
Recent Progress Using De Novo Design to Study Protein Structure, Design and Binding Interactions

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
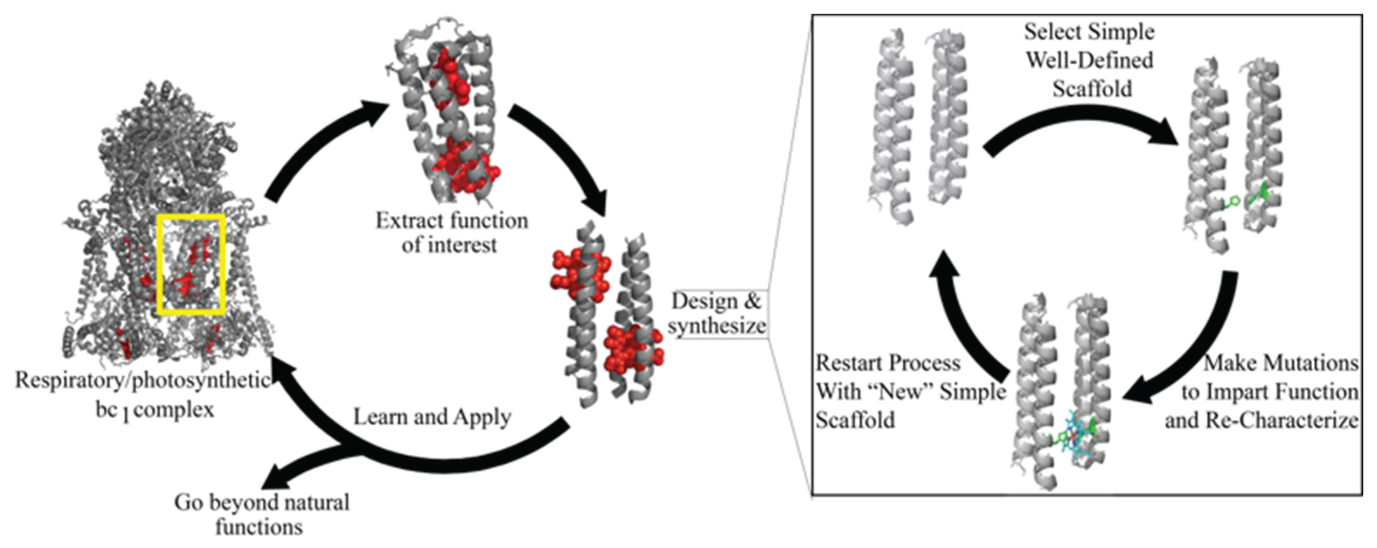
:1. Introduction
2. Small Molecule Binding Proteins
2.1. Metal Binding Proteins and Related Functions
2.2. Porphyrin Cofactors
2.3. Hydrophobic Molecules
3. Transmembrane Proteins
4. Design of DNA/RNA Binding Proteins
5. Packing of the Hydrophobic Core and Structural Stability
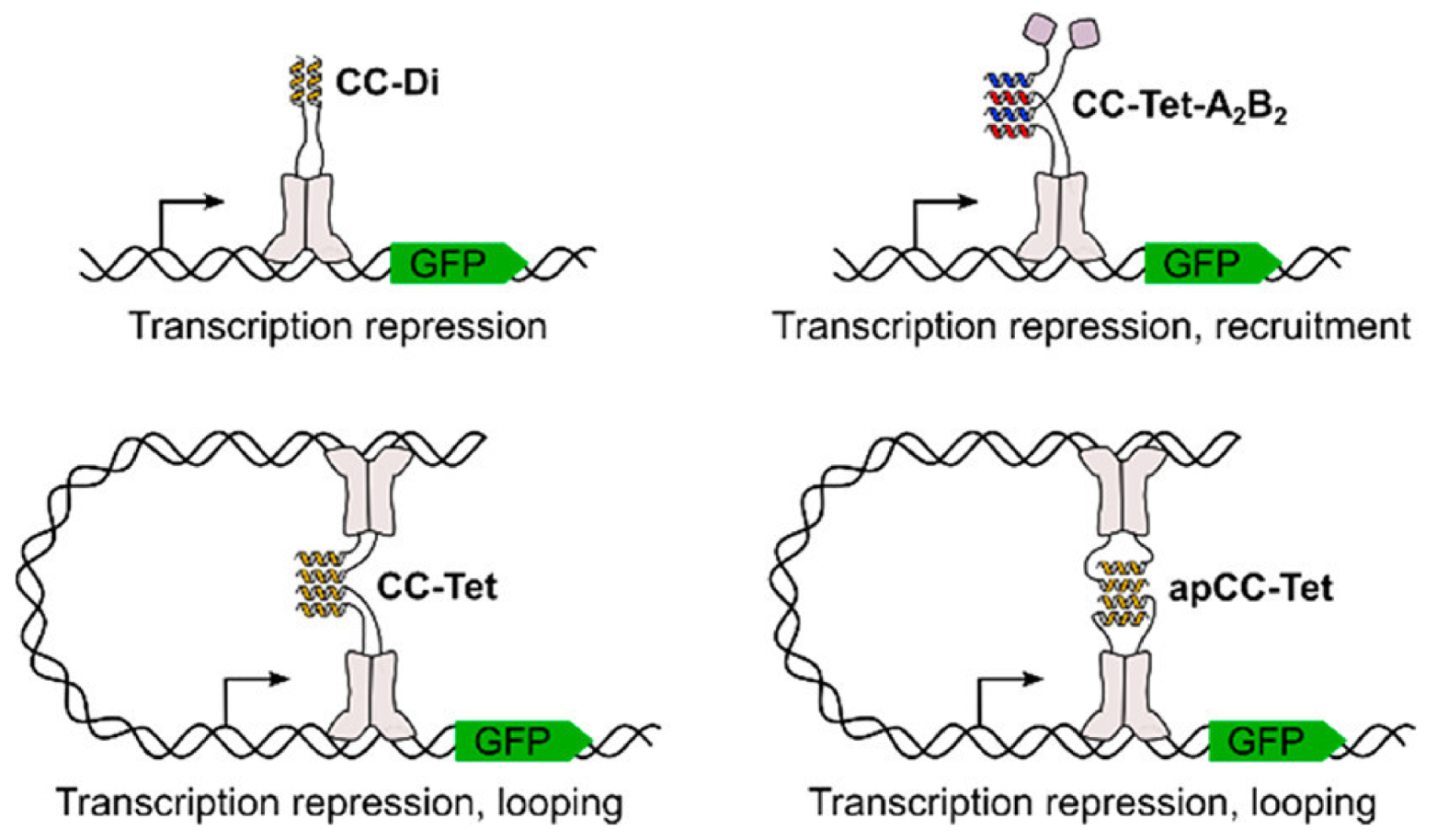
6. Design of Protein Interfaces
6.1. Protein-Protein Interactions
6.2. Antimicrobial Peptides and Other Therapeutics
6.3. Using Protein Design to Combat COVID-19
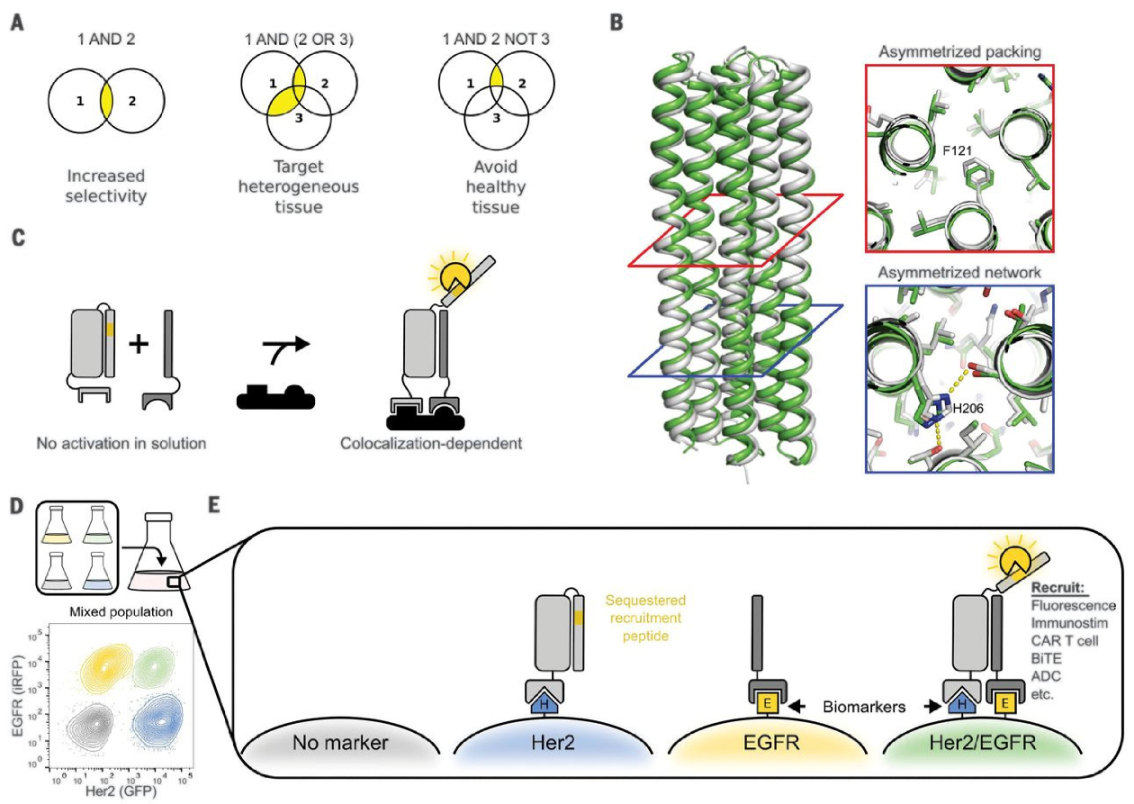
6.4. Using De Novo Designed PPIs to Fight Cancer
6.5. Methods of Identifying PPIs
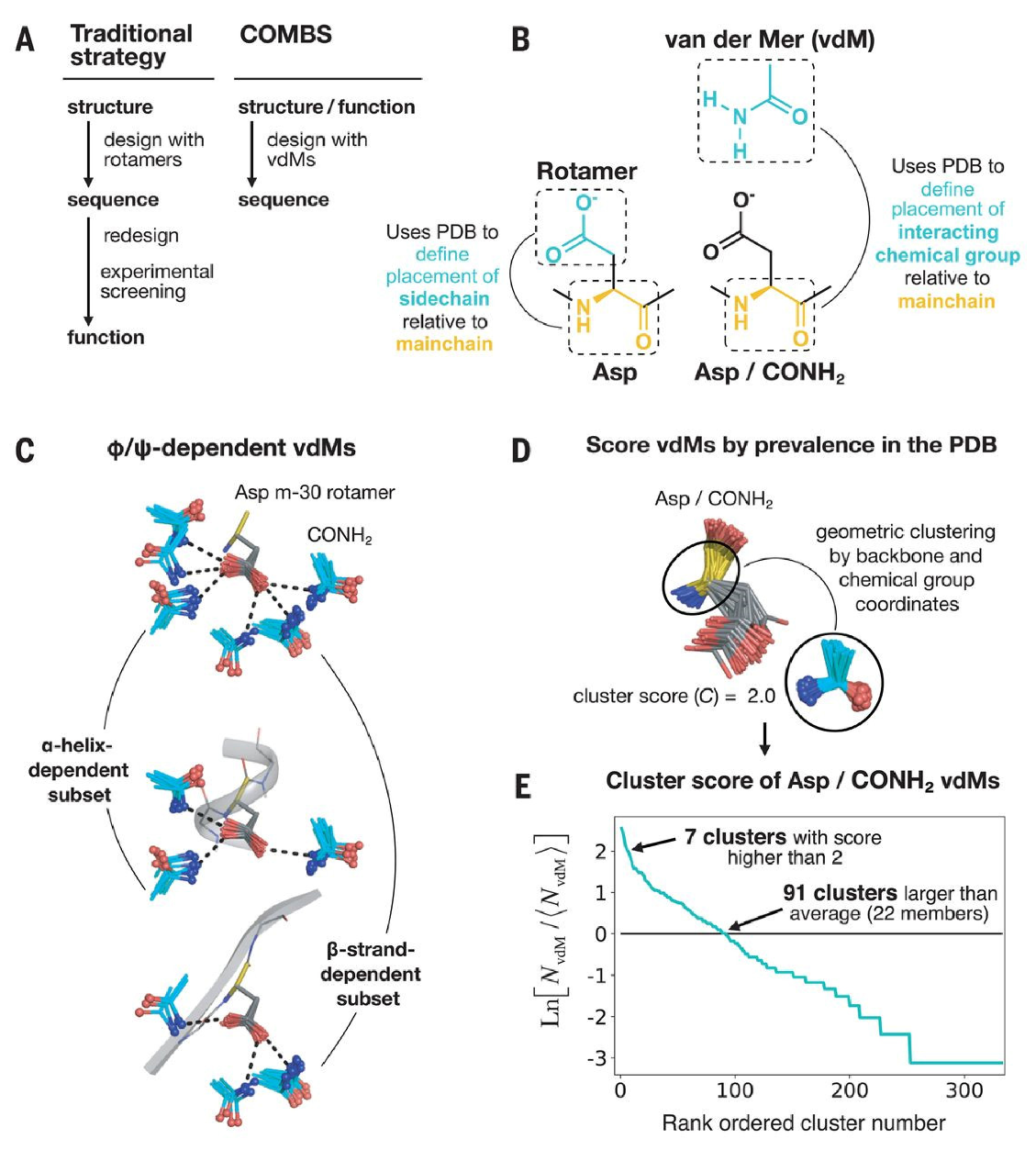
7. New Algorithms for Protein Design
8. Design from the Perspective of the Binding Pair
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Vuignier, K.; Schappler, J.; Veuthey, J.-L.; Carrupt, P.-A.; Martel, S. Drug–protein binding: A critical review of analytical tools. Anal. Bioanal. Chem. 2010, 398, 53–66. [Google Scholar] [CrossRef] [PubMed]
- Siggers, T.; Gordân, R. Protein–DNA binding: Complexities and multi-protein codes. Nucleic Acids Res. 2014, 42, 2099–2111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, Y.; Walsh, C.A. Protein–Protein interactions, cytoskeletal regulation and neuronal migration. Nat. Rev. Neurosci. 2001, 2, 408–416. [Google Scholar] [CrossRef] [PubMed]
- Dutton, P.L.; Moser, C.C. Engineering enzymes. Faraday Discuss. 2011, 148, 443–448. [Google Scholar] [CrossRef] [PubMed]
- Lichtenstein, B.R.; Farid, T.A.; Kodali, G.; Solomon, L.A.; Anderson, J.R.; Sheehan, M.M.; Ennist, N.M.; Fry, B.A.; Chobot, S.E.; Bialas, C.; et al. Engineering oxidoreductases: Maquette proteins designed from scratch. Biochem. Soc. Trans. 2012, 40, 561–566. [Google Scholar] [CrossRef] [PubMed]
- Macdonald, J.R.; Johnson, W.C. Environmental features are important in determining protein secondary structure. Protein Sci. 2001, 10, 1172–1177. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.-S.; Cobessi, D.; Tung, E.Y.; Berry, E.A. Binding of the Respiratory Chain Inhibitor Antimycin to the Mitochondrial bc1 Complex: A New Crystal Structure Reveals an Altered Intramolecular Hydrogen-bonding Pattern. J. Mol. Biol. 2005, 351, 573–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, S.P.; DeGrado, W.F. Design of a 4-helix bundle protein: Synthesis of peptides which self-associate into a helical protein. J. Am. Chem. Soc. 1987, 109, 6751–6758. [Google Scholar] [CrossRef]
- Regan, L.; DeGrado, W.F.; Landegren, U.; Kaiser, R.; Caskey, C.; Hood, L. Characterization of a helical protein designed from first principles. Science 1988, 241, 976–978. [Google Scholar] [CrossRef]
- Koder, R.L.; Anderson, J.L.R.; Solomon, L.A.; Reddy, K.S.; Moser, C.C.; Dutton, P.L. Design and engineering of an O2 transport protein. Nat. Cell Biol. 2009, 458, 305–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, L.; Althoff, E.A.; Clemente, F.R.; Doyle, L.; Röthlisberger, D.; Zanghellini, A.; Gallaher, J.L.; Betker, J.L.; Tanaka, F.; Barbas, C.F.; et al. De Novo Computational Design of Retro-Aldol Enzymes. Science 2008, 319, 1387–1391. [Google Scholar] [CrossRef] [Green Version]
- Anderson, J.L.R.; Armstrong, C.T.; Kodali, G.; Lichtenstein, B.R.; Watkins, D.W.; Mancini, J.A.; Boyle, A.L.; Farid, T.A.; Crump, M.P.; Moser, C.C.; et al. Constructing a man-made c-type cytochrome maquette in vivo: Electron transfer, oxygen transport and conversion to a photoactive light harvesting maquette. Chem. Sci. 2014, 5, 507–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farid, A.T.; Kodali, G.; Solomon, A.L.; Lichtenstein, B.R.; Sheehan, M.M.; Fry, A.B.; Bialas, C.; Ennist, N.M.; Siedlecki, A.J.; Zhao, Z.; et al. Elementary tetrahelical protein design for diverse oxidoreductase functions. Nat. Chem. Biol. 2013, 9, 826–833. [Google Scholar] [CrossRef] [Green Version]
- Currin, A.; Swainston, N.; Day, P.J.; Kell, D.B. Synthetic biology for the directed evolution of protein biocatalysts: Navigating sequence space intelligently. Chem. Soc. Rev. 2015, 44, 1172–1239. [Google Scholar] [CrossRef] [Green Version]
- Packer, M.S.; Liu, D.R. Methods for the directed evolution of proteins. Nat. Rev. Genet. 2015, 16, 379–394. [Google Scholar] [CrossRef]
- Neylon, C. Chemical and biochemical strategies for the randomization of protein encoding DNA sequences: Library construction methods for directed evolution. Nucleic Acids Res. 2004, 32, 1448–1459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kan, S.B.J.; Lewis, R.D.; Chen, K.; Arnold, F.H. Directed evolution of cytochrome c for carbon–silicon bond formation: Bringing silicon to life. Science 2016, 354, 1048–1051. [Google Scholar] [CrossRef] [Green Version]
- Karanicolas, J.; Corn, J.E.; Chen, I.; Joachimiak, L.A.; Dym, O.; Peck, S.H.; Albeck, S.; Unger, T.; Hu, W.; Liu, G.; et al. A De Novo Protein Binding Pair By Computational Design and Directed Evolution. Mol. Cell 2011, 42, 250–260. [Google Scholar] [CrossRef] [Green Version]
- Grayson, K.J.; Anderson, J.L.R. Designed for life: Biocompatible de novo designed proteins and components. J. R. Soc. Interface 2018, 15, 20180472. [Google Scholar] [CrossRef]
- Huang, P.-S.; Boyken, S.E.; Baker, P.-S.H.S.E.B.D. The coming of age of de novo protein design. Nat. Cell Biol. 2016, 537, 320–327. [Google Scholar] [CrossRef] [PubMed]
- Korendovych, I.V.; DeGrado, W.F. De novoprotein design, a retrospective. Q. Rev. Biophys. 2020, 53, e3. [Google Scholar] [CrossRef]
- Calhoun, J.R.; Nastri, F.; Maglio, O.; Pavone, V.; Lombardi, A.; DeGrado, W.F. Artificial diiron proteins: From structure to function. Biopolymer 2005, 80, 264–278. [Google Scholar] [CrossRef]
- Maglio, O.; Nastri, F.; De Rosales, R.T.M.; Faiella, M.; Pavone, V.; DeGrado, W.F.; Lombardi, A. Diiron-containing metalloproteins: Developing functional models. Comptes Rendus Chim. 2007, 10, 703–720. [Google Scholar] [CrossRef]
- Kaplan, J.; DeGrado, W.F. De novo design of catalytic proteins. Proc. Natl. Acad. Sci. USA 2004, 101, 11566–11570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paredes, A.; Loh, B.M.; Peduzzi, O.M.; Reig, A.J.; Buettner, K.M. DNA Cleavage by a De Novo Designed Protein–Titanium Complex. Inorg. Chem. 2020, 59, 11248–11252. [Google Scholar] [CrossRef] [PubMed]
- Olson, T.L.; Espiritu, E.; Edwardraja, S.; Canarie, E.; Flores, M.; Williams, J.C.; Ghirlanda, G.; Allen, J.P. Biochemical and spectroscopic characterization of dinuclear Mn-sites in artificial four-helix bundle proteins. Biochim. Biophys. Acta (BBA) Bioenerg. 2017, 1858, 945–954. [Google Scholar] [CrossRef]
- Chino, M.; Zhang, S.-Q.; Pirro, F.; Leone, L.; Maglio, O.; Lombardi, A.; DeGrado, W.F. Spectroscopic and metal binding properties of a de novo metalloprotein binding a tetrazinc cluster. Biopolymer 2018, 109, e23339. [Google Scholar] [CrossRef]
- Zhang, S.-Q.; Chino, M.; Liu, L.; Tang, Y.; Hu, X.; DeGrado, W.F.; Lombardi, A. De Novo Design of Tetranuclear Transition Metal Clusters Stabilized by Hydrogen-Bonded Networks in Helical Bundles. J. Am. Chem. Soc. 2018, 140, 1294–1304. [Google Scholar] [CrossRef] [Green Version]
- Pirro, F.; Schmidt, N.; Lincoff, J.; Widel, Z.X.; Polizzi, N.F.; Liu, L.; Therien, M.J.; Grabe, M.; Chino, M.; Lombardi, A.; et al. Allosteric cooperation in a de novo-designed two-domain protein. Proc. Natl. Acad. Sci. USA 2020, 117, 33246–33253. [Google Scholar] [CrossRef]
- Mancini, J.A.; Pike, D.H.; Tyryshkin, A.M.; Haramaty, L.; Wang, M.S.; Poudel, S.; Hecht, M.; Nanda, V. Design of a Fe 4 S 4 cluster into the core of a de novo four-helix bundle. Biotechnol. Appl. Biochem. 2020, 67, 574–585. [Google Scholar] [CrossRef] [PubMed]
- Selvan, D.; Prasad, P.; Farquhar, E.R.; Shi, Y.; Crane, S.; Zhang, Y.; Chakraborty, S. Redesign of a Copper Storage Protein into an Artificial Hydrogenase. ACS Catal. 2019, 9, 5847–5859. [Google Scholar] [CrossRef]
- Mutter, A.C.; Tyryshkin, A.M.; Campbell, I.J.; Poudel, S.; Bennett, G.N.; Silberg, J.J.; Nanda, V.; Falkowski, P.G. De novo design of symmetric ferredoxins that shuttle electrons in vivo. Proc. Natl. Acad. Sci. USA 2019, 116, 14557–14562. [Google Scholar] [CrossRef] [Green Version]
- Boyken, S.E.; Benhaim, M.A.; Busch, F.; Jia, M.; Bick, M.J.; Choi, H.; Klima, J.C.; Chen, Z.; Walkey, C.; Mileant, A.; et al. De novo design of tunable, pH-driven conformational changes. Science 2019, 364, 658–664. [Google Scholar] [CrossRef]
- Robertson, D.E.; Farid, R.S.; Moser, C.C.; Urbauer, J.L.; Mulholland, S.E.; Pidikiti, R.; Lear, J.D.; Wand, A.J.; DeGrado, W.F.; Dutton, P.L. Design and synthesis of multi-haem proteins. Nat. Cell Biol. 1994, 368, 425–432. [Google Scholar] [CrossRef]
- Sykes, A.G. Advances in Inorganic Chemistry: Heme-Fe Proteins; Academic Press: San Diego, CA, USA, 2000; Volume 51. [Google Scholar]
- Nagarajan, D.; Sukumaran, S.; Deka, G.; Krishnamurthy, K.; Atreya, H.S.; Chandra, N. Design of a heme-binding peptide motif adopting a β-hairpin conformation. J. Biol. Chem. 2018, 293, 9412–9422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zambrano, G.; Nastri, F.; Pavone, V.; Lombardi, A.; Chino, M. Use of an Artificial Miniaturized Enzyme in Hydrogen Peroxide Detection by Chemiluminescence. Sensors 2020, 20, 3793. [Google Scholar] [CrossRef]
- Polizzi, N.F.; Wu, Y.; Lemmin, T.; Maxwell, A.M.; Zhang, S.-Q.; Rawson, J.; Beratan, D.N.; Therien, M.J.; DeGrado, W.F. De novo design of a hyperstable non-natural protein–ligand complex with sub-Å accuracy. Nat. Chem. 2017, 9, 1157–1164. [Google Scholar] [CrossRef] [Green Version]
- Kodali, G.; Mancini, J.A.; Solomon, L.A.; Episova, T.V.; Roach, N.; Hobbs, C.J.; Wagner, P.; Mass, O.A.; Aravindu, K.; Barnsley, J.E.; et al. Design and engineering of water-soluble light-harvesting protein maquettes. Chem. Sci. 2016, 8, 316–324. [Google Scholar] [CrossRef] [Green Version]
- Polizzi, N.F.; DeGrado, W.F. A defined structural unit enables de novo design of small-molecule–binding proteins. Science 2020, 369, 1227–1233. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Selvaraj, B.; McShan, A.C.; Boyken, S.E.; Wei, K.Y.; Oberdorfer, G.; DeGrado, W.; Sgourakis, N.G.; Cuneo, M.J.; Myles, D.A.; et al. De novo design of a homo-trimeric amantadine-binding protein. eLife 2019, 8, 47839. [Google Scholar] [CrossRef]
- Dou, J.; Vorobieva, A.A.; Sheffler, W.; Doyle, L.A.; Park, H.; Bick, M.J.; Mao, B.; Foight, G.W.; Lee, M.Y.; Gagnon, L.A.; et al. De novo design of a fluorescence-activating β-barrel. Nat. Cell Biol. 2018, 561, 485–491. [Google Scholar] [CrossRef] [PubMed]
- Thomas, F.; Dawson, W.M.; Lang, E.J.M.; Burton, A.J.; Bartlett, G.J.; Rhys, G.G.; Mulholland, A.J.; Woolfson, D.N. De Novo-Designed α-Helical Barrels as Receptors for Small Molecules. ACS Synth. Biol. 2018, 7, 1808–1816. [Google Scholar] [CrossRef] [Green Version]
- Caldwell, S.J.; Haydon, I.C.; Piperidou, N.; Huang, P.-S.; Bick, M.J.; Sjöström, H.S.; Hilvert, D.; Baker, D.; Zeymer, C. Tight and specific lanthanide binding in a de novo TIM barrel with a large internal cavity designed by symmetric domain fusion. Proc. Natl. Acad. Sci. USA 2020, 117, 30362–30369. [Google Scholar] [CrossRef]
- Karas, C.; Hecht, M. A Strategy for Combinatorial Cavity Design in De Novo Proteins. Life 2020, 10, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, S.; Davidsen, K.; Gomez-Castillo, L.; Jiang, H.; Fu, X.; Li, Z.; Liang, Y.; Jahn, M.; Moussa, M.; DiMaio, F.; et al. COMBINES-CID: An Efficient Method for De Novo Engineering of Highly Specific Chemically Induced Protein Dimerization Systems. J. Am. Chem. Soc. 2019, 141, 10948–10952. [Google Scholar] [CrossRef] [PubMed]
- Prakash, V.; Ranbhor, R.; Ramakrishnan, V. De Novo Designed Heterochiral Blue Fluorescent Protein. ACS Omega 2020, 5, 26382–26388. [Google Scholar] [CrossRef] [PubMed]
- Stenner, R.; Steventon, J.W.; Seddon, A.; Anderson, J.L.R. A de novo peroxidase is also a promiscuous yet stereoselective carbene transferase. Proc. Natl. Acad. Sci. USA 2020, 117, 1419–1428. [Google Scholar] [CrossRef] [Green Version]
- Stenner, R.; Anderson, J.L.R. Chemoselective N−H insertion catalyzed by a de novo carbene transferase. Biotechnol. Appl. Biochem. 2020, 67, 527–535. [Google Scholar] [CrossRef]
- Gromiha, M.M.; Ou, Y.-Y. Bioinformatics approaches for functional annotation of membrane proteins. Brief. Bioinform. 2013, 15, 155–168. [Google Scholar] [CrossRef] [Green Version]
- Cournia, Z.; Allen, T.W.; Andricioaei, I.; Antonny, B.; Baum, D.; Brannigan, G.; Buchete, N.-V.; Deckman, J.T.; Delemotte, L.; Del Val, C.; et al. Membrane Protein Structure, Function, and Dynamics: A Perspective from Experiments and Theory. J. Membr. Biol. 2015, 248, 611–640. [Google Scholar] [CrossRef] [Green Version]
- Engel, A.; Gaub, H.E. Structure and Mechanics of Membrane Proteins. Annu. Rev. Biochem. 2008, 77, 127–148. [Google Scholar] [CrossRef] [Green Version]
- Mravic, M.; Thomaston, J.L.; Tucker, M.; Solomon, P.E.; Liu, L.; DeGrado, W.F. Packing of apolar side chains enables accurate design of highly stable membrane proteins. Science 2019, 363, 1418–1423. [Google Scholar] [CrossRef] [PubMed]
- Curnow, P.; Hardy, B.J.; Dufour, V.; Arthur, C.J.; Stenner, R.; Hodgson, L.R.; Verkade, P.; Williams, C.; Shoemark, D.K.; Sessions, R.B.; et al. Small-residue packing motifs modulate the structure and function of a minimal de novo membrane protein. Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Dong, J.; Viviani, M.; Tulini, I.; Pontillo, N.; Maity, S.; Zhou, Y.; Roos, W.H.; Liu, K.; Herrmann, A.; et al. De novo rational design of a freestanding, supercharged polypeptide, proton-conducting membrane. Sci. Adv. 2020, 6, eabc0810. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Lu, P.; El-Din, T.M.G.; Pei, X.Y.; Johnson, M.C.; Uyeda, A.; Bick, M.J.; Xu, Q.; Jiang, D.; Bai, H.; et al. Computational design of transmembrane pores. Nat. Cell Biol. 2020, 585, 129–134. [Google Scholar] [CrossRef]
- Rohs, R.; Jin, X.; West, S.M.; Joshi, R.; Honig, B.; Mann, R.S. Origins of Specificity in Protein-DNA Recognition. Annu. Rev. Biochem. 2010, 79, 233–269. [Google Scholar] [CrossRef] [Green Version]
- Corley, M.; Burns, M.C.; Yeo, G.W. How RNA-Binding Proteins Interact with RNA: Molecules and Mechanisms. Mol. Cell 2020, 78, 9–29. [Google Scholar] [CrossRef]
- Inamoto, I.; Sheoran, I.; Popa, S.C.; Hussain, M.; Shin, J.A. Combining Rational Design and Continuous Evolution on Minimalist Proteins That Target the E-box DNA Site. ACS Chem. Biol. 2021, 16, 35–44. [Google Scholar] [CrossRef]
- Lebar, T.; Lainšček, D.; Merljak, E.; Aupič, J.; Jerala, R. A tunable orthogonal coiled-coil interaction toolbox for engineering mammalian cells. Nat. Chem. Biol. 2020, 16, 513–519. [Google Scholar] [CrossRef] [PubMed]
- Walker, M.J.; Varani, G. Design of RNA-targeting macrocyclic peptides. Methods Enzym. 2019, 623, 339–372. [Google Scholar] [CrossRef]
- Smith, A.J.; Thomas, F.; Shoemark, D.; Woolfson, D.N.; Savery, N.J. Guiding Biomolecular Interactions in Cells Using de Novo Protein–Protein Interfaces. ACS Synth. Biol. 2019, 8, 1284–1293. [Google Scholar] [CrossRef]
- Edgell, C.L.; Smith, A.J.; Beesley, J.L.; Savery, N.J.; Woolfson, D.N. De NovoDesigned Protein-Interaction Modules for In-Cell Applications. ACS Synth. Biol. 2020, 9, 427–436. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, C.; Young, J.T.; Slade, G.G.; Oliveira, R.J.; McCully, M.E. A Dynamic Hydrophobic Core and Surface Salt Bridges Thermostabilize a Designed Three-Helix Bundle. Biophys. J. 2019, 116, 621–632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koga, R.; Yamamoto, M.; Kosugi, T.; Kobayashi, N.; Sugiki, T.; Fujiwara, T.; Koga, N. Robust folding of a de novo designed ideal protein even with most of the core mutated to valine. Proc. Natl. Acad. Sci. USA 2020, 117, 31149–31156. [Google Scholar] [CrossRef] [PubMed]
- Banach, M.; Fabian, P.; Stapor, K.; Konieczny, L.; Roterman, A.I. Structure of the Hydrophobic Core Determines the 3D Protein Structure—Verification by Single Mutation Proteins. Biomolecules 2020, 10, 767. [Google Scholar] [CrossRef]
- Kimura, N.; Mochizuki, K.; Umezawa, K.; Hecht, M.H.; Arai, R. Hyperstable De Novo Protein with a Dimeric Bisecting Topology. ACS Synth. Biol. 2019, 9, 254–259. [Google Scholar] [CrossRef]
- Edgell, C.L.; Savery, N.J.; Woolfson, D.N. Robust De Novo-Designed Homotetrameric Coiled Coils. Biochemistry 2020, 59, 1087–1092. [Google Scholar] [CrossRef]
- Chen, Z.; Johnson, M.C.; Chen, J.; Bick, M.J.; Boyken, S.E.; Lin, B.; De Yoreo, J.J.; Kollman, J.M.; Baker, D.; DiMaio, F. Self-Assembling 2D Arrays with de Novo Protein Building Blocks. J. Am. Chem. Soc. 2019, 141, 8891–8895. [Google Scholar] [CrossRef]
- Pagel, P.; Kovac, S.; Oesterheld, M.; Brauner, B.; Dunger-Kaltenbach, I.; Frishman, D.; Montrone, C.; Mark, P.; Stümpflen, V.; Mewes, H.-W.; et al. The MIPS mammalian protein-protein interaction database. Bioinformatics 2004, 21, 832–834. [Google Scholar] [CrossRef] [Green Version]
- Langan, R.A.; Boyken, S.E.; Ng, A.H.; Samson, J.A.; Dods, G.; Westbrook, A.M.; Nguyen, T.H.; Lajoie, M.J.; Chen, Z.; Berger, S.; et al. De novo design of bioactive protein switches. Nat. Cell Biol. 2019, 572, 205–210. [Google Scholar] [CrossRef]
- Lajoie, M.J.; Boyken, S.E.; Salter, A.I.; Bruffey, J.; Rajan, A.; Langan, R.A.; Olshefsky, A.; Muhunthan, V.; Bick, M.J.; Gewe, M.; et al. Designed protein logic to target cells with precise combinations of surface antigens. Science 2020, 369, eaba6527. [Google Scholar] [CrossRef]
- Xie, M.; Lu, P. When de novo-designed protein logics meet CAR-T therapies. Cell Res. 2020, 30, 946–947. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Kibler, R.D.; Hunt, A.; Busch, F.; Pearl, J.; Jia, M.; VanAernum, Z.L.; Wicky, B.I.M.; Dods, G.; Liao, H.; et al. De novo design of protein logic gates. Science 2020, 368, 78–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glasgow, A.A.; Huang, Y.-M.; Mandell, D.J.; Thompson, M.; Ritterson, R.; Loshbaugh, A.L.; Pellegrino, J.; Krivacic, C.; Pache, R.A.; Barlow, K.A.; et al. Computational design of a modular protein sense-response system. Science 2019, 366, 1024–1028. [Google Scholar] [CrossRef]
- Schnatz, P.J.; Brisendine, J.M.; Laing, C.C.; Everson, B.H.; French, C.A.; Molinaro, P.M.; Koder, R.L. Designing heterotropically activated allosteric conformational switches using supercharging. Proc. Natl. Acad. Sci. USA 2020, 117, 5291–5297. [Google Scholar] [CrossRef] [PubMed]
- Chevalier, A.; Silva, D.-A.; Rocklin, G.J.; Hicks, D.R.; Vergara, R.; Murapa, P.; Bernard, S.M.; Zhang, L.; Lam, K.-H.; Yao, G.; et al. Massively parallel de novo protein design for targeted therapeutics. Nature 2017, 550, 74–79. [Google Scholar] [CrossRef]
- Wang, Y.; Fan, Y.; Zhou, Z.; Tu, H.; Ren, Q.; Wang, X.; Ding, L.; Zhou, X.; Zhang, L. De novo synthetic short antimicrobial peptides against cariogenic bacteria. Arch. Oral Biol. 2017, 80, 41–50. [Google Scholar] [CrossRef]
- Chen, C.H.; Starr, C.G.; Troendle, E.P.; Wiedman, G.; Wimley, W.C.; Ulmschneider, J.P.; Ulmschneider, M.B. Simulation-Guided Rational de Novo Design of a Small Pore-Forming Antimicrobial Peptide. J. Am. Chem. Soc. 2019, 141, 4839–4848. [Google Scholar] [CrossRef] [PubMed]
- Vishnepolsky, B.; Zaalishvili, G.; Karapetian, M.; Nasrashvili, T.; Kuljanishvili, N.; Gabrielian, A.; Rosenthal, A.; Hurt, D.E.; Tartakovsky, M.; Grigolava, M.; et al. De Novo Design and In Vitro Testing of Antimicrobial Peptides against Gram-Negative Bacteria. Pharmaceuticals 2019, 12, 82. [Google Scholar] [CrossRef] [Green Version]
- CDC. Antibiotic Resistance Threats in the United States, 2019; U.S. Department of Health and Human Services, CDC: Atlanta, GA, USA, 2019.
- Haque, S.M.; Ashwaq, O.; Sarief, A.; Mohamed, A.K.A.J. A comprehensive review about SARS-CoV-2. Futur. Virol. 2020, 15, 625–648. [Google Scholar] [CrossRef] [PubMed]
- Yan, R.; Zhang, Y.; Li, Y.; Xia, L.; Guo, Y.; Zhou, Q. Structural basis for the recognition of SARS-CoV-2 by full-length human ACE2. Science 2020, 367, 1444–1448. [Google Scholar] [CrossRef] [Green Version]
- Linsky, T.W.; Vergara, R.; Codina, N.; Nelson, J.W.; Walker, M.J.; Su, W.; Barnes, C.O.; Hsiang, T.-Y.; Esser-Nobis, K.; Yu, K.; et al. De novo design of potent and resilient hACE2 decoys to neutralize SARS-CoV-2. Science 2020, 370, eabe0075. [Google Scholar] [CrossRef] [PubMed]
- LaRue, R.C.; Xing, E.; Kenney, A.D.; Zhang, Y.; Tuazon, J.A.; Li, J.; Yount, J.S.; Li, P.-K.; Sharma, A. Rationally Designed ACE2-Derived Peptides Inhibit SARS-CoV-2. Bioconjugate Chem. 2021, 32, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Goreshnik, I.; Coventry, B.; Case, J.B.; Miller, L.; Kozodoy, L.; Chen, R.E.; Carter, L.; Walls, A.C.; Park, Y.-J.; et al. De novo design of picomolar SARS-CoV-2 miniprotein inhibitors. Science 2020, 370. [Google Scholar] [CrossRef]
- Ibarra, A.A.; Bartlett, G.J.; Hegedüs, Z.; Dutt, S.; Hobor, F.; Horner, K.A.; Hetherington, K.; Spence, K.; Nelson, A.; Edwards, T.A.; et al. Predicting and Experimentally Validating Hot-Spot Residues at Protein–Protein Interfaces. ACS Chem. Biol. 2019, 14, 2252–2263. [Google Scholar] [CrossRef]
- Kelly, G.L.; Strasser, A. Toward Targeting Antiapoptotic MCL-1 for Cancer Therapy. Annu. Rev. Cancer Biol. 2020, 4, 299–313. [Google Scholar] [CrossRef] [Green Version]
- Johannes, J.W.; Bates, S.; Beigie, C.; Belmonte, M.A.; Breen, J.; Cao, S.; Centrella, P.A.; Clark, M.A.; Cuozzo, J.W.; Dumelin, C.E.; et al. Structure Based Design of Non-Natural Peptidic Macrocyclic Mcl-1 Inhibitors. ACS Med. Chem. Lett. 2017, 8, 239–244. [Google Scholar] [CrossRef] [Green Version]
- Fletcher, J.M.; Horner, K.A.; Bartlett, G.J.; Rhys, G.G.; Wilson, A.J.; Woolfson, D.N. De novocoiled-coil peptides as scaffolds for disrupting protein–protein interactions. Chem. Sci. 2018, 9, 7656–7665. [Google Scholar] [CrossRef] [Green Version]
- Kamagata, K.; Mano, E.; Itoh, Y.; Wakamoto, T.; Kitahara, R.; Kanbayashi, S.; Takahashi, H.; Murata, A.; Kameda, T. Rational design using sequence information only produces a peptide that binds to the intrinsically disordered region of p53. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef]
- Liu, Q.; Zhou, J.; Gao, J.; Ma, W.; Wang, S.; Xing, L. Rational design of EGFR dimerization-disrupting peptides: A new strategy to combat drug resistance in targeted lung cancer therapy. Biochimie 2020, 176, 128–137. [Google Scholar] [CrossRef] [PubMed]
- Silva, D.-A.; Yu, S.; Ulge, U.Y.; Spangler, J.B.; Jude, K.M.; Labão-Almeida, C.; Ali, L.R.; Quijano-Rubio, A.; Ruterbusch, M.; Leung, I.; et al. De novo design of potent and selective mimics of IL-2 and IL-15. Nat. Cell Biol. 2019, 565, 186–191. [Google Scholar] [CrossRef]
- Grisoni, F.; Neuhaus, C.S.; Hishinuma, M.; Gabernet, G.; Hiss, J.A.; Kotera, M.; Schneider, G. De novo design of anticancer peptides by ensemble artificial neural networks. J. Mol. Model. 2019, 25, 112. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Schneider, J.P. De novo Design of Selective Membrane-Active Peptides by Enzymatic Control of Their Conformational Bias on the Cell Surface. Angew. Chem. Int. Ed. 2019, 58, 13706–13710. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Roose, B.W.; Zemerov, S.D.; Stringer, M.A.; Dmochowski, I.J. Detecting protein–protein interactions by Xe-129 NMR. Chem. Commun. 2020, 56, 11122–11125. [Google Scholar] [CrossRef] [PubMed]
- Yudenko, A.; Smolentseva, A.; Maslov, I.; Semenov, O.; Goncharov, I.M.; Nazarenko, V.V.; Maliar, N.L.; Borshchevskiy, V.; Gordeliy, V.; Remeeva, A.; et al. Rational Design of a Split Flavin-Based Fluorescent Reporter. ACS Synth. Biol. 2021, 10, 72–83. [Google Scholar] [CrossRef] [PubMed]
- Baker, D.A. What has de novo protein design taught us about protein folding and biophysics? Protein Sci. 2019, 28, 678–683. [Google Scholar] [CrossRef] [Green Version]
- Kuhlman, B.; Bradley, P. Advances in protein structure prediction and design. Nat. Rev. Mol. Cell Biol. 2019, 20, 681–697. [Google Scholar] [CrossRef]
- Basanta, B.; Bick, M.J.; Bera, A.K.; Norn, C.; Chow, C.M.; Carter, L.P.; Goreshnik, I.; DiMaio, F.; Baker, D. An enumerative algorithm for de novo design of proteins with diverse pocket structures. Proc. Natl. Acad. Sci. USA 2020, 117, 22135–22145. [Google Scholar] [CrossRef]
- Strokach, A.; Becerra, D.; Corbi-Verge, C.; Perez-Riba, A.; Kim, P.M. Fast and Flexible Protein Design Using Deep Graph Neural Networks. Cell Syst. 2020, 11, 402–411.e4. [Google Scholar] [CrossRef]
- Skalic, M.; Jiménez, J.; Sabbadin, D.; De Fabritiis, G. Shape-Based Generative Modeling for de Novo Drug Design. J. Chem. Inf. Model. 2019, 59, 1205–1214. [Google Scholar] [CrossRef] [PubMed]
- Lucas, J.E.; Kortemme, T. New computational protein design methods for de novo small molecule binding sites. PLoS Comput. Biol. 2020, 16, e1008178. [Google Scholar] [CrossRef] [PubMed]
- Sesterhenn, F.; Yang, C.; Bonet, J.; Cramer, J.T.; Wen, X.; Wang, Y.; Chiang, C.-I.; Abriata, L.A.; Kucharska, I.; Castoro, G.; et al. De novo protein design enables the precise induction of RSV-neutralizing antibodies. Science 2020, 368, eaay5051. [Google Scholar] [CrossRef]
- Liu, J.; Zhou, X.-G.; Zhang, Y.; Zhang, G.-J. CGLFold: A contact-assisted de novo protein structure prediction using global exploration and loop perturbation sampling algorithm. Bioinformatics 2019, 36, 2443–2450. [Google Scholar] [CrossRef]
- Leal, E.S.; Adler, N.S.; Fernández, G.A.; Gebhard, L.G.; Battini, L.; Aucar, M.G.; Videla, M.; Monge, M.E.; Ríos, A.H.D.L.; Dávila, J.A.A.; et al. De novo design approaches targeting an envelope protein pocket to identify small molecules against dengue virus. Eur. J. Med. Chem. 2019, 182, 111628. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Gupta, S.T.P.; Bhimsaria, D.; Reed, J.L.; A Rodríguez-Martínez, J.; Ansari, A.Z.; Raman, S. De novo design of programmable inducible promoters. Nucleic Acids Res. 2019, 47, 10452–10463. [Google Scholar] [CrossRef] [PubMed]
- Callaway, E. ‘It will change everything’: DeepMind’s AI makes gigantic leap in solving protein structures. Nat. Cell Biol. 2020, 588, 203–204. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferrando, J.; Solomon, L.A. Recent Progress Using De Novo Design to Study Protein Structure, Design and Binding Interactions. Life 2021, 11, 225. https://doi.org/10.3390/life11030225
Ferrando J, Solomon LA. Recent Progress Using De Novo Design to Study Protein Structure, Design and Binding Interactions. Life. 2021; 11(3):225. https://doi.org/10.3390/life11030225
Chicago/Turabian StyleFerrando, Juan, and Lee A. Solomon. 2021. "Recent Progress Using De Novo Design to Study Protein Structure, Design and Binding Interactions" Life 11, no. 3: 225. https://doi.org/10.3390/life11030225
APA StyleFerrando, J., & Solomon, L. A. (2021). Recent Progress Using De Novo Design to Study Protein Structure, Design and Binding Interactions. Life, 11(3), 225. https://doi.org/10.3390/life11030225