
Transcriptomic Analysis of Genes Involved in Plant Defense Response to the Cucumber Green Mottle Mosaic Virus Infection

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
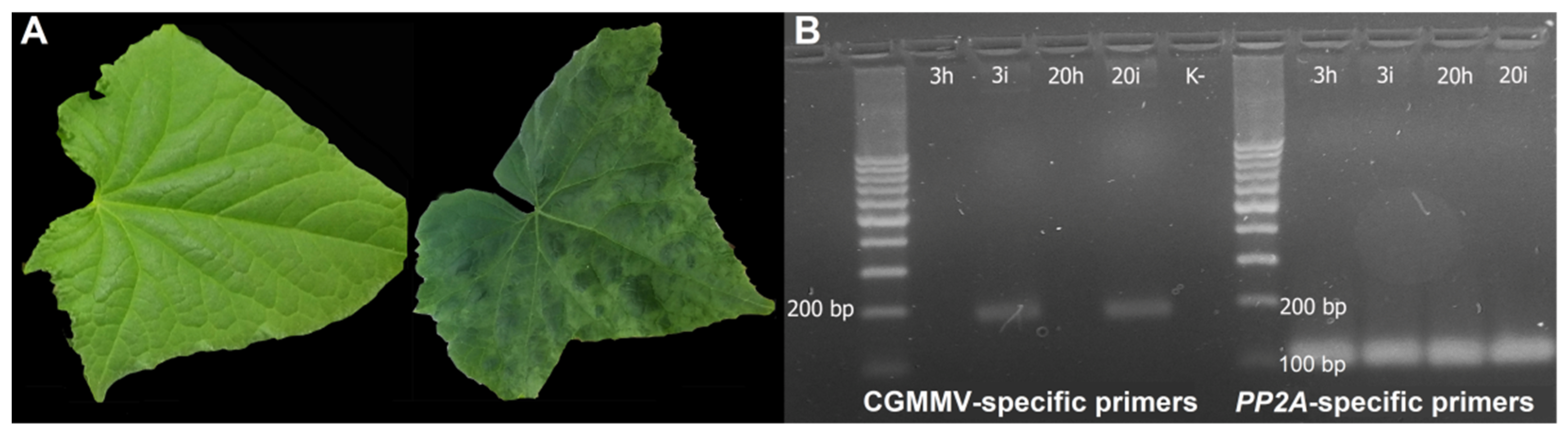
2.1. Plant Samples, CGMMV Treatment and Confirmation of Viral Infection
2.2. RNA Isolation
2.3. Library Preparation and Sequencing
2.4. Transcript Abundance Estimation and Differential Expression Analysis
2.5. Quantitative RT-PCR (qRT-PCR)
3. Results
3.1. CGMMV Infection Confirmation and Plant Phenotypes
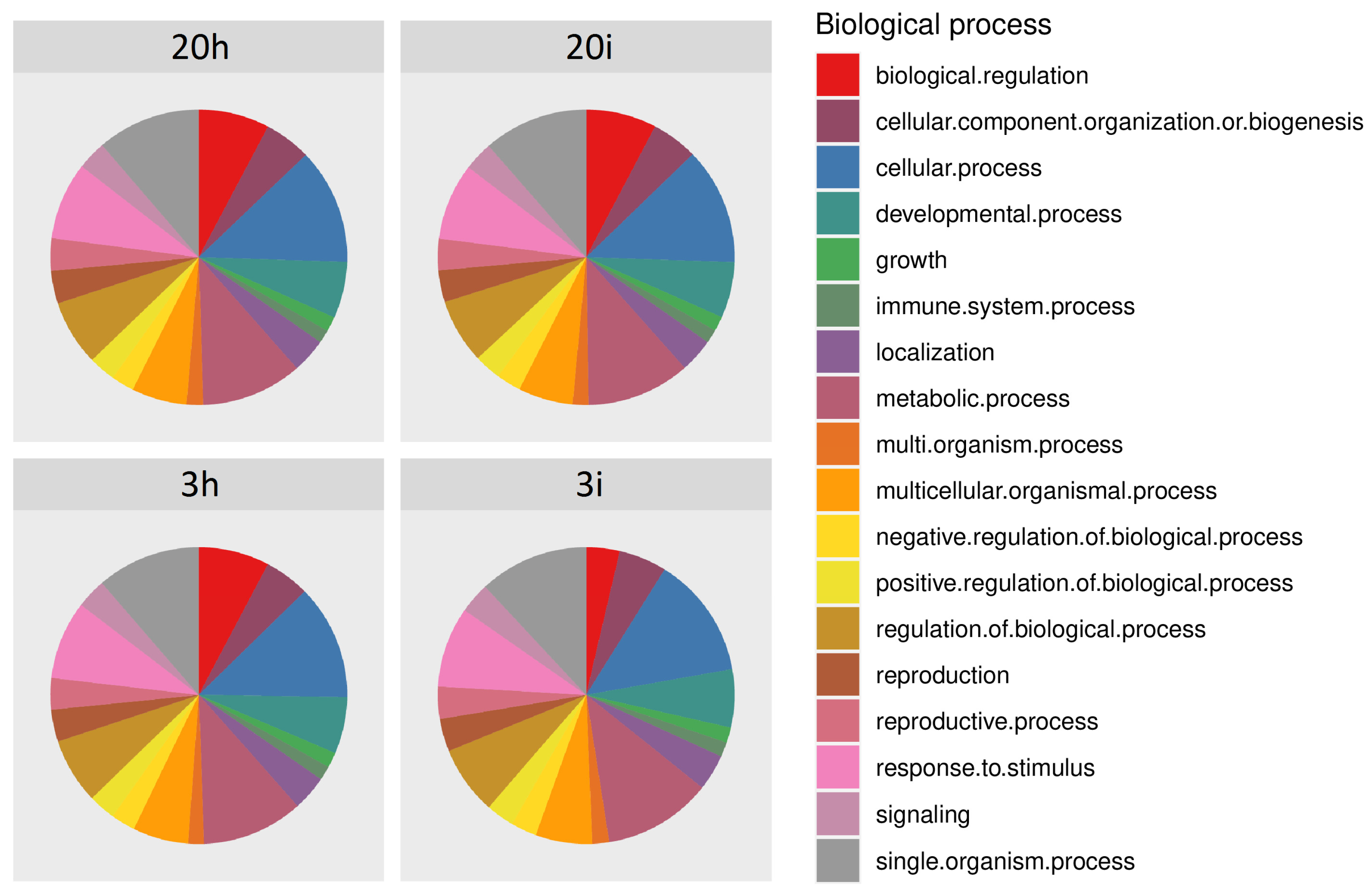
3.2. Overview of the RNA-seq Results
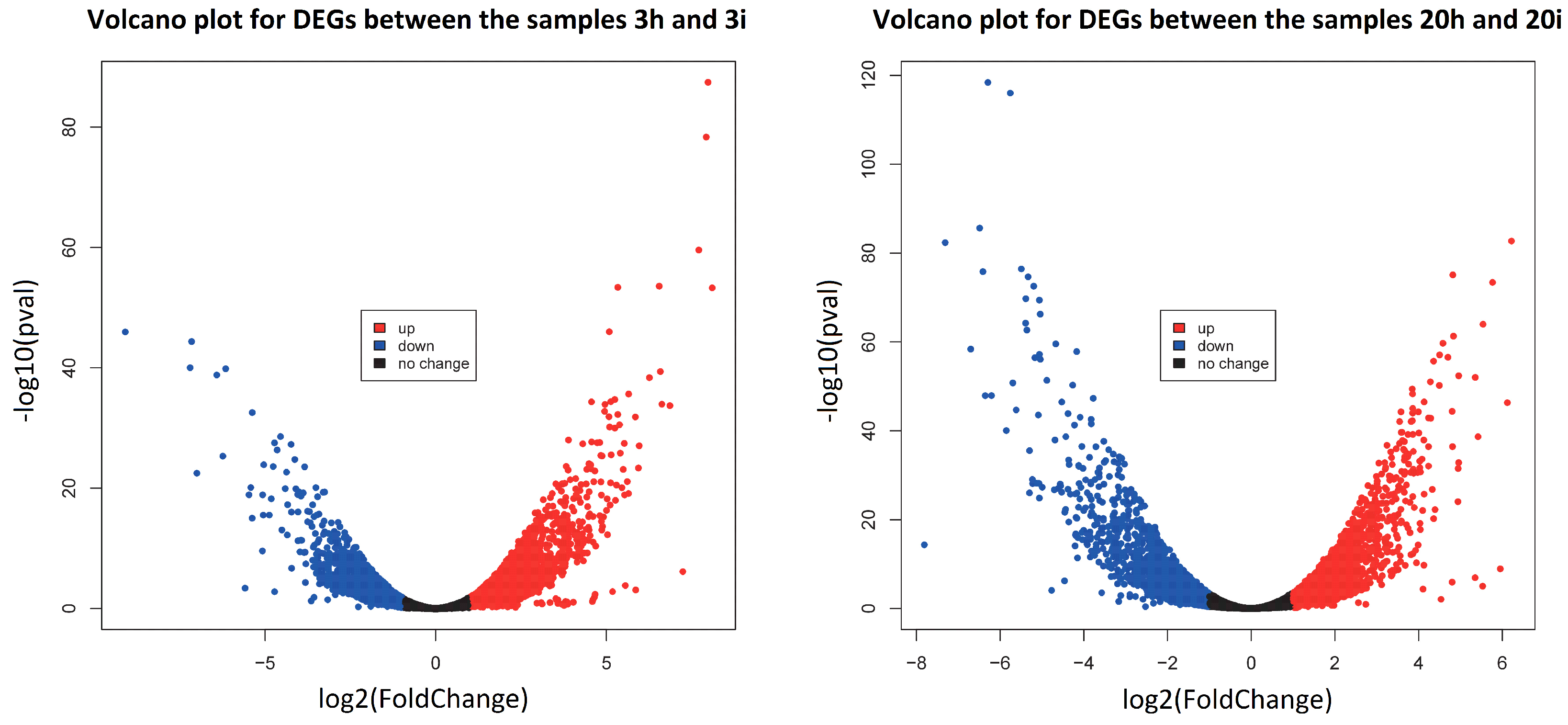
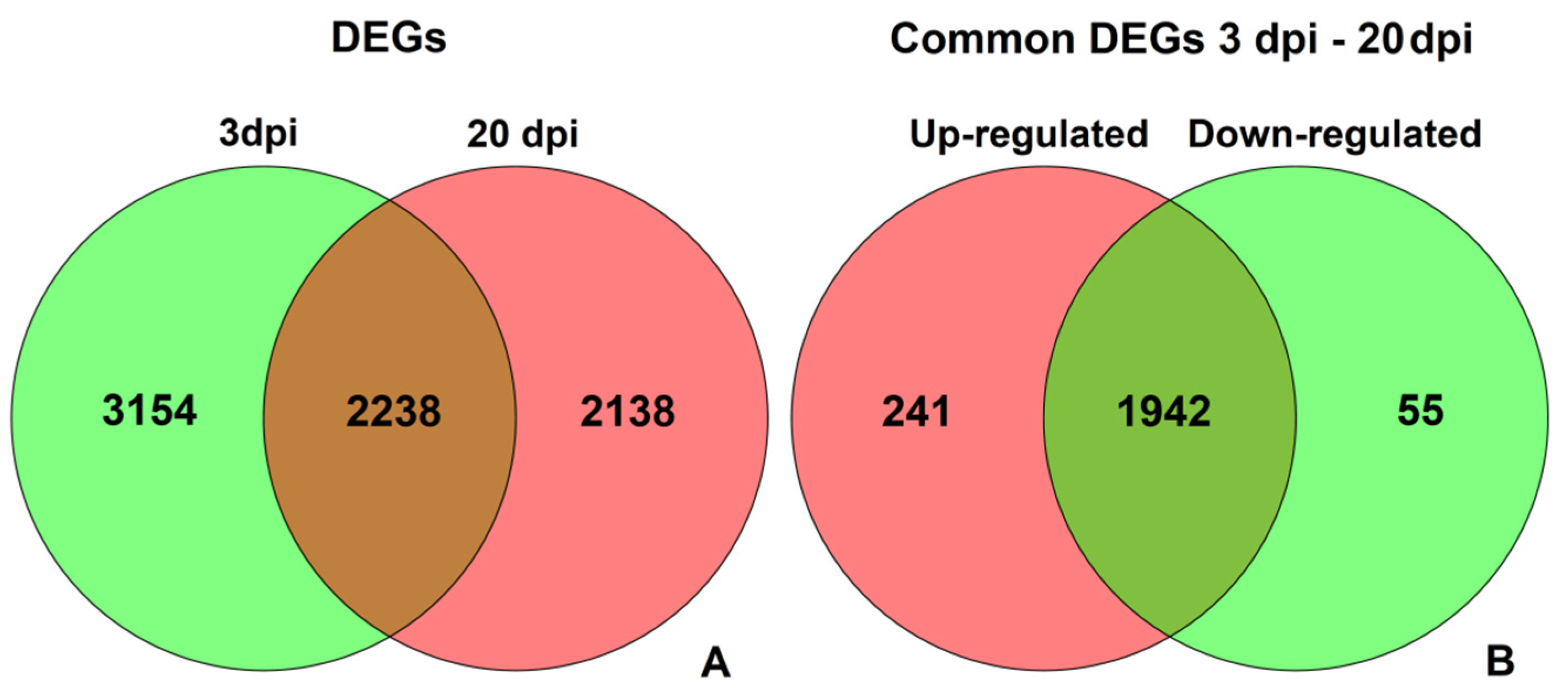
3.3. DEGs in Response to CGMMV-Induced Stress
3.4. DEGs Involved in Host Defense Response
3.5. RNA-seq Validation by qRT-PCR
4. Discussion
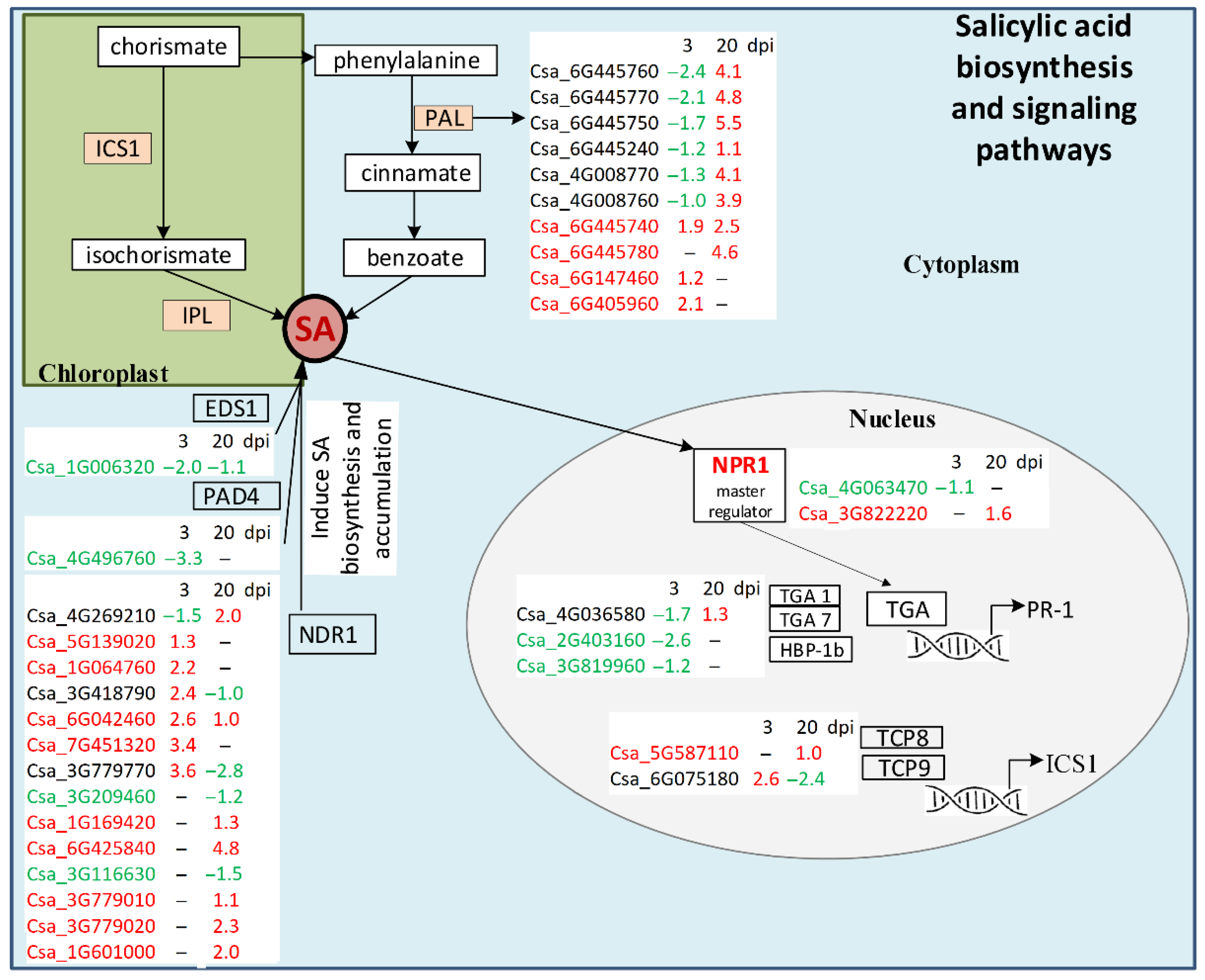
4.1. Salicylic Acid Biosynthesis and Signaling
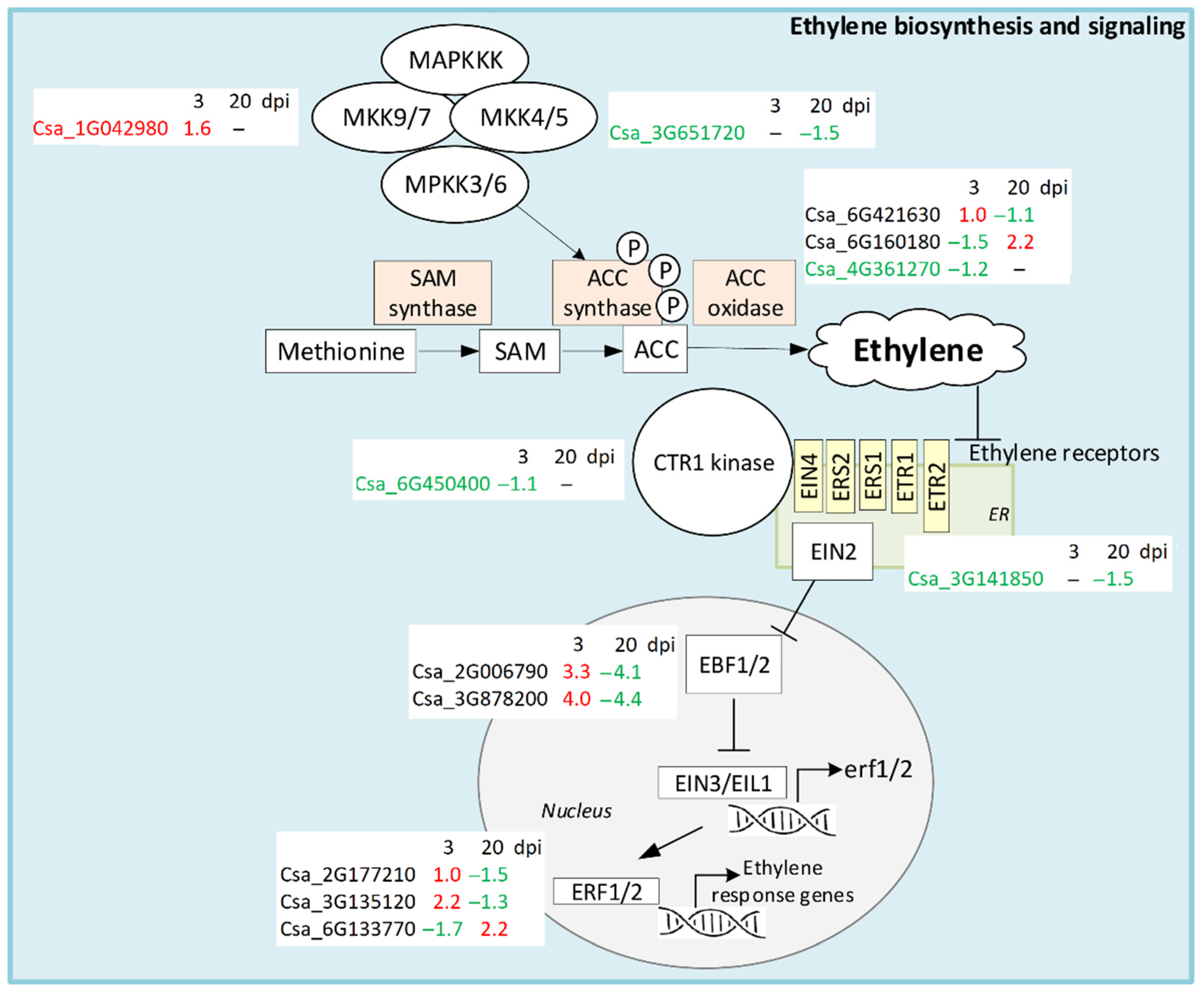
4.2. Ethylene Biosynthesis and Signaling
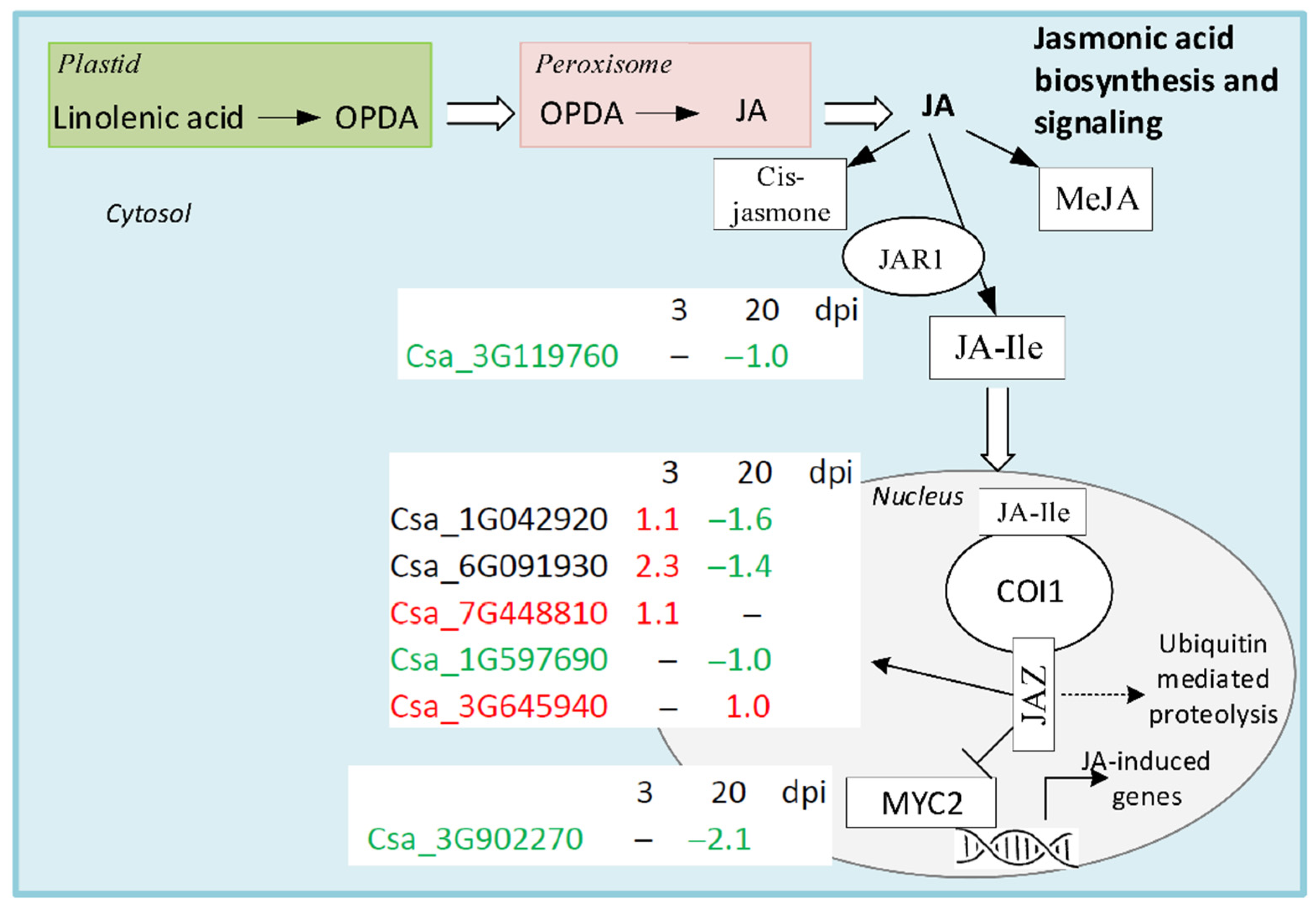
4.3. Jasmonic acid Biosynthesis and Signaling
4.4. Expression of PR-Protein Genes in Response to CGMMV Infection
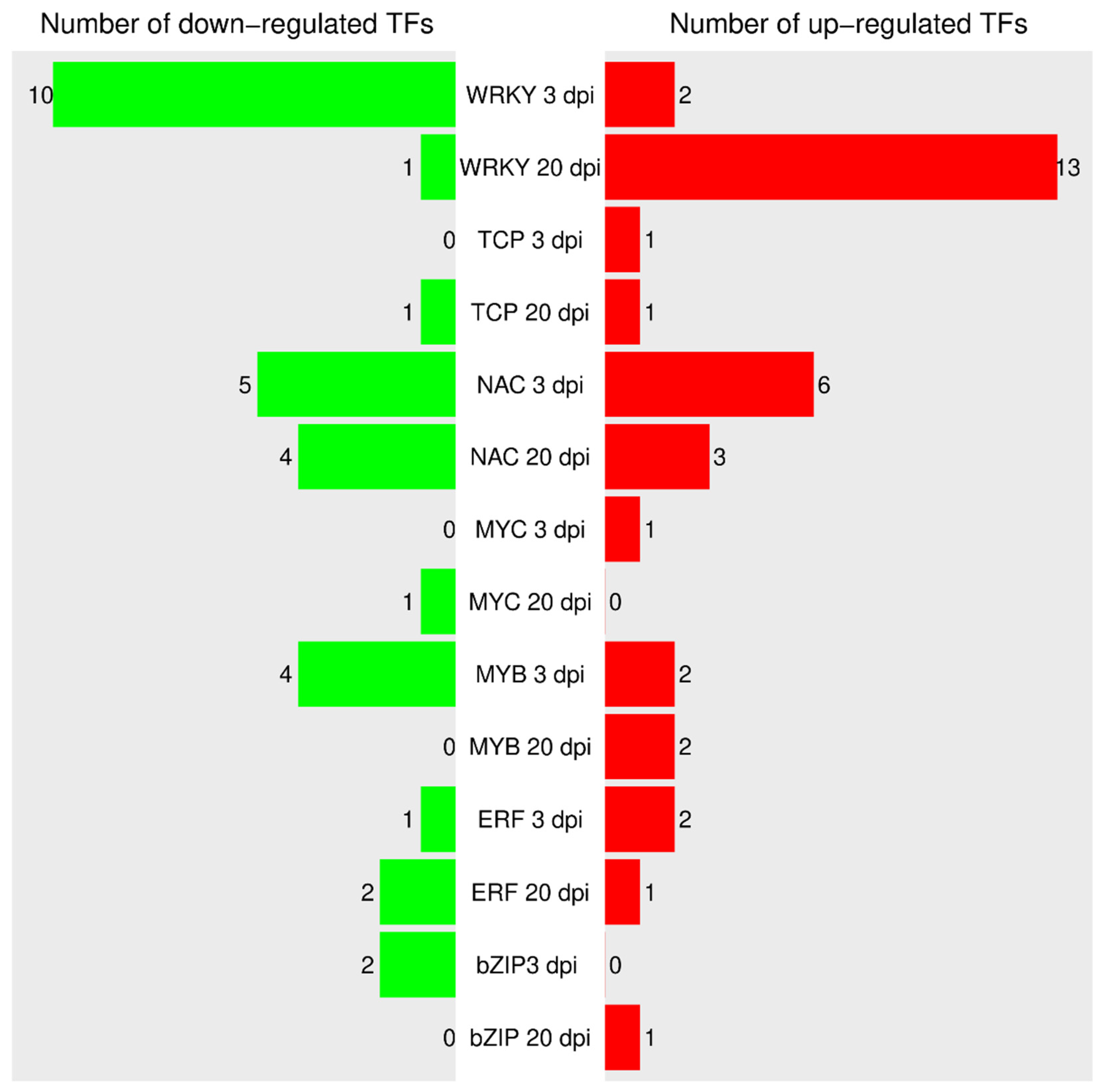
4.5. Transcription Factors Associated with Biotic Stress
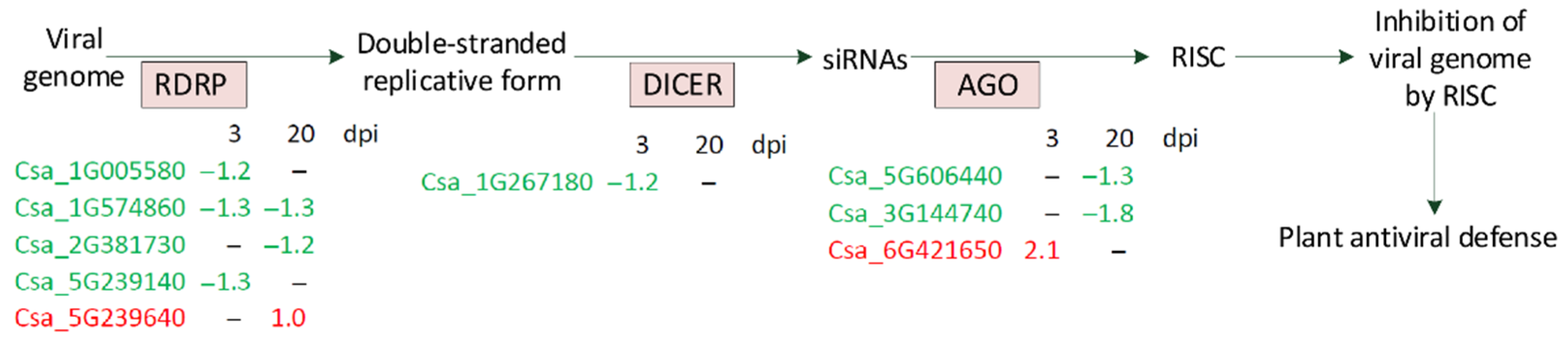
4.6. Silencing
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Osbourn, A.E. Plant mechanisms that give defence against soilborne diseases. Australas. Plant Pathol. 2001, 30, 99–102. [Google Scholar] [CrossRef]
- Zipfel, C.; Felix, G. Plants and animals: A different taste for microbes? Curr. Opin. Plant Biol. 2005, 8, 353–360. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Ganai, B.A.; Kamili, A.N.; Bhat, A.A.; Mir, Z.A.; Bhat, J.A.; Tyagi, A.; Islam, S.T.; Mushtaq, M.; Yadav, P.; et al. Pathogenesis-related proteins and peptides as promising tools for engineering plants with multiple stress tolerance. Microbiol. Res. 2018, 212, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Amorim, L.L.B.; da Fonseca Dos Santos, R.; Neto, J.P.B.; Guida-Santos, M.; Crovella, S.; Benko-Iseppon, A.M. Transcription Factors Involved in Plant Resistance to Pathogens. Curr. Protein Pept. Sci. 2017, 18, 335–351. [Google Scholar] [CrossRef] [PubMed]
- Wasternack, C.; Song, S. Jasmonates: Biosynthesis, metabolism, and signaling by proteins activating and repressing transcription. J. Exp. Bot. 2017, 68, 1303–1321. [Google Scholar] [CrossRef] [PubMed]
- Pumplin, N.; Voinnet, O. RNA silencing suppression by plant pathogens: Defence, counter-defence and counter-counter-defence. Nature reviews. Microbiology 2013, 11, 745–760. [Google Scholar] [CrossRef] [PubMed]
- Pasin, F.; Shan, H.; Garcia, B.; Muller, M.; San Leon, D.; Ludman, M.; Fresno, D.H.; Fatyol, K.; Munne-Bosch, S.; Rodrigo, G.; et al. Abscisic Acid Connects Phytohormone Signaling with RNA Metabolic Pathways and Promotes an Antiviral Response that Is Evaded by a Self-Controlled RNA Virus. Plant Commun. 2020, 1, 100099. [Google Scholar] [CrossRef] [PubMed]
- Dombrovsky, A.; Tran-Nguyen, L.T.T.; Jones, R.A.C. Cucumber green mottle mosaic virus: Rapidly increasing global distribution, etiology, epidemiology, and management. Annu. Rev. Phytopathol. 2017, 55, 231–256. [Google Scholar] [CrossRef]
- Slavokhotova, A.A.; Istomina, E.A.; Andreeva, E.N.; Korostyleva, T.V.; Pukhalskij, V.A.; Shijan, A.N.; Odintsova, T.I. An Attenuated Strain of Cucumber Green Mottle Mosaic Virus as a Biological Control Agent against Pathogenic Viral Strains. Am. J. Plant Sci. 2016, 7, 724–732. [Google Scholar] [CrossRef] [Green Version]
- Ainsworth, G.C. Mosaic Diseases of the Cucumber. Ann. Appl. Biol. 1935, 22, 55–67. [Google Scholar] [CrossRef]
- Varveri, C.; Vassilakos, N.; Bem, F. Characterization and detection ofCucumber green mottle mosaic virus in Greece. Phytoparasitica 2002, 30, 493–501. [Google Scholar] [CrossRef]
- Tesoriero, L.A.; Chambers, G.; Srivastava, M.; Smith, S.; Conde, B.; Tran-Nguyen, L.T.T. First report of cucumber green mottle mosaic virus in Australia. Australas. Plant Dis. Notes 2015, 11. [Google Scholar] [CrossRef]
- Matasci, N.; Hung, L.H.; Yan, Z.; Carpenter, E.J.; Wickett, N.J.; Mirarab, S.; Nguyen, N.; Warnow, T.; Ayyampalayam, S.; Barker, M.; et al. Data access for the 1,000 Plants (1KP) project. Gigascience 2014, 3, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adhikary, D.; Khatri-Chhetri, U.; Tymm, F.J.M.; Murch, S.J.; Deyholos, M.K. A virus-induced gene-silencing system for functional genetics in a betalainic species, Amaranthus tricolor (Amaranthaceae). Appl. Plant Sci. 2019, 7, e01221. [Google Scholar] [CrossRef] [Green Version]
- Shelenkov, A.; Slavokhotova, A.; Odintsova, T. Predicting Antimicrobial and Other Cysteine-Rich Peptides in 1267 Plant Transcriptomes. Antibiotics 2020, 9, 60. [Google Scholar] [CrossRef] [Green Version]
- Nelson, D.R. Cytochrome P450 diversity in the tree of life. Biochim. Biophys. Acta Proteins Proteom. 2018, 1866, 141–154. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Fan, M.; He, Y. Transcriptome Analysis of Watermelon Leaves Reveals Candidate Genes Responsive to Cucumber green mottle mosaic virus Infection. Int. J. Mol. Sci. 2019, 20, 610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; An, M.; Xia, Z.; Bai, X.; Wu, Y. Transcriptome analysis of watermelon (Citrullus lanatus) fruits in response to Cucumber green mottle mosaic virus (CGMMV) infection. Sci. Rep. 2017, 7, 16747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, L.; Zong, X.; Wang, J.; Wei, H.; Chen, X.; Liu, Q. Transcriptomic analysis reveals insights into the response to Hop stunt viroid (HSVd) in sweet cherry (Prunus avium L.) fruits. Peer J. 2020, 8, e10005. [Google Scholar] [CrossRef] [PubMed]
- Slavokhotova, A.A.; Andreeva, E.N.; Shiian, A.N.; Odintsova, T.I.; Pukhalskij, V.A. Specifics of the coat protein gene in Russian strains of the cucumber green mottle mosaic virus. Genetika 2007, 43, 1461–1467. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shelenkov, A.A.; Slavokhotova, A.A.; Odintsova, T.I. Cysmotif Searcher Pipeline for Antimicrobial Peptide Identification in Plant Transcriptomes. Biochemistry 2018, 83, 1424–1432. [Google Scholar] [CrossRef]
- Slavokhotova, A.A.; Shelenkov, A.A.; Korostyleva, T.V.; Rogozhin, E.A.; Melnikova, N.V.; Kudryavtseva, A.V.; Odintsova, T.I. Defense peptide repertoire of Stellaria media predicted by high throughput next generation sequencing. Biochimie 2017, 135, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Ricachenevsky, F.K.; Menguer, P.K.; Sperotto, R.A.; Williams, L.E.; Fett, J.P. Roles of plant metal tolerance proteins (MTP) in metal storage and potential use in biofortification strategies. Front. Plant Sci. 2013, 4, 144. [Google Scholar] [CrossRef] [Green Version]
- D’Auria, J.C. Acyltransferases in plants: A good time to be BAHD. Curr. Opin. Plant Biol. 2006, 9, 331–340. [Google Scholar] [CrossRef] [PubMed]
- Gujjar, R.S.; Pathak, A.D.; Karkute, S.G.; Supaibulwatana, K. Multifunctional proline rich proteins and their role in regulating cellular proline content in plants under stress. Biol. Plant 2019, 63, 448–454. [Google Scholar] [CrossRef]
- Stein, O.; Granot, D. An Overview of Sucrose Synthases in Plants. Front. Plant Sci. 2019, 10, 95. [Google Scholar] [CrossRef] [Green Version]
- Gonzales-Vigil, E.; Bianchetti, C.M.; Phillips, G.N.; Howe, G.A. Adaptive evolution of threonine deaminase in plant defense against insect herbivores. Proc. Natl. Acad. Sci. USA 2011, 108, 5897–5902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalunke, R.M.; Tundo, S.; Benedetti, M.; Cervone, F.; De Lorenzo, G.; D’Ovidio, R. An update on polygalacturonase-inhibiting protein (PGIP), a leucine-rich repeat protein that protects crop plants against pathogens. Front. Plant Sci. 2015, 6, 146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, W.L.; Hsiao, Y.Y.; Lu, H.C.; Liang, C.K.; Fu, C.H.; Huang, T.H.; Chuang, M.H.; Chen, L.J.; Liu, Z.J.; Tsai, W.C. Expression regulation of MALATE SYNTHASE involved in glyoxylate cycle during protocorm development in Phalaenopsis aphrodite (Orchidaceae). Sci. Rep. 2020, 10, 10123. [Google Scholar] [CrossRef]
- Langenkamper, G.; Manac’h, N.; Broin, M.; Cuine, S.; Becuwe, N.; Kuntz, M.; Rey, P. Accumulation of plastid lipid-associated proteins (fibrillin/CDSP34) upon oxidative stress, ageing and biotic stress in Solanaceae and in response to drought in other species. J. Exp. Bot. 2001, 52, 1545–1554. [Google Scholar] [CrossRef] [Green Version]
- Boatright, J.; Negre, F.; Chen, X.; Kish, C.M.; Wood, B.; Peel, G.; Orlova, I.; Gang, D.; Rhodes, D.; Dudareva, N. Understanding in vivo benzenoid metabolism in petunia petal tissue. Plant Physiol. 2004, 135, 1993–2011. [Google Scholar] [CrossRef] [Green Version]
- Qi, G.; Chen, J.; Chang, M.; Chen, H.; Hall, K.; Korin, J.; Liu, F.; Wang, D.; Fu, Z.Q. Pandemonium Breaks Out: Disruption of Salicylic Acid-Mediated Defense by Plant Pathogens. Mol. Plant 2018, 11, 1427–1439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dempsey, D.A.; Vlot, A.C.; Wildermuth, M.C.; Klessig, D.F. Salicylic Acid biosynthesis and metabolism. Arab. Book 2011, 9, e0156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dao, T.T.; Linthorst, H.J.; Verpoorte, R. Chalcone synthase and its functions in plant resistance. Phytochem. Rev. 2011, 10, 397–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Correa, L.G.; Riano-Pachon, D.M.; Schrago, C.G.; dos Santos, R.V.; Mueller-Roeber, B.; Vincentz, M. The role of bZIP transcription factors in green plant evolution: Adaptive features emerging from four founder genes. PLoS ONE 2008, 3, e2944. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Liu, D. Ethylene and plant responses to phosphate deficiency. Front. Plant Sci. 2015, 6, 796. [Google Scholar] [CrossRef] [Green Version]
- Iwai, T.; Miyasaka, A.; Seo, S.; Ohashi, Y. Contribution of ethylene biosynthesis for resistance to blast fungus infection in young rice plants. Plant Physiol. 2006, 142, 1202–1215. [Google Scholar] [CrossRef] [Green Version]
- Chang, C. Q&A: How do plants respond to ethylene and what is its importance? BMC Biol. 2016, 14, 7. [Google Scholar] [CrossRef] [Green Version]
- Jagodzik, P.; Tajdel-Zielinska, M.; Ciesla, A.; Marczak, M.; Ludwikow, A. Mitogen-Activated Protein Kinase Cascades in Plant Hormone Signaling. Front. Plant Sci. 2018, 9, 1387. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.S.; Baek, K.H. Jasmonic Acid Signaling Pathway in Response to Abiotic Stresses in Plants. Int. J. Mol. Sci. 2020, 21, 621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wager, A.; Browse, J. Social Network: JAZ Protein Interactions Expand Our Knowledge of Jasmonate Signaling. Front. Plant Sci. 2012, 3, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akbudak, M.A.; Yildiz, S.; Filiz, E. Pathogenesis related protein-1 (PR-1) genes in tomato (Solanum lycopersicum L.): Bioinformatics analyses and expression profiles in response to drought stress. Genomics 2020, 112, 4089–4099. [Google Scholar] [CrossRef]
- Oide, S.; Bejai, S.; Staal, J.; Guan, N.; Kaliff, M.; Dixelius, C. A novel role of PR2 in abscisic acid (ABA) mediated, pathogen-induced callose deposition in Arabidopsis thaliana. New Phytol. 2013, 200, 1187–1199. [Google Scholar] [CrossRef] [PubMed]
- Pusztahelyi, T. Chitin and chitin-related compounds in plant-fungal interactions. Mycology 2018, 9, 189–201. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Xiao, B.; Xiong, L. Identification of a cluster of PR4-like genes involved in stress responses in rice. J. Plant Physiol. 2011, 168, 2212–2224. [Google Scholar] [CrossRef]
- van Loon, L.C.; Rep, M.; Pieterse, C.M. Significance of inducible defense-related proteins in infected plants. Annu. Rev. Phytopathol. 2006, 44, 135–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bovie, C.; Ongena, M.; Thonart, P.; Dommes, J. Cloning and expression analysis of cDNAs corresponding to genes activated in cucumber showing systemic acquired resistance after BTH treatment. BMC Plant Biol. 2004, 4, 15. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Fan, M.; He, Y. DNA Methylation Analysis of the Citrullus lanatus Response to Cucumber Green Mottle Mosaic Virus Infection by Whole-Genome Bisulfite Sequencing. Genes 2019, 10, 344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, S.P.; Somssich, I.E. The role of WRKY transcription factors in plant immunity. Plant Physiol. 2009, 150, 1648–1655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, S.K.; Baek, K.H.; Park, J.M.; Yi, S.Y.; Yu, S.H.; Kamoun, S.; Choi, D. Capsicum annuum WRKY protein CaWRKY1 is a negative regulator of pathogen defense. New Phytol. 2008, 177, 977–989. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Li, M.Y.; Wu, P.; Xu, Z.S.; Que, F.; Wang, F.; Xiong, A.S. Members of WRKY Group III transcription factors are important in TYLCV defense signaling pathway in tomato (Solanum lycopersicum). BMC Genom. 2016, 17, 788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naoumkina, M.A.; He, X.; Dixon, R.A. Elicitor-induced transcription factors for metabolic reprogramming of secondary metabolism in Medicago truncatula. BMC Plant Biol. 2008, 8, 132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meraj, T.A.; Fu, J.; Raza, M.A.; Zhu, C.; Shen, Q.; Xu, D.; Wang, Q. Transcriptional Factors Regulate Plant Stress Responses through Mediating Secondary Metabolism. Genes 2020, 11, 346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, X.; Wang, H.; Cai, J.; Li, D.; Song, F. NAC transcription factors in plant immunity. Phytopathol. Res. 2019, 1, 3. [Google Scholar] [CrossRef]
- Nakashima, K.; Tran, L.S.; Van Nguyen, D.; Fujita, M.; Maruyama, K.; Todaka, D.; Ito, Y.; Hayashi, N.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Functional analysis of a NAC-type transcription factor OsNAC6 involved in abiotic and biotic stress-responsive gene expression in rice. Plant J. Cell Mol. Biol. 2007, 51, 617–630. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Yuan, Y.; Yang, C.; Geng, S.; Sun, Q.; Long, L.; Cai, C.; Chu, Z.; Liu, X.; Wang, G.; et al. Characterization, Expression, and Functional Analysis of a Novel NAC Gene Associated with Resistance to Verticillium Wilt and Abiotic Stress in Cotton. G3 2016, 6, 3951–3961. [Google Scholar] [CrossRef]
- Zhang, T.; Lv, W.; Zhang, H.; Ma, L.; Li, P.; Ge, L.; Li, G. Genome-wide analysis of the basic Helix-Loop-Helix (bHLH) transcription factor family in maize. BMC Plant Biol. 2018, 18, 235. [Google Scholar] [CrossRef] [Green Version]
- Ambawat, S.; Sharma, P.; Yadav, N.R.; Yadav, R.C. MYB transcription factor genes as regulators for plant responses: An overview. Physiol. Mol. Biol. Plants 2013, 19, 307–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raffaele, S.; Vailleau, F.; Leger, A.; Joubes, J.; Miersch, O.; Huard, C.; Blee, E.; Mongrand, S.; Domergue, F.; Roby, D. A MYB transcription factor regulates very-long-chain fatty acid biosynthesis for activation of the hypersensitive cell death response in Arabidopsis. Plant Cell 2008, 20, 752–767. [Google Scholar] [CrossRef] [Green Version]
- Vannini, C.; Iriti, M.; Bracale, M.; Locatelli, F.; Faoro, F.; Croce, P.; Pirona, R.; Di Maro, A.; Coraggio, I.; Genga, A. The ectopic expression of the rice Osmyb4 gene in Arabidopsis increases tolerance to abiotic, environmental and biotic stresses. Physiol. Mol. Plant Pathol. 2006, 69, 26–42. [Google Scholar] [CrossRef]
- Shulga, O.A.; Nedoluzhko, A.V.; Shchennikova, A.V.; Gruzdeva, N.M.; Shelenkov, A.A.; Sharko, F.S.; Sokolov, A.S.; Pantiukh, E.S.; Rastorguev, S.M.; Prokhortchouk, E.B.; et al. Profiling of microRNAs in wild type and early flowering transgenic Chrysanthemum morifolium by deep sequencing. Plant Cell Tissue Organ Cult. (PCTOC) 2016, 128, 283–301. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Transcriptome id | Days Post Inoculation | Number of Reads | Number of Transcripts | Median Transcript Length |
|---|---|---|---|---|
| 3h | 3 | 94,646,812 | 51,956 | 800 |
| 3i | 3 | 100,708,366 | 54,781 | 846 |
| Gene ID | log2(FoldChange) | Gene Annotation |
|---|---|---|
| 3 dpi | ||
| Csa_3G105950 | −9.1 | MTP-3 metal tolerance protein type 3-like |
| Csa_6G191570 | 8.1 | Histone H4 |
| Csa_2G030610 | 8 | Probable histone H2A.5 |
| Csa_4G290220 | 7.9 | Histone H3.2 |
| Csa_1G612925 | 7.7 | Histone H3.2 |
| Csa_2G429030 | 7.2 | Benzylalcohol O-benzoyl transferase |
| Csa_2G176690 | −7.2 | PRP1 proline-rich protein |
| Csa_4G001950 | −7.1 | Sucrose synthase 2-like |
| Csa_6G084600 | −7 | Uncharacterized |
| Csa_7G064050 | 6.9 | Uncharacterized |
| 20 dpi | ||
| Csa_6G448740 | −7.8 | Threonine dehydratase biosynthetic, chloroplastic |
| Csa_4G154320 | −7.3 | Polygalacturonase-inhibiting protein |
| Csa_2G429030 | −6.7 | Benzylalcohol O-benzoyltransferase |
| Csa_3G778180 | −6.5 | Uncharacterized |
| Csa_4G308490 | −6.4 | Benzylalcohol O-benzoyltransferase |
| Csa_4G000820 | −6.3 | Uncharacterized |
| Csa_1G050360 | −6.3 | Malate synthase, glyoxysomal-like |
| Csa_2G176190 | 6.2 | Repetitive proline-rich cell wall protein 3-like |
| Csa_3G076580 | −6.2 | Plastid lipid- associated protein |
| Csa_2G287040 | 6.1 | Uncharacterized |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Slavokhotova, A.; Korostyleva, T.; Shelenkov, A.; Pukhalskiy, V.; Korottseva, I.; Slezina, M.; Istomina, E.; Odintsova, T. Transcriptomic Analysis of Genes Involved in Plant Defense Response to the Cucumber Green Mottle Mosaic Virus Infection. Life 2021, 11, 1064. https://doi.org/10.3390/life11101064
Slavokhotova A, Korostyleva T, Shelenkov A, Pukhalskiy V, Korottseva I, Slezina M, Istomina E, Odintsova T. Transcriptomic Analysis of Genes Involved in Plant Defense Response to the Cucumber Green Mottle Mosaic Virus Infection. Life. 2021; 11(10):1064. https://doi.org/10.3390/life11101064
Chicago/Turabian StyleSlavokhotova, Anna, Tatyana Korostyleva, Andrey Shelenkov, Vitalii Pukhalskiy, Irina Korottseva, Marina Slezina, Ekaterina Istomina, and Tatyana Odintsova. 2021. "Transcriptomic Analysis of Genes Involved in Plant Defense Response to the Cucumber Green Mottle Mosaic Virus Infection" Life 11, no. 10: 1064. https://doi.org/10.3390/life11101064
APA StyleSlavokhotova, A., Korostyleva, T., Shelenkov, A., Pukhalskiy, V., Korottseva, I., Slezina, M., Istomina, E., & Odintsova, T. (2021). Transcriptomic Analysis of Genes Involved in Plant Defense Response to the Cucumber Green Mottle Mosaic Virus Infection. Life, 11(10), 1064. https://doi.org/10.3390/life11101064