
Pro-Oxidative Effect of KIO3 and Protective Effect of Melatonin in the Thyroid—Comparison to Other Tissues



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Animals
2.3. Incubation of Tissue Homogenates
2.4. Measurement of Lipid Peroxidation Products
2.5. Statistical Analyses
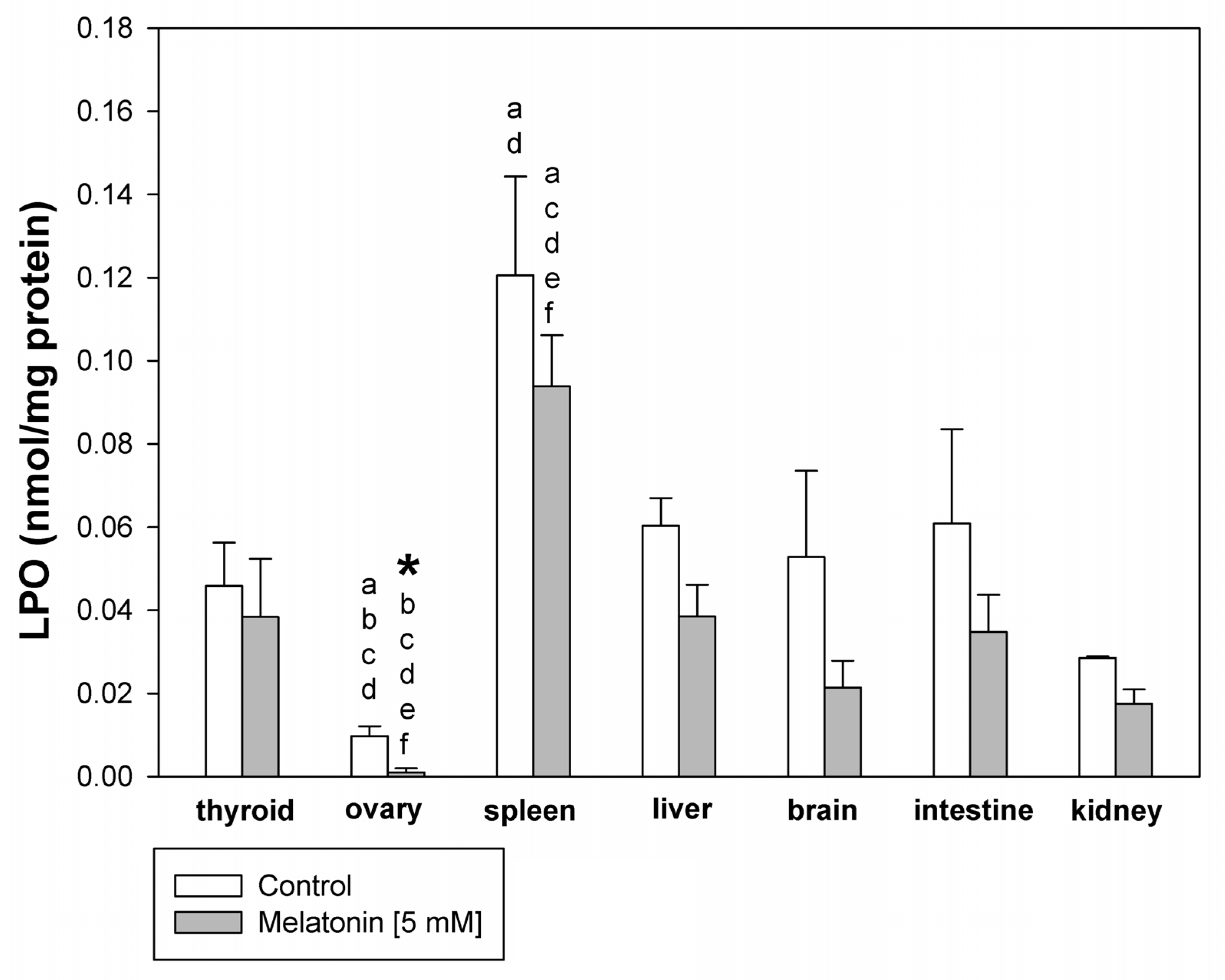
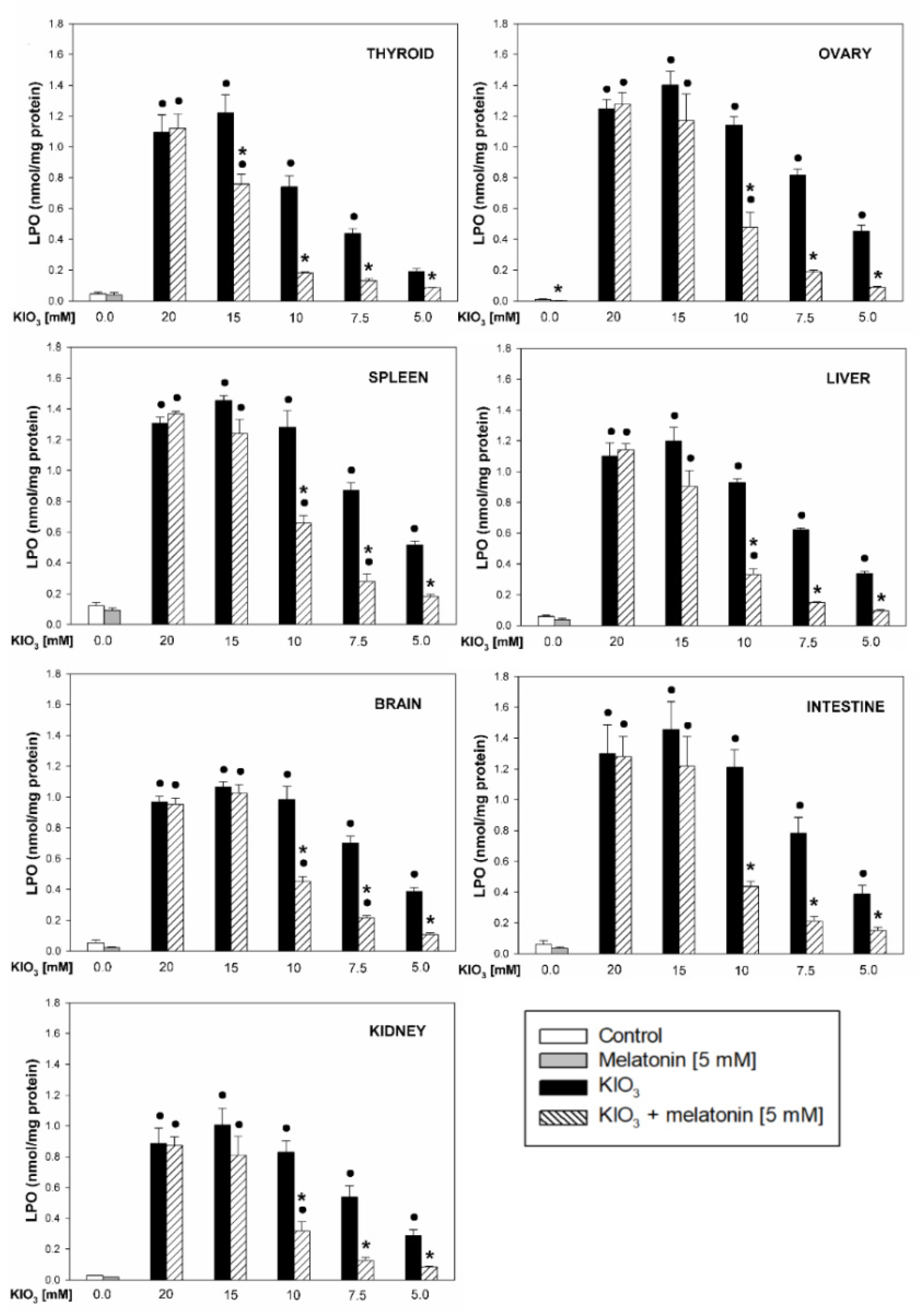
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sies, H.; Jones, D.P. Reactive oxygen species (ROS) as pleiotropic physiological signalling agents. Nat. Rev. Mol. Cell Biol. 2020, 21, 363–383. [Google Scholar] [CrossRef]
- Hunyadi, A. The mechanism(s) of action of antioxidants: From scavenging reactive oxygen/nitrogen species to redox signaling and the generation of bioactive secondary metabolites. Med. Res. Rev. 2019, 39, 2505–2533. [Google Scholar] [CrossRef] [Green Version]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef] [Green Version]
- Su, L.-J.; Zhang, J.-H.; Gomez, H.; Murugan, R.; Hong, X.; Xu, D.; Jiang, F.; Peng, Z.-Y. Reactive oxygen species-induced lipid peroxidation in apoptosis, autophagy, and ferroptosis. Oxid Med. Cell Longev. 2019, 2019, 5080843. [Google Scholar] [CrossRef] [Green Version]
- Pisoschi, A.M.; Pop, A. The role of antioxidants in the chemistry of oxidative stress: A review. Eur. J. Med. Chem. 2015, 97, 55–74. [Google Scholar] [CrossRef]
- Ursini, F.; Maiorino, M. Lipid peroxidation and ferroptosis: The role of GSH and GPx4. Free Radic. Biol. Med. 2020, 152, 175–185. [Google Scholar] [CrossRef]
- Karbownik-Lewinska, M.; Kokoszko-Bilska, A. Oxidative damage to macromolecules in the thyroid—Experimental evidence. Thyroid Res. 2012, 5, 25. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, D.P.; Dupuy, C. Thyroid hormone biosynthesis and release. Mol. Cell Endocrinol. 2017, 458, 6–15. [Google Scholar] [CrossRef] [PubMed]
- Vitale, M.; Di Matola, T.; D’Ascoli, F.; Salzano, S.; Bogazzi, F.; Fenzi, G.; Martino, E.; Rossi, G. Iodide excess induces apoptosis in thyroid cells through a p53-independent mechanism involving oxidative stress. Endocrinology 2000, 141, 598–605. [Google Scholar] [CrossRef]
- Portulano, C.; Paroder-Belenitsky, M.; Carrasco, N. The Na+/I- symporter (NIS): Mechanism and medical impact. Endocr. Rev. 2014, 35, 106–149. [Google Scholar] [CrossRef] [Green Version]
- Filetti, S.; Bidart, J.M.; Arturi, F.; Caillou, B.; Russo, D.; Schlumberger, M. Sodium/iodide symporter: A key transport system in thyroid cancer cell metabolism. Eur. J. Endocrinol. 1999, 141, 443–457. [Google Scholar] [CrossRef] [Green Version]
- Dohán, O.; De la Vieja, A.; Paroder, V.; Riedel, C.; Artani, M.; Reed, M.; Ginter, C.S.; Carrasco, N. The sodium/iodide Symporter (NIS): Characterization, regulation, and medical significance. Endocr. Rev. 2003, 24, 48–77. [Google Scholar] [CrossRef] [Green Version]
- Southern, A.P.; Jwayyed, S. Iodine Toxicity. StatPearls. Available online: https://www.statpearls.com/ArticleLibrary/viewarticle/40905 (accessed on 11 April 2021).
- Shahid, M.A.; Ashraf, M.A.; Sharma, S. Physiology, Thyroid Hormone. StatPearls. Available online: https://www.statpearls.com/ArticleLibrary/viewarticle/30145 (accessed on 18 May 2020).
- Zimmermann, M.B.; Boelaert, K. Iodine deficiency and thyroid disorders. Lancet Diabetes Endocrinol. 2015, 3, 286–295. [Google Scholar] [CrossRef]
- Blankenship, J.L.; Garrett, G.S.; Khan, N.A.; De-Regil, L.M.; Spohrer, R.; Gorstein, J. Effect of iodized salt on organoleptic properties of processed foods: A systematic review. J. Food Sci. Technol. 2018, 55, 3341–3352. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Ma, W.; Liu, L.; Xu, J.; Wang, H.; Li, X.; Wang, J.; Hang, J.; Wang, Z.; Gu, Y. Analysis of potassium iodate reduction in tissue homogenates using high performance liquid chromatography-inductively coupled plasma-mass spectrometry. J. Trace Elem. Med. Biol. 2015, 32, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Milczarek, M.; Stepniak, J.; Lewinski, A.; Karbownik-Lewinska, M. Potassium iodide, but not potassium iodate, as a potential protective agent against oxidative damage to membrane lipids in porcine thyroid. Thyroid Res. 2013, 6, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karbownik-Lewinska, M.; Stepniak, J.; Milczarek, M.; Lewinski, A. Protective effect of KI in mtDNA in porcine thyroid: Comparison with KIO3 and nDNA. Eur. J. Nutr. 2015, 54, 319–323. [Google Scholar] [CrossRef] [Green Version]
- Iwan, P.; Stepniak, J.; Karbownik-Lewinska, M. Melatonin reduces high levels of lipid peroxidation induced by potassium iodate in porcine thyroid. Int. J. Vitam. Nutr. Res. 2019, 17, 1–7. [Google Scholar] [CrossRef]
- Iwan, P.; Karbownik-Lewinska, M. Indole-3-propionic acid reduces lipid peroxidation induced by potassium iodate in porcine thyroid. Interdiscip. Toxicol. 2020, 13, 101–105. [Google Scholar]
- Iwan, P.; Stepniak, J.; Karbownik-Lewinska, M. Cumulative protective effect of melatonin and indole-3-propionic acid against KIO3-induced oxidative damage to membrane lipids in porcine thyroid homogenates. Toxics 2021, 9, 89. [Google Scholar] [CrossRef]
- Tiran, B.; Karpf, E.; Tiran, A.; Lax, S.; Langsteger, W.; Eber, O.; Lorenz, O. Iodine content of thyroid tissue in the Styrian population. Acta Med. Austriaca 1993, 20, 6–8. [Google Scholar]
- Taurog, A.; Chaikoff, I.L.; Feller, D.D. The mechanism of iodine concentration by the thyroid gland: Its non-organic iodine-binding capacity in the normal and propylthiouracil-treated rat. J. Biol. Chem. 1947, 171, 189–201. [Google Scholar] [CrossRef]
- Taurog, A.; Tong, W.; Chaikoff, I.L. Non-thyroglobulin iodine of the thyroid gland II. Inorganic iodide. J. Biol. Chem. 1951, 191, 677–682. [Google Scholar] [CrossRef]
- Hays, M.T. Estimation of total body iodine content in normal young men. Thyroid 2001, 11, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Franke, K.; Schöne, F.; Berk, A.; Leiterer, M.; Flachowsky, G. Influence of dietary iodine on the iodine content of pork and the distribution of the trace element in the body. Eur. J. Nutr. 2008, 47, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Mair, C.; Schedle, K.; Hammerl, S.; Schodl, K.; Windisch, W. Effect of iodine source and dose on growth and iodine content in tissue and plasma thyroid hormones in fattening pigs. Eur. J. Nutr. 2012, 51, 685–691. [Google Scholar] [CrossRef] [PubMed]
- Bürgi, H.; Schaffner, T.H.; Seiler, J.P. The toxicology of iodate: A review of the literature. Thyroid 2001, 11, 449–456. [Google Scholar] [CrossRef] [Green Version]
- Reiter, R.J.; Mayo, J.C.; Tan, D.X.; Sainz, R.M.; Alatorre-Jimenez, M.; Qin, L. Melatonin as an antioxidant: Under promises but over delivers. J. Pineal Res. 2016, 61, 253–278. [Google Scholar] [CrossRef]
- Manchester, L.C.; Coto-Montes, A.; Boga, J.A.; Andersen, L.P.H.; Zhou, Z.; Galano, A.; Vriend, J.; Tan, D.X.; Reiter, R.J. Melatonin: An ancient molecule that makes oxygen metabolically tolerable. J. Pineal Res. 2015, 59, 403–419. [Google Scholar] [CrossRef]
- Baş, E.; Nazıroğlu, M. Treatment with melatonin and selenium attenuates docetaxel-induced apoptosis and oxidative injury in kidney and testes of mice. Andrologia 2019, 51, e13320. [Google Scholar] [CrossRef]
- Sutradhar, S.; Deb, A.; Singh, S.S. Melatonin attenuates diabetes-induces oxidative stress in spleen and suppression of splenocyte proliferation in laboratory mice. Arch. Physiol. Biochem. 2020, 5, 1–12. [Google Scholar] [CrossRef]
- Behram-Kandemir, Y.; Aydin, C.; Gorgisen, G. The effects of melatonin on oxidative stress and prevention of primordial follicle loss via activation of mTOR pathway in the rat ovary. Cell Mol. Biol. 2017, 63, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Karbownik, M.; Reiter, R.J.; Garcia, J.J.; Tan, D.X.; Qi, W.; Manchester, L.C. Melatonin reduces rat hepatic macromolecular damage due to oxidative stress caused by delta-aminolevulinic acid. Biochim. Biophys. Acta. 2000, 1523, 140–146. [Google Scholar] [CrossRef]
- Morabito, R.; Remigante, A.; Marino, A. Melatonin protects band 3 protein in human erythrocytes against H2O2-induced oxidative stress. Molecules 2019, 24, 2741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hang, H.-M.; Zhang, Y. Melatonin: A well documented antioxidant with conditional pro-oxidant actions. J. Pineal Res. 2014, 57, 131–146. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Kuzmuk, K.N.; Schook, L.B. Pigs as a model for biomedical sciences. In The Genetics of the Pig, 2nd ed.; Rothschild, M.F., Ruvinsky, A., Eds.; CAB International: Wallingford, UK, 2011; pp. 426–444. [Google Scholar]
- Bürgi, H. Iodine excess. Best Pract Res. Clin. Endocrinol. Metab. 2010, 24, 107–115. [Google Scholar] [CrossRef]
- Darrouzet, E.; Lindenthal, S.; Marcellin, D.; Pellequer, J.-L.; Pourcher, T. The sodium/iodide symporter: State of the art of its molecular characterization. Biochim. Biophys. Acta 2014, 1838, 244–253. [Google Scholar] [CrossRef] [Green Version]
- Bizhanova, A.; Kopp, P. The sodium-iodide symporter NIS and pendrin in iodide homeostasis of the thyroid. Endocrinology 2009, 150, 1084–1090. [Google Scholar] [CrossRef] [Green Version]
- Ravera, S.; Reyna-Neyra, A.; Ferrandino, G.; Amzel, L.M.; Carrasco, N. The sodium/iodide symporter (NIS): Molecular physiology and preclinical and clinical applications. Annu. Rev. Physiol. 2017, 79, 261–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rynkowska, A.; Stepniak, J.; Karbownik-Lewinska, M. Fenton reaction-induced oxidative damage to membrane lipids and protective effects of 17β-estradiol in porcine ovary and thyroid homogenates. Int. J. Environ. Res. Public Health. 2020, 17, 6841. [Google Scholar] [CrossRef]
- Karbownik-Lewinska, M.; Stepniak, J.; Krawczyk, J.; Zasada, K.; Szosland, J.; Gesing, A.; Lewinski, A. External hydrogen peroxide is not indispensable for experimental induction of lipid peroxidation via Fenton reaction in porcine ovary homogenates. Neuro Endocrinol. Lett. 2010, 31, 343–347. [Google Scholar]
- Stepniak, J.; Lewinski, A.; Karbownik-Lewinska, M. Membrane lipids and nuclear DNA are differently susceptive to Fenton reaction substrates in porcine thyroid. Toxicol. In Vitro 2013, 27, 71–78. [Google Scholar] [CrossRef]
- Kurokawa, Y.; Maekawa, A.; Takahashi, M.; Hayashi, Y. Toxicity and carcinogenicity of potassium bromate—A new renal carcinogen. Environ. Health Perspect. 1990, 87, 309–335. [Google Scholar] [CrossRef] [PubMed]
- IARC Monographs—Potassium Bromate. Available online: https://monographs.iarc.who.int/wp-content/uploads/2018/06/mono73-22.pdf (accessed on 14 March 2021).
- Osha Select Carcinogens. Available online: https://memphis.edu/ehs/pdfs/carlist.pdf (accessed on 21 June 2021).
- Vriend, J. The pineal and melatonin in the regulation of pituitary thyroid axis. Life Sci. 1981, 29, 1929–1936. [Google Scholar] [CrossRef]
- Lewiński, A.; Karbownik, M. REVIEW. Melatonin and the thyroid gland. Neuro Endocrinol. Lett. 2002, 23 (Suppl. 1), 73–78. [Google Scholar]
- Galano, A.; Tan, D.X.; Reiter, R.J. On the free radical scavenging activities of melatonin’s metabolites, AFMK and AMK. J. Pineal Res. 2013, 54, 245–257. [Google Scholar] [CrossRef]
- Reiter, R.J.; Tan, D.X.; Galano, A. Melatonin: Exceeding expectations. Physiology (Bethesda) 2014, 29, 325–333. [Google Scholar] [CrossRef] [Green Version]
- Tordjman, S.; Chokron, S.; Delorme, R.; Charrier, A.; Bellissant, E.; Jaafari, N.; Fougerou, C. Melatonin: Pharmacology, functions and therapeutic benefits. Curr. Neuropharmacol. 2017, 15, 434–443. [Google Scholar] [CrossRef] [PubMed]
- Andersen, L.P.H.; Gögenur, I.; Rosenberg, J.; Reiter, R.J. The safety of melatonin in humans. Clin. Drug Investig. 2015, 36, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Waldhauser, F.; Weiszenbacher, G.; Frisch, H.; Zeitlhuber, U.; Waldhauser, M.; Wurtman, R.J. Fall in nocturnal serum of melatonin during puberty and pubescence. Lancet. 1984, 1, 362–365. [Google Scholar] [CrossRef]
- Zetner, D.; Andersen, L.P.K.; Alder, R.; Jessen, M.L.; Tolstrup, A.; Rosenberg, J. Pharmacokinetics and safety of intravenous, intravesical, rectal, transdermal, and vaginal melatonin in healthy female volunteers: A cross-over study. Pharmacology 2021, 106, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Vural, E.M.S.; van Munster, B.C.; de Rooij, S.E. Optimal dosages for melatonin supplementation therapy in older adults: A systematic review of current literature. Drugs Aging 2014, 31, 441–451. [Google Scholar] [CrossRef] [Green Version]
- Sato, K.; Meng, F.; Francis, H.; Wu, N.; Chen, L.; Kennedy, L.; Zhou, T.; Franchitto, A.; Onori, P.; Gaudio, E.; et al. Melatonin and circadian rhythms in liver diseases: Functional roles and potential therapies. J. Pineal Res. 2020, 68, e12639. [Google Scholar] [CrossRef] [PubMed]
- Salim, S. Oxidative stress and the Central Nervous System. J. Pharmacol. Exp. Ther. 2017, 360, 201–205. [Google Scholar] [CrossRef]
- Behrman, H.R.; Kodaman, P.H.; Preston, S.L.; Gao, S. Oxidative stress and the ovary. J. Soc. Gynecol. Investig. 2001, 8 (Suppl. 1), S40–S42. [Google Scholar] [CrossRef]
- Diaz de Barboza, G.; Guizzardi, S.; Moine, L.; Tolosa de Talamoni, N. Oxidative stress, antioxidants and intestinal calcium absorption. World J. Gastroenterol. 2017, 23, 2841–2853. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iwan, P.; Stepniak, J.; Karbownik-Lewinska, M. Pro-Oxidative Effect of KIO3 and Protective Effect of Melatonin in the Thyroid—Comparison to Other Tissues. Life 2021, 11, 592. https://doi.org/10.3390/life11060592
Iwan P, Stepniak J, Karbownik-Lewinska M. Pro-Oxidative Effect of KIO3 and Protective Effect of Melatonin in the Thyroid—Comparison to Other Tissues. Life. 2021; 11(6):592. https://doi.org/10.3390/life11060592
Chicago/Turabian StyleIwan, Paulina, Jan Stepniak, and Malgorzata Karbownik-Lewinska. 2021. "Pro-Oxidative Effect of KIO3 and Protective Effect of Melatonin in the Thyroid—Comparison to Other Tissues" Life 11, no. 6: 592. https://doi.org/10.3390/life11060592
APA StyleIwan, P., Stepniak, J., & Karbownik-Lewinska, M. (2021). Pro-Oxidative Effect of KIO3 and Protective Effect of Melatonin in the Thyroid—Comparison to Other Tissues. Life, 11(6), 592. https://doi.org/10.3390/life11060592