
The Inhibitory Effect of Noscapine on the In Vitro Cathepsin G-Induced Collagen Expression in Equine Endometrium

 ,
,  , , ,
, , ,  and
and 
Abstract
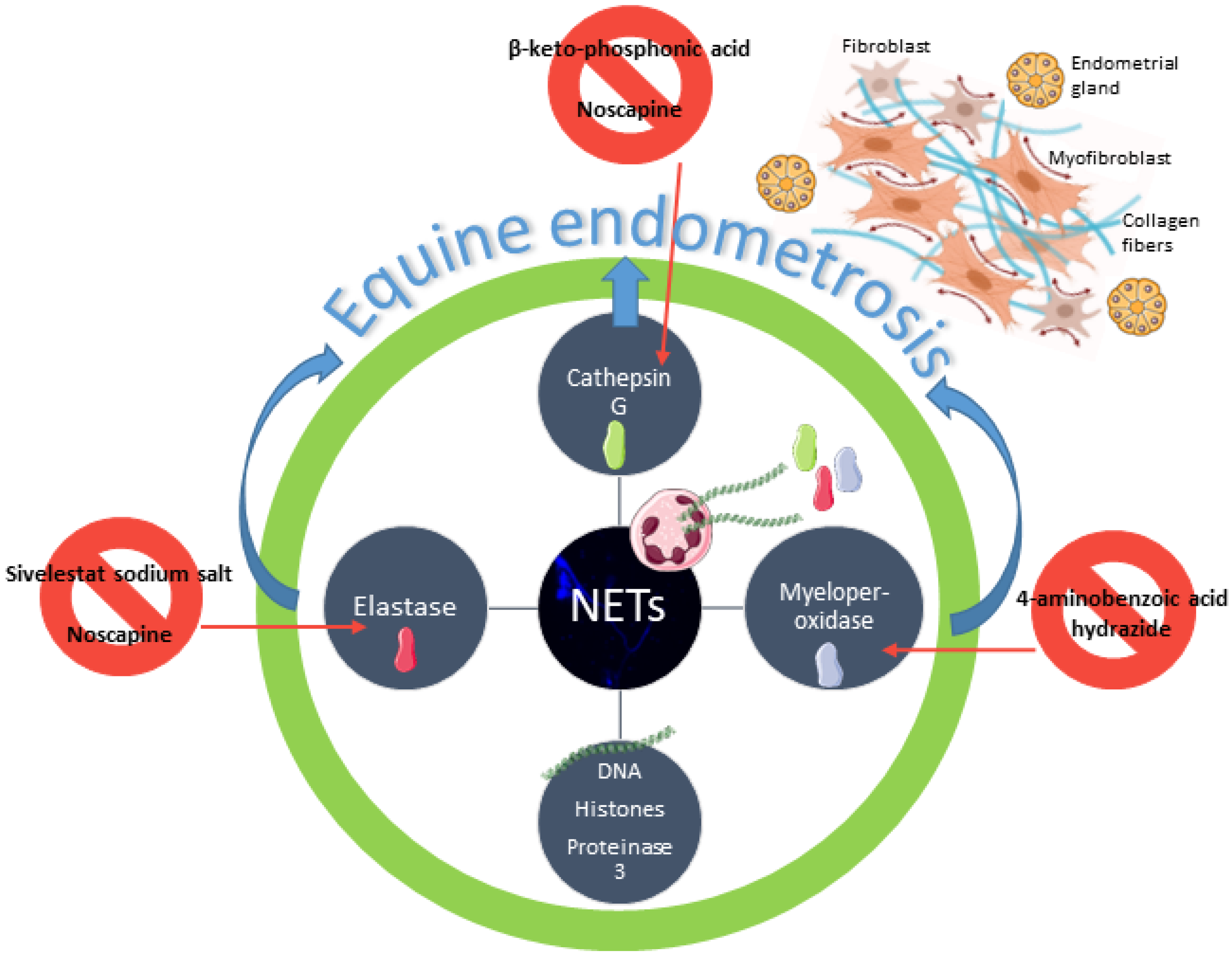
:1. Introduction
2. Materials and Methods
2.1. Mares
2.2. In Vitro Culture of Mare Endometrial Explants
2.3. Viability of Equine Endometrial Explants
2.4. Determination of COL1A2 mRNA Transcription by Real-Time Polymerase Chain Reaction (qPCR)
2.5. Quantification of COL1 Protein Relative Abundance by Western Blot
2.6. Statistical Analysis
3. Results
3.1. Evaluation of Equine Endometrial Explant Viability
3.2. The Isolated Effect of CAT, NOSC, Time of Treatment, and Estrous Cycle Phase and Their Interaction Combinations
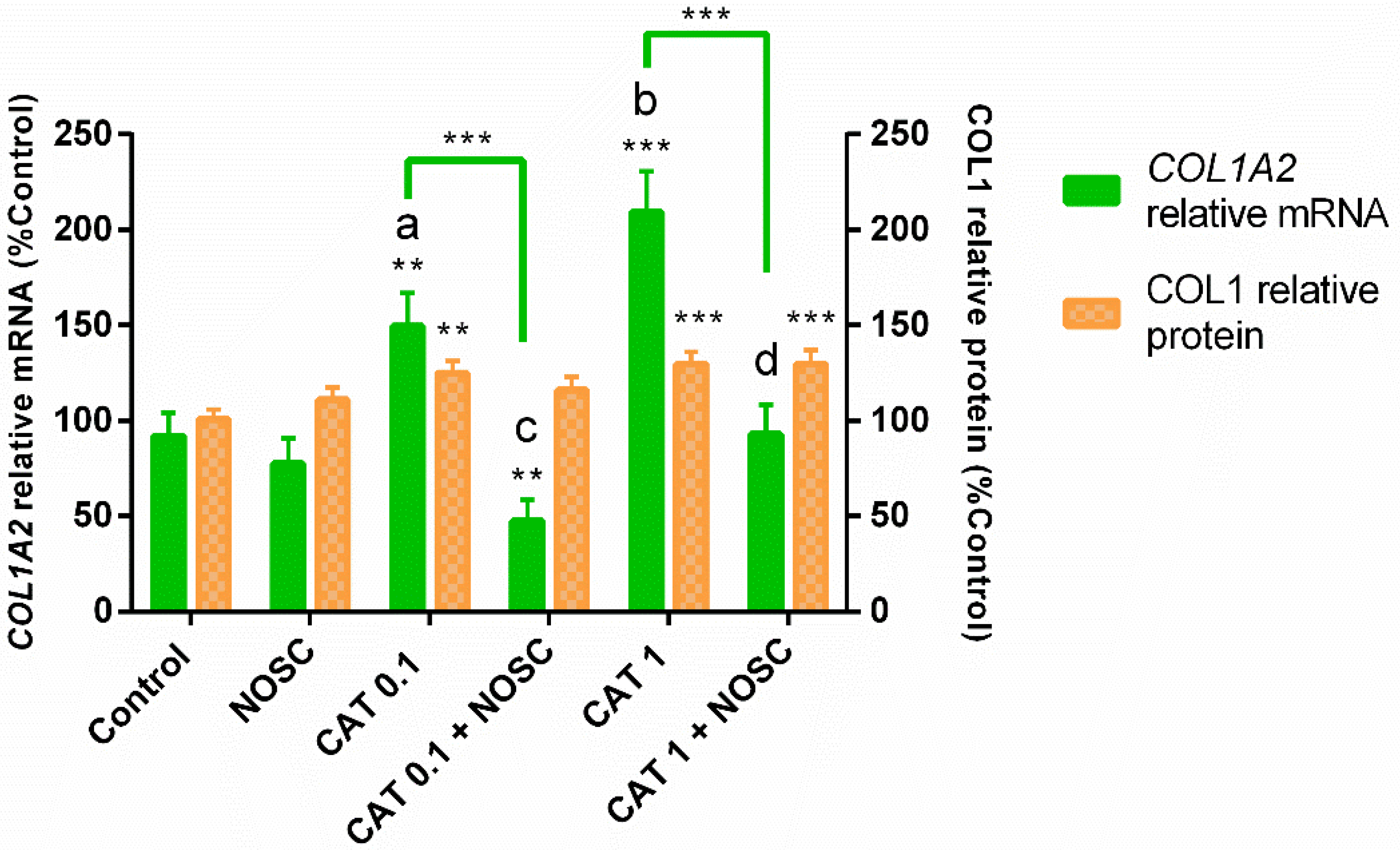
3.3. The Noscapine Inhibition of COL1 Induced by CAT Is Independent of Estrous Cycle Phase and Time of Treatment
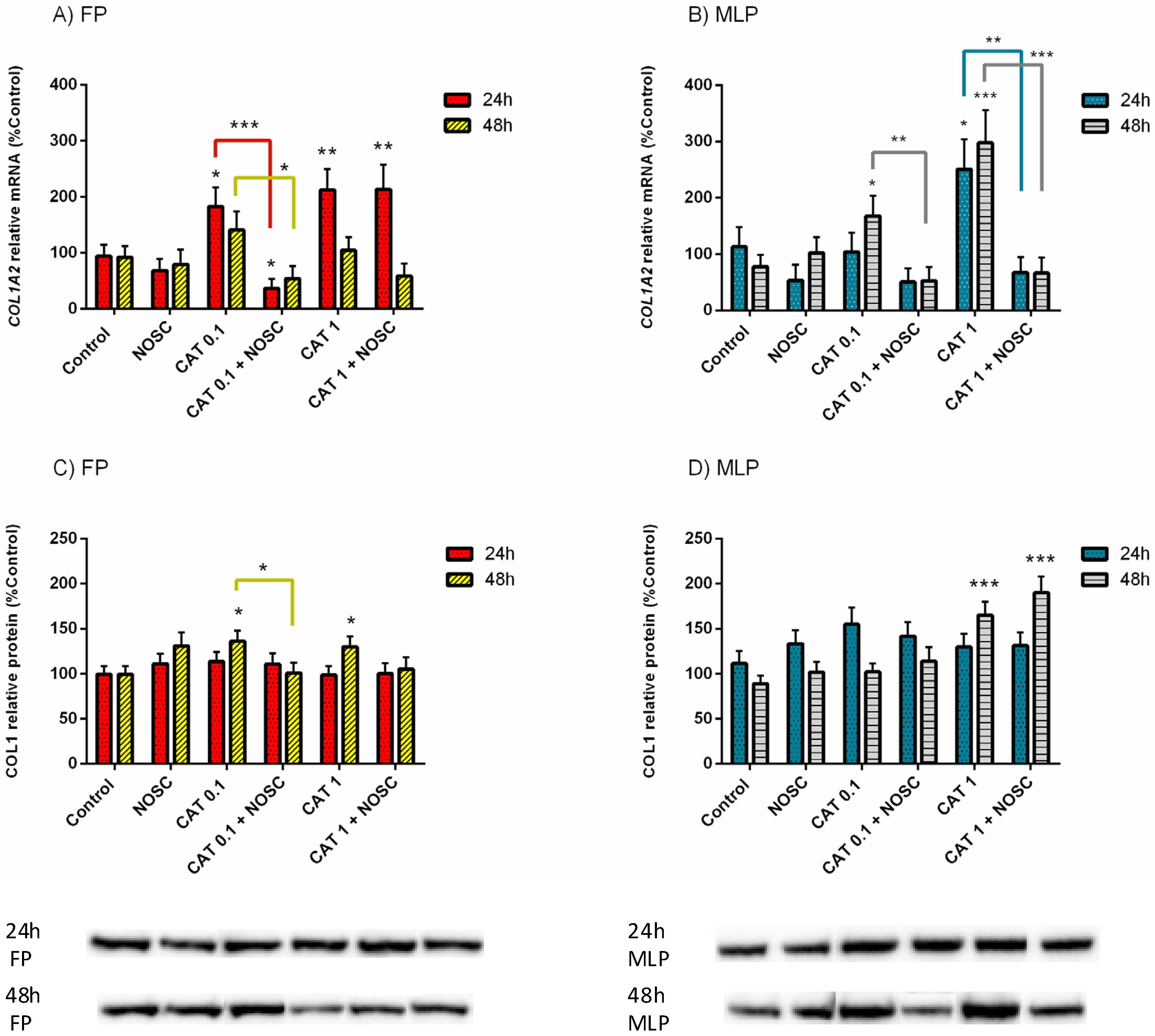
3.4. Noscapine Inhibition of CAT-Induced COL1 Expression Is Different Depending on the Estrous Cycle Phase and Treatment Time
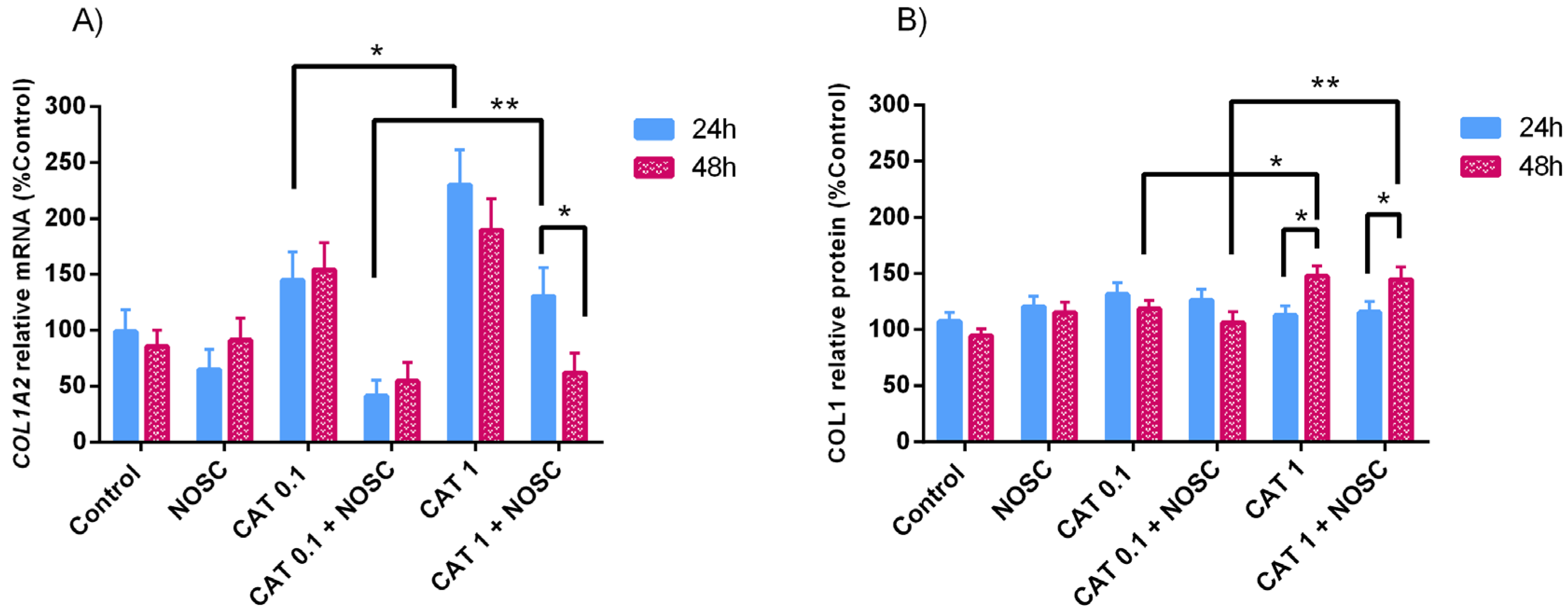
3.5. Noscapine Inhibition on CAT-Induced COL1 Expression Is Dependent of Treatment Time
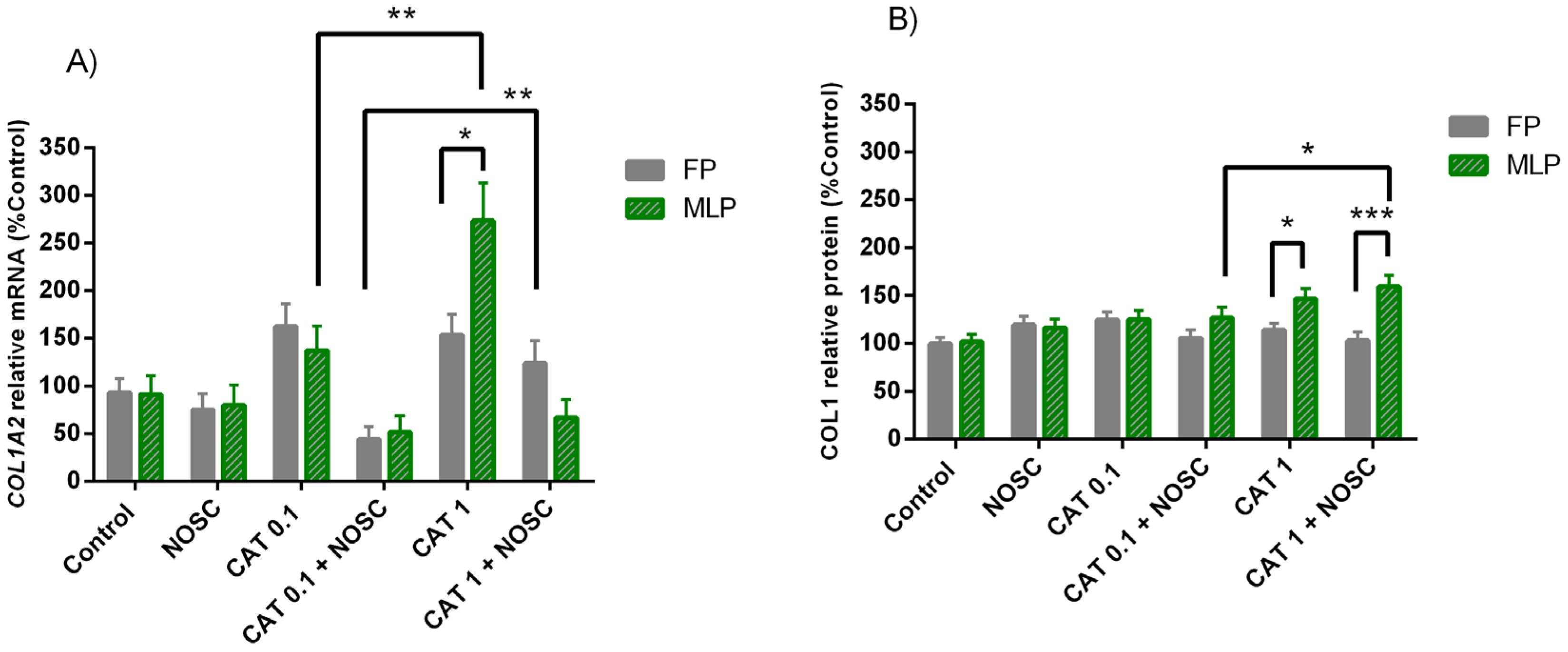
3.6. The Noscapine Inhibition on CAT-Induced COL1 Expression Is Dependent on Estrous Cycle Phase
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brinkmann, V. Neutrophil Extracellular Traps Kill Bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef]
- Averhoff, P.; Kolbe, M.; Zychlinsky, A.; Weinrauch, Y. Single Residue Determines the Specificity of Neutrophil Elastase for Shigella Virulence Factors. J. Mol. Biol. 2008, 377, 1053–1066. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Tu, J.; Hu, Y.; Song, G.; Yin, Z. Cathepsin G Cleaves and Activates IL-36γ and Promotes the Inflammation of Psoriasis. Drug Des. Devel. Ther. 2019, 13, 581–588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrmann, S.-M.; Funke-Kaiser, H.; Schmidt-Petersen, K.; Nicaud, V.; Gautier-Bertrand, M.; Evans, A.; Kee, F.; Arveiler, D.; Morrison, C.; Orzechowski, H.-D.; et al. Characterization of Polymorphic Structure of Cathepsin G Gene: Role in Cardiovascular and Cerebrovascular Diseases. Arterioscler. Thromb. Vasc. Biol. 2001, 21, 1538–1543. [Google Scholar] [CrossRef]
- Bhojak, T.J.; DeKosky, S.T.; Ganguli, M.; Kamboh, M.I. Genetic Polymorphism in the Cathepsin G Gene and the Risk of Alzheimer’s Disease. Neurosci. Lett. 2001, 309, 138–140. [Google Scholar] [CrossRef]
- Pérez-Is, L.; Ocaña, M.G.; Montes, A.H.; Carton, J.A.; Álvarez, V.; Meana, Á.; Fierer, J.; Valle-Garay, E.; Asensi, V. The N125S Polymorphism in the Cathepsin G Gene (Rs45567233) Is Associated with Susceptibility to Osteomyelitis in a Spanish Population. PLoS ONE 2019, 14, e0220022. [Google Scholar] [CrossRef] [PubMed]
- Brehm, A.; Geraghty, P.; Campos, M.; Garcia-Arcos, I.; Dabo, A.J.; Gaffney, A.; Eden, E.; Jiang, X.; D’Armiento, J.; Foronjy, R. Cathepsin G Degradation of Phospholipid Transfer Protein (PLTP) Augments Pulmonary Inflammation. FASEB J. 2014, 28, 2318–2331. [Google Scholar] [CrossRef] [Green Version]
- Shimoda, N.; Fukazawa, N.; Nonomura, K.; Fairchild, R.L. Cathepsin G Is Required for Sustained Inflammation and Tissue Injury after Reperfusion of Ischemic Kidneys. Am. J. Pathol. 2007, 170, 930–940. [Google Scholar] [CrossRef] [Green Version]
- Twigg, M.S.; Brockbank, S.; Lowry, P.; FitzGerald, S.P.; Taggart, C.; Weldon, S. The Role of Serine Proteases and Antiproteases in the Cystic Fibrosis Lung. Mediat. Inflamm. 2015, 2015, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Guerra, M.; Frey, D.; Hagner, M.; Dittrich, S.; Paulsen, M.; Mall, M.A.; Schultz, C. Cathepsin G Activity as a New Marker for Detecting Airway Inflammation by Microscopy and Flow Cytometry. ACS Cent. Sci. 2019, 5, 539–548. [Google Scholar] [CrossRef] [Green Version]
- Gudmann, N.S.; Manon-Jensen, T.; Sand, J.M.B.; Diefenbach, C.; Sun, S.; Danielsen, A.; Karsdal, M.A.; Leeming, D.J. Lung Tissue Destruction by Proteinase 3 and Cathepsin G Mediated Elastin Degradation Is Elevated in Chronic Obstructive Pulmonary Disease. Biochem. Biophys. Res. Commun. 2018, 503, 1284–1290. [Google Scholar] [CrossRef]
- Tan, K.; Brasch, H.D.; van Schaijik, B.; Armstrong, J.R.; Marsh, R.W.; Davis, P.F.; Tan, S.T.; Itinteang, T. Expression and Localization of Cathepsins B, D, and G in Dupuytren’s Disease. Plast. Reconstr. Surg. Glob. Open 2018, 6, e1686. [Google Scholar] [CrossRef] [PubMed]
- Rebordão, M.R.; Amaral, A.; Lukasik, K.; Szóstek-Mioduchowska, A.; Pinto-Bravo, P.; Galvão, A.; Skarzynski, D.J.; Ferreira-Dias, G. Constituents of Neutrophil Extracellular Traps Induce in Vitro Collagen Formation in Mare Endometrium. Theriogenology 2018, 113, 8–18. [Google Scholar] [CrossRef]
- Amaral, A.; Fernandes, C.; Lukasik, K.; Szóstek-Mioduchowska, A.; Baclawska, A.; Rebordão, M.R.; Aguiar-Silva, J.; Pinto-Bravo, P.; Skarzynski, D.J.; Ferreira-Dias, G. Elastase Inhibition Affects Collagen Transcription and Prostaglandin Secretion in Mare Endometrium during the Estrous Cycle. Reprod. Domest. Anim. 2018, 53, 66–69. [Google Scholar] [CrossRef] [PubMed]
- Amaral, A.; Fernandes, C.; Rebordão, M.R.; Szóstek-Mioduchowska, A.; Lukasik, K.; Gawronska-Kozak, B.; Telo da Gama, L.; Skarzynski, D.J.; Ferreira-Dias, G. The In Vitro Inhibitory Effect of Sivelestat on Elastase Induced Collagen and Metallopeptidase Expression in Equine Endometrium. Animals 2020, 10, 863. [Google Scholar] [CrossRef]
- Amaral, A.; Fernandes, C.; Morazzo, S.; Rebordão, M.R.; Szóstek-Mioduchowska, A.; Lukasik, K.; Gawronska-Kozak, B.; Telo da Gama, L.; Skarzynski, D.J.; Ferreira-Dias, G. The Inhibition of Cathepsin G on Endometrial Explants with Endometrosis in the Mare. Front. Vet. Sci. 2020, 7, 582211. [Google Scholar] [CrossRef] [PubMed]
- Amaral, A.; Fernandes, C.; Rebordão, M.R.; Szóstek-Mioduchowska, A.; Lukasik, K.; Pinto-Bravo, P.; Telo da Gama, L.; Jan Skarzynski, D.; Ferreira-Dias, G. Myeloperoxidase Inhibition Decreases the Expression of Collagen and Metallopeptidase in Mare Endometria under In Vitro Conditions. Animals 2021, 11, 208. [Google Scholar] [CrossRef]
- Katila, T. Onset and Duration of Uterine Inflammatory Response of Mares after Insemination with Fresh Semen. Biol. Reprod. 1995, 52, 515–517. [Google Scholar] [CrossRef]
- Christoffersen, M.; Woodward, E.; Bojesen, A.M.; Jacobsen, S.; Petersen, M.R.; Troedsson, M.H.; Lehn-Jensen, H. Inflammatory Responses to Induced Infectious Endometritis in Mares Resistant or Susceptible to Persistent Endometritis. BMC Vet. Res. 2012, 8, 41. [Google Scholar] [CrossRef] [Green Version]
- Christoffersen, M.; Troedsson, M. Inflammation and Fertility in the Mare. Reprod. Domest. Anim. 2017, 52 (Suppl. 3), 14–20. [Google Scholar] [CrossRef]
- Canisso, I.F.; Segabinazzi, L.G.T.M.; Fedorka, C.E. Persistent Breeding-Induced Endometritis in Mares—A Multifaceted Challenge: From Clinical Aspects to Immunopathogenesis and Pathobiology. Int. J. Mol. Sci. 2020, 21, 1432. [Google Scholar] [CrossRef] [Green Version]
- Troedsson, M.H.T.; Liu, I.K.M.; Thurmond, M. Function of Uterine and Blood-Derived Polymorphonuclear Neutrophils in Mares Susceptible and Resistant to Chronic Uterine Infection: Phagocytosis and Chemotaxis1. Biol. Reprod. 1993, 49, 507–514. [Google Scholar] [CrossRef] [Green Version]
- LeBLANC, M.M.; Neuwirth, L.; Asbury, A.C.; Tran, T.; Mauragis, D.; Klapstein, E. Scintigraphic Measurement of Uterine Clearance in Normal Mares and Mares with Recurrent Endometritis. Equine Vet. J. 1994, 26, 109–113. [Google Scholar] [CrossRef] [PubMed]
- Rebordão, M.R.; Amaral, A.; Fernandes, C.; Silva, E.; Lukasik, K.; Szóstek-Mioduchowska, A.; Pinto-Bravo, P.; Galvão, A.; Skarzynski, D.J.; Ferreira-Dias, G. Enzymes Present in Neutrophil Extracellular Traps May Stimulate the Fibrogenic PGF2α Pathway in the Mare Endometrium. Animals 2021, 11, 2615. [Google Scholar] [CrossRef]
- Kenney, R.M. Cyclic and pathological changes of the mare endometrium as detected by biopsy, with a note on early embryonic death. J. Am. Vet. Med. Assoc. 1978, 172, 241–262. [Google Scholar] [PubMed]
- Hoffmann, C.; Ellenberger, C.; Mattos, R.C.; Aupperle, H.; Dhein, S.; Stief, B.; Schoon, H.-A. The Equine Endometrosis: New Insights into the Pathogenesis. Anim. Reprod. Sci. 2009, 111, 261–278. [Google Scholar] [CrossRef]
- Lehmann, J.; Ellenberger, C.; Hoffmann, C.; Bazer, F.W.; Klug, J.; Allen, W.R.; Sieme, H.; Schoon, H.-A. Morpho-Functional Studies Regarding the Fertility Prognosis of Mares Suffering from Equine Endometrosis. Theriogenology 2011, 76, 1326–1336. [Google Scholar] [CrossRef]
- Ley, W.B.; Bowen, J.M.; Sponenberg, D.P.; Lessard, P.N. Dimethyl Sulfoxide Intrauterine Therapy in the Mare: Effects upon Endometrial Histological Features and Biopsy Classification. Theriogenology 1989, 32, 263–276. [Google Scholar] [CrossRef]
- Buczkowska, J.; Kozdrowski, R.; Nowak, M.; Raś, A.; Mrowiec, J. Endometrosis—Significance for Horse Reproduction, Pathogenesis, Diagnosis, and Proposed Therapeutic Methods. Pol. J. Vet. Sci. 2014, 17, 547–554. [Google Scholar] [CrossRef] [Green Version]
- Podico, G.; Canisso, I.F.; Roady, P.J.; Austin, S.M.; Carossino, M.; Balasuriya, U.; Ellerbrock, R.E.; Lima, F.S.; Ferreira-Dias, G.; Douglas, R.H. Uterine Responses and Equine Chorionic Gonadotropin Concentrations after Two Intrauterine Infusions with Kerosene Post Early Fetal Loss in Mares. Theriogenology 2020, 147, 202–210. [Google Scholar] [CrossRef] [PubMed]
- Navarrete, F.; Saravia, F.; Cisterna, G.; Rojas, F.; Silva, P.P.; Rodríguez-Alvarez, L.; Rojas, D.; Cabezas, J.; Mançanares, A.C.F.; Castro, F.O. Assessment of the Anti-Inflammatory and Engraftment Potential of Horse Endometrial and Adipose Mesenchymal Stem Cells in an in Vivo Model of Post Breeding Induced Endometritis. Theriogenology 2020, 155, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Cabezas, J.; Rojas, D.; Wong, Y.; Telleria, F.; Manriquez, J.; Mançanares, A.C.F.; Rodriguez-Alvarez, L.; Castro, F.O. In Vitro Preconditioning of Equine Adipose Mesenchymal Stem Cells with Prostaglandin E2, Substance P and Their Combination Changes the Cellular Protein Secretomics and Improves Their Immunomodulatory Competence without Compromising Stemness. Vet. Immunol. Immunopathol. 2020, 228, 110100. [Google Scholar] [CrossRef]
- Karlsson, M.O.; Dahlström, B.; Eckernäs, S.A.; Johansson, M.; Alm, A.T. Pharmacokinetics of Oral Noscapine. Eur. J. Clin. Pharmacol. 1990, 39, 275–279. [Google Scholar] [CrossRef] [PubMed]
- Rida, P.C.G.; LiVecche, D.; Ogden, A.; Zhou, J.; Aneja, R. The Noscapine Chronicle: A Pharmaco-Historic Biography of the Opiate Alkaloid Family and Its Clinical Applications: THE NOSCAPINE CHRONICLE. Med. Res. Rev. 2015, 35, 1072–1096. [Google Scholar] [CrossRef] [Green Version]
- Esnaashari, S.S.; Muhammadnejad, S.; Amanpour, S.; Amani, A. A Combinational Approach Towards Treatment of Breast Cancer: An Analysis of Noscapine-Loaded Polymeric Nanoparticles and Doxorubicin. AAPS PharmSciTech 2020, 21, 166. [Google Scholar] [CrossRef]
- Ke, Y.; Ye, K.; Grossniklaus, H.E.; Archer, D.R.; Joshi, H.C.; Kapp, J.A. Noscapine Inhibits Tumor Growth with Little Toxicity to Normal Tissues or Inhibition of Immune Responses. Cancer Immunol. Immunother. 2000, 49, 217–225. [Google Scholar] [CrossRef]
- Aneja, R.; Dhiman, N.; Idnani, J.; Awasthi, A.; Arora, S.K.; Chandra, R.; Joshi, H.C. Preclinical Pharmacokinetics and Bioavailability of Noscapine, a Tubulin-Binding Anticancer Agent. Cancer Chemother. Pharmacol. 2007, 60, 831–839. [Google Scholar] [CrossRef]
- Newcomb, E.W.; Lukyanov, Y.; Smirnova, I.; Schnee, T.; Zagzag, D. Noscapine Induces Apoptosis in Human Glioma Cells by an Apoptosis-Inducing Factor-Dependent Pathway. Anti-Cancer Drugs 2008, 19, 553–563. [Google Scholar] [CrossRef]
- Liu, M.; Luo, X.-J.; Liao, F.; Lei, X.-F.; Dong, W.-G. Noscapine Induces Mitochondria-Mediated Apoptosis in Gastric Cancer Cells in Vitro and in Vivo. Cancer Chemother. Pharmacol. 2011, 67, 605–612. [Google Scholar] [CrossRef]
- Yang, Z.-R.; Liu, M.; Peng, X.-L.; Lei, X.-F.; Zhang, J.-X.; Dong, W.-G. Noscapine Induces Mitochondria-Mediated Apoptosis in Human Colon Cancer Cells in Vivo and in Vitro. Biochem. Biophys. Res. Commun. 2012, 421, 627–633. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Liu, M.; Huang, X.; Zhu, Q.; Liu, W.; Chen, W.; Zou, Y.; Cai, Y.; Huang, S.; Chen, A.; et al. Noscapine Induces Apoptosis in Human Colon Cancer Cells by Regulating Mitochondrial Damage and Warburg Effect via PTEN/PI3K/MTOR Signaling Pathway. Onco Targets Ther. 2020, 13, 5419–5428. [Google Scholar] [CrossRef] [PubMed]
- Idänpään-Heikkilä, J.E. Studies on the Fate of 3H-Noscapine in Mice and Rats. Ann. Med. Exp. Biol. Fenn. 1968, 46, 201–216. [Google Scholar] [PubMed]
- Lasagna, L.; Owens, A.H.; Shnider, B.I.; Gold, G.L. Toxicity after Large Doses of Noscapine. Cancer Chemother. Rep. 1961, 15, 33–34. [Google Scholar]
- Sebak, S.; Mirzaei, M.; Malhotra, M.; Kulamarva, A.; Prakash, S. Human Serum Albumin Nanoparticles as an Efficient Noscapine Drug Delivery System for Potential Use in Breast Cancer: Preparation and in Vitro Analysis. Int. J. Nanomed. 2010, 5, 525–532. [Google Scholar] [CrossRef] [Green Version]
- Kach, J.; Sandbo, N.; La, J.; Denner, D.; Reed, E.B.; Akimova, O.; Koltsova, S.; Orlov, S.N.; Dulin, N.O. Antifibrotic Effects of Noscapine through Activation of Prostaglandin E2 Receptors and Protein Kinase A. J. Biol. Chem. 2014, 289, 7505–7513. [Google Scholar] [CrossRef] [Green Version]
- Doddapaneni, R.; Patel, K.; Chowdhury, N.; Singh, M. Noscapine Chemosensitization Enhances Docetaxel Anticancer Activity and Nanocarrier Uptake in Triple Negative Breast Cancer. Exp. Cell Res. 2016, 346, 65–73. [Google Scholar] [CrossRef] [Green Version]
- Amaral, A.; Fernandes, C.; Szóstek-Mioduchowska, A.; Rebordão, M.R.; Skarzynski, D.J.; Ferreira-Dias, G. Noscapine Acts as a Protease Inhibitor of In Vitro Elastase-Induced Collagen Deposition in Equine Endometrium. Int. J. Mol. Sci. 2021, 22, 5333. [Google Scholar] [CrossRef]
- Roberto da Costa, R.P.; Ferreira-Dias, G.; Mateus, L.; Korzekwa, A.; Andronowska, A.; Platek, R.; Skarzynski, D.J. Endometrial Nitric Oxide Production and Nitric Oxide Synthases in the Equine Endometrium: Relationship with Microvascular Density during the Estrous Cycle. Domest. Anim. Endocrinol. 2007, 32, 287–302. [Google Scholar] [CrossRef]
- Rebordão, M.R.; Amaral, A.; Lukasik, K.; Szóstek-Mioduchowska, A.; Pinto-Bravo, P.; Galvão, A.; Skarzynski, D.J.; Ferreira-Dias, G. Impairment of the Antifibrotic Prostaglandin E2 Pathway May Influence Neutrophil Extracellular Traps–Induced Fibrosis in the Mare Endometrium. Domest. Anim. Endocrinol. 2019, 67, 1–10. [Google Scholar] [CrossRef]
- Kenney, R.M.; Doig, P.A. Equine endometrial biopsy. In Current Therapy in Theriogenology 2: Diagnosis, Treatment, and Prevention of Reproductive Diseases in Small and Large Animals, 2nd ed.; Morrow, D.A., Ed.; Saunders WB: Philadelphia, PA, USA, 1986; pp. 723–729. [Google Scholar]
- Schafer, W.R.; Fischer, L.; Roth, K.; Jullig, A.K.; Stuckenschneider, J.E.; Schwartz, P.; Weimer, M.; Orlowska-Volk, M.; Hanjalic-Beck, A.; Kranz, I.; et al. Critical Evaluation of Human Endometrial Explants as an Ex Vivo Model System: A Molecular Approach. Mol. Hum. Reprod. 2011, 17, 255–265. [Google Scholar] [CrossRef]
- Zhao, S.; Fernald, R.D. Comprehensive Algorithm for Quantitative Real-Time Polymerase Chain Reaction. J. Comput. Biol. 2005, 12, 1047–1064. [Google Scholar] [CrossRef]
- PREDICTED: Equus Caballus Collagen Type I Alpha 2 Chain (COL1A2), mRNA. Available online: https://www.ncbi.nlm.nih.gov/nucleotide/XM_001492939.4?report=genbank&log$=nuclalign&blast_rank=19&RID=NHF70AB501N (accessed on 9 October 2021).
- PREDICTED: Equus Caballus Ribosomal Protein L32 (RPL32), mRNA. Available online: https://www.ncbi.nlm.nih.gov/nucleotide/XM_001492042.6?report=genbank&log$=nuclalign&blast_rank=1&RID=NHFCEW9V01N (accessed on 9 October 2021).
- Posch, A.; Kohn, J.; Oh, K.; Hammond, M.; Liu, N. V3 Stain-Free Workflow for a Practical, Convenient, and Reliable Total Protein Loading Control in Western Blotting. J. Vis. Exp. 2013, 50948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szóstek-Mioduchowska, A.Z.; Lukasik, K.; Skarzynski, D.J.; Okuda, K. Effect of Transforming Growth Factor -Β1 on α-Smooth Muscle Actin and Collagen Expression in Equine Endometrial Fibroblasts. Theriogenology 2019, 124, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Szóstek-Mioduchowska, A.Z.; Baclawska, A.; Okuda, K.; Skarzynski, D.J. Effect of Proinflammatory Cytokines on Endometrial Collagen and Metallopeptidase Expression during the Course of Equine Endometrosis. Cytokine 2019, 123, 154767. [Google Scholar] [CrossRef]
- Szóstek-Mioduchowska, A.Z.; Baclawska, A.; Rebordão, M.R.; Ferreira-Dias, G.; Skarzynski, D.J. Prostaglandins Effect on Matrix Metallopeptidases and Collagen in Mare Endometrial Fibroblasts. Theriogenology 2020, 153, 74–84. [Google Scholar] [CrossRef]
- Ganjam, V.K.; McLeod, C.; Klesius, P.H.; Washburn, S.M.; Kwapien, R.; Brown, B.; Fazeli, M.H. Effect of Ovarian Hormones on the Phagocytic Response of Ovariectomized Mares. J. Reprod. Fertil. Suppl. 1982, 32, 169–174. [Google Scholar]
- Kumar, N.; Sood, D.; van der Spek, P.J.; Sharma, H.S.; Chandra, R. Molecular Binding Mechanism and Pharmacology Comparative Analysis of Noscapine for Repurposing against SARS-CoV-2 Protease. J. Proteome Res. 2020, 19, 4678–4689. [Google Scholar] [CrossRef]
- Zughaier, S.; Karna, P.; Stephens, D.; Aneja, R. Potent Anti-Inflammatory Activity of Novel Microtubule-Modulating Brominated Noscapine Analogs. PLoS ONE 2010, 5, e9165. [Google Scholar] [CrossRef] [PubMed]
- Khanmoradi, M.; Ali Mard, S.; Aboutaleb, N.; Nobakht, M.; Mahmoudian, M. The Protective Activity of Noscapine on Renal Ischemia-Reperfusion Injury in Male Wistar Rat. Iran. J. Basic Med. Sci. 2014, 17, 244–249. [Google Scholar]
- Quisbert-Valenzuela, E.O.; Calaf, G.M. Apoptotic Effect of Noscapine in Breast Cancer Cell Lines. Int. J. Oncol. 2016, 48, 2666–2674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aneja, R.; Zhou, J.; Zhou, B.; Chandra, R.; Joshi, H.C. Treatment of Hormone-Refractory Breast Cancer: Apoptosis and Regression of Human Tumors Implanted in Mice. Mol. Cancer Ther. 2006, 5, 2366–2377. [Google Scholar] [CrossRef] [Green Version]
- Altinoz, M.A.; Topcu, G.; Hacimuftuoglu, A.; Ozpinar, A.; Ozpinar, A.; Hacker, E.; Elmaci, İ. Noscapine, a Non-Addictive Opioid and Microtubule-Inhibitor in Potential Treatment of Glioblastoma. Neurochem. Res. 2019, 44, 1796–1806. [Google Scholar] [CrossRef]
- Landen, J.W.; Hau, V.; Wang, M.; Davis, T.; Ciliax, B.; Wainer, B.H.; Van Meir, E.G.; Glass, J.D.; Joshi, H.C.; Archer, D.R. Noscapine Crosses the Blood-Brain Barrier and Inhibits Glioblastoma Growth. Clin. Cancer Res. 2004, 10, 5187–5201. [Google Scholar] [CrossRef] [Green Version]
- Sandbo, N.; Ngam, C.; Torr, E.; Kregel, S.; Kach, J.; Dulin, N. Control of Myofibroblast Differentiation by Microtubule Dynamics through a Regulated Localization of MDia2. J. Biol. Chem. 2013, 288, 15466–15473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sung, B.; Ahn, K.S.; Aggarwal, B.B. Noscapine, a Benzylisoquinoline Alkaloid, Sensitizes Leukemic Cells to Chemotherapeutic Agents and Cytokines by Modulating the NF-ΚB Signaling Pathway. Cancer Res. 2010, 70, 3259–3268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chougule, M.B.; Patel, A.R.; Jackson, T.; Singh, M. Antitumor Activity of Noscapine in Combination with Doxorubicin in Triple Negative Breast Cancer. PLoS ONE 2011, 6, e17733. [Google Scholar] [CrossRef] [PubMed]
- Chandra, R. Noscapine as Anticancer Agent & Its Role in Ovarian Cancer. Org. Med. Chem. Int. J. 2019, 9, 48–56. [Google Scholar] [CrossRef]
- Dong, J.; Ma, Q. In Vivo Activation and Pro-Fibrotic Function of NF-ΚB in Fibroblastic Cells During Pulmonary Inflammation and Fibrosis Induced by Carbon Nanotubes. Front. Pharmacol. 2019, 10, 1140. [Google Scholar] [CrossRef]
- Shen, W.; Liang, B.; Yin, J.; Li, X.; Cheng, J. Noscapine Increases the Sensitivity of Drug-Resistant Ovarian Cancer Cell Line SKOV3/DDP to Cisplatin by Regulating Cell Cycle and Activating Apoptotic Pathways. Cell Biochem. Biophys. 2015, 72, 203–213. [Google Scholar] [CrossRef]
- Domino, M.; Jasinski, T.; Kautz, E.; Juszczuk-Kubiak, E.; Ferreira-Dias, G.; Zabielski, R.; Sady, M.; Gajewski, Z. Expression of Genes Involved in the NF-ΚB-Dependent Pathway of the Fibrosis in the Mare Endometrium. Theriogenology 2020, 147, 18–24. [Google Scholar] [CrossRef]
- Doddapaneni, R.; Patel, K.; Chowdhury, N.; Singh, M. Reversal of Drug-Resistance by Noscapine Chemo-Sensitization in Docetaxel Resistant Triple Negative Breast Cancer. Sci. Rep. 2017, 7, 15824. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene (Accession Number) | Sequence 5′-3′ | Amplicon | References |
|---|---|---|---|
| COL1A2 (XM_001492939.3) | Forward: CAAGGGCATTAGGGGACACA | 196 | [13,15,53] |
| Reverse: ACCCACACTTCCATCGCTTC | |||
| RPL32 (XM_001492042.6) | Forward: AGCCATCTACTCGGCGTCA | 144 | [13,15,54] |
| Reverse: GTCAATGCCTCTGGGTTTCC |
| Time of Incubation | LDH Activity (%) |
|---|---|
| 1 h | 94.33 ± 0.91 a |
| 24 h | 89.96 ± 0.73 b |
| 48 h | 87.88 ± 0.85 b |
| Evaluated Variable | ||
|---|---|---|
| Isolated Factor/Interaction between Factors | COL1A2 Gene | COL1 Protein |
| CAT | 0.0003 | 0.0094 |
| NOSC | <0.0001 | 0.4065 |
| Time of treatment | 0.4375 | 0.6498 |
| Estrous cycle phase | 0.8009 | 0.0013 |
| CAT × NOSC | 0.0028 | 0.0794 |
| CAT × time of treatment | 0.0707 | 0.0011 |
| CAT × estrous cycle phase | 0.8579 | 0.001 |
| NOSC × time of treatment | 0.8591 | 0.9252 |
| NOSC × estrous cycle phase | 0.1535 | 0.2856 |
| Time of treatment × estrous cycle phase | 0.0281 | 0.0359 |
| CAT × NOSC × time of treatment | 0.3104 | 0.7706 |
| CAT × NOSC × estrous cycle phase | 0.0089 | 0.3501 |
| CAT × time of treatment × estrous cycle phase | 0.0817 | 0.0126 |
| NOSC × time of treatment × estrous cycle phase | 0.9812 | 0.297 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amaral, A.; Fernandes, C.; Szóstek-Mioduchowska, A.; Lukasik, K.; Rebordão, M.R.; Pinto-Bravo, P.; Skarzynski, D.J.; Ferreira-Dias, G. The Inhibitory Effect of Noscapine on the In Vitro Cathepsin G-Induced Collagen Expression in Equine Endometrium. Life 2021, 11, 1107. https://doi.org/10.3390/life11101107
Amaral A, Fernandes C, Szóstek-Mioduchowska A, Lukasik K, Rebordão MR, Pinto-Bravo P, Skarzynski DJ, Ferreira-Dias G. The Inhibitory Effect of Noscapine on the In Vitro Cathepsin G-Induced Collagen Expression in Equine Endometrium. Life. 2021; 11(10):1107. https://doi.org/10.3390/life11101107
Chicago/Turabian StyleAmaral, Ana, Carina Fernandes, Anna Szóstek-Mioduchowska, Karolina Lukasik, Maria Rosa Rebordão, Pedro Pinto-Bravo, Dariusz Jan Skarzynski, and Graça Ferreira-Dias. 2021. "The Inhibitory Effect of Noscapine on the In Vitro Cathepsin G-Induced Collagen Expression in Equine Endometrium" Life 11, no. 10: 1107. https://doi.org/10.3390/life11101107
APA StyleAmaral, A., Fernandes, C., Szóstek-Mioduchowska, A., Lukasik, K., Rebordão, M. R., Pinto-Bravo, P., Skarzynski, D. J., & Ferreira-Dias, G. (2021). The Inhibitory Effect of Noscapine on the In Vitro Cathepsin G-Induced Collagen Expression in Equine Endometrium. Life, 11(10), 1107. https://doi.org/10.3390/life11101107