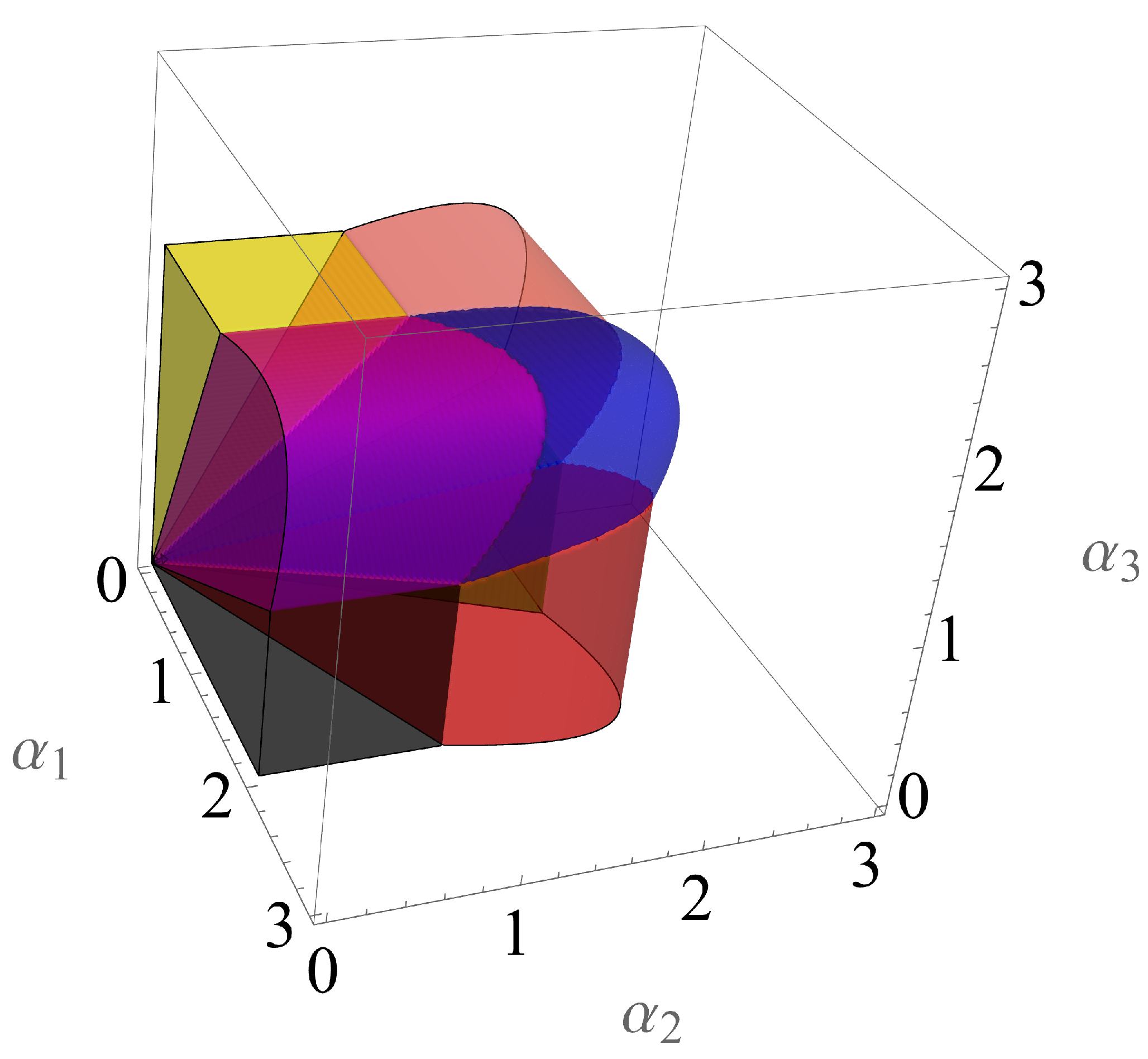
4.2. Multiple Species
5. Conclusions
Funding
Data Availability Statement
Conflicts of Interest
References
- Darwin, C. The Origin of Species; Avenel Books: London, UK, 1859. [Google Scholar]
- Ackleh, A.S.; Cushing, J.M.; Salceanu, P.L. On the dynamics of ecolutionary competition models. Nat. Resurce Model. 2015, 28, 380–397. [Google Scholar] [CrossRef]
- Cushing, J.M. An Evolutionary Beverton-Holt Model. In Springer Proceedings in Mathematics & Statistics, Proceedings of the Theory and Applications of Difference Equations and Discrete Dynamical Systems: ICDEA, Muscat, Oman, 26–30 May 2013; AlSharawi, Z., Cushing, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar]
- Cushing, J.M. Difference Equations as models of evolutionary population dynamics. J. Biol. Dyn. 2019, 13, 103–127. [Google Scholar] [CrossRef] [PubMed]
- Cushing, J.M. A Darwinian Ricker equation. In Springer Proceedings in Mathematics & Statistics, Proceedings of the Progress on Difference Equations and Discrete Dynamical Systems: 25th ICDEA, London, UK, 24–28 June 2019; Baigent, S., Bhoner, M., Eds.; Switzerland AG: Zurich, Switzerland, 2020; pp. 231–243. [Google Scholar]
- Cushing, J.M.; Stefanko, K. A Darwinian dynamic model for the evolution of post-reproduction survival. J. Biol. Syst. 2021, 29, 433–450. [Google Scholar] [CrossRef]
- Cushing, J.M. The evolutionary dynamics of a population model with a strong Allee effect. Math. Biosci. Eng. 2015, 12, 643–660. [Google Scholar] [CrossRef]
- Rael, R.C.; Vincent, T.L.; Cushing, J.M. Competitive outcomes changed by evolution. J. Biol. Dyn. 2011, 5, 227–252. [Google Scholar] [CrossRef]
- Vincent, T.; Brown, J. Evolutionary Game Theory, Natural Selection, and Darwinian Dynamics; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- Ackleh, A.S.; Hossain, M.I.; Veprauskas, A.; Zhang, A. Persistence and stability analysis of discrete-time predator–prey models: A study of population and evolutionary dynamics. J. Differ. Equ. Appl. 2019, 25, 1568–1603. [Google Scholar] [CrossRef]
- Elaydi, S.; Kang, Y.; Luís, R. The effects of evolution on the stability of competing species. J. Biol. Dyn. 2022, 16, 816–839. [Google Scholar] [CrossRef] [PubMed]
- Mokni, K.; Ch-Chaoui, M. Asymptotic Stability, Bifurcation Analysis and Chaos Control in a Discrete Evolutionary Ricker Population Model with Immigration. In Springer Proceedings in Mathematics & Statistics, Proceedings of the Advances in Discrete Dynamical Systems, Difference Equuation and Applications: 26th ICDEA, Sarajevo, Bosnia and Herzegovina, 26–30 July 2021; Elaydi, S., Kulenović, M.R.S., Kalabušić, R., Eds.; Springer International Publishing: Cham, Switzerland, 2023; pp. 363–403. [Google Scholar]
- Mokni, K.; Elaydi, S.; CH-Chaoui, M.; Eladdadi, A. Discrete evolutionary population models: A new approach. J. Biol. Dyn. 2020, 14, 454–478. [Google Scholar] [CrossRef] [PubMed]
- Ch-Chaoui, M.; Mokni, K. A discrete evolutionary Beverton–Holt population model. Int. J. Dyn. Control. 2023, 11, 1060–1075. [Google Scholar] [CrossRef]
- Elaydi, S.; Kang, Y.; Luís, R. Global asymptotic stability of evolutionary periodic Ricker competition models. J. Differ. Equ. Appl. 2023, 1–26. [Google Scholar] [CrossRef]
- Kulenovic, M.; Merino, O. A global attractivity result for maps with invariant boxes. Discret. Contin. Dyn. Syst.-B 2006, 6, 97–110. [Google Scholar] [CrossRef]
- Smith, H.L. The Discrete Dynamics of Monotonically Decomposable Maps. J. Math. Biol. 2006, 53, 747–758. [Google Scholar] [CrossRef] [PubMed]
- Smith, H.L. Global stability for mixed monotone systems. J. Differ. Equ. Appl. 2008, 14, 1159–1164. [Google Scholar] [CrossRef]
- Dercole, F.; Rinaldi, S. Analysis of Evolutionary Processes: The Adaptive Dynamics Approach and Its Applications; Princeton Series in Theoretical and Computational Biology; Princeton University Press: Princeton, NY, USA, 2008. [Google Scholar]
- Lande, R. Natural selection and random genetic drift in phenotypic evolution. Evolution 1976, 30, 314–334. [Google Scholar] [CrossRef] [PubMed]
- Lande, R. A Quantitative Genetic Theory of Life History Evolution. Ecology 1982, 63, 607–615. [Google Scholar] [CrossRef]
- Abrams. Modelling the adaptive dynamics of traits involved in inter- and intraspecific interactions: An assessment of three methods. Ecol. Lett. 2001, 4, 166–175. [Google Scholar] [CrossRef]
- Luís, R.; Elaydi, S.; Oliveira, H. Stability of a Ricker-type competition model and the competitive exclusion principle. J. Biol. Dyn. 2011, 5, 636–660. [Google Scholar] [CrossRef]
- Luís, R.; Rodrigues, E. Local Stability in 3D Discrete Dynamical Systems: Application to a Ricker Competition Model. Discret. Dyn. Nat. Soc. 2017, 2017, 16. [Google Scholar] [CrossRef]
- Baigent, S.; Hou, Z.; Elaydi, S.; Balreira, E.C.; Luís, R. A global picture for the planar Ricker map: Convergence to fixed points and identification of the stable/unstable manifolds. J. Differ. Equ. Appl. 2023, 29, 575–591. [Google Scholar] [CrossRef]
- Balreira, E.C.; Elaydi, S.; Luís, R. Local stability implies global stability for the planar Ricker competition model. Discret. Contin. Dyn. Syst.-Ser. B 2014, 19, 323–351. [Google Scholar] [CrossRef]
- Ryals, B.; Sacker, R.J. Global stability in the 2D Ricker equation. J. Differ. Equ. Appl. 2015, 21, 1068–1081. [Google Scholar] [CrossRef]
- Ryals, B.; Sacker, R.J. Global Stability in the 2D Ricker Equation-Revisited. Discret. Contin. Dyn. Syst.-Ser. B 2017, 22, 585–604. [Google Scholar] [CrossRef]
- Gyllenberg, M.; Jiang, J.; Niu, L. A note on global stability of three-dimensional Ricker models. J. Differ. Equ. Appl. 2019, 25, 142–150. [Google Scholar] [CrossRef]
- Balreira, E.C.; Elaydi, S.; Luís, R. Global stability of higher dimensional monotone maps. J. Differ. Equ. Appl. 2017, 23, 2037–2071. [Google Scholar] [CrossRef]
- Balreira, E.C.; Luís, R. Geometry and Global Stability of 2D Periodic Monotone Maps. J. Dyn. Differ. Equations 2021, 35, 2185–2198. [Google Scholar] [CrossRef]
- Luís, R. Linear Stability Conditions for a First Order n-Dimensional Mapping. Qual. Theory Dyn. Syst. 2021, 20, 20. [Google Scholar] [CrossRef]
- Jury, E. A Simplified Stability Criterion for Linear Discrete Systems. Proc. IRE 1962, 50, 1493–1500. [Google Scholar] [CrossRef]
- Jury, E. On the roots of a real polynomial inside the unit circle and a stability criterion for linear discrete systems. Ifac Proc. Vol. 1963, 1, 142–153. [Google Scholar] [CrossRef]
- Sacker, R. A note on periodic Ricker map. J. Differ. Equ. Appl. 2007, 13, 89–92. [Google Scholar] [CrossRef]
- Liz, E. On the global stability of periodic Ricker maps. Electron. J. Qual. Theory Differ. Equ. 2016, 2016, 1–8. [Google Scholar] [CrossRef]
- Anagnostopoulou, V.; Pötzsche, C.; Rasmussen, M. Nonautonomous Bifurcation Theory: Concepts and Tools; Frontiers in Applied Dynamical Systems: Reviews and Tutorials; Springer: Berlin/Heidelberg, Germany, 2023. [Google Scholar] [CrossRef]
- Pötzsche, C. Bifurcations in nonautonomous dynamical systems: Results and tools in discrete time. In Proceedings of the International Workshop Future Directions in Difference Equations, Vigo, Spain, 13–17 June 2011; pp. 163–212. [Google Scholar]
- Pötzsche, C. Bifurcations in a periodic discrete-time environment. Nonlinear Anal. Real World Appl. 2013, 14, 53–82. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luís, R. Open Problems and Conjectures in the Evolutionary Periodic Ricker Competition Model. Axioms 2024, 13, 246. https://doi.org/10.3390/axioms13040246
Luís R. Open Problems and Conjectures in the Evolutionary Periodic Ricker Competition Model. Axioms. 2024; 13(4):246. https://doi.org/10.3390/axioms13040246
Chicago/Turabian StyleLuís, Rafael. 2024. "Open Problems and Conjectures in the Evolutionary Periodic Ricker Competition Model" Axioms 13, no. 4: 246. https://doi.org/10.3390/axioms13040246
APA StyleLuís, R. (2024). Open Problems and Conjectures in the Evolutionary Periodic Ricker Competition Model. Axioms, 13(4), 246. https://doi.org/10.3390/axioms13040246