AFM Images of Viroid-Sized Rings That Self-Assemble from Mononucleotides through Wet–Dry Cycling: Implications for the Origin of Life




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
- Oligomers produced by wet-dry cycling of mononucleotides form pellets when isolated by precipitation in 70% ethanol or with spin tubes designed to purify nucleic acids. The oligomers exhibit UV spectra that are characteristic of the mononucleotides composing them.
- The RNA-like oligomers synthesized by wet–dry cycles are recognized by the enzymes used to label RNA with radioactive phosphate. When the labeled material is analyzed by standard methods of gel electrophoresis, it moves through the gel as expected for polyanions and shows up as a band ranging from 20 to >100 nucleotides in length [16]. The oligomers can also bind dyes such as ethidium bromide and SYBR-SAFE that are used to stain RNA polymers in gels [17].
- When tested by nanopore analysis with the alpha-hemolysin pore, the oligomers produce blockade signals virtually identical to those caused by single stranded RNA molecules. This demonstrates that at least some of the products are linear polyanionic strands that impede ionic currents as they are driven through the nanopore by an applied voltage [16,17,18].
- If a 1:1 mole ratio of AMP and UMP is exposed to wet–dry cycling, the oligomeric products exhibit hyperchromicity, but the products from AMP alone do not [17]. This result is consistent with hairpin structures forming in random sequence linear polymers of RNA composed of monomers capable of Watson-Crick base pairing.
- An X-ray diffraction study of AMP-UMP mixtures revealed that linear arrays of stacked bases are present with 3.4 Å distances between the bases [19]. The arrays are referred to as pre-polymers which presumably can form phosphodiester bonds during wet–dry cycles that link them into polymers by condensation reactions.
- We also note that cyclic guanosine monophosphate has been shown to undergo spontaneous polymerization in the dry state resulting in oligomeric products up to 80 nucleotides in length [20].
2. Materials and Methods
2.1. Atomic Force Microscopy
2.2. Compounds and Materials
2.3. Wet-Dry Cycles on Glass Substrates
2.4. Experiments on Mica Surfaces
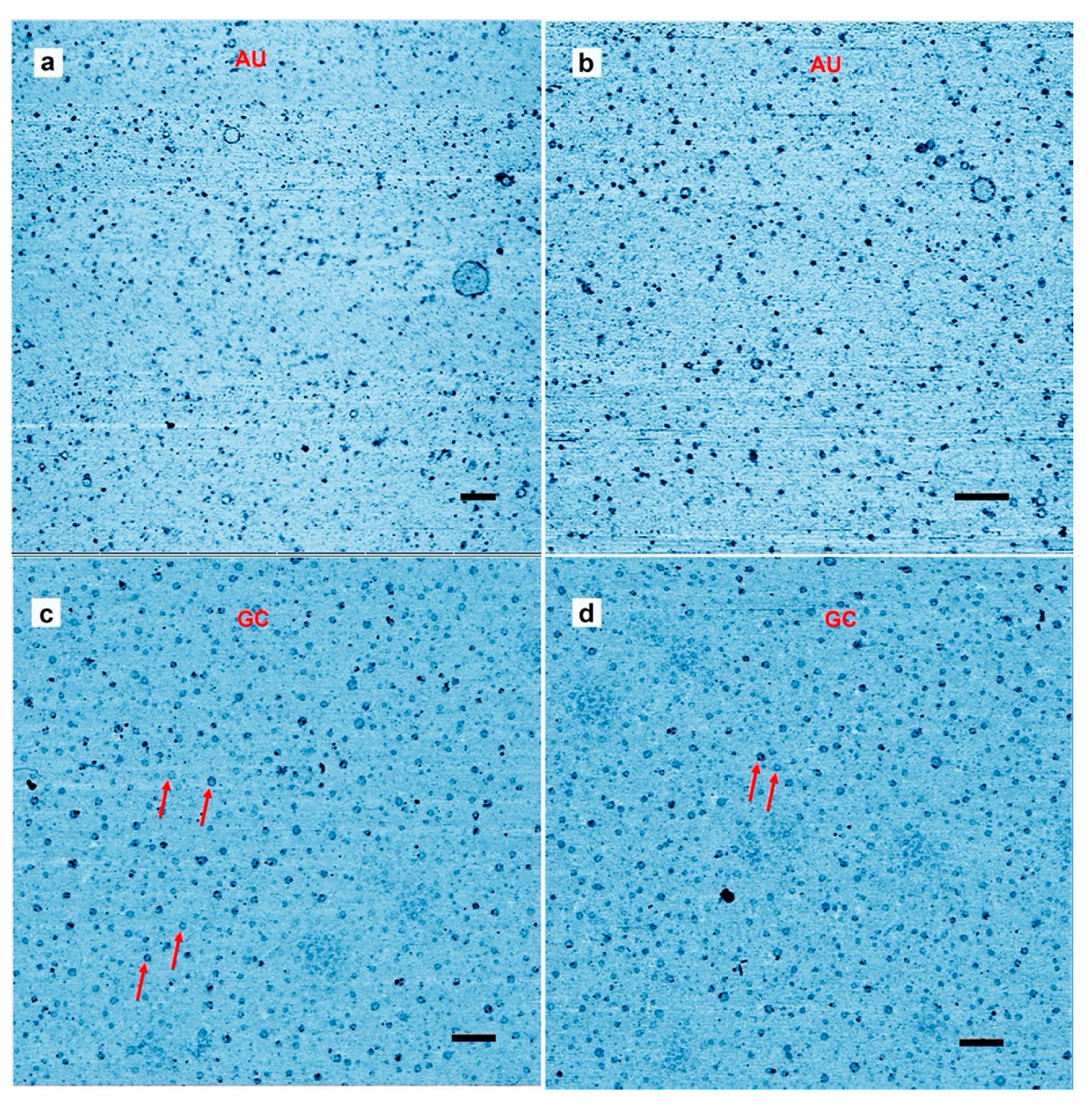
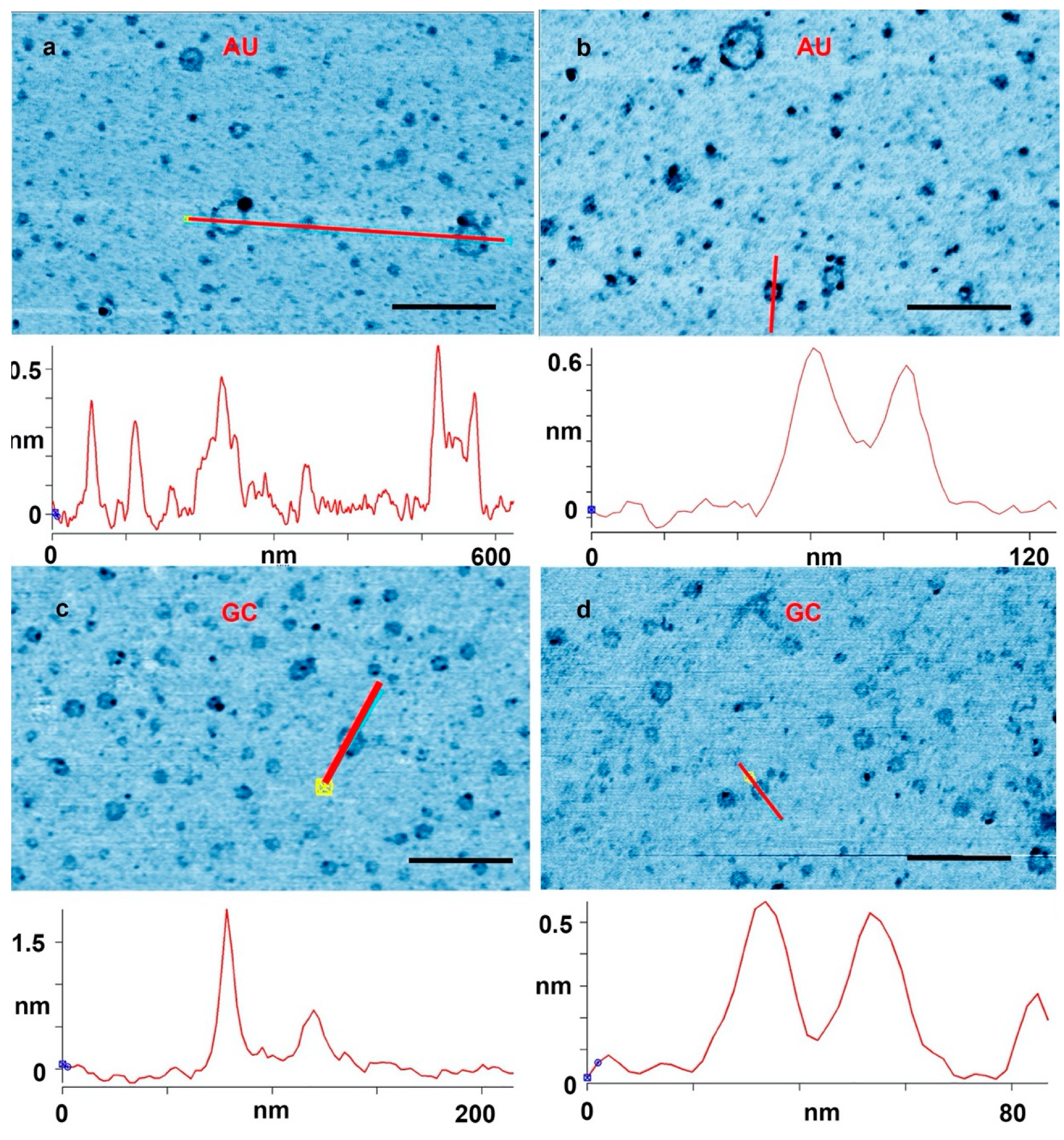
3. Results and Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hassenkam, T.; Andersson, M.P.; Dalby, K.N.; Mackenzie, D.M.A.; Rosing, M.T. Elements of Eoarchean life trapped in mineral inclusions. Nature 2017, 548, 78. [Google Scholar] [CrossRef] [PubMed]
- Rosing, M.T. C-13-depleted carbon microparticles in > 3700-Ma sea-floor sedimentary rocks from west Greenland. Science 1999, 283, 674–676. [Google Scholar] [CrossRef] [PubMed]
- Djokic, T.; Van Kranendonk, M.J.; Campbell, K.A.; Walter, M.R.; Ward, C.R. Earliest signs of life on land preserved in ca. 3.5 Ga hot spring deposits. Nat. Commun. 2017, 8, 15263. [Google Scholar] [CrossRef] [Green Version]
- Krishnamurthy, R. Experimentally investigating the origin of DNA/RNA on early Earth. Nat. Commun. 2018, 9, 5175. [Google Scholar] [CrossRef] [PubMed]
- Robertson, M.P.; Joyce, G.F. The Origins of the RNA World. Cold Spring Harb. Perspect. Biol. 2012, 4. [Google Scholar] [CrossRef]
- Furukawa, Y.; Chikaraishi, Y.; Ohkouchi, N.; Ogawa, N.O.; Glavin, D.P.; Dworkin, J.P.; Abe, C.; Nakamura, T. Extraterrestrial ribose and other sugars in primitive meteorites. Proc. Natl. Acad. Sci. USA 2019. [Google Scholar] [CrossRef] [Green Version]
- Pearce, B.K.D.; Pudritz, R.E.; Semenov, D.A.; Henning, T.K. Origin of the RNA world: The fate of nucleobases in warm little ponds. Proc. Natl. Acad. Sci. USA 2017, 114, 11327. [Google Scholar] [CrossRef] [Green Version]
- Deamer, D. Assembling Life; Oxford University Press: Oxford, UK, 2019. [Google Scholar]
- Deamer, D.; Damer, B. The hot spring hypothesis for an origin of life. Astrobiology 2019, 20. [Google Scholar] [CrossRef] [Green Version]
- Ross, D.; Deamer, D. Dry/Wet Cycling and the Thermodynamics and Kinetics of Prebiotic Polymer Synthesis. Life 2016, 6, 28. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Garcia, M.; Surman, A.J.; Cooper, G.J.T.; Suarez-Marina, I.; Hosni, Z.; Lee, M.P.; Cronin, L. Formation of oligopeptides in high yield under simple programmable conditions. Nat. Commun. 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Forsythe, J.G.; Yu, S.S.; Mamajanov, I.; Grover, M.A.; Krishnamurthy, R.; Fernandez, F.M.; Hud, N.V. Ester-Mediated Amide Bond Formation Driven by Wet–dry Cycles: A Possible Path to Polypeptides on the Prebiotic Earth. Angew. Chem. Int. Ed. 2015, 54, 9871–9875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayer, C.; Schreiber, U.; Davila, M.J.; Schmitz, O.J.; Bronja, A.; Meyer, M.; Klein, J.; Meckelmann, S.W. Molecular Evolution in a Peptide-Vesicle System. Life 2018, 8, 16. [Google Scholar] [CrossRef] [Green Version]
- Becker, S.; Feldmann, J.; Wiedemann, S.; Okamura, H.; Schneider, C.; Iwan, K.; Crisp, A.; Rossa, M.; Amatov, T.; Carell, T. Unified prebiotically plausible synthesis of pyrimidine and purine RNA ribonucleotides. Science 2019, 366, 76–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, T.; Ghule, S.; Vanka, K. Insights Into the Origin of Life: Did It Begin from HCN and H2O? ACS Cent. Sci. 2019, 5, 1532–1540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajamani, S.; Vlassov, A.; Benner, S.; Coombs, A.; Olasagasti, F.; Deamer, D. Lipid-assisted Synthesis of RNA-like Polymers from Mononucleotides. Orig. Life Evol. Biosph. 2008, 38, 57–74. [Google Scholar] [CrossRef]
- DeGuzman, V.; Vercoutere, W.; Shenasa, H.; Deamer, D. Generation of Oligonucleotides Under Hydrothermal Conditions by Non-enzymatic Polymerization. J. Mol. Evol. 2014, 78, 251–262. [Google Scholar] [CrossRef]
- Da Silva, L.; Maurel, M.C.; Deamer, D. Salt-Promoted Synthesis of RNA-like Molecules in Simulated Hydrothermal Conditions. J. Mol. Evol. 2014, 80, 86–97. [Google Scholar] [CrossRef]
- Himbert, S.; Chapman, M.; Deamer, D.W.; Rheinstadter, M.C. Organization of Nucleotides in Different Environments and the Formation of Pre-Polymers. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef] [Green Version]
- Morasch, M.; Mast, C.B.; Langer, J.K.; Schilcher, P.; Braun, D. Dry Polymerization of 3′,5′-Cyclic GMP to Long Strands of RNA. ChemBioChem 2014, 15, 879–883. [Google Scholar] [CrossRef]
- Deamer, D.; Damer, B.; Kompanichenko, V. Hydrothermal Chemistry and the Origin of Cellular Life. Astrobiology 2019, 19, 1523–1537. [Google Scholar] [CrossRef]
- Bizard, A.H.; Allemand, J.F.; Hassenkam, T.; Paramasivam, M.; Sarlos, K.; Singh, M.I.; Hickson, I.D. PICH and TOP3A cooperate to induce positive DNA supercoiling. Nat. Struct. Mol. Biol. 2019, 26, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Kunstelj, K.; Federiconi, F.; Spindler, L.; Drevenšek-Olenik, I. Self-organization of guanosine 5′-monophosphate on mica. Colloids Surfaces B Biointerfaces 2007, 59, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Hansma, H.G. The Power of Crowding for the Origins of Life. Orig. Life Evol. Biosph. 2014, 44, 307–311. [Google Scholar] [CrossRef] [PubMed]
- Hansma, H.G. Possible origin of life between mica sheets. J. Theor. Biol. 2010, 266, 175–188. [Google Scholar] [CrossRef]
- Li, J.; Mohammed-Elsabagh, M.; Paczkowski, F.; Li, Y. Circular Nucleic Acids: Discovery, Functions and Applications. ChemBioChem 2020, 21, 1547–1566. [Google Scholar] [CrossRef]
- Han, W.H.; Dlakic, M.; Zhu, Y.W.J.; Lindsay, S.M.; Harrington, R.E. Strained DNA is kinked by low concentrations of Zn2+. Proc. Natl. Acad. Sci. USA 1997, 94, 10565–10570. [Google Scholar] [CrossRef] [Green Version]
- Diener, T.O. The viroid: Biological oddity or evolutionary fossil? In Advances in Virus Research; Academic Press: Cambridge, MA, USA, 2001; Volume 57, pp. 137–184. [Google Scholar] [CrossRef]
- Darwin, C. Letter No. 7471. In Darwin Correspondence Project; University of Cambridge; Available online: https://www.darwinproject.ac.uk/letter/DCP-LETT-7471.xml (accessed on 30 November 2020).





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hassenkam, T.; Damer, B.; Mednick, G.; Deamer, D. AFM Images of Viroid-Sized Rings That Self-Assemble from Mononucleotides through Wet–Dry Cycling: Implications for the Origin of Life. Life 2020, 10, 321. https://doi.org/10.3390/life10120321
Hassenkam T, Damer B, Mednick G, Deamer D. AFM Images of Viroid-Sized Rings That Self-Assemble from Mononucleotides through Wet–Dry Cycling: Implications for the Origin of Life. Life. 2020; 10(12):321. https://doi.org/10.3390/life10120321
Chicago/Turabian StyleHassenkam, Tue, Bruce Damer, Gabriel Mednick, and David Deamer. 2020. "AFM Images of Viroid-Sized Rings That Self-Assemble from Mononucleotides through Wet–Dry Cycling: Implications for the Origin of Life" Life 10, no. 12: 321. https://doi.org/10.3390/life10120321
APA StyleHassenkam, T., Damer, B., Mednick, G., & Deamer, D. (2020). AFM Images of Viroid-Sized Rings That Self-Assemble from Mononucleotides through Wet–Dry Cycling: Implications for the Origin of Life. Life, 10(12), 321. https://doi.org/10.3390/life10120321