
Primitive Compartmentalization for the Sustainable Replication of Genetic Molecules



Abstract
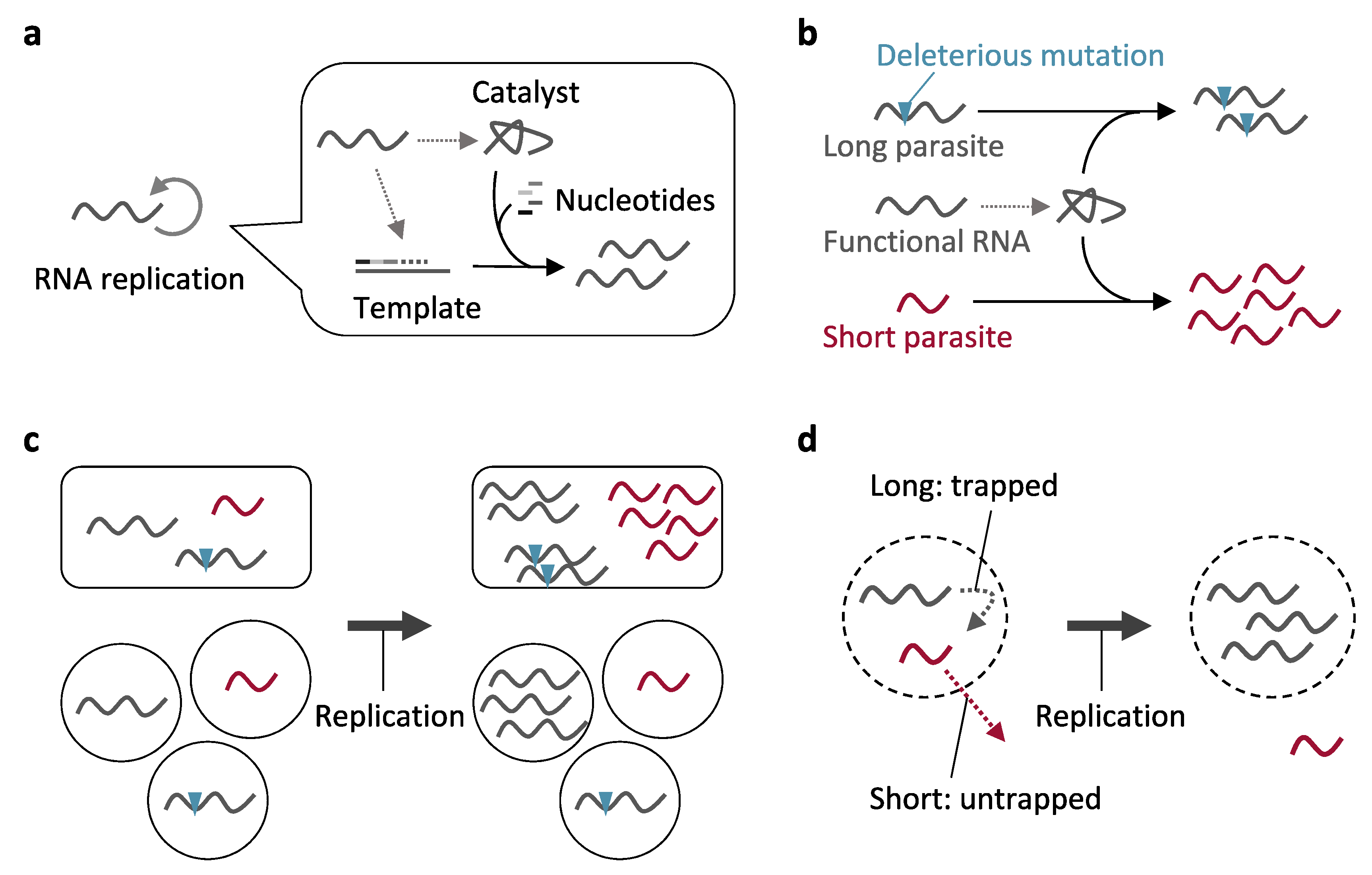
:1. Introduction
2. Parasitic Molecules and Prevention of Their Replication through Compartmentalization
3. Primitive Compartmentalization
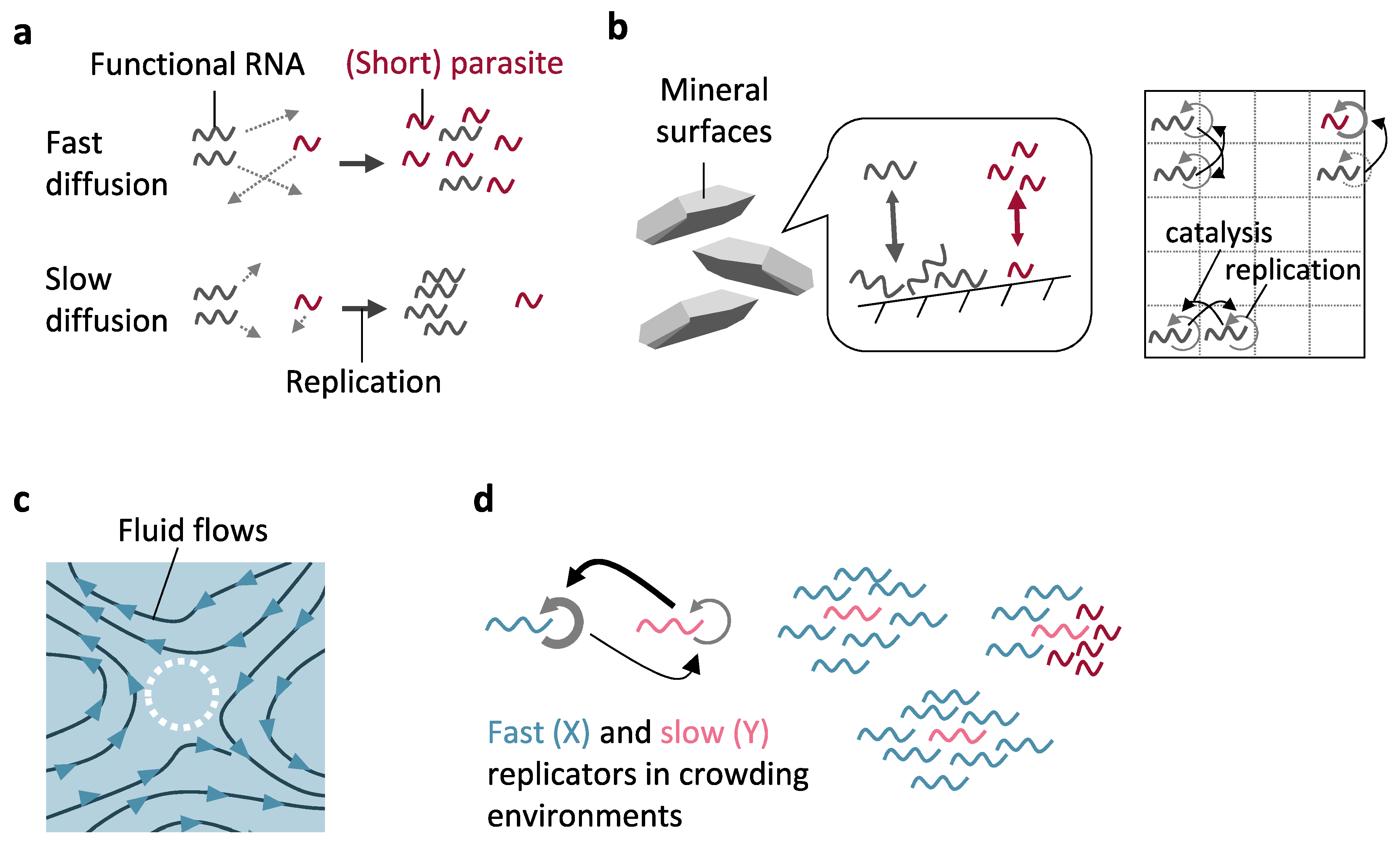
3.1. Compartments without Boundaries
3.1.1. Mineral Surfaces
3.1.2. Active Fluid Environments
3.1.3. Crowded Environments
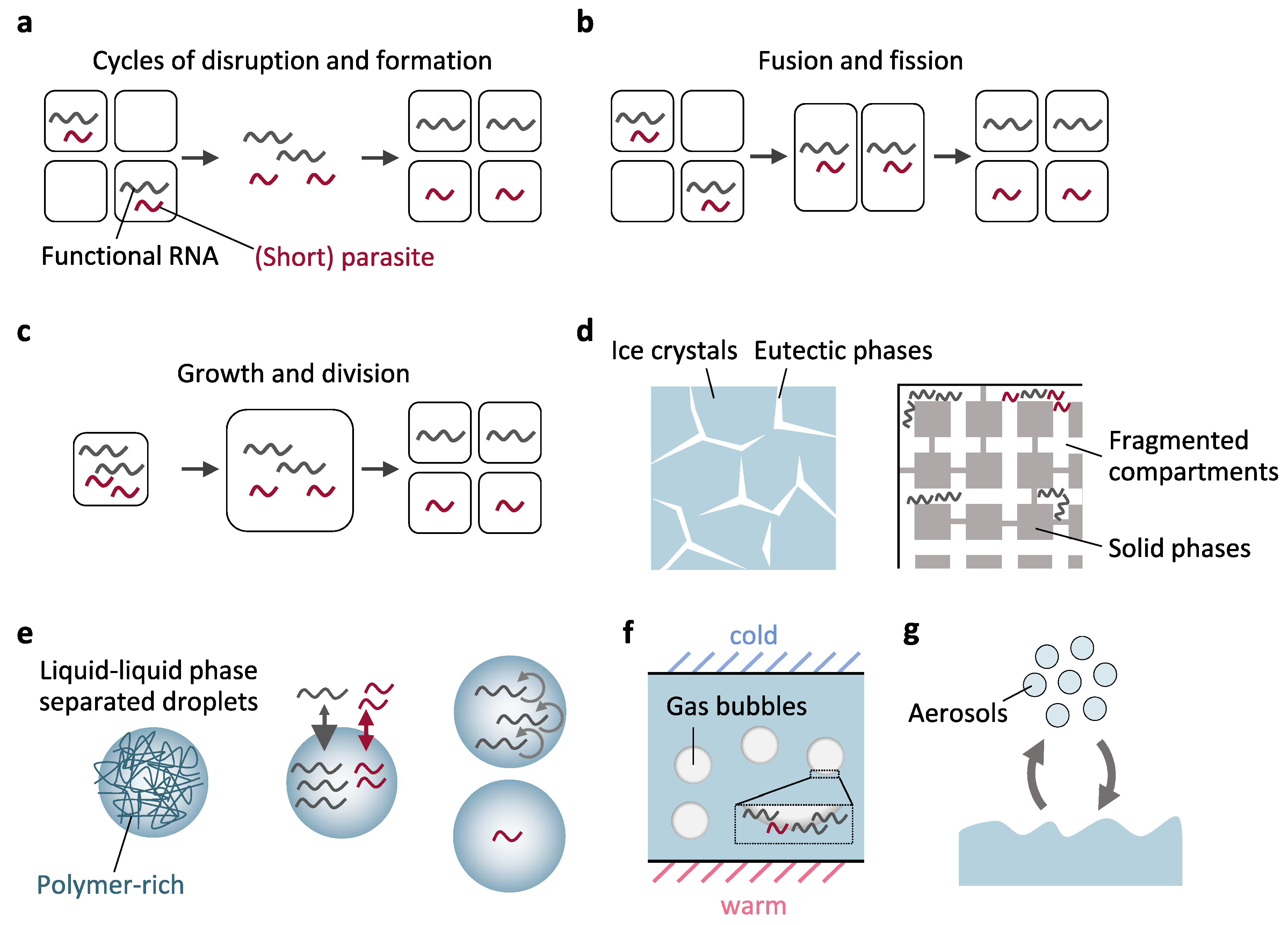
3.2. Compartments with Boundaries
3.2.1. Ice Eutectic Phases
3.2.2. Membranous Compartments
3.2.3. Membraneless Cell-Like Compartments
3.2.4. Gas Bubbles
3.2.5. Atmospheric Compartments
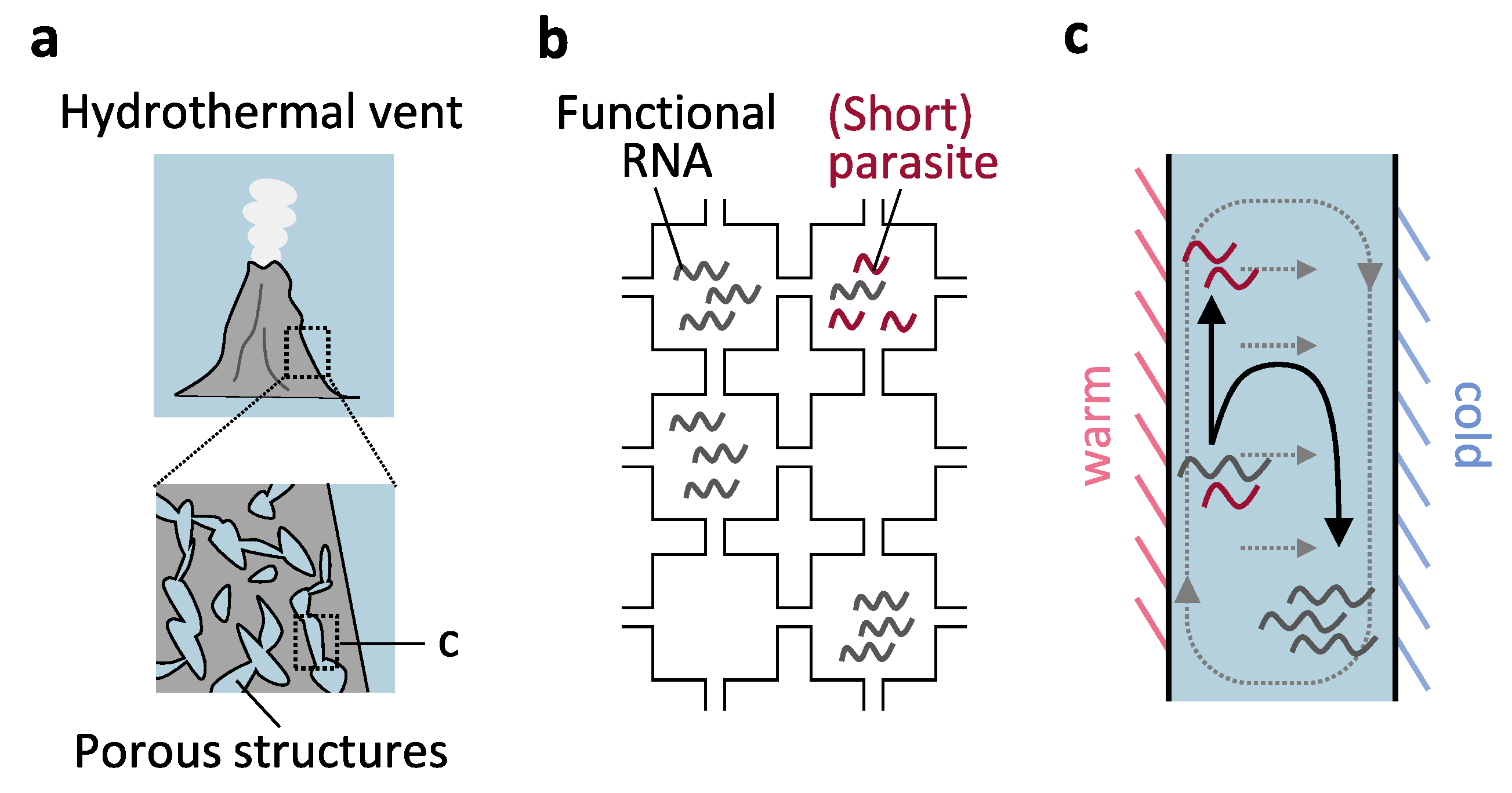
3.3. Compartments with Incomplete Boundaries
Porous Structures of Hydrothermal Vents
4. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Maynard-Smith, J.; Szathmáry, E. The Major Transitions in Evolution; Oxford University Press: Oxford, UK, 1997. [Google Scholar]
- Ichihashi, N. What can we learn from the construction of in vitro replication systems? Ann. N. Y. Acad. Sci. 2019, 1447, 144–156. [Google Scholar] [CrossRef]
- Monnard, P.A.; Walde, P. Current ideas about prebiological compartmentalization. Life 2015, 5, 1239–1263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wachowius, F.; Attwater, J.; Holliger, P. Nucleic acids: Function and potential for abiogenesis. Q. Rev. Biophys. 2017, 50. [Google Scholar] [CrossRef] [PubMed]
- Joyce, G.F.; Szostak, J.W. Protocells and RNA self-replication. Cold Spring Harb. Perspect. Biol. 2018, 10, a034801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanczyc, M.M.; Monnard, P.A. Primordial membranes: More than simple container boundaries. Curr. Opin. Chem. Biol. 2017, 40, 78–86. [Google Scholar] [CrossRef]
- Sarkar, S.; Das, S.; Dagar, S.; Joshi, M.P.; Mungi, C.V.; Sawant, A.A.; Patki, G.M.; Rajamani, S. Prebiological membranes and their role in the emergence of early cellular life. J. Membr. Biol. 2020, 253, 589–608. [Google Scholar] [CrossRef]
- Gilbert, W. The RNA world. Nature 1986, 319, 618. [Google Scholar] [CrossRef]
- Joyce, G.F. The antiquity of RNA-based evolution. Nature 2002, 418, 214–221. [Google Scholar] [CrossRef]
- Higgs, P.G.; Lehman, N. The RNA world: Molecular cooperation at the origins of life. Nat. Rev. Genet. 2014, 16, 7–17. [Google Scholar] [CrossRef]
- Mills, D.R.; Peterson, R.L.; Spiegelman, S. An extracellular Darwinian experiment with a self-duplicating nucleic acid molecule. Proc. Natl. Acad. Sci. USA 1967, 58, 217–224. [Google Scholar] [CrossRef] [Green Version]
- Maynard Smith, J. Hypercycles and the origin of life. Nature 1979, 280, 445–446. [Google Scholar] [CrossRef]
- Ichihashi, N.; Usui, K.; Kazuta, Y.; Sunami, T.; Matsuura, T.; Yomo, T. Darwinian evolution in a translation-coupled RNA replication system within a cell-like compartment. Nat. Commun. 2013, 4, 2494. [Google Scholar] [CrossRef]
- Matsumura, S.; Kun, Á.; Ryckelynck, M.; Coldren, F.; Szilágyi, A.; Jossinet, F.; Rick, C.; Nghe, P.; Szathmáry, E.; Griffiths, A.D. Transient compartmentalization of RNA replicators prevents extinction due to parasites. Science 2016, 354, 1293–1296. [Google Scholar] [CrossRef] [Green Version]
- Bansho, Y.; Furubayashi, T.; Ichihashi, N.; Yomo, T. Host-parasite oscillation dynamics and evolution in a compartmentalized RNA replication system. Proc. Natl. Acad. Sci. USA 2016, 113, 4045–4050. [Google Scholar] [CrossRef] [Green Version]
- Mizuuchi, R.; Ichihashi, N.; Usui, K.; Kazuta, Y.; Yomo, T. Adaptive evolution of an artificial RNA genome to a reduced ribosome Environment. ACS Synth. Biol. 2015, 4, 292–298. [Google Scholar] [CrossRef]
- Mizuuchi, R.; Ichihashi, N.; Yomo, T. Adaptation and diversification of an RNA replication system under initiation- or termination-impaired translational conditions. ChemBioChem 2016, 17, 1229–1232. [Google Scholar] [CrossRef]
- Furubayashi, T.; Ueda, K.; Bansho, Y.; Daisuke, M.; Shota, N.; Mizuuchi, R.; Ichihashi, N. Emergence and diversification of a host-parasite RNA ecosystem through Darwinian evolution. Elife 2020, 9, e56038. [Google Scholar] [CrossRef] [PubMed]
- Yoshiyama, T.; Ichii, T.; Yomo, T.; Ichihashi, N. Automated in vitro evolution of a translation-coupled RNA replication system in a droplet flow reactor. Sci. Rep. 2018, 8, 11867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizuuchi, R.; Ichihashi, N. Sustainable replication and coevolution of cooperative RNAs in an artificial cell-like system. Nat. Ecol. Evol. 2018, 2, 1654–1660. [Google Scholar] [CrossRef]
- Eigen, M.; Schuster, P. A principle of natural self-organization - Part A: Emergence of the hypercycle. Naturwissenschaften 1977, 64, 541–565. [Google Scholar] [CrossRef] [PubMed]
- Mizuuchi, R.; Blokhuis, A.; Vincent, L.; Nghe, P.; Lehman, N.; Baum, D. Mineral surfaces select for longer RNA molecules. Chem. Commun. 2019, 55, 2090–2093. [Google Scholar] [CrossRef] [PubMed]
- Gibbs, D.; Lohrmann, R.; Orgel, L.E. Template-directed synthesis and selective adsorption of oligoadenylates on hydroxyapatite. J. Mol. Evol. 1980, 15, 347–354. [Google Scholar] [CrossRef]
- Boerlijst, M.C.; Hogeweg, P. Spiral wave structure in pre-biotic evolution – hypercycles stable against parasites. Phys. D 1991, 48, 17–28. [Google Scholar] [CrossRef]
- Czárán, T.; Szathmáry, E. Coexistence of replicators in prebiotic evolution. In The Geometry of Ecological Interactions: Simplifying Spatial Complexity; Dieckmann, U., Law, R., Metz, J.A., Eds.; Cambridge University Press: Cambridge, UK, 2000; pp. 116–134. [Google Scholar]
- Szabó, P.; Scheuring, I.; Czárán, T.; Szathmáry, E. In silico simulations reveal that replicators with limited dispersal evolve towards higher efficiency and fidelity. Nature 2002, 420, 340–343. [Google Scholar] [CrossRef] [PubMed]
- Hogeweg, P.; Takeuchi, N. Multilevel selection in models of prebiotic evolution: Compartments and spatial self-organization. Orig. Life Evol. Biosph. 2003, 33, 375–403. [Google Scholar] [CrossRef] [PubMed]
- Könnyu, B.; Czárán, T.; Szathmáry, E. Prebiotic replicase evolution in a surface-bound metabolic system: Parasites as a source of adaptive evolution. BMC Evol. Biol. 2008, 8, 267. [Google Scholar] [CrossRef] [Green Version]
- Takeuchi, N.; Hogeweg, P. Multilevel selection in models of prebiotic evolution II: A direct comparison of compartmentalization and spatial self-organization. PLoS Comput. Biol. 2009, 5, e1000542. [Google Scholar] [CrossRef] [Green Version]
- Shay, J.A.; Huynh, C.; Higgs, P.G. The origin and spread of a cooperative replicase in a prebiotic chemical system. J. Theor. Biol. 2015, 364, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Colizzi, E.S.; Hogeweg, P. Parasites sustain and enhance RNA-like replicators through spatial self-organisation. PLoS Comput. Biol. 2016, 12, e1004902. [Google Scholar] [CrossRef]
- Kim, Y.E.; Higgs, P.G. Co-operation between polymerases and nucleotide synthetases in the RNA world. PLoS Comput. Biol. 2016, 12, e1005161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tupper, A.S.; Higgs, P.G. Error thresholds for RNA replication in the presence of both point mutations and premature termination errors. J. Theor. Biol. 2017, 428, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Hanczyc, M.M.; Fujikawa, S.M.; Szostak, J.W. Experimental models of primitive cellular compartments: Encapsulation, growth, and division. Science 2003, 302, 618–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanczyc, M.M.; Mansy, S.S.; Szostak, J.W. Mineral surface directed membrane assembly. Orig. Life Evol. Biosph. 2007, 37, 67–82. [Google Scholar] [CrossRef]
- Sahai, N.; Kaddour, H.; Dalai, P.; Wang, Z.; Bass, G.; Gao, M. Mineral surface chemistry and nanoparticle-aggregation control membrane self-assembly. Sci. Rep. 2017, 7, 43418. [Google Scholar] [CrossRef] [Green Version]
- Krieger, M.S.; Sinai, S.; Nowak, M.A. Turbulent coherent structures and early life below the Kolmogorov scale. Nat. Commun. 2020, 11, 2192. [Google Scholar] [CrossRef]
- Kamimura, A.; Kaneko, K. Reproduction of a protocell by replication of a minority molecule in a catalytic reaction network. Phys. Rev. Lett. 2010, 105, 268103. [Google Scholar] [CrossRef] [Green Version]
- Attwater, J.; Wochner, A.; Pinheiro, V.B.; Coulson, A.; Holliger, P. Ice as a protocellular medium for RNA replication. Nat. Commun. 2010, 1, 76. [Google Scholar] [CrossRef]
- Blokhuis, A.; Lacoste, D.; Nghe, P.; Peliti, L. Selection dynamics in transient compartmentalization. Phys. Rev. Lett. 2018, 120, 158101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laurent, G.; Peliti, L.; Lacoste, D. Survival of self-replicating molecules under transient compartmentalization with natural selection. Life 2019, 9, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blokhuis, A.; Nghe, P.; Peliti, L.; Lacoste, D. The generality of transient compartmentalization and its associated error thresholds. J. Theor. Biol. 2020, 487, 110110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niesert, U.; Harnasch, D.; Bresch, C. Origin of life between Scylla and Charybdis. J. Mol. Evol. 1981, 17, 348–353. [Google Scholar] [CrossRef]
- Furubayashi, T.; Ichihashi, N. Sustainability of a compartmentalized host-parasite replicator system under periodic washout-mixing cycles. Life 2018, 8, 3. [Google Scholar] [CrossRef] [Green Version]
- Szathmáry, E.; Demeter, L. Group selection of early replicators and the origin of life. J. Theor. Biol. 1987, 128, 463–486. [Google Scholar] [CrossRef]
- Markvoort, A.J.; Sinai, S.; Nowak, M.A. Computer simulations of cellular group selection reveal mechanism for sustaining cooperation. J. Theor. Biol. 2014, 357, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Shah, V.; de Bouter, J.; Pauli, Q.; Tupper, A.S.; Higgs, P.G. Survival of RNA replicators is much easier in protocells than in surface-based, spatial systems. Life 2019, 9, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monnard, P.A.; Oberholzer, T.; Luisi, P. Entrapment of nucleic acids in liposomes. Biochim. Biophys. Acta - Biomembr. 1997, 1329, 39–50. [Google Scholar] [CrossRef] [Green Version]
- Qiao, H.; Hu, N.; Bai, J.; Ren, L.; Liu, Q.; Fang, L.; Wang, Z. Encapsulation of nucleic acids into giant unilamellar vesicles by freeze-thaw: A way protocells may form. Orig. Life Evol. Biosph. 2017, 47, 499–510. [Google Scholar] [CrossRef]
- Drobot, B.; Iglesias-Artola, J.M.; Le Vay, K.; Mayr, V.; Kar, M.; Kreysing, M.; Mutschler, H.; Tang, T.Y.D. Compartmentalised RNA catalysis in membrane-free coacervate protocells. Nat. Commun. 2018, 9, 3643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strulson, C.A.; Molden, R.C.; Keating, C.D.; Bevilacqua, P.C. RNA catalysis through compartmentalization. Nat. Chem. 2012, 4, 941–946. [Google Scholar] [CrossRef]
- Mizuuchi, R.; Ichihashi, N. Translation-coupled RNA replication and parasitic replicators in membrane-free compartments. Chem. Commun. 2020, 56, 13453–13456. [Google Scholar] [CrossRef]
- Dora Tang, T.Y.; Rohaida Che Hak, C.; Thompson, A.J.; Kuimova, M.K.; Williams, D.S.; Perriman, A.W.; Mann, S. Fatty acid membrane assembly on coacervate microdroplets as a step towards a hybrid protocell model. Nat. Chem. 2014, 6, 527–533. [Google Scholar] [CrossRef] [Green Version]
- Sakuta, H.; Fujita, F.; Hamada, T.; Hayashi, M.; Takiguchi, K.; Tsumoto, K.; Yoshikawa, K. Self-emergent protocells generated in an aqueous solution with binary macromolecules through liquid-liquid phase separation. ChemBioChem 2020, 21, 3323–3328. [Google Scholar] [CrossRef]
- Morasch, M.; Liu, J.; Dirscherl, C.F.; Ianeselli, A.; Kühnlein, A.; Le Vay, K.; Schwintek, P.; Islam, S.; Corpinot, M.K.; Scheu, B.; et al. Heated gas bubbles enrich, crystallize, dry, phosphorylate and encapsulate prebiotic molecules. Nat. Chem. 2019, 11, 779–788. [Google Scholar] [CrossRef]
- Dobson, C.M.; Ellison, G.B.; Tuck, A.F.; Vaida, V. Atmospheric aerosols as prebiotic chemical reactors. Proc. Natl. Acad. Sci. USA 2000, 97, 11864–11868. [Google Scholar] [CrossRef] [Green Version]
- Kreysing, M.; Keil, L.; Lanzmich, S.; Braun, D. Heat flux across an open pore enables the continuous replication and selection of oligonucleotides towards increasing length. Nat. Chem. 2015, 7, 203–208. [Google Scholar] [CrossRef]
- Branciamore, S.; Gallori, E.; Szathmáry, E.; Czárán, T. The origin of life: Chemical evolution of a metabolic system in a mineral honeycomb? J. Mol. Evol. 2009, 69, 458–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Budin, I.; Bruckner, R.J.; Szostak, J.W. Formation of protocell-like vesicles in a thermal diffusion column. J. Am. Chem. Soc. 2009, 131, 9628–9629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cleaves, H.J.; Scott, A.M.; Hill, F.C.; Leszczynski, J.; Sahai, N.; Hazen, R. Mineral–organic interfacial processes: Potential roles in the origins of life. Chem. Soc. Rev. 2012, 41, 5502–5525. [Google Scholar] [CrossRef]
- Yu, W.H.; Li, N.; Tong, D.S.; Zhou, C.H.; Lin, C.X.; Xu, C.Y. Adsorption of proteins and nucleic acids on clay minerals and their interactions: A review. Appl. Clay Sci. 2013, 80–81, 443–452. [Google Scholar] [CrossRef]
- Biondi, E.; Furukawa, Y.; Kawai, J.; Benner, S.A. Adsorption of RNA on mineral surfaces and mineral precipitates. Beilstein J. Org. Chem. 2017, 13, 393–404. [Google Scholar] [CrossRef] [Green Version]
- Swadling, J.B.; Coveney, P.V.; Greenwell, H.C. Clay minerals mediate folding and regioselective interactions of RNA: A large-scale atomistic simulation study. J. Am. Chem. Soc. 2010, 132, 13750–13764. [Google Scholar] [CrossRef] [PubMed]
- Acevedo, O.L.; Orgel, L.E. Template-directed oligonucleotide ligation on hydroxylapatite. Nature 1986, 321, 790–792. [Google Scholar] [CrossRef] [PubMed]
- Ferris, J.P.; Hill, A.R.; Liu, R.; Orgel, L.E. Synthesis of long prebiotic oligomers on mineral surfaces. Nature 1996, 381, 59–61. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Ferris, J.P. Synthesis of 35-40 mers of RNA oligomers from unblocked monomers. A simple approach to the RNA world. Chem. Commun. 2003, 3, 1458–1459. [Google Scholar] [CrossRef]
- Joshi, P.C.; Aldersley, M.F.; Delano, J.W.; Ferris, J.P. Mechanism of montmorillonite catalysis in the formation of RNA oligomers. J. Am. Chem. Soc. 2009, 131, 13369–13374. [Google Scholar] [CrossRef]
- Biondi, E.; Branciamore, S.; Maurel, M.C.; Gallori, E. Montmorillonite protection of an UV-irradiated hairpin ribozyme: Evolution of the RNA world in a mineral environment. BMC Evol. Biol. 2007, 7, S2. [Google Scholar] [CrossRef]
- Ricardo, A.; Carrigan, M.A.; Olcott, A.N.; Benner, S.A. Borate minerals stabilize ribose. Science 2004, 303, 196. [Google Scholar] [CrossRef] [Green Version]
- Costanzo, G.; Saladino, R.; Crestini, C.; Ciciriello, F.; Di Mauro, E. Nucleoside phosphorylation by phosphate minerals. J. Biol. Chem. 2007, 282, 16729–16735. [Google Scholar] [CrossRef] [Green Version]
- Gull, M.; Mojica, M.A.; Fernández, F.M.; Gaul, D.A.; Orlando, T.M.; Liotta, C.L.; Pasek, M.A. Nucleoside phosphorylation by the mineral schreibersite. Sci. Rep. 2015, 5, 17198. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.-J.; Furukawa, Y.; Kakegawa, T.; Bita, A.; Scorei, R.; Benner, S.A. Evaporite borate-containing mineral ensembles make phosphate available and regiospecifically phosphorylate ribonucleosides: Borate as a multifaceted problem solver in prebiotic chemistry. Angew. Chemie Int. Ed. 2016, 128, 16048–16052. [Google Scholar] [CrossRef]
- Biondi, E.; Branciamore, S.; Fusi, L.; Gago, S.; Gallori, E. Catalytic activity of hammerhead ribozymes in a clay mineral environment: Implications for the RNA world. Gene 2007, 389, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Stephenson, J.D.; Popović, M.; Bristow, T.F.; Ditzler, M.A. Evolution of ribozymes in the presence of a mineral surface. RNA 2016, 22, 1893–1901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dix, J.A.; Verkman, A.S. Crowding effects on diffusion in solutions and cells. Annu. Rev. Biophys. 2008, 37, 247–263. [Google Scholar] [CrossRef]
- Bada, J.L.; Bigham, C.; Miller, S.L. Impact melting of frozen oceans on the early Earth: Implications for the origin of life. Proc. Natl. Acad. Sci. USA 1994, 91, 1248–1250. [Google Scholar] [CrossRef] [Green Version]
- Zahnle, K.; Arndt, N.; Cockell, C.; Halliday, A.; Nisbet, E.; Selsis, F.; Sleep, N.H. Emergence of a habitable planet. Space Sci. Rev. 2007, 129, 35–78. [Google Scholar] [CrossRef]
- Kanavarioti, A.; Monnard, P.A.; Deamer, D.W. Eutectic phases in ice facilitate nonenzymatic nucleic acid synthesis. Astrobiology 2001, 1, 271–281. [Google Scholar] [CrossRef]
- Monnard, P.A.; Kanavarioti, A.; Deamer, D.W. Eutectic phase polymerization of activated ribonucleotide mixtures yields quasi-equimolar incorporation of purine and pyrimidine nucleobases. J. Am. Chem. Soc. 2003, 125, 13734–13740. [Google Scholar] [CrossRef]
- Monnard, P.A.; Szostak, J.W. Metal-ion catalyzed polymerization in the eutectic phase in water-ice: A possible approach to template-directed RNA polymerization. J. Inorg. Biochem. 2008, 102, 1104–1111. [Google Scholar] [CrossRef]
- Mutschler, H.; Taylor, A.I.; Lightowler, A.; Houlihan, G.; Abramov, M.; Herdewijn, P.; Holliger, P. Innate potential of random genetic oligomer pools for recombination. Elife 2018, 7, e43022. [Google Scholar] [CrossRef]
- Smail, B.A.; Clifton, B.E.; Mizuuchi, R.; Lehman, N. Spontaneous advent of genetic diversity in RNA populations through multiple recombination mechanisms. RNA 2019, 25, 453–464. [Google Scholar] [CrossRef]
- Vlassov, A.V.; Johnston, B.H.; Landweber, L.F.; Kazakov, S.A. Ligation activity of fragmented ribozymes in frozen solution: Implications for the RNA world. Nucleic Acids Res. 2004, 32, 2966–2974. [Google Scholar] [CrossRef] [Green Version]
- Kazakov, S.A.; Balatskaya, S.V.; Johnston, B.H. Ligation of the hairpin ribozyme in cis induced by freezing and dehydration. RNA 2006, 12, 446–456. [Google Scholar] [CrossRef] [Green Version]
- Mutschler, H.; Wochner, A.; Holliger, P. Freeze-thaw cycles as drivers of complex ribozyme assembly. Nat. Chem. 2015, 7, 502–508. [Google Scholar] [CrossRef] [Green Version]
- Litschel, T.; Ganzinger, K.A.; Movinkel, T.; Heymann, M.; Robinson, T.; Mutschler, H.; Schwille, P. Freeze-thaw cycles induce content exchange between cell-sized lipid vesicles. New J. Phys. 2018, 20, 055008. [Google Scholar] [CrossRef]
- Tsuji, G.; Fujii, S.; Sunami, T.; Yomo, T. Sustainable proliferation of liposomes compatible with inner RNA replication. Proc. Natl. Acad. Sci. USA 2016, 113, 590–595. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Huang, X.; Tang, T.Y.D.; Mann, S. Synthetic cellularity based on non-lipid micro-compartments and protocell models. Curr. Opin. Chem. Biol. 2014, 22, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Subramaniam, A.B.; Wan, J.; Gopinath, A.; Stone, H.A. Semi-permeable vesicles composed of natural clay. Soft Matter 2011, 7, 2600–2612. [Google Scholar] [CrossRef] [Green Version]
- Oparin, A.I. Origin of Life; Macmillan: New York, NY, USA, 1938. [Google Scholar]
- Keating, C.D. Aqueous phase separation as a possible route to compartmentalization of biological molecules. Acc. Chem. Res. 2012, 45, 2114–2124. [Google Scholar] [CrossRef]
- Mann, S. Systems of creation: The emergence of life from nonliving matter. Acc. Chem. Res. 2012, 45, 2131–2141. [Google Scholar] [CrossRef] [PubMed]
- Poudyal, R.R.; Pir Cakmak, F.; Keating, C.D.; Bevilacqua, P.C. Physical principles and extant biology reveal roles for RNA-containing membraneless compartments in origins of life chemistry. Biochemistry 2018, 57, 2509–2519. [Google Scholar] [CrossRef]
- Yoshizawa, T.; Nozawa, R.S.; Jia, T.Z.; Saio, T.; Mori, E. Biological phase separation: Cell biology meets biophysics. Biophys. Rev. 2020, 12, 519–539. [Google Scholar] [CrossRef] [Green Version]
- Koga, S.; Williams, D.S.; Perriman, A.W.; Mann, S. Peptide-nucleotide microdroplets as a step towards a membrane-free protocell model. Nat. Chem. 2011, 3, 720–724. [Google Scholar] [CrossRef]
- Cakmak, F.P.; Choi, S.; Meyer, M.C.O.; Bevilacqua, P.C.; Keating, C.D. Prebiotically-relevant low polyion multivalency can improve functionality of membraneless compartments. Nat. Commun. 2020, 11, 5949. [Google Scholar] [CrossRef]
- Long, M.S.; Jones, C.D.; Helfrich, M.R.; Mangeney-Slavin, L.K.; Keating, C.D. Dynamic microcompartmentation in synthetic cells. Proc. Natl. Acad. Sci. USA 2005, 102, 5920–5925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, A.C.; Steinkühler, J.; Gonzales, D.T.; Robinson, T.; Dimova, R.; Dora, T.Y. Reversible pH responsive coacervate formation in lipid vesicles activates dormant enzymatic reactions. Angew. Chemie Int. Ed. 2020, 59, 5950–5957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fares, H.M.; Marras, A.E.; Ting, J.M.; Tirrell, M.V.; Keating, C.D. Impact of wet-dry cycling on the phase behavior and compartmentalization properties of complex coacervates. Nat. Commun. 2020, 11, 5423. [Google Scholar] [CrossRef]
- Zwicker, D.; Seyboldt, R.; Weber, C.A.; Hyman, A.A.; Jülicher, F. Growth and division of active droplets provides a model for protocells. Nat. Phys. 2017, 13, 408–413. [Google Scholar] [CrossRef]
- Donau, C.; Späth, F.; Sosson, M.; Kriebisch, B.A.K.; Schnitter, F.; Tena-Solsona, M.; Kang, H.S.; Salibi, E.; Sattler, M.; Mutschler, H.; et al. Active coacervate droplets as a model for membraneless organelles and protocells. Nat. Commun. 2020, 11, 5167. [Google Scholar] [CrossRef] [PubMed]
- Jia, T.Z.; Hentrich, C.; Szostak, J.W. Rapid RNA exchange in aqueous two-phase system and coacervate droplets. Orig. Life Evol. Biosph. 2014, 44, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poudyal, R.R.; Guth-Metzler, R.M.; Veenis, A.J.; Frankel, E.A.; Keating, C.D.; Bevilacqua, P.C. Template-directed RNA polymerization and enhanced ribozyme catalysis inside membraneless compartments formed by coacervates. Nat. Commun. 2019, 10, 490. [Google Scholar] [CrossRef]
- Poudyal, R.R.; Keating, C.D.; Bevilacqua, P.C. Polyanion-assisted ribozyme catalysis inside complex coacervates. ACS Chem. Biol. 2019, 14, 1243–1248. [Google Scholar] [CrossRef]
- Trevors, J.T.; Pollack, G.H. Hypothesis: The origin of life in a hydrogel environment. Prog. Biophys. Mol. Biol. 2005, 89, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Peng, S.; Hartman, M.R.; Gupton-Campolongo, T.; Rice, E.J.; Chang, A.K.; Gu, Z.; Lu, G.Q.; Luo, D. Enhanced transcription and translation in clay hydrogel and implications for early life evolution. Sci. Rep. 2013, 3, 3165. [Google Scholar] [CrossRef]
- Jia, T.Z.; Chandru, K.; Hongo, Y.; Afrin, R.; Usui, T.; Myojo, K.; James Cleaves, H. Membraneless polyester microdroplets as primordial compartments at the origins of life. Proc. Natl. Acad. Sci. USA 2019, 116, 15830–15835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, T.Z.; Fraccia, T.P. Liquid crystal peptide/DNA coacervates in the context of prebiotic molecular evolution. Crystals 2020, 10, 964. [Google Scholar] [CrossRef]
- Frommer, M.A.; Miller, I.R. Adsorption of DNA at the air-water interface. J. Phys. Chem. 1968, 72, 2862–2866. [Google Scholar] [CrossRef]
- Eickbush, T.H.; Moudrianakis, E.N. A mechanism for the entrapment of DNA at an air-water interface. Biophys. J. 1977, 18, 275–288. [Google Scholar] [CrossRef] [Green Version]
- Woose, C.R. A proposal concerning the origin of life on the planet Earth. J. Mol. Evol. 1979, 13, 95–101. [Google Scholar] [CrossRef]
- Griffith, E.C.; Tuck, A.F.; Vaida, V. Ocean–atmosphere interactions in the emergence of complexity in simple chemical systems. Acc. Chem. Res. 2012, 45, 2106–2113. [Google Scholar] [CrossRef]
- Martin, W.; Baross, J.; Kelley, D.; Russell, M.J. Hydrothermal vents and the origin of life. Nat. Rev. Microbiol. 2008, 6, 805–814. [Google Scholar] [CrossRef]
- Sojo, V.; Herschy, B.; Whicher, A.; Camprubí, E.; Lane, N. The origin of life in alkaline hydrothermal vents. Astrobiology 2016, 16, 181–197. [Google Scholar] [CrossRef] [PubMed]
- Kelley, D.S.; Karson, J.A.; Blackman, D.K.; Früh-Green, G.L.; Butterfield, D.A.; Lilley, M.D.; Olson, E.J.; Schrenk, M.O.; Roe, K.K.; Lebon, G.T.; et al. An off-axis hydrothermal vent field near the mid-atlantic ridge at 30° n. Nature 2001, 412, 145–149. [Google Scholar] [CrossRef]
- Kelley, D.S.; Karson, J.A.; Früh-Green, G.L.; Yoerger, D.R.; Shank, T.M.; Butterfield, D.A.; Hayes, J.M.; Schrenk, M.O.; Olson, E.J.; Proskurowski, G.; et al. A serpentinite-hosted ecosystem: The Lost City hydrothermal field. Science 2005, 307, 1428–1434. [Google Scholar] [CrossRef]
- Russell, M.J.; Daniel, R.M.; Hall, A.J.; Sherringham, J.A. A hydrothermally precipitated catalytic iron sulphide membrane as a first step toward life. J. Mol. Evol. 1994, 39, 231–243. [Google Scholar] [CrossRef]
- Martin, W.; Russell, M.J.; Horner, D.; Blankenship, R.; Cavalier-Smith, T.; Nisbet, E. On the origins of cells: A hypothesis for the evolutionary transitions from abiotic geochemistry to chemoautotrophic prokaryotes, and from prokaryotes to nucleated cells. Philos. Trans. R. Soc. B Biol. Sci. 2003, 358, 59–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koonin, E.V.; Martin, W. On the origin of genomes and cells within inorganic compartments. Trends Genet. 2005, 21, 647–654. [Google Scholar] [CrossRef]
- Baaske, P.; Weinert, F.M.; Duhr, S.; Lemke, K.H.; Russell, M.J.; Braun, D. Extreme accumulation of nucleotides in simulated hydrothermal pore systems. Proc. Natl. Acad. Sci. USA 2007, 104, 9346–9351. [Google Scholar] [CrossRef] [Green Version]
- Mast, C.B.; Schink, S.; Gerland, U.; Braun, D. Escalation of polymerization in a thermal gradient. Proc. Natl. Acad. Sci. USA 2013, 110, 8030–8035. [Google Scholar] [CrossRef] [Green Version]
- Salditt, A.; Keil, L.M.R.; Horning, D.P.; Mast, C.B.; Joyce, G.F.; Braun, D. Thermal habitat for RNA amplification and accumulation. Phys. Rev. Lett. 2020, 125, 048104. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Steinbock, O. Materials synthesis and catalysis in microfluidic devices: Prebiotic chemistry in mineral membranes. ChemCatChem 2020, 12, 63–74. [Google Scholar] [CrossRef] [Green Version]
- Ogasawara, H.; Yoshida, A.; Imai, E.I.; Honda, H.; Hatori, K.; Matsuno, K. Synthesizing oligomers from monomeric nucleotides in simulated hydrothermal environments. Orig. Life Evol. Biosph. 2000, 30, 519–526. [Google Scholar] [CrossRef] [PubMed]
- Burcar, B.T.; Barge, L.M.; Trail, D.; Watson, E.B.; Russell, M.J.; McGown, L.B. RNA oligomerization in laboratory analogues of alkaline hydrothermal vent systems. Astrobiology 2015, 15, 509–522. [Google Scholar] [CrossRef] [PubMed]
- Pir Cakmak, F.; Keating, C.D. Combining catalytic microparticles with droplets formed by phase coexistence: Adsorption and activity of natural clays at the aqueous/aqueous interface. Sci. Rep. 2017, 7, 3215. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class (and Major Mechanism for Separating Non-Parasitic Molecules from Parasites) | Elements for Compartmentalization | Boundary | Additional Mechanism that Contributes to Segregating Parasitic Molecules | Mechanism of Substrate Uptake from Environments | Selective Accumulation of Longer Genetic Molecules | Selective Replication of Non-Parasitic Genetic Molecules | Possible Transition to Lipid Vesicles | |
|---|---|---|---|---|---|---|---|---|
| Using an Analogous Computational Model | In a Laboratory Experiment | |||||||
| 1. Without boundaries (limited diffusion) | Mineral surfaces | Diffusion | Yes [22,23] | Yes [24,25,26,27,28,29,30,31,32,33] | Mineral-assisted [34,35,36] | |||
| Active fluid environments | Apparent group formation | Diffusion | Yes [37] | |||||
| Crowded environments | Apparent group formation | Diffusion | Yes [38] | |||||
| 2. With boundaries (group formation) | Ice eutectic phases | Ice | Limited diffusion | Disruption and reformation | Yes [20,29,39,40,41,42,43,44,45,46,47], but mostly by using general protocell models that have one or more features of each compartmentalizaiton with boundaries | Freeze-thaw-assisted [48,49] | ||
| Membranous compartments 1 | Membranes | Diffusion through semi-permeable membranes | ||||||
| Membraneless cell-like compartments 1 | Distinct aqueous phases | Diffusion through semi-permeable interfaces | Yes [50,51] | Yes [52] | Interface-assisted [53,54] | |||
| Gas bubbles | Aqueous phases | Diffusion | Likely 2 [55] | Interface-assisted [55] | ||||
| Atmospheric compartments 1 | Atmospheric gas phases | Disruption and reformation | Interface-assisted [56] | |||||
| 3. With incomplete boundaries (limited group formation) | Porous structures of hydrothermal vents | Minerals | Diffusion through incomplete boundaries | Yes [57] | Yes [47,58] | Thermal gradient-assisted [59] | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mizuuchi, R.; Ichihashi, N. Primitive Compartmentalization for the Sustainable Replication of Genetic Molecules. Life 2021, 11, 191. https://doi.org/10.3390/life11030191
Mizuuchi R, Ichihashi N. Primitive Compartmentalization for the Sustainable Replication of Genetic Molecules. Life. 2021; 11(3):191. https://doi.org/10.3390/life11030191
Chicago/Turabian StyleMizuuchi, Ryo, and Norikazu Ichihashi. 2021. "Primitive Compartmentalization for the Sustainable Replication of Genetic Molecules" Life 11, no. 3: 191. https://doi.org/10.3390/life11030191
APA StyleMizuuchi, R., & Ichihashi, N. (2021). Primitive Compartmentalization for the Sustainable Replication of Genetic Molecules. Life, 11(3), 191. https://doi.org/10.3390/life11030191