
Comparing Native Crystal Structures and AlphaFold2 Predicted Water-Soluble G Protein-Coupled Receptor QTY Variants

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
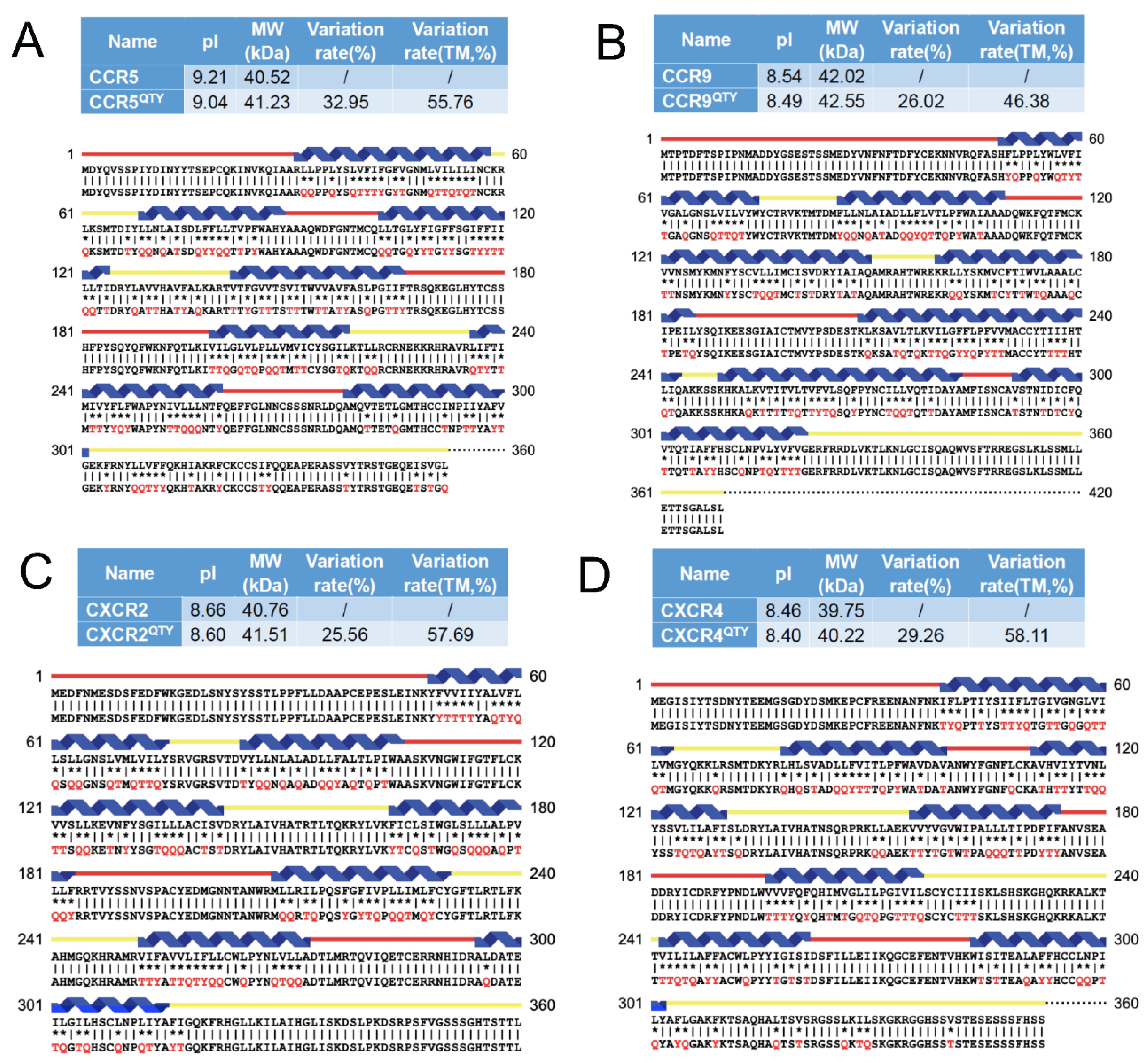
2.1. Protein Sequence Alignments
2.2. AlphaFold2 Predictions
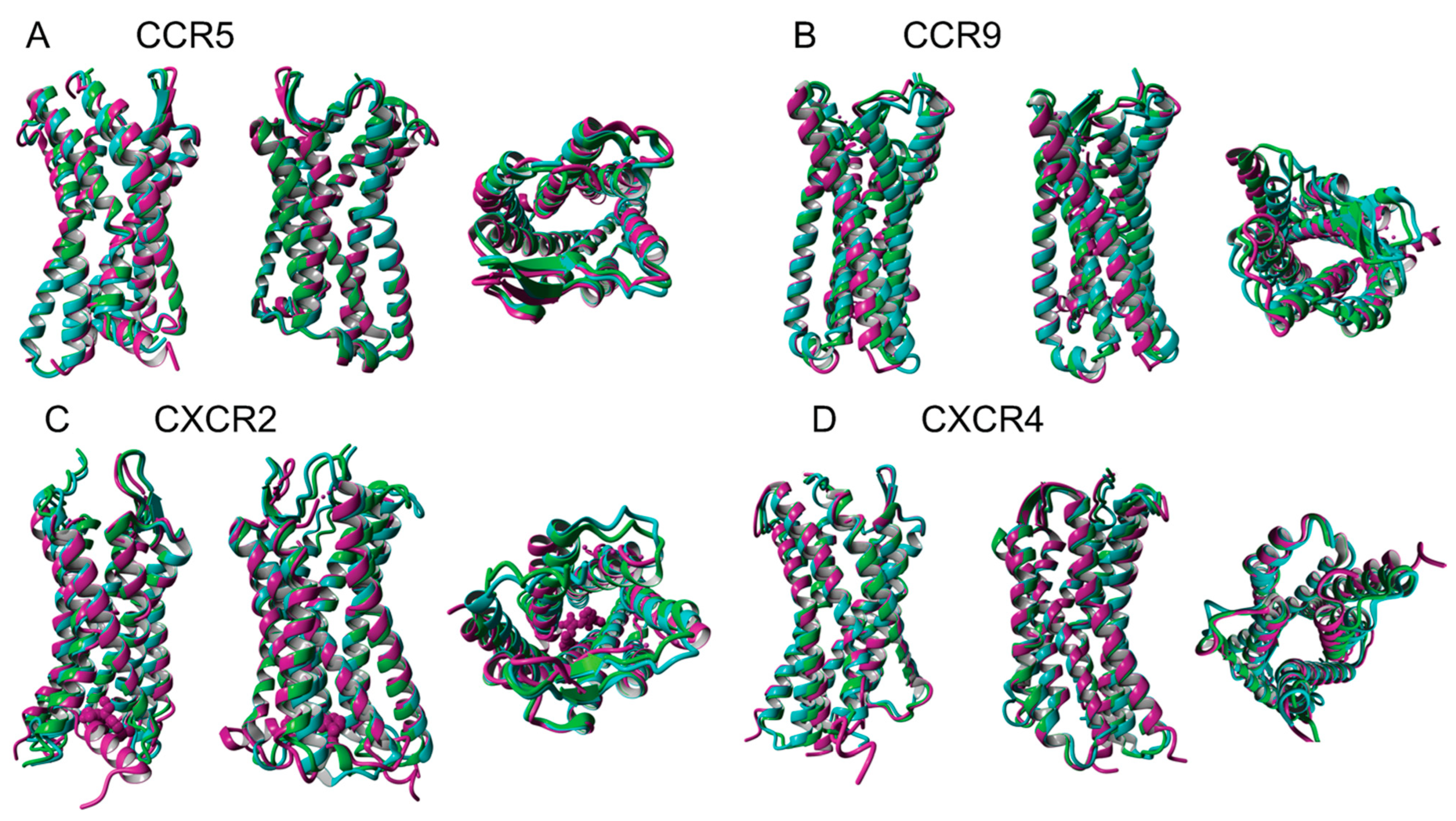
2.3. Superimpose the Structures
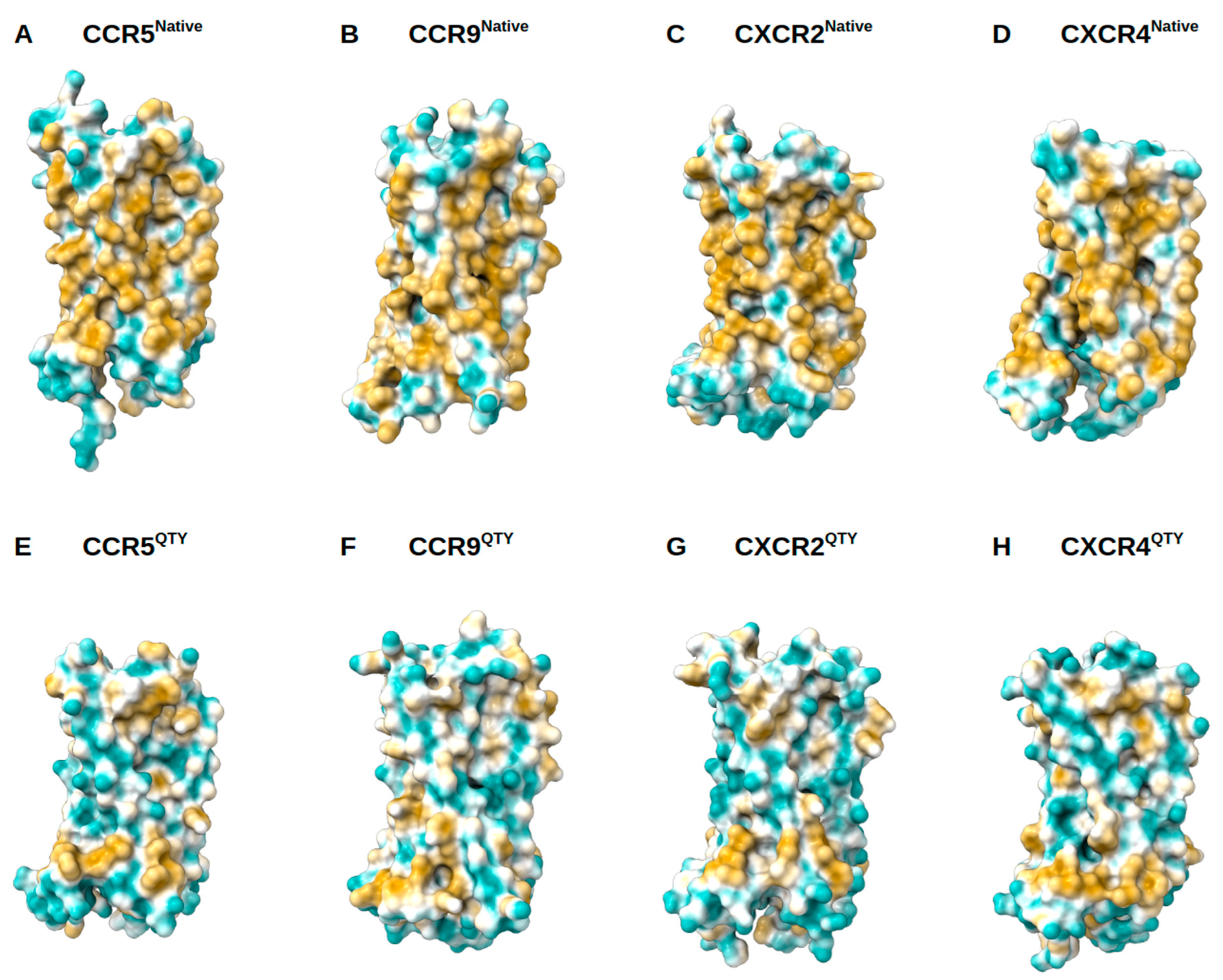
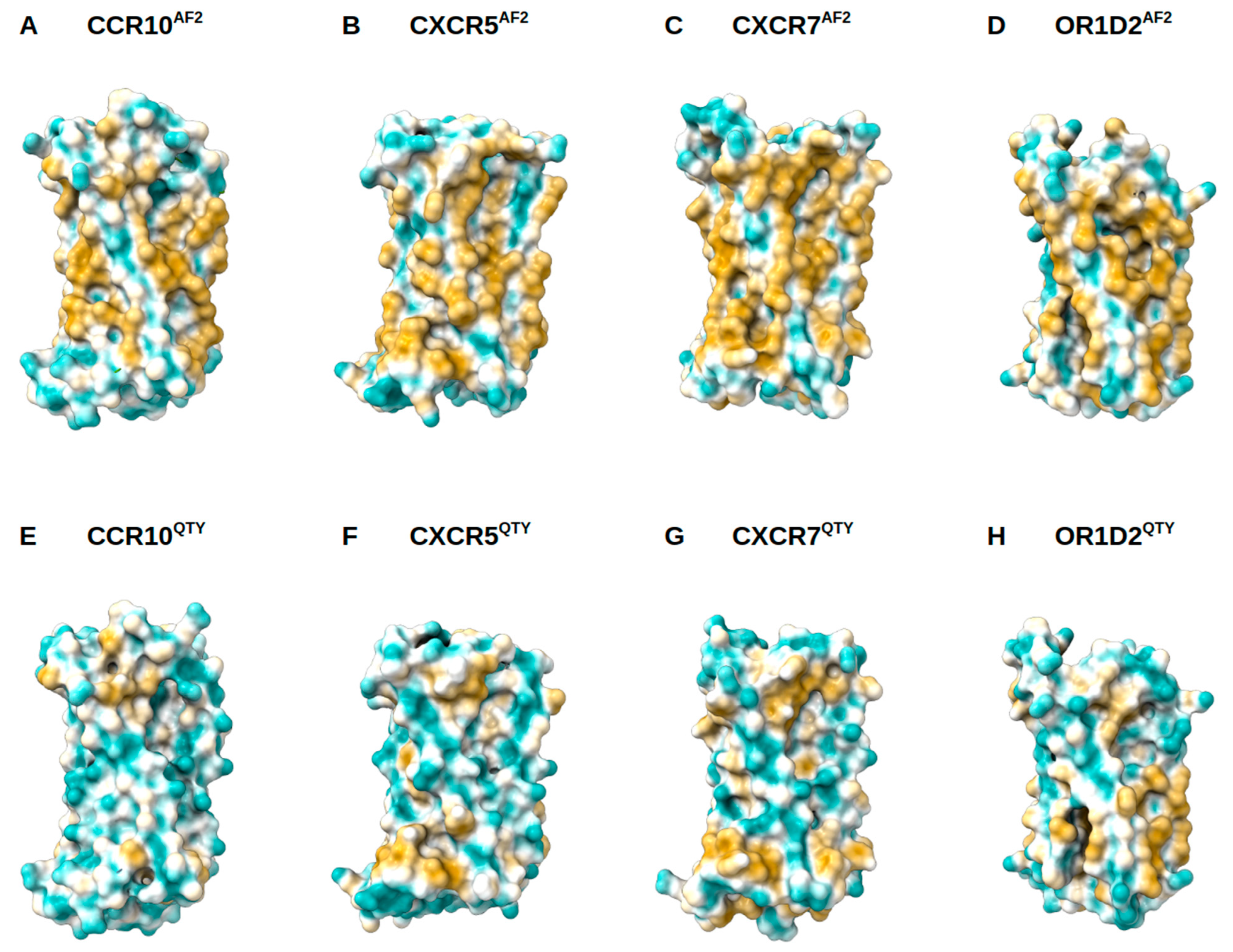
2.4. Structure Visualization
3. Results and Discussions
3.1. Protein Sequence Alignments
3.2. AlphaFold2 Predictions
3.3. Superimpositon of Native and Water-Soluble QTY Variants
3.4. Analysis of the Hydrophobic Surface of Native and Water-Soluble QTY Variants
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Competing Financial Interests
References
- Senior, A.W.; Evans, R.; Jumper, J.; Kirkpatrick, J.; Sifre, L.; Green, T.; Qin, C.; Žídek, A.; Nelson, A.W.R.; Bridgland, A.; et al. Improved protein structure prediction using potentials from deep learning. Nature 2020, 577, 706–710. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nat. Cell Biol. 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Tunyasuvunakool, K.; Adler, J.; Wu, Z.; Green, T.; Zielinski, M.; Žídek, A.; Bridgland, A.; Cowie, A.; Meyer, C.; Laydon, A.; et al. Highly accurate protein structure prediction for the human proteome. Nat. Cell Biol. 2021, 596, 590–596. [Google Scholar] [CrossRef]
- Baek, M.; DiMaio, F.; Anishchenko, I.; Dauparas, J.; Ovchinnikov, S.; Lee, G.R.; Wang, J.; Cong, Q.; Kinch, L.N.; Schaeffer, R.D. Accurate prediction of protein structures and interactions using a three-track neural network. Science 2021, 373, 871–876. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Tao, F.; Qing, R.; Tang, H.; Skuhersky, M.; Corin, K.; Tegler, L.; Wassie, A.; Wassie, B.; Kwon, Y.; et al. QTY code enables design of detergent-free chemokine receptors that retain ligand-binding activities. Proc. Natl. Acad. Sci. USA 2018, 115, E8652–E8659. [Google Scholar] [CrossRef] [Green Version]
- Qing, R.; Han, Q.; Fei, T.; Skuhersky, M.; Badr, M.; Chung, H.; Schubert, T.; Zhang, S. QTY code designed thermostable and water-soluble chimeric chemokine receptors with tunable ligand-binding activities. Proc. Natl. Acad. Sci. USA 2019, 116, 25668–25676. [Google Scholar] [CrossRef]
- Tegler, L.; Corin, K.; Pick, H.; Brookes, J.; Skuhersky, M.; Vogel, H.; Zhang, S. The G protein coupled receptor CXCR4 designed by the QTY code becomes more hydrophilic and retains cell signaling activity. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Hao, S.; Jin, D.; Zhang, S.; Qing, R. QTY Code-designed Water-soluble Fc-fusion Cytokine Receptors Bind to their Respective Ligands. QRB Discov. 2020, 1, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qing, R.; Tao, F.; Chatterjee, P.; Yang, G.; Han, Q.; Chung, H.; Ni, J.; Suter, B.P.; Kubicek, J.; Maertens, B.; et al. Non-full-length Water-Soluble CXCR4QTY and CCR5QTY Chemokine Receptors: Implication for Overlooked Truncated but Functional Membrane Receptors. iScience 2020, 23, 101670. [Google Scholar] [CrossRef]
- Proudfoot, A.E.I. Chemokine receptors: Multifaceted therapeutic targets. Nat. Rev. Immunol. 2002, 2, 106–115. [Google Scholar] [CrossRef]
- Dorsam, R.T.; Gutkind, J.S. G-protein-coupled receptors and cancer. Nat. Rev. Cancer 2007, 7, 79–94. [Google Scholar] [CrossRef]
- O’Hayre, M.; Degese, M.S.; Gutkind, J.S. Novel insights into G protein and G protein-coupled receptor signaling in cancer. Curr. Opin. Cell Biol. 2014, 27, 126–135. [Google Scholar] [CrossRef] [Green Version]
- Poeta, V.M.; Massara, M.; Capucetti, A.; Bonecchi, R. Chemokines and Chemokine Receptors: New Targets for Cancer Immunotherapy. Front. Immunol. 2019, 10, 379. [Google Scholar] [CrossRef] [Green Version]
- Wu, B.; Chien, E.Y.T.; Mol, C.D.; Fenalti, G.; Liu, W.; Katritch, V.; Abagyan, R.; Brooun, A.; Wells, P.; Bi, F.C.; et al. Structures of the CXCR4 Chemokine GPCR with Small-Molecule and Cyclic Peptide Antagonists. Science 2010, 330, 1066–1071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, Q.; Zhu, Y.; Li, J.; Chen, Z.; Han, G.W.; Kufareva, I.; Li, T.; Ma, L.; Fenalti, G.; Li, J.; et al. Structure of the CCR5 Chemokine Receptor–HIV Entry Inhibitor Maraviroc Complex. Science 2013, 341, 1387–1390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oswald, C.; Rappas, M.; Kean, J.; Doré, A.S.; Errey, J.C.; Bennett, K.; Deflorian, F.; Christopher, J.A.; Jazayeri, A.; Mason, J.S.; et al. Intracellular allosteric antagonism of the CCR9 receptor. Nat. Cell Biol. 2016, 540, 462–465. [Google Scholar] [CrossRef]
- Liu, K.; Wu, L.; Yuan, S.; Wu, M.; Xu, Y.; Sun, Q.; Li, S.; Zhao, S.; Hua, T.; Liu, Z.-J. Structural basis of CXC chemokine receptor 2 activation and signalling. Nat. Cell Biol. 2020, 585, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Korbecki, J.; Grochans, S.; Gutowska, I.; Barczak, K.; Baranowska-Bosiacka, I. CC Chemokines in a Tumor: A Review of Pro-Cancer and Anti-Cancer Properties of Receptors CCR5, CCR6, CCR7, CCR8, CCR9, and CCR10 Ligands. Int. J. Mol. Sci. 2020, 21, 7619. [Google Scholar] [CrossRef]
- Karnezis, T.; Farnsworth, R.H.; Harris, N.C.; Williams, S.P.; Caesar, C.; Byrne, D.J.; Herle, P.; Macheda, M.L.; Shayan, R.; Zhang, Y.F.; et al. CCL27/CCL28-CCR10 Chemokine Signaling Mediates Migration of Lymphatic Endothelial Cells. Cancer Res. 2019, 79, 1558–1572. [Google Scholar] [CrossRef]
- Ehrlich, A.T.; Semache, M.; Couvineau, P.; Wojcik, S.; Kobayashi, H.; Thelen, M.; Gross, F.; Hogue, M.; Le Gouill, C.; Darcq, E.; et al. Ackr3-Venus knock-in mouse lights up brain vasculature. Mol. Brain 2021, 14, 151. [Google Scholar] [CrossRef]
- Sun, X.; Cheng, G.; Hao, M.; Zheng, J.; Zhou, X.; Zhang, J.; Taichman, R.S.; Pienta, K.J.; Wang, J. CXCL12/CXCR4/CXCR7 chemokine axis and cancer progression. Cancer Metastasis Rev. 2010, 29, 709–722. [Google Scholar] [CrossRef] [Green Version]
- Gustavsson, M.; Wang, L.; Van Gils, N.; Stephens, B.; Zhang, P.; Schall, T.J.; Yang, S.; Abagyan, R.; Chance, M.R.; Kufareva, I.; et al. Structural basis of ligand interaction with atypical chemokine receptor 3. Nat. Commun. 2017, 8, 14135. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Riese, D.J.I.; Shen, J. The Role of the CXCL12/CXCR4/CXCR7 Chemokine Axis in Cancer. Front. Pharmacol. 2020, 11, 574667. [Google Scholar] [CrossRef]
- Buck, L.; Axel, R. A novel multigene family may encode odorant receptors: A molecular basis for odor recognition. Cell 1991, 65, 175–187. [Google Scholar] [CrossRef]
- Mombaerts, P.; Wang, F.; Dulac, C.; Vassar, R.; Chao, S.K.; Nemes, A.; Mendelsohn, M.; Edmondson, J.; Axel, R. The molecular biology of olfactory perception. Cold Spring Harb. Symp. Quant. Biol. 1996, 61, 135–145. [Google Scholar] [PubMed]
- Buck, L.B. Unraveling smell. Harvey Lect. 2007, 101, 117–134. [Google Scholar]
- Touhara, K. Odor discrimination by G protein-coupled olfactory receptors. Microsc. Res. Tech. 2002, 58, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Sanz, G.; Schlegel, C.; Pernollet, J.-C.; Briand, L. Comparison of Odorant Specificity of Two Human Olfactory Receptors from Different Phylogenetic Classes and Evidence for Antagonism. Chem. Senses 2005, 30, 69–80. [Google Scholar] [CrossRef]
- Konagurthu, A.; Whisstock, J.; Stuckey, P.; Lesk, A. MUSTANG: A multiple structural alignment algorithm. Proteins Struct. Funct. Bioinform. 2006, 64, 559–574. [Google Scholar] [CrossRef] [PubMed]
- Almén, M.S.; Nordström, K.J.; Fredriksson, R.; Schiöth, H.B. Mapping the human membrane proteome: A majority of the human membrane proteins can be classified according to function and evolutionary origin. BMC Biol. 2009, 7, 50. [Google Scholar] [CrossRef] [Green Version]
- Pauling, L.; Corey, R.B. Atomic Coordinates and Structure Factors for Two Helical Configurations of Polypeptide Chains. Proc. Natl. Acad. Sci. USA 1951, 37, 235–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fersht, A. Structure and Mechanism in Protein Science: A Guide to Enzyme Catalysis and Protein Folding; W.H. Freeman: New York, NY, USA, 1998; pp. 523–532. [Google Scholar]
- Brändén, C.-I.; Tooze, J. Introduction to Protein Structure, 2nd ed.; Garland Publishing: London, UK; New York, NY, USA, 1999; pp. 15–17. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Value |
|---|---|
| homooligomer | 1 |
| msa_method | mmseqs2 |
| msa_format | fas |
| pair_mode | unpaired |
| pair_cov | 50 |
| pair_qid | 20 |
| rank_by | pLDDT |
| use_turbo | True |
| max_msa | 512:1024 |
| show_images | True |
| num_models | 5 |
| use_ptm | True |
| num_ensemble | 1 |
| max_recycles | 3 |
| tol | 0 |
| num_samples | 1 |
| subsample_msa | True |
| num_relax | None |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Skuhersky, M.A.; Tao, F.; Qing, R.; Smorodina, E.; Jin, D.; Zhang, S. Comparing Native Crystal Structures and AlphaFold2 Predicted Water-Soluble G Protein-Coupled Receptor QTY Variants. Life 2021, 11, 1285. https://doi.org/10.3390/life11121285
Skuhersky MA, Tao F, Qing R, Smorodina E, Jin D, Zhang S. Comparing Native Crystal Structures and AlphaFold2 Predicted Water-Soluble G Protein-Coupled Receptor QTY Variants. Life. 2021; 11(12):1285. https://doi.org/10.3390/life11121285
Chicago/Turabian StyleSkuhersky, Michael A., Fei Tao, Rui Qing, Eva Smorodina, David Jin, and Shuguang Zhang. 2021. "Comparing Native Crystal Structures and AlphaFold2 Predicted Water-Soluble G Protein-Coupled Receptor QTY Variants" Life 11, no. 12: 1285. https://doi.org/10.3390/life11121285
APA StyleSkuhersky, M. A., Tao, F., Qing, R., Smorodina, E., Jin, D., & Zhang, S. (2021). Comparing Native Crystal Structures and AlphaFold2 Predicted Water-Soluble G Protein-Coupled Receptor QTY Variants. Life, 11(12), 1285. https://doi.org/10.3390/life11121285