1. Introduction
The size of our pupils changes continuously in response to variations in ambient light levels to regulate the amount of light entering our eyes. This process is known as the pupillary light reflex (PLR) [
1]. The PLR, while considered an example of a basic neurological reflex, is not a simple reflex as its function is modulated by cognitive brain function and interaction with the superior colliculus (SC), pretectal olivary nucleus (PON), and locus coeruleus that regulate the PLR [
2]. The PLR has probably been observed since the beginning of time and has become a standard examination tool used by all health care professionals after it was first described by Rhazes of Baghdad in the ninth century [
3].
It is essential to understand that the PLR is not a linear reaction of the pupil to light because pupil size is dramatically confounded by modest changes in attention, accommodation, and environmental ambient light [
4]. The attentional state of an individual, along with the size, luminance, and eccentricity of a stimulus, contributes to what was previously considered a simple reflex [
5]. The PLR has been widely used to evaluate the activity of the autonomic nervous system [
6]. Interestingly, constriction and dilation of the pupil may be more sensitive than other measures of autonomic function, with constriction dominated by the parasympathetic pathway and pupillary dilation dominated by the sympathetic system [
7].
There are gender differences in autonomic function that are important clinically. Interestingly, the iris muscle is modulated by light stimulation activation of the parasympathetic and sympathetic nervous system resulting in higher parasympathetic and lower sympathetic activity in females compared to males, similar to the cardiovascular system [
8]. This is due to central processing as a constant amount of light stimulation at a maintained luminance results in a different PLR response for males and females. PLR function is also correlated with other tests of autonomic function. For instance, heart rate variability (HRV) is significantly correlated with PLR variables for evaluating the activity of the autonomic nervous system with the most robust measures of parasympathetic activity [
9].
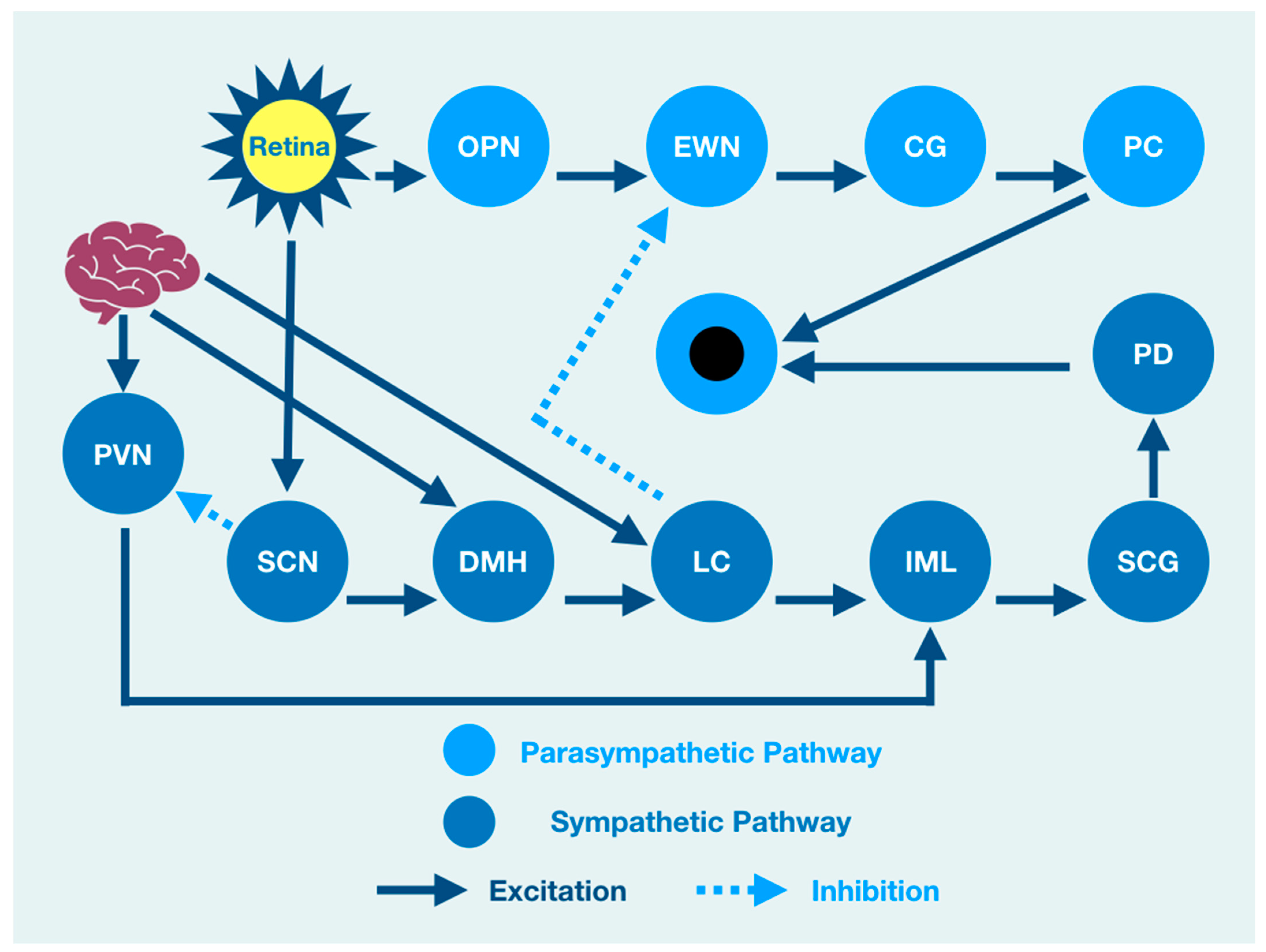
The PLR represents a complexity of integration at many brain, brainstem, and spinal cord levels such that it should not be considered as a simple response to a light source stimulation. There is, therefore, a need to understand the neurophysiology associated with brain function that might modulate the PLR. When light enters the eye, it is immediately converted to electrical impulses by specialized retinal rod and cone cells in the neuroretina [
10,
11,
12]. These impulses are integrated by six types of neuronal cells in a layered functional unit of photoreceptors and glial cells that form the optic nerve [
13]. The nasal retinal fibers of each eye contribute to the optic nerve and cross at the optic chiasm joining temporal retinal fibers from the other eye where they continue as the optic tract to the visual cortex of the occipital lobe at the back of the brain [
14]. The optic tract gives off fibers that synapse and excite the mesencephalic olivary pretectal nucleus (OPN) [
15,
16], which is excited by light from the retina but is also modulated by output from the cerebral cortex [
2]. OPN discharge results in bilateral activation of the Edinger Westphal nucleus (EWN), a preganglionic parasympathetic pupil constricting center [
17,
18]. While the OPN excites the EWN, the locus coeruleus (LC), the principal synthesizer of norepinephrine, inhibits the EWN. Its output is the result of a combination of excitation and inhibition [
19,
20,
21,
22].
The resultant discharge rate of the EWN results in pupil constriction because of its output and excitation of the ciliary ganglion (CG) that excites the pupiloconstrictor muscle [
18,
23]. The EWN excitation of the CG results from mathematical modeling of the direct activation of the EWN by the OPN and its direct inhibition by the LC [
24]. However, the LC output is modulated by multisynaptic activators from the brain and brainstem that affect the amount of pupil constriction via its effect on the EWN [
25]. A crucial clinical understanding of function is that pupil constriction is constantly changing in response to retinal activation by a light source. However, at the same time, the PLR is under the influence of other areas of the nervous system that are independent of the light source [
22,
26]. As such, the function of the brain affects or modulates the size of the pupil and its reaction to light, even though many of the structures that influence the PLR are not associated with the light-mediated PLR. For instance, cognitive processes that are often impaired after traumatic brain injuries are central to the prefrontal cortex (PFC) function, which modulates the PLR [
27].
Brain injury changes affect the PLR latency of its activation, pupil size, the velocity of constriction and dilation, and recovery times. The brain excites the paraventricular nucleus (PVN), which excites the sympathetic intermediolateral cell nucleus (IML) in the spinal cord [
28]. This cascade of events is such that the IML activates the suprachiasmatic nucleus (SCN), which activates the pupillodilator muscle (PD), causing pupil dilation. At the same time, the IML also receives excitation from the LC, which is itself excited by the cerebral cortex [
29]. From a functional viewpoint, cortical lesions of function or pathology may affect both parasympathetic pupillary constriction via a change in activation of the LC, resulting in changes of EWN inhibition and sympathetic pupillary dilation via IML activation.
All of this activity is happening at the same time such that the central role of the brain in modulating the PLR is further confounded because the LC is also activated by the dorsomedial hypothalamus (DMH), which is activated by the cerebral cortex, affecting both parasympathetic and sympathetic influences of the PLR [
2]. While functional brain connectivity drives the integration of target areas involved in the PLR, light activation of the retina is associated with direct activation of the OPN and the SCN that excites the DMH while inhibiting the PVN excitation of the IML. At the same time, the DMH is activated by the cerebral cortex that also activates the DMH target (LC) that excites the IML. While the DMH can be considered a part of the sympathetic pupil constriction pathway to the IML because it activates the LC excitation of the IML, it is also under the influence of the SCN that excites it while at the same time inhibiting the PVN activation of the IML [
30,
31].
An overview cartoon of the integrated PLR demonstrates the functional clinical integration of brain, brainstem, and spinal cord structures that contribute to pupil size after light stimulation (
Figure 1).
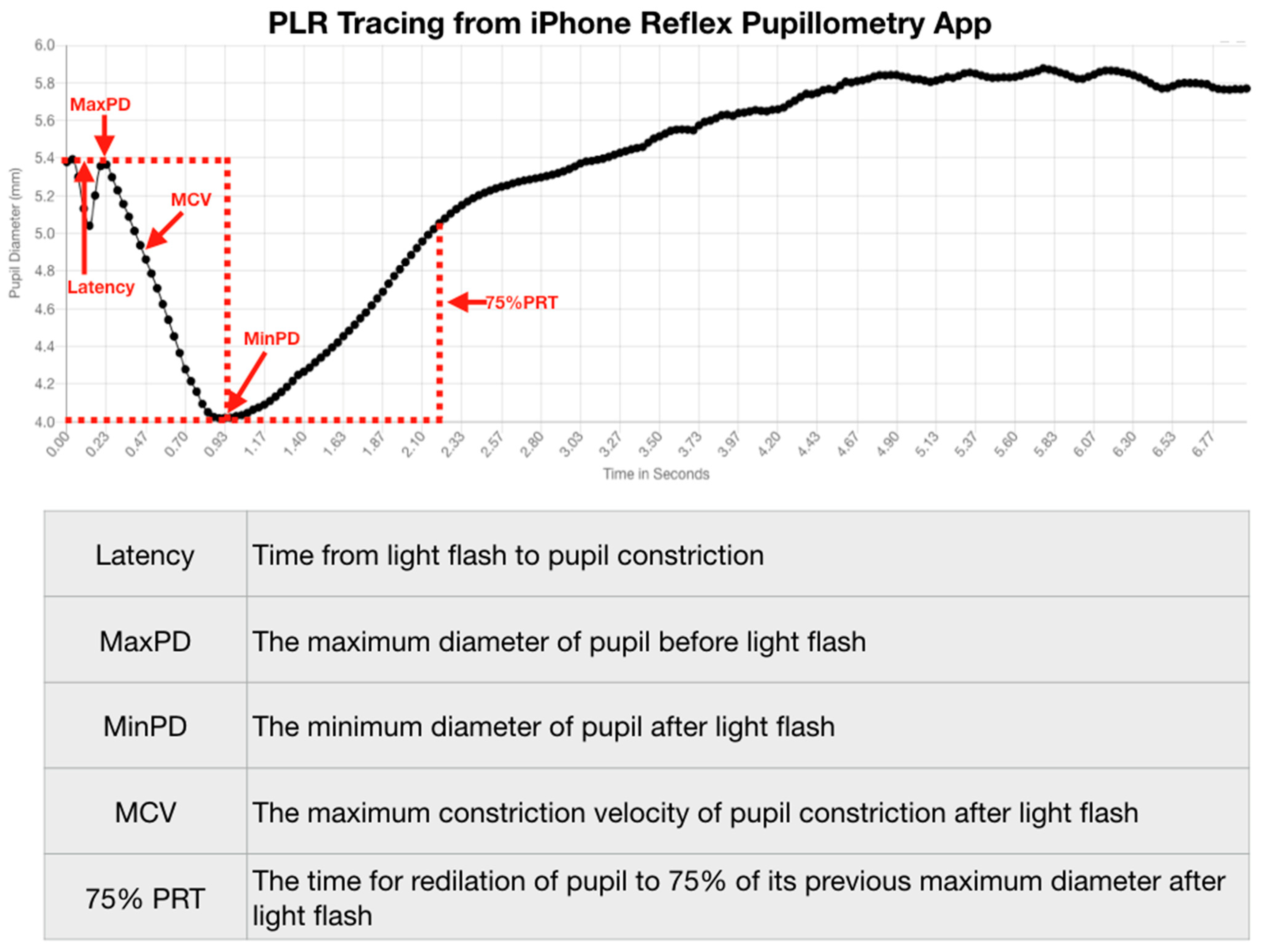
The PLR represents much more than a simple neurological reflex. Pupillometry represents a diagnostic technology that provides a reliable quantitative evaluation of the PLR that is reproducible [
32]. Reproducibility of a diagnostic test is necessary if clinicians desire to use the test to quantify function over time. Fortunately, the assessment of pupil function has been demonstrated to be reliable and vital in the evaluation of all patients, including neurocritical patients [
33]. The value of the PLR is, therefore, more than a simple quantification of a reflex. Even acute brain-injured patients will benefit from quantitative pupillometry for various brain functions, including delirium [
34].
The global incidence of traumatic brain injury (TBI) represents 64–74 million cases annually, resulting in one of the world’s most significant incidences of morbidity and mortality [
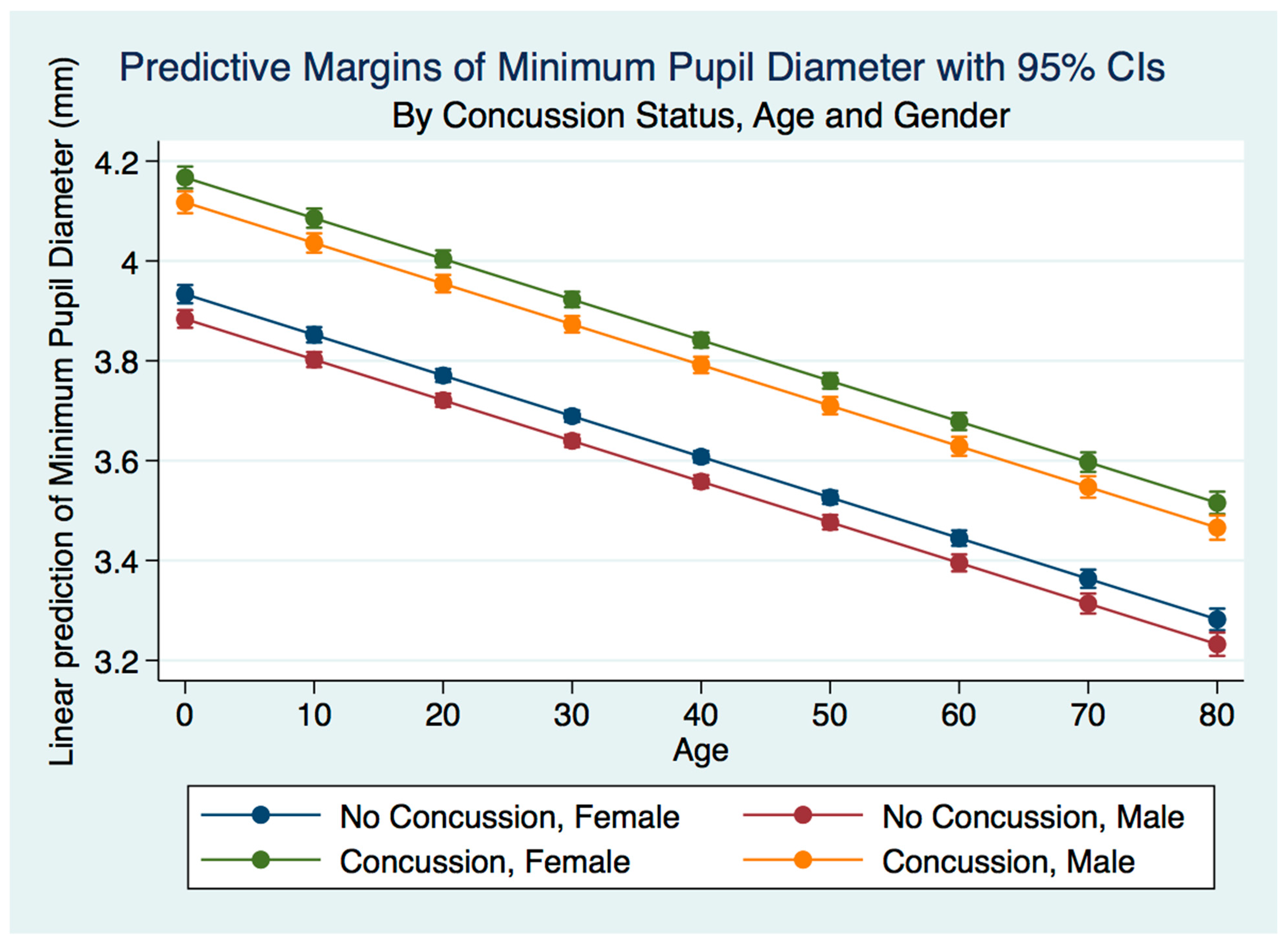
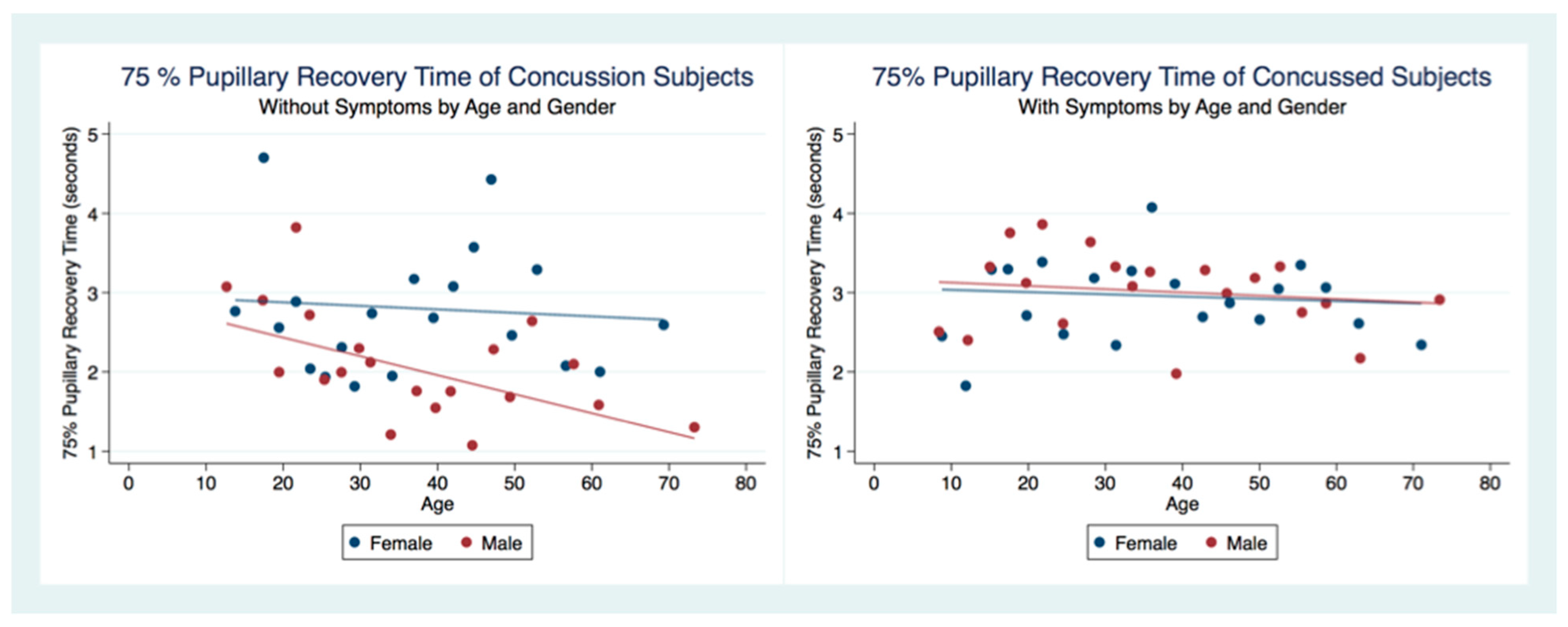
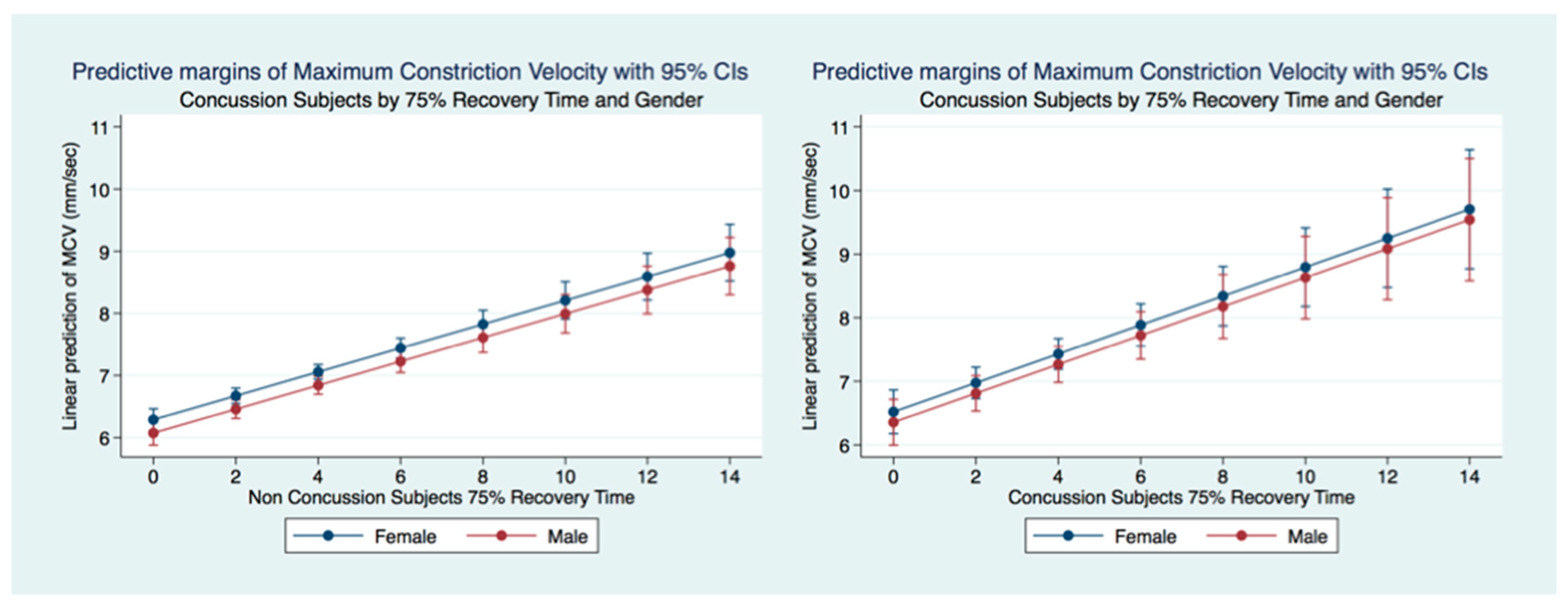
35]. We wondered whether people that have had a concussion with and without symptoms might have functional brain changes that the PLR might measure. As the PLR is modulated by central brain function, any long-term changes in brain function secondary to injury should cause a change in the parameters of the PLR compared to non-concussion subjects. We hypothesized that there would be differences in PLR components between concussion and non-concussion subjects that also would manifest in both gender and age effects.
4. Discussion
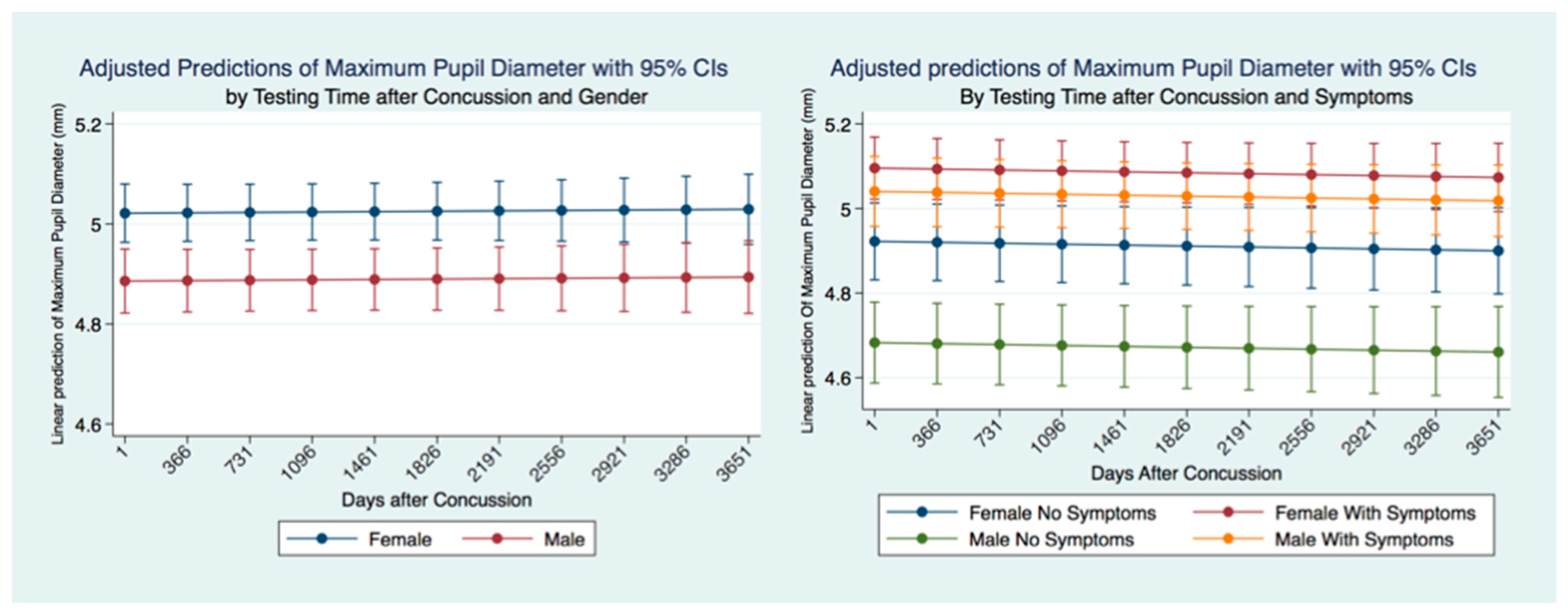
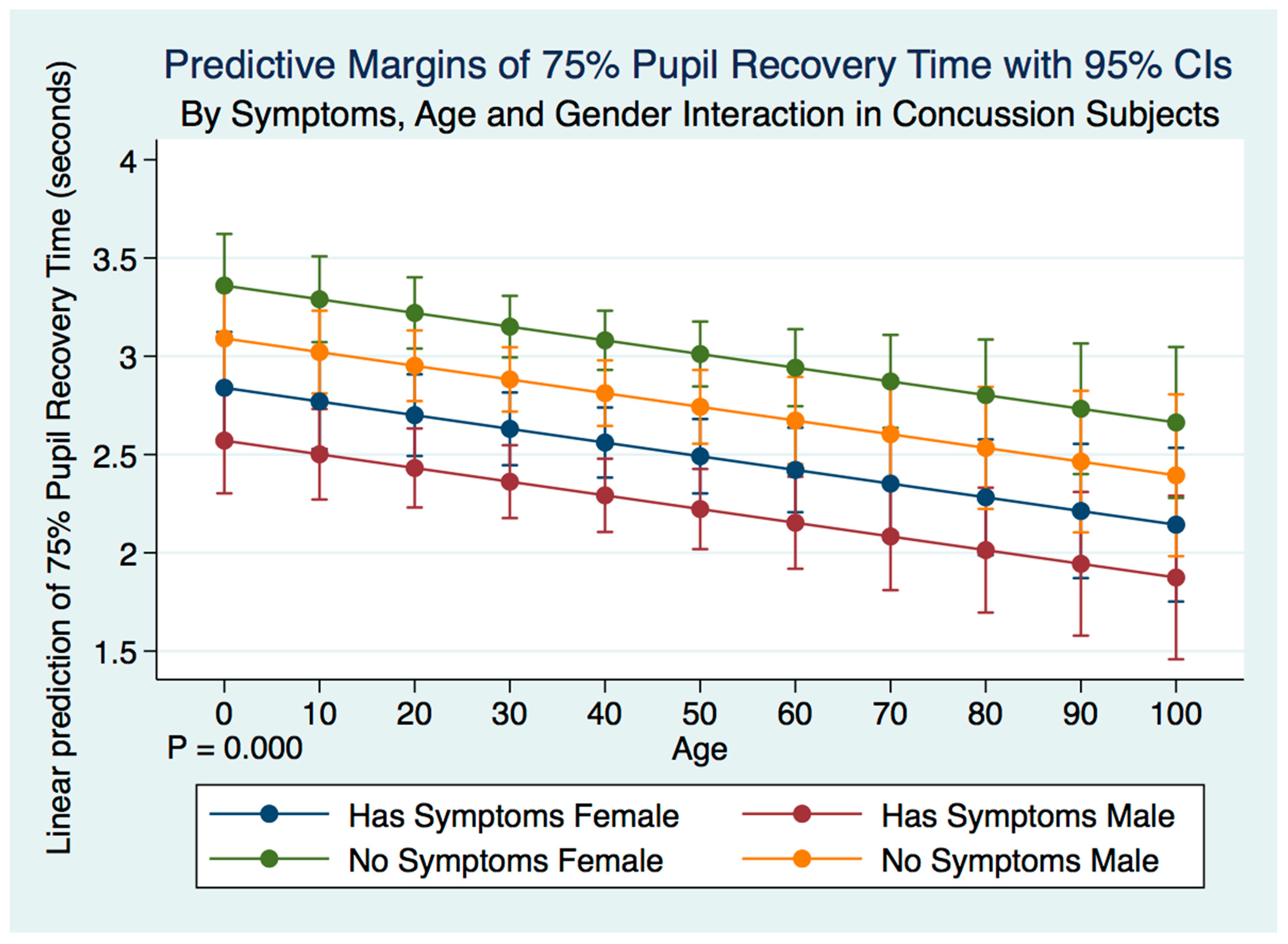
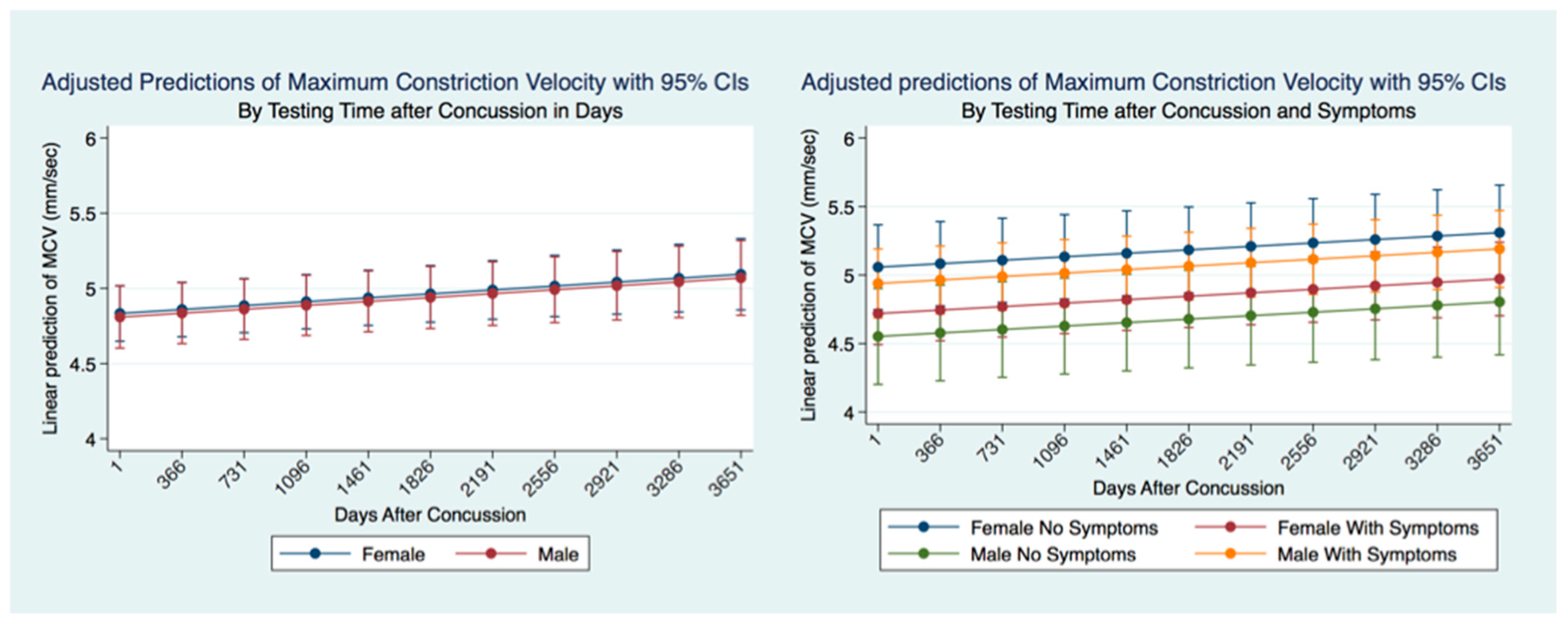
This investigation has provided information about the pupil’s reaction to light in subjects that have suffered a concussion compared to those that have not. The differences between subjects in all metrics of the PLR after concussion contribute to a greater understanding of the functional integration of the brain when it is injured. These functional differences are pronounced when considering the PLR function between genders. While most subjects that suffer a concussion will become symptom-free in a relatively short period [
37], it is clear from our findings that the lack of symptoms after a concussion does not represent a return to typical PLR metrics. This is alarming, as it appears that functional brain states in individuals who have suffered a concussion might be similar to those associated with other neurodegenerative disorders such as Alzheimer’s and Parkinson’s disease [
38]. These observations also suggest that the PLR might be used as a diagnostic biomarker of the success or failure of treatment strategies.
The PLR is often considered a simple reflex representative of a quantum of light evoking Cartesian pathways, resulting in a change of pupil diameter. As such, clinicians have been concerned that changes in ambient light observed in different settings might make a baseline measurement of pupil size impossible in anything but laboratory settings that might control for environmental light. However, our findings suggest that light is but one of many integration variables that govern the pupil’s size. Even the magnitude of the PLR in a talking parrot, while affected by light, is modulated more by the attention required for vocalization or recognition of known words by humans [
39].
Our findings show that the variables associated with brain function that modulates the PLR might, as in the parrot, be more critical in pupillary functional reaction to light than the light source. The differences in the PLR after concussion support this, and we were able to observe trends in changes of PLR metrics in non-laboratory controlled ambient lighting situations that mirrored the observations of others in a controlled laboratory setting [
40]. The reality of this situation translates to the ability to use the PLR in non-traditional settings such as on the sporting field, in the gym, clinic, or hospital wards without great concern for the control of ambient environmental light.
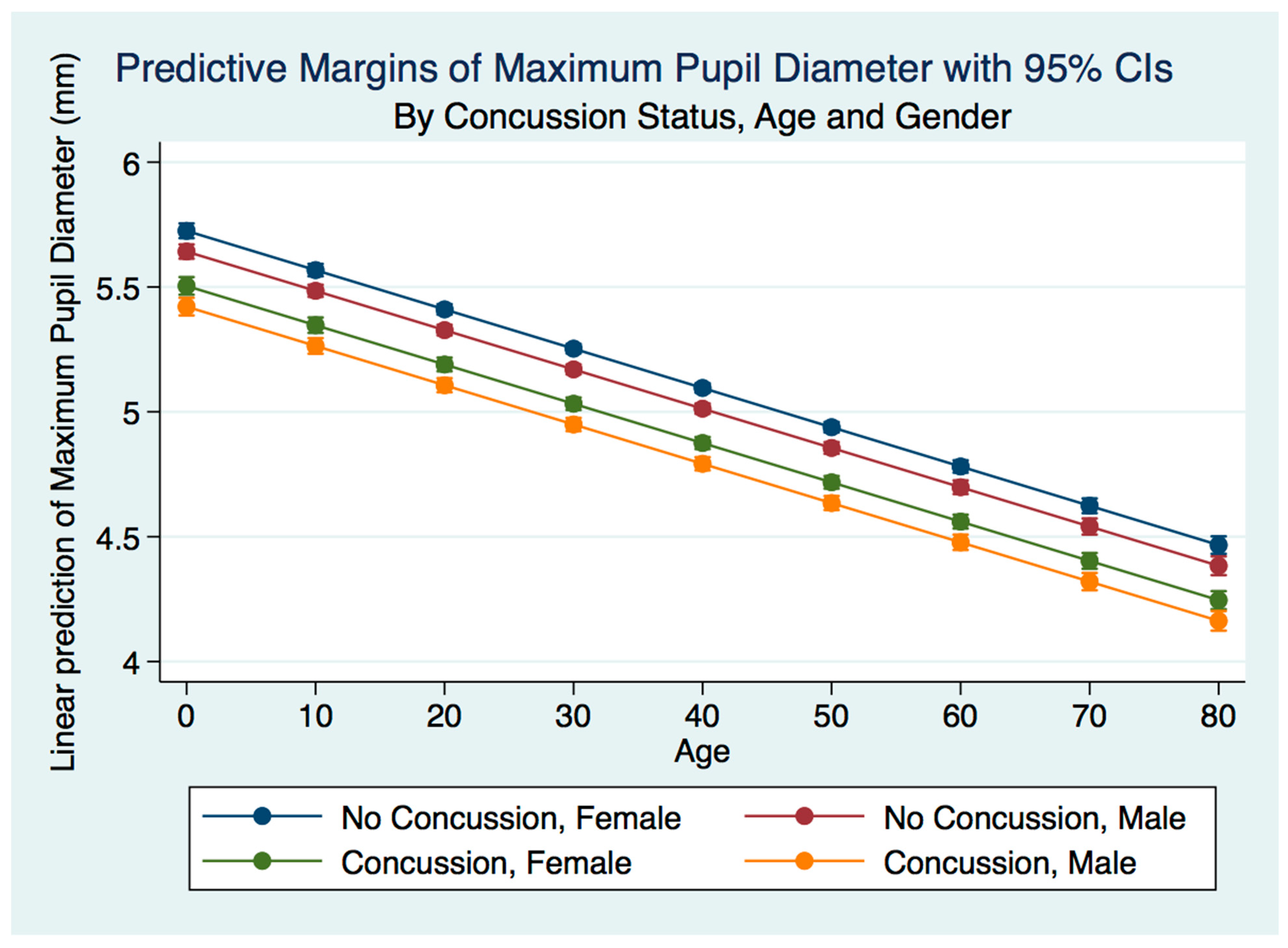
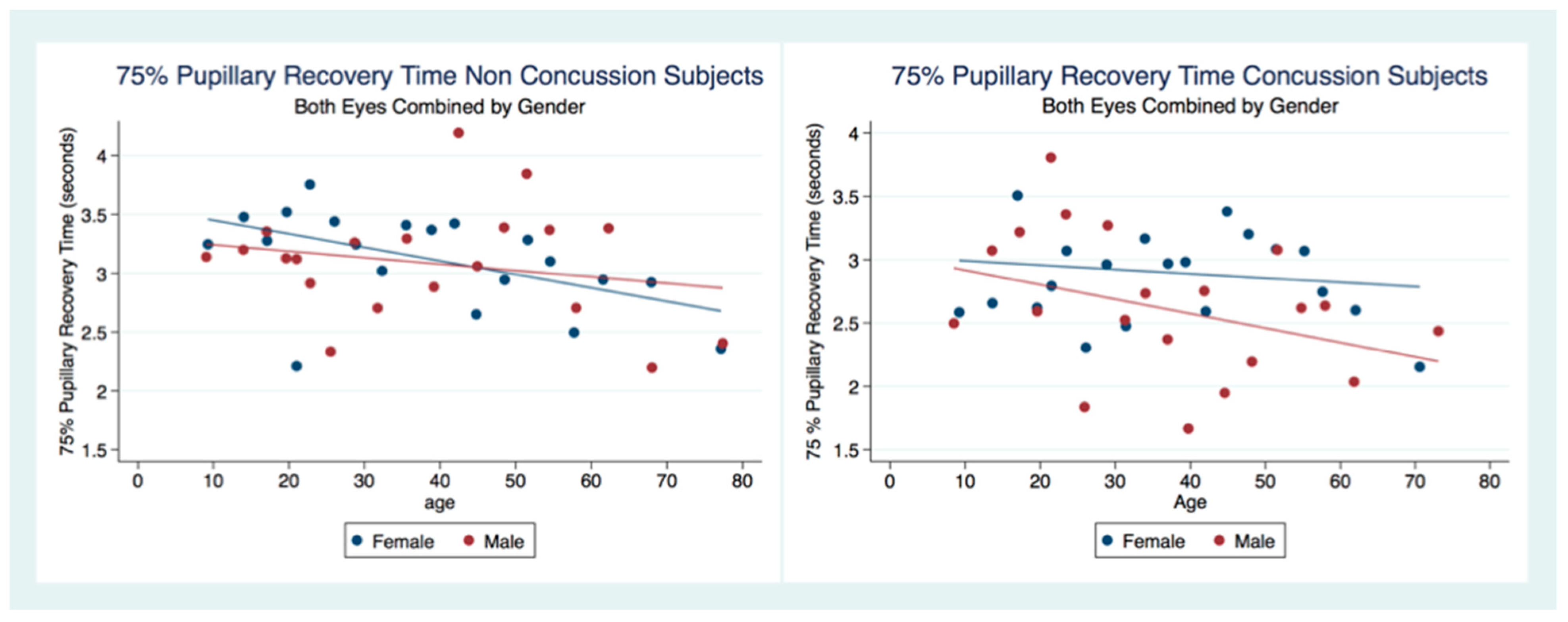
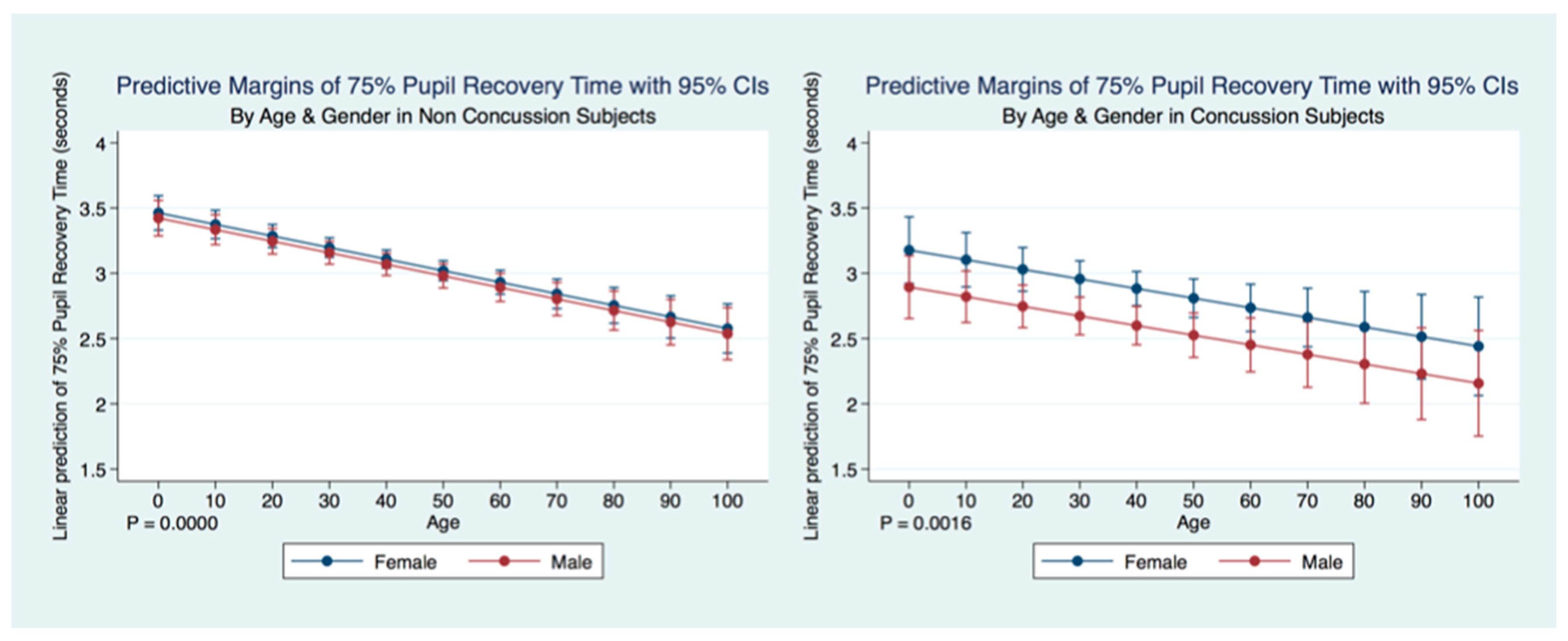
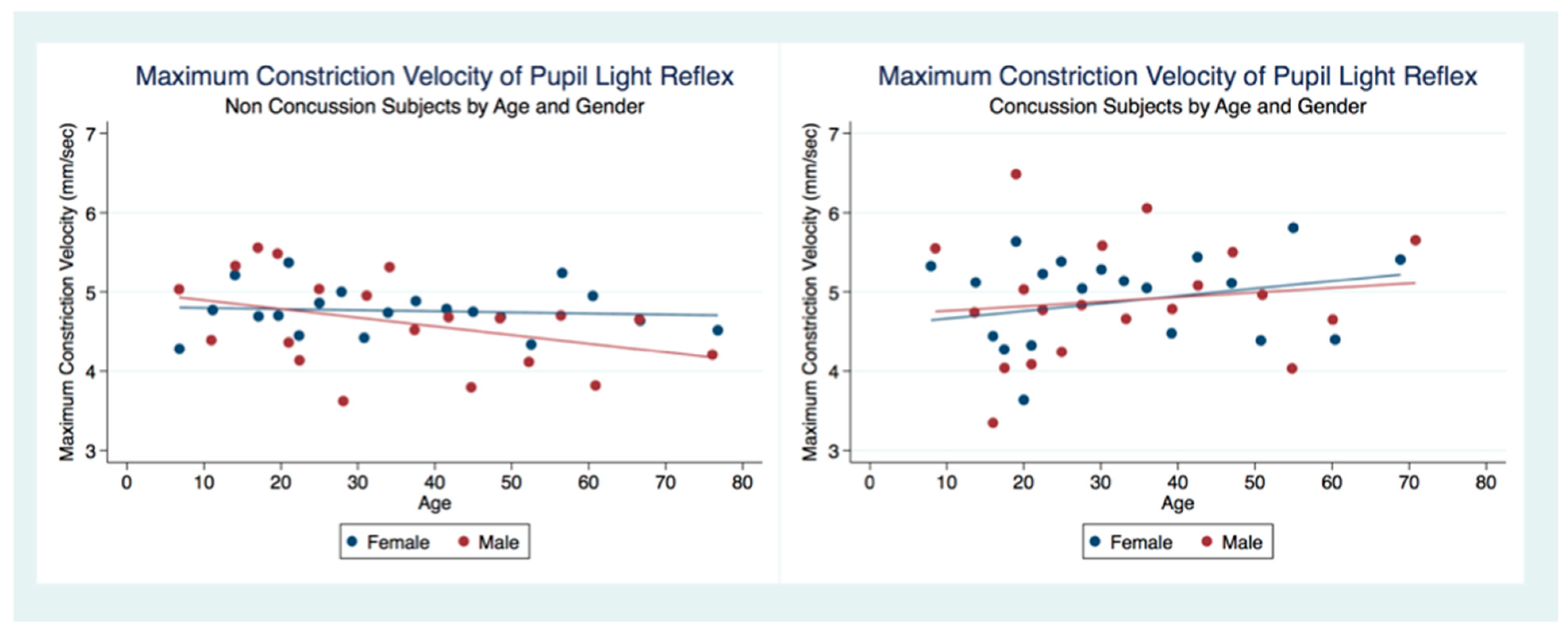
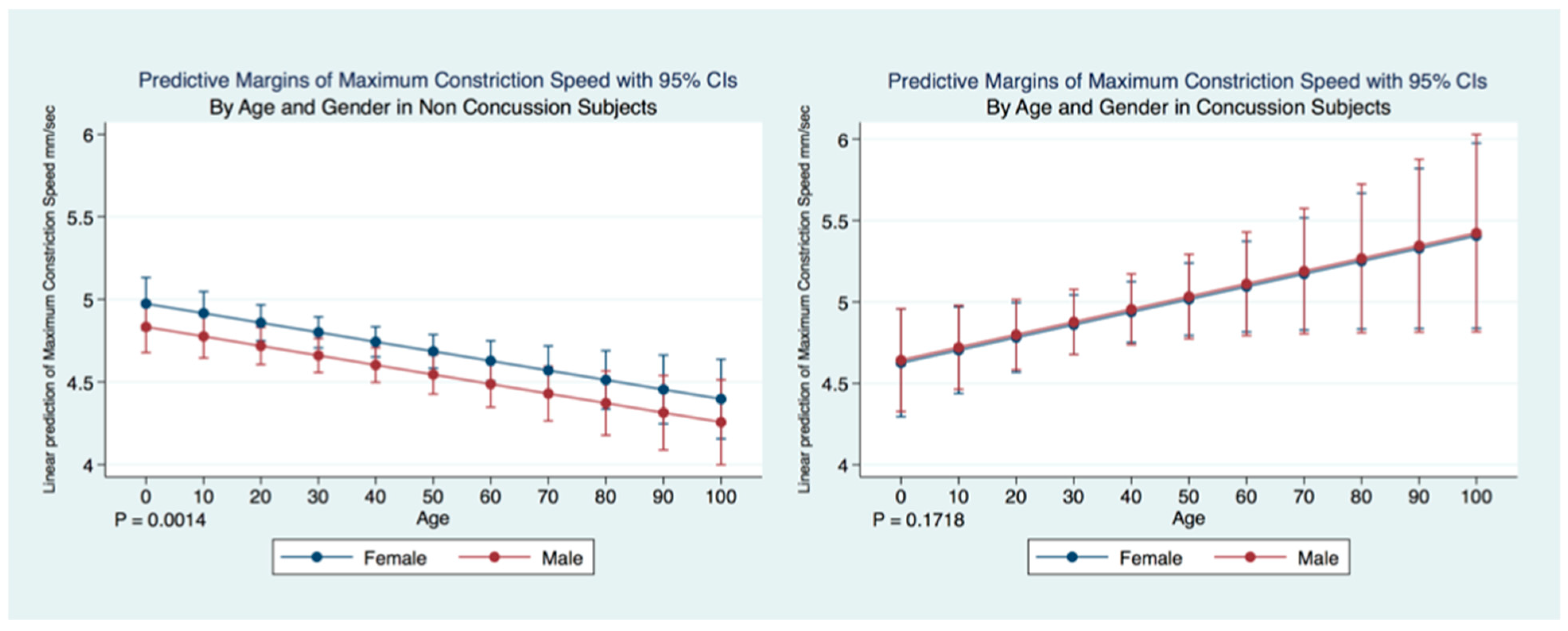
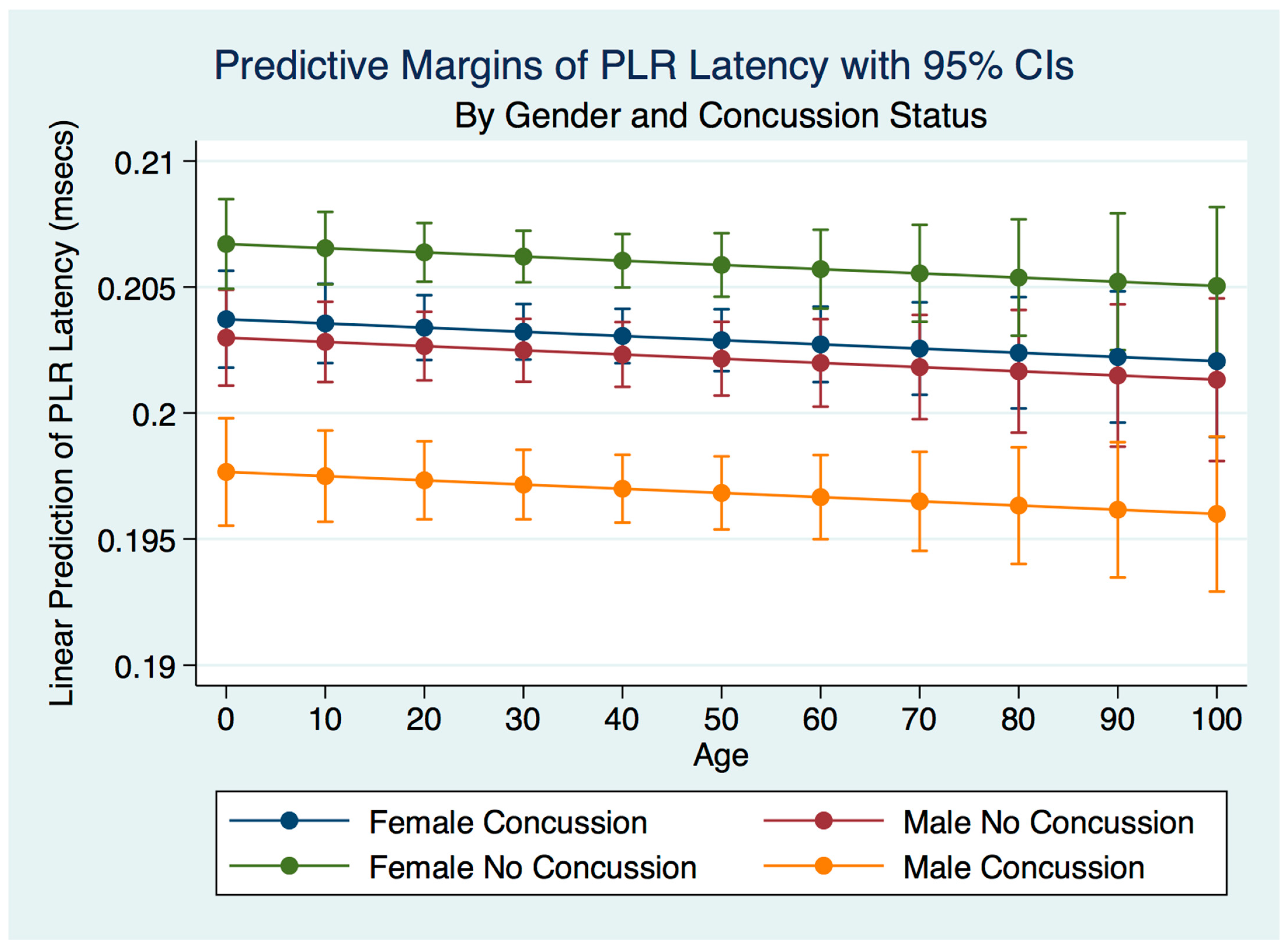
We have demonstrated that the PLR is different between genders, and that it changes with age. Fortunately, normative age-related data have been calculated for the PLR and can be used as a baseline for patient evaluation [
40]. We demonstrated that head injuries affect the PLR differently across age groups, complementing the work of others that have noted that adolescents demonstrate visual disorders with autonomic concomitants that are easily identified by the PLR after concussion [
41]. The complexity of cognitive disorders and neurodevelopmental syndromes also have identifiable PLR functions. Patients with attention deficit disorder (ADD) have a greater pupillary diameter and a faster maximum constriction velocity than normal age-matched controls and older subjects [
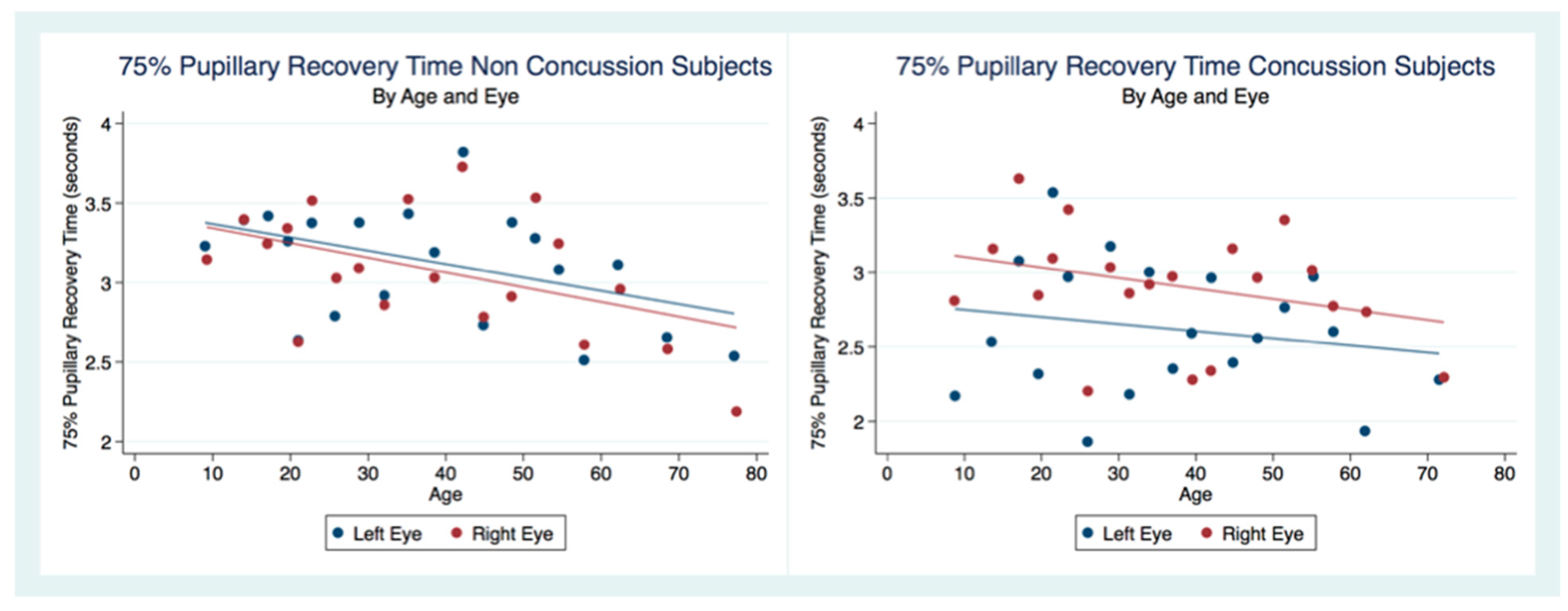
42]. Our findings have delineated the changes in pupillary diameter and constriction velocity controlled by gender, age and the presence or absence of concussion symptoms. We have also demonstrated that the latency of the PLR may serve as a sensitive biomarker that also changes with age. For example, the time delay between a light stimulation and the latency of the PLR is increased in older subjects (46–78 years) compared to younger subjects (18–45 years old) [
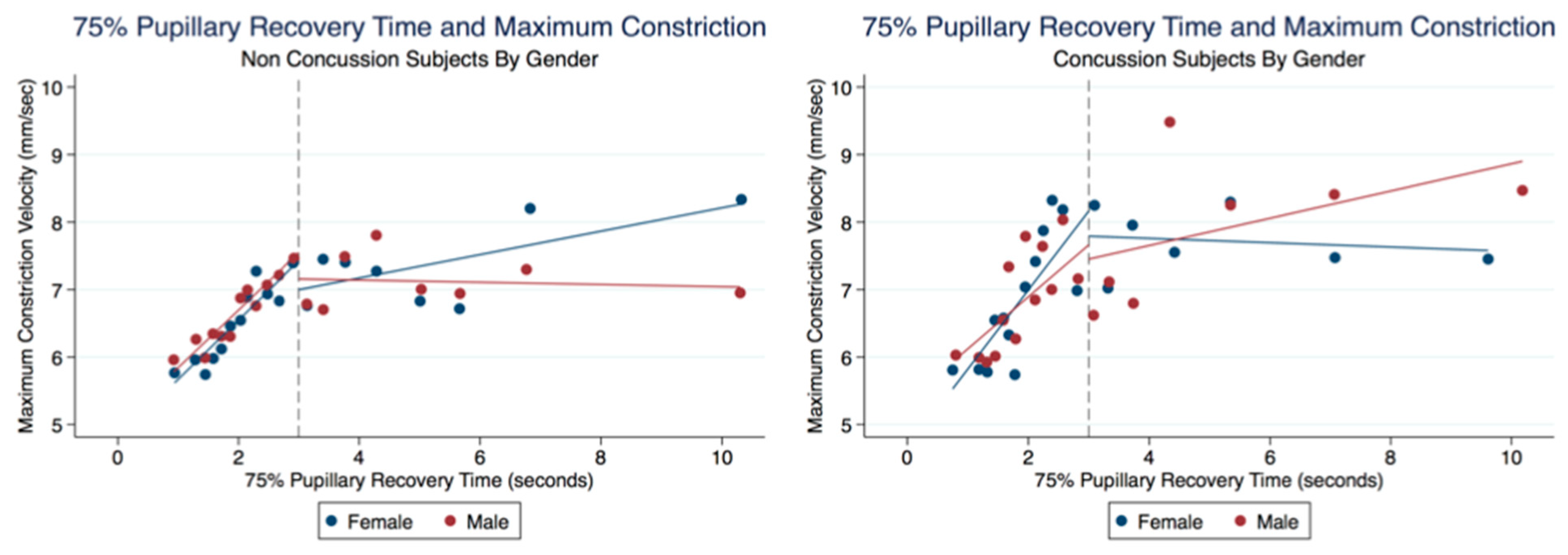
43]. We have demonstrated that brain injury changes affect the PLR latency of its activation, pupil size, the velocity of constriction and dilation, and recovery times.
Advances in technology have improved the accuracy of diagnostic instruments and the portability and ease of their use. We have demonstrated that the size of the pupil and its reaction to light can be reliably and reproducibly quantified by an advanced portable instrument using an iPhone medical app for clinical use [
44,
45]. Furthermore, this quantitative pupillometry has provided us with immediate evaluation and management of traumatic brain-injured patients over time and should contribute to decreasing the associated morbidity and mortality of these injuries [
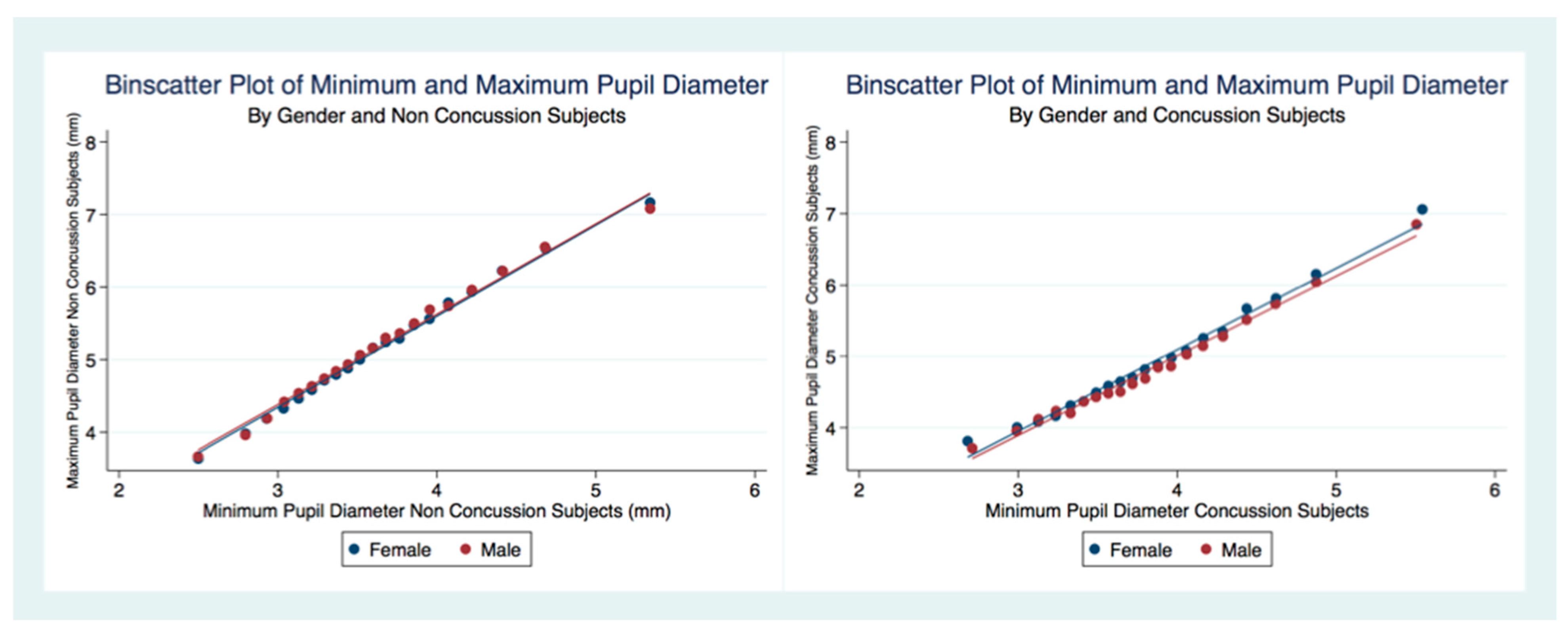
46]. We have provided reliable biomarkers that identify athletes with a concussion because of their larger maximum and minimum pupil diameter compared to healthy controls [
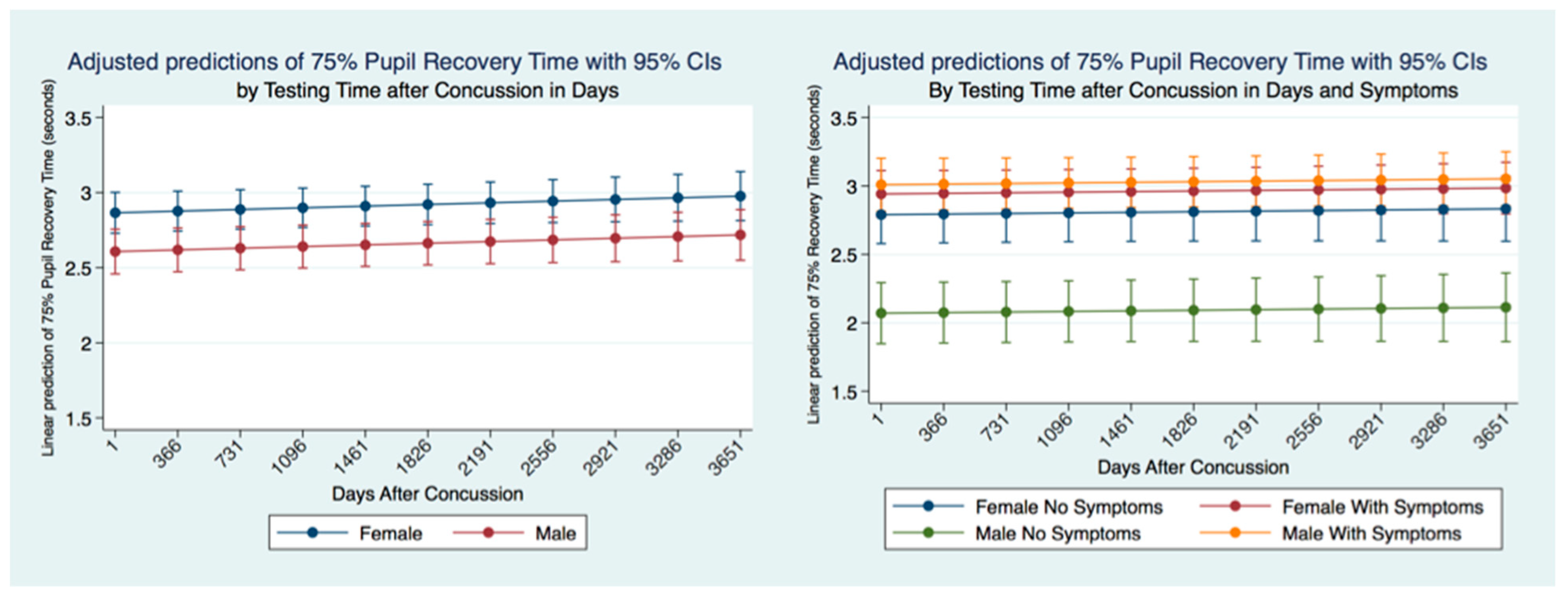
47]. Most importantly we have quantified the long-term metrics of the PLR in individuals that have suffered concussions. Most people that suffer from a concussion have a resolution of their symptoms within a short time of days to weeks. At the same time, a smaller number maintain their symptoms and are classified as suffering from post-concussion syndrome (PCS) [
48]. We have demonstrated the apparent permanency of functional changes of the PLR after concussion. These life-long changes in neurological function are supported by recent advances in quantification of increased ventricular size and decreased white matter measurements apparent later in life after a concussion [
49].


 ,
, 





















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}