
A Novel, Easy Assay Method for Human Cysteine Sulfinic Acid Decarboxylase

 ,
,  ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
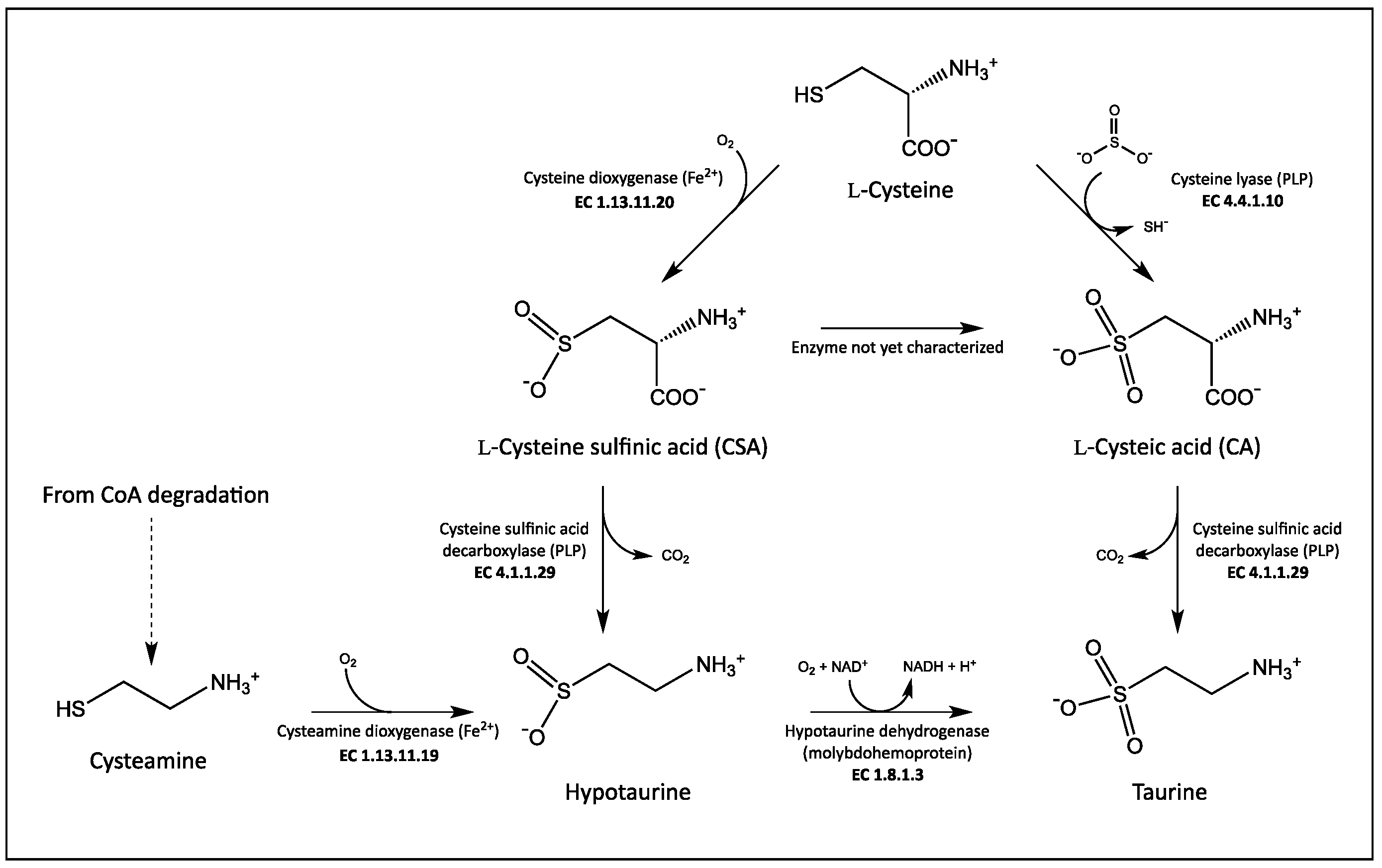
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Purification of CSAD
2.3. Spectroscopic Measurements
2.4. Thin Layer Chromatography
2.5. Activity Assay of CSAD with CSA and CA
2.6. Activity Assay of CSAD with Glutamate
2.7. Data Analysis
3. Results
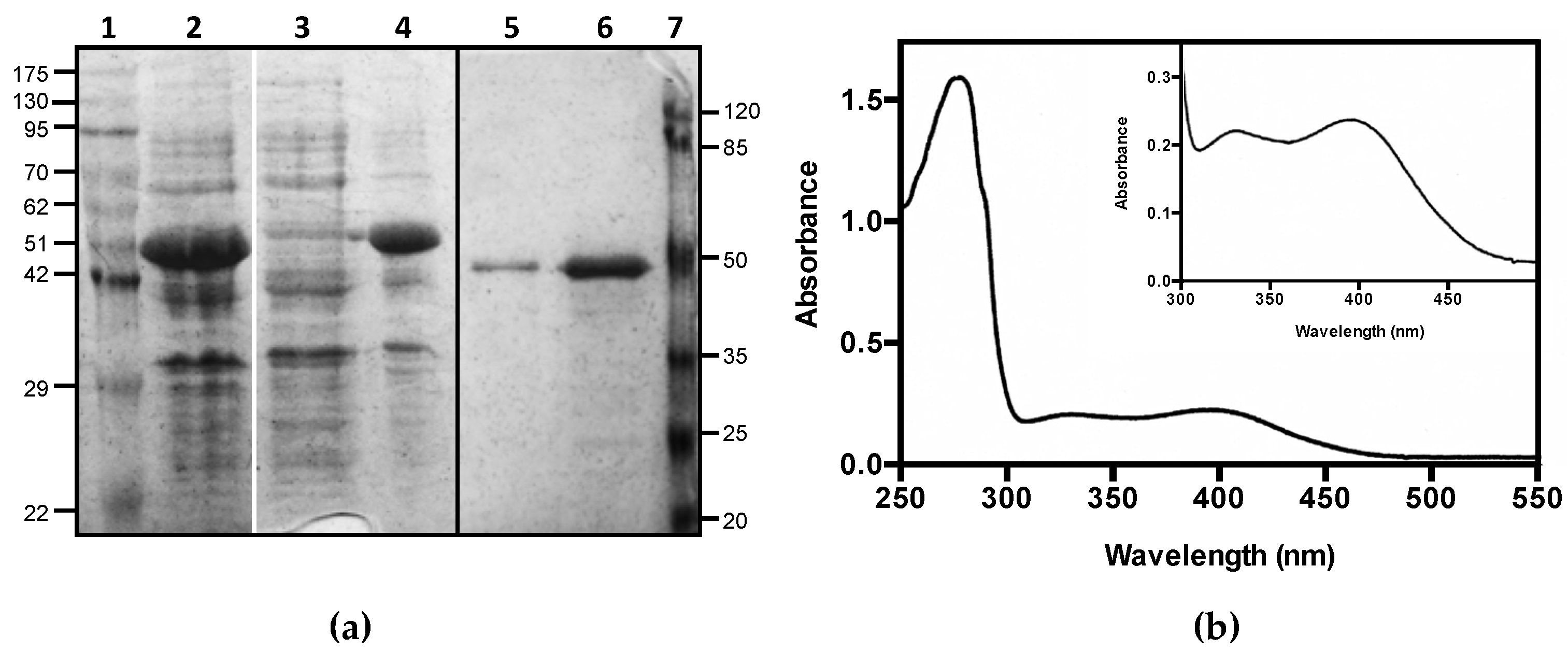
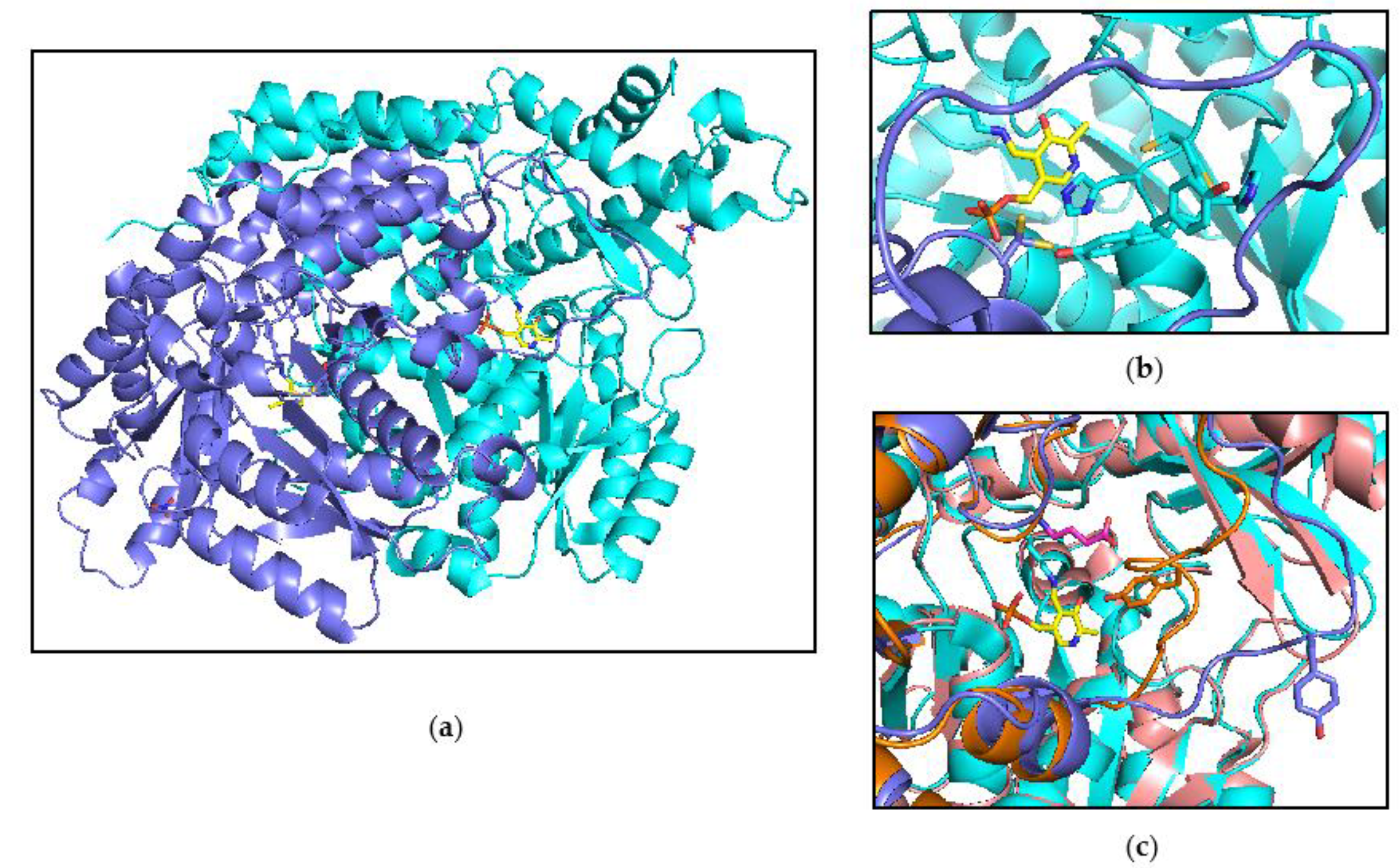
3.1. Expression, Purification, and Spectrophotometric Analysis of Human CSAD
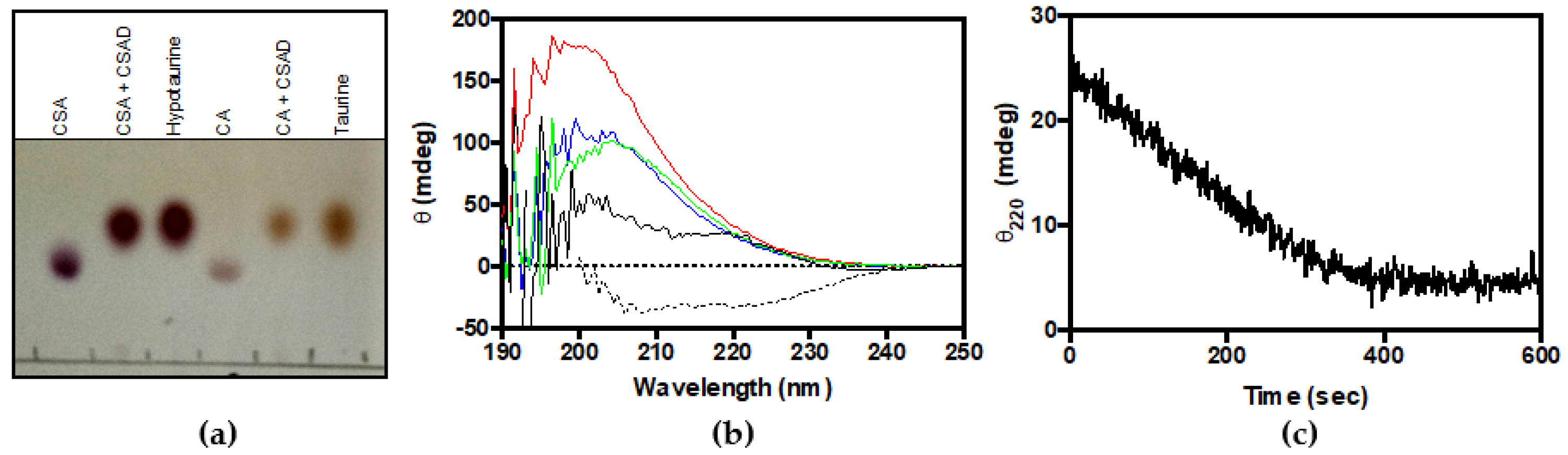
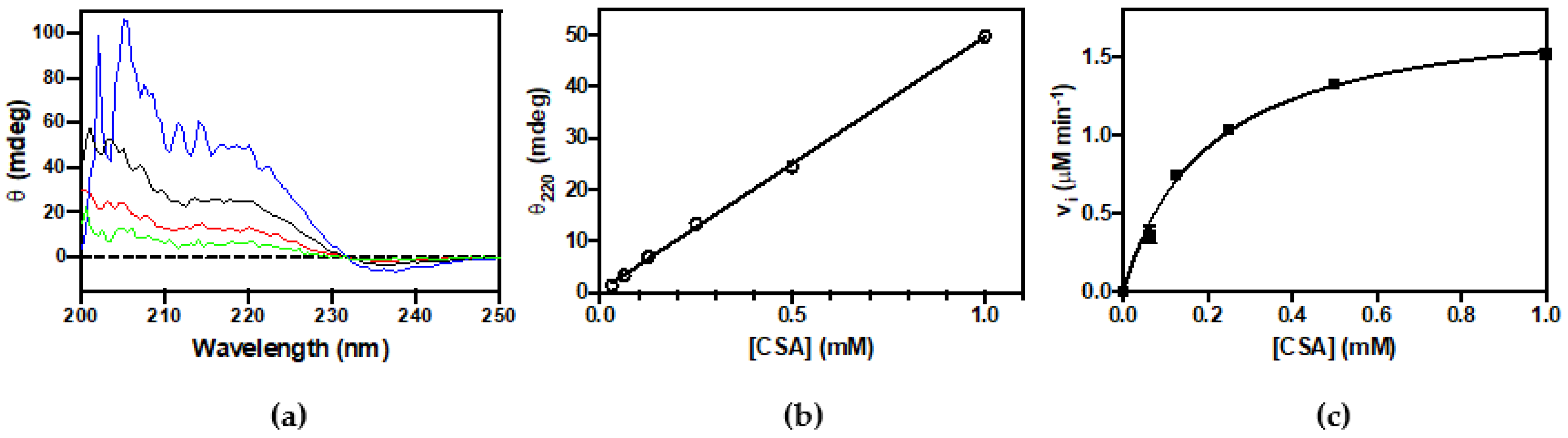
3.2. Activity Assay: Decarboxylation of CSA and CA
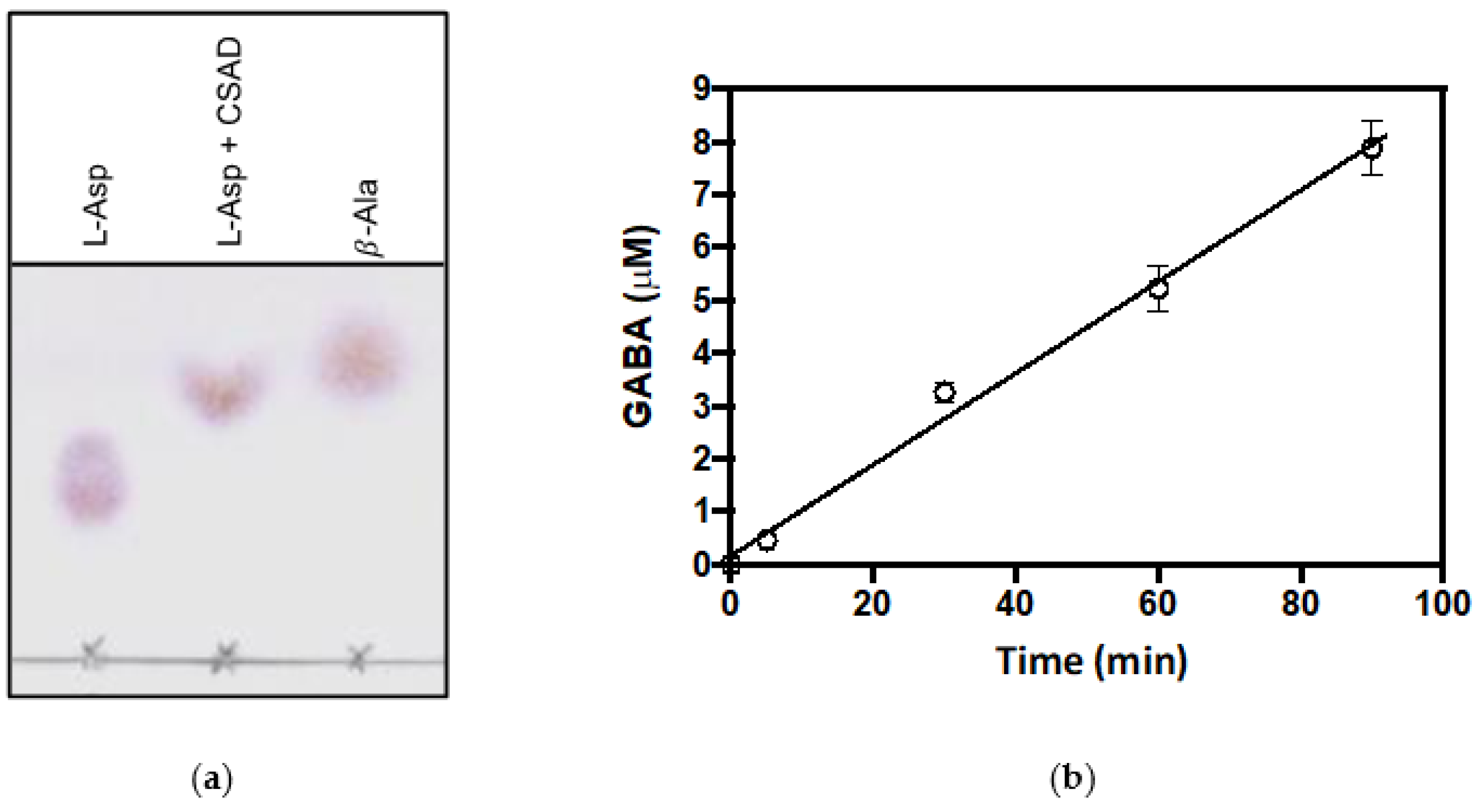
3.3. Decarboxylation of L-Aspartate and L-Glutamate
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brosnan, J.T.; Brosnan, M.E. The Sulfur-Containing Amino Acids: An Overview. J. Nutr. 2006, 136, 1636S–1640S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huxtable, R.J. Physiological actions of taurine. Physiol. Rev. 1992, 72, 101–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falany, C.N.; Johnson, M.R.; Barnes, S.; Diasio, R.B. Glycine and taurine conjugation of bile acids by a single enzyme. Molecular cloning and expression of human liver bile acid CoA:amino acid N-acyltransferase. J. Biol. Chem. 1994, 269, 19375–19379. [Google Scholar] [CrossRef]
- Marcinkiewicz, J.; Kontny, E. Taurine and inflammatory diseases. Amino Acids 2014, 46, 7–20. [Google Scholar] [CrossRef] [Green Version]
- Seidel, U.; Huebbe, P.; Rimbach, G. Taurine: A Regulator of Cellular Redox Homeostasis and Skeletal Muscle Function. Mol. Nutr. Food Res. 2019, 63, e1800569. [Google Scholar] [CrossRef]
- Lambert, I.H. Regulation of the cellular content of the organic osmolyte taurine in mammalian cells. Neurochem. Res. 2004, 29, 27–63. [Google Scholar] [CrossRef]
- Moran, J.; Salazar, P.; Pasantes-Morales, H. Effect of tocopherol and taurine on membrane fluidity of retinal rod outer segments. Exp. Eye Res. 1987, 45, 769–776. [Google Scholar] [CrossRef]
- Wu, J.-Y.; Prentice, H. Role of taurine in the central nervous system. J. Biomed. Sci. 2010, 17, S1. [Google Scholar] [CrossRef] [Green Version]
- Schaffer, S.W.; Shimada-Takaura, K.; Jong, C.J.; Ito, T.; Takahashi, K. Impaired energy metabolism of the taurinedeficient heart. Amino Acids 2016, 48, 549–558. [Google Scholar] [CrossRef] [PubMed]
- Hayes, K.C.; Carey, R.; Schmidt, S.Y. Retinal degeneration associated with taurine deficiency in the cat. Science 1975, 188, 949–951. [Google Scholar] [CrossRef]
- Verner, A.M.; McGuire, W.; Craig, J.S. Effect of taurine supplementation on growth and development in preterm or low birth weight infants. Cochrane Database Syst. Rev. 2007. [Google Scholar] [CrossRef] [PubMed]
- Murakami, S. Role of taurine in the pathogenesis of obesity. Mol. Nutr. Food Res. 2015, 59, 1353–1363. [Google Scholar] [CrossRef] [PubMed]
- Chesney, R.W. Taurine: Its biological role and clinical implications. Adv. Pediatr. 1985, 32, 1–42. [Google Scholar] [PubMed]
- Siefken, W.; Carstensen, S.; Springmann, G.; Wittern, K.-P.; Wenck, H.; Stäb, F.; Sauermann, G.; Schreiner, V.; Doering, T.; Janeke, G.; et al. Role of Taurine Accumulation in Keratinocyte Hydration. J. Investig. Dermatol. 2003, 121, 354–361. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.Y. Purification and characterization of cysteic acid and cysteine sulfinic acid decarboxylase and L-glutamate decarboxylase from bovine brain. Proc. Natl. Acad. Sci. USA 1982, 79, 4270–4274. [Google Scholar] [CrossRef] [Green Version]
- Jansonius, J.N. Structure, evolution and action of vitamin B6-dependent enzymes. Curr. Opin. Struct. Biol. 1998, 8, 759–769. [Google Scholar] [CrossRef]
- Tappaz, M.; Bitoun, M.; Reymond, I.; Sergeant, A. Characterization of the cDNA coding for rat brain cysteine sul-finate decarboxylase: Brain and liver enzymes are identical proteins encoded by two distinct mRNAs. J. Neurochem. 1999, 73, 903–912. [Google Scholar] [CrossRef]
- Tang, X.W.; Hsu, C.C.; Schloss, J.V.; Faiman, M.D.; Wu, E.; Yang, C.Y.; Wu, J.Y. Protein phosphorylation and taurine biosynthesis in vivo and in vitro. J. Neurosci. 1997, 17, 6947–6951. [Google Scholar] [CrossRef] [Green Version]
- Agnello, G.; Chang, L.L.; Lamb, C.M.; Georgiou, G.; Stone, E.M. Discovery of a Substrate Selectivity Motif in Amino Acid Decarboxylases Unveils a Taurine Biosynthesis Pathway in Prokaryotes. ACS Chem. Biol. 2013, 8, 2264–2271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tevatia, R.; Allen, J.; Rudrappa, D.; White, D.; Clemente, T.E.; Cerutti, H.; Demirel, Y.; Blum, P. The taurine biosynthetic pathway of microalgae. Algal Res. 2015, 9, 21–26. [Google Scholar] [CrossRef] [Green Version]
- Gill, S.C.; von Hippel, P.H. Calculation of protein extinction coefficients from amino acid sequence data. Anal. Biochem. 1989, 182, 319–326. [Google Scholar] [CrossRef]
- Tramonti, A.; De Biase, D.; Giartosio, A.; Bossa, F.; John, R.A. The Roles of His-167 and His-275 in the Reaction Catalyzed by Glutamate Decarboxylase from Escherichia coli. J. Biol. Chem. 1998, 273, 1939–1945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, C.M.; Johnson, R.J.; Metzler, D.E. Band-shape analysis and resolution of electronic spectra of pyridoxal phosphate and other 3-hydroxypyridine-4-aldehydes. Biochim. Biophys. Acta Gen. Subj. 1976, 421, 181–194. [Google Scholar] [CrossRef]
- Hayashi, H.; Mizuguchi, H.; Kagamiyama, H. Rat liver aromatic L-amino acid decarboxylase: Spectroscopic and kinetic analysis of the coenzyme and reaction intermediates. Biochemistry 1993, 32, 812–818. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, H.; Tsukiyama, F.; Ishii, S.; Mizuguchi, H.; Kagamiyama, H. Acid-base chemistry of the reaction of ar-omatic L-amino acid decarboxylase and dopa analyzed by transient and steady-state kinetics: Preferential binding of the substrate with its amino group unprotonated. Biochemistry 1999, 38, 15615–15622. [Google Scholar] [CrossRef]
- Voltattorni, C.B.; Minelli, A.; Vecchini, P.; Fiori, A.; Turano, C. Purification and characterization of 3,4-dihydroxyphenylalanine decarboxyase from pig kidney. Eur. J. Biochem. 1979, 93, 181–188. [Google Scholar] [CrossRef]
- Brooks, H.B.; Phillips, M.A. Circular Dichroism Assay for Decarboxylation of Optically Pure Amino Acids: Application to Ornithine Decarboxylase. Anal. Biochem. 1996, 238, 191–194. [Google Scholar] [CrossRef]
- Fogle, E.J.; Toney, M.D. Analysis of catalytic determinants of diaminopimelate and ornithine decarboxylases using alternate substrates. Biochim. Biophys. Acta Proteins Proteom. 2011, 1814, 1113–1119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winge, I.; Teigen, K.; Fossbakk, A.; Mahootchi, E.; Kleppe, R.; Sköldberg, F.; Kämpe, O.; Haavik, J. Mammalian CSAD and GADL1 have distinct biochemical properties and patterns of brain expression. Neurochem. Int. 2015, 90, 173–184. [Google Scholar] [CrossRef]
- Noda, M.; Matoba, Y.; Kumagai, T.; Sugiyama, M. A novel assay method for an amino acid racemase reaction based on circular dichroism. Biochem. J. 2005, 389, 491–496. [Google Scholar] [CrossRef] [Green Version]
- Blindermann, J.-M.; Maitre, M.; Ossola, L.; Mandel, P. Purification and Some Properties of l-Glutamate Decarboxylase from Human Brain. J. Biol. Inorg. Chem. 1978, 86, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Siaterli, M.Z.; Vassilacopoulou, D.; Fragoulis, E.G. Cloning and expression of human placental L-Dopa decar-boxylase. Neurochem. Res. 2003, 28, 797–803. [Google Scholar] [CrossRef] [PubMed]
- Oertel, W.H.; Schmechel, D.E.; Weise, V.K.; Ransom, D.H.; Tappaz, M.L.; Krutzsch, H.C.; Kopin, I.J. Comparison of cysteine sulphinic acid decarboxylase isoenzymes and glutamic acid decarboxylase in rat liver and brain. Neuroscience 1981, 6, 2701–2714. [Google Scholar] [CrossRef]
- Fenalti, G.; Law, R.H.P.; Buckle, A.M.; Langendorf, C.; Tuck, K.; Rosado, C.J.; Faux, N.G.; Mahmood, K.; Hampe, C.S.; Banga, J.P.; et al. GABA production by glutamic acid decarboxylase is regulated by a dynamic catalytic loop. Nat. Struct. Mol. Biol. 2007, 14, 280–286. [Google Scholar] [CrossRef] [PubMed]
- Bertoldi, M.; Gonsalvi, M.; Contestabile, R.; Voltattorni, C.B. Mutation of Tyrosine 332 to Phenylalanine Converts Dopa Decarboxylase into a Decarboxylation-dependent Oxidative Deaminase. J. Biol. Chem. 2002, 277, 36357–36362. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tramonti, A.; Contestabile, R.; Florio, R.; Nardella, C.; Barile, A.; Di Salvo, M.L. A Novel, Easy Assay Method for Human Cysteine Sulfinic Acid Decarboxylase. Life 2021, 11, 438. https://doi.org/10.3390/life11050438
Tramonti A, Contestabile R, Florio R, Nardella C, Barile A, Di Salvo ML. A Novel, Easy Assay Method for Human Cysteine Sulfinic Acid Decarboxylase. Life. 2021; 11(5):438. https://doi.org/10.3390/life11050438
Chicago/Turabian StyleTramonti, Angela, Roberto Contestabile, Rita Florio, Caterina Nardella, Anna Barile, and Martino L. Di Salvo. 2021. "A Novel, Easy Assay Method for Human Cysteine Sulfinic Acid Decarboxylase" Life 11, no. 5: 438. https://doi.org/10.3390/life11050438
APA StyleTramonti, A., Contestabile, R., Florio, R., Nardella, C., Barile, A., & Di Salvo, M. L. (2021). A Novel, Easy Assay Method for Human Cysteine Sulfinic Acid Decarboxylase. Life, 11(5), 438. https://doi.org/10.3390/life11050438