
Stress-Related Regulation Is Abnormal in the Psoriatic Uninvolved Skin

 , , ,
, , ,  and
and 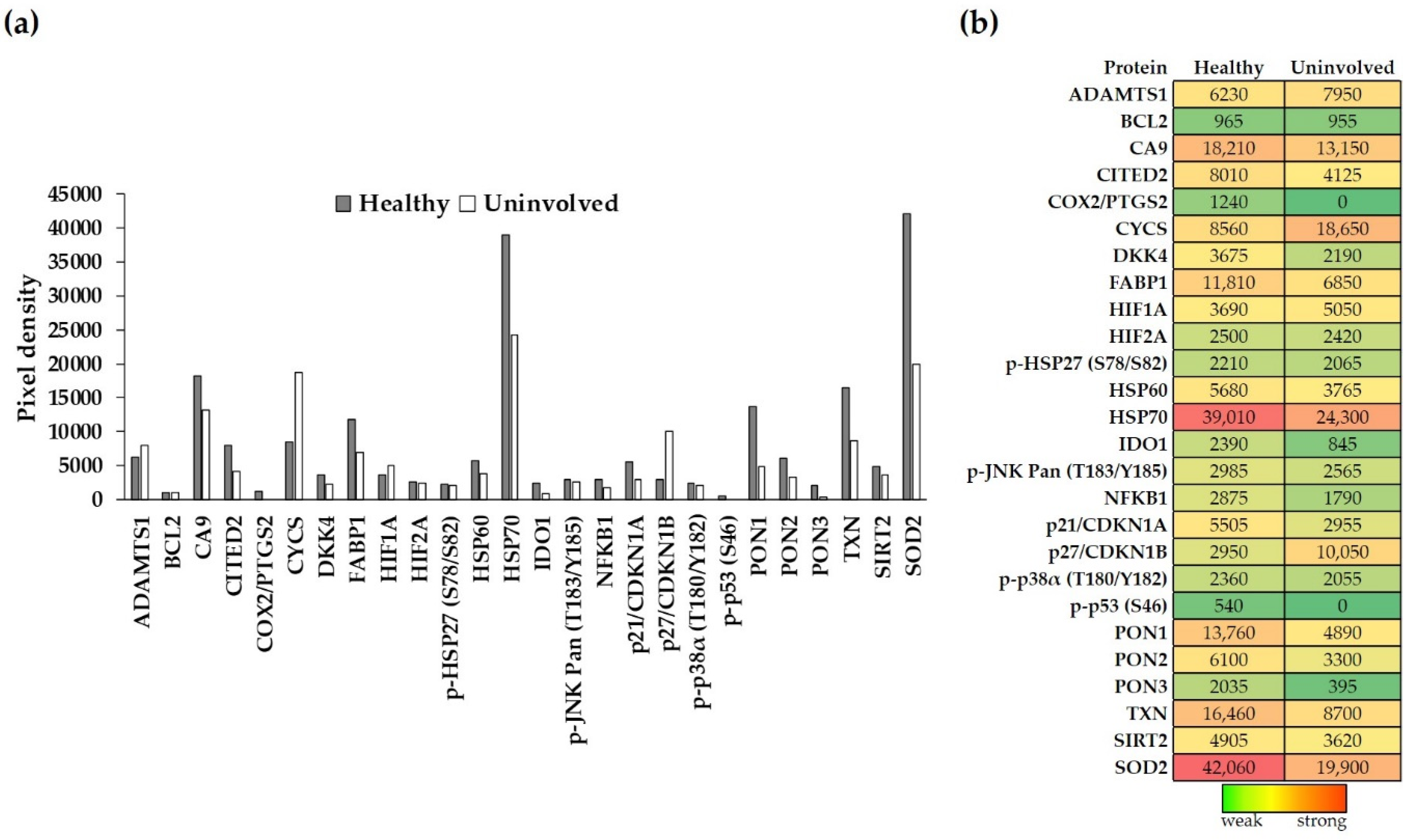{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Skin Samples and Ethics
2.2. Protein Isolation from Skin Biopsies
2.3. Cell-Stress Protein Array
2.4. Data Analysis
2.5. Immunofluorescence Staining
2.6. Examination of p27/CDKN1B and FOXO1 Localization in Normal Human Epidermal Keratinocytes (Nheks)
3. Results
3.1. Annotated Cell-Stress-Related Proteins (CSRPs) Are Expressed in Healthy Aand Psoriatic Uninvolved Skin
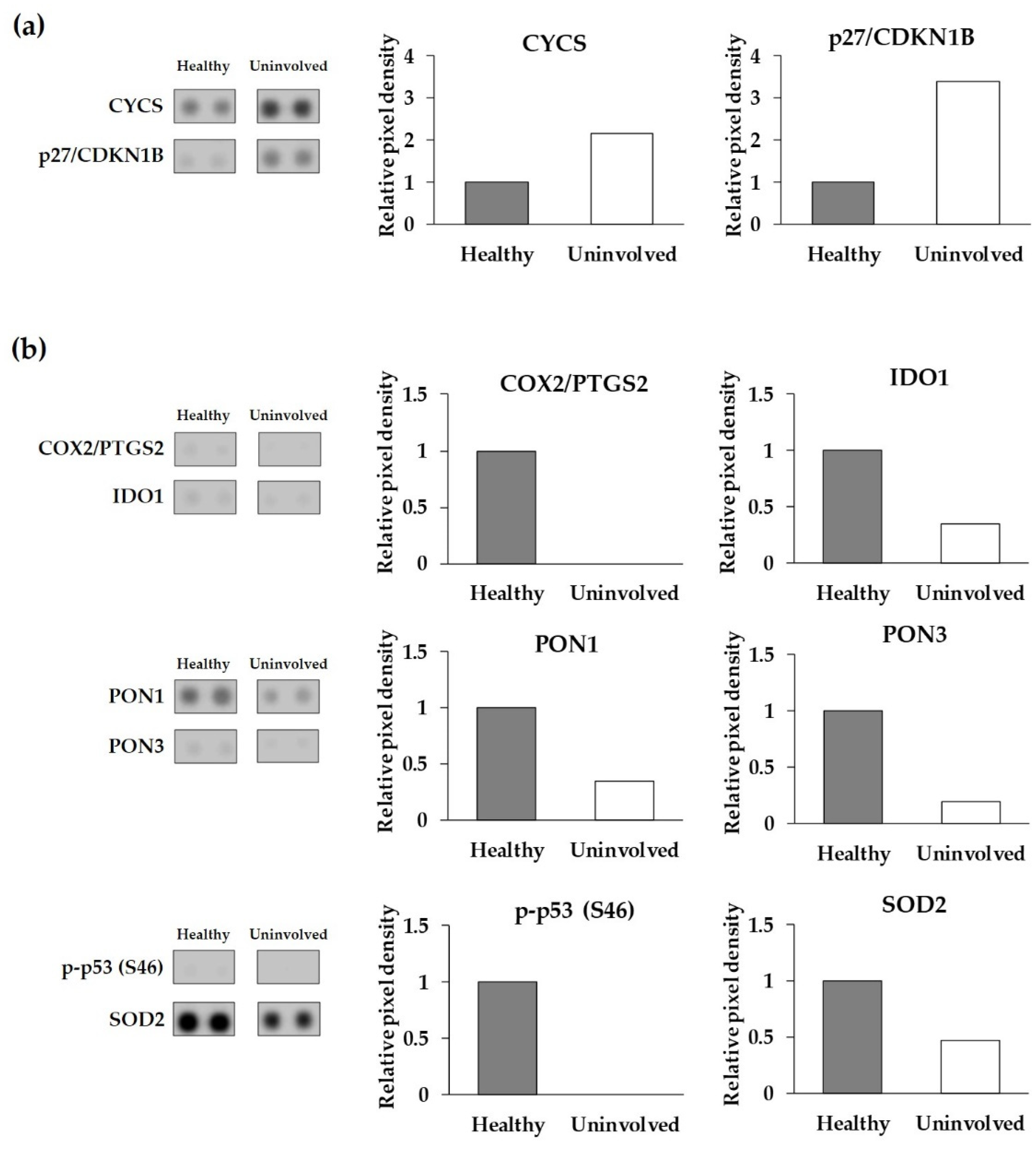
3.2. The Expression of Cell-Stress-Related Proteins in Psoriatic Uninvolved Skin Is Altered in Comparison with Healthy Skin
3.3. The FOXO-Mediated Transcription Is Over-Represented by the Differentially Expressed Cell-Stress-Related Proteins in the Psoriatic Uninvolved Tissue
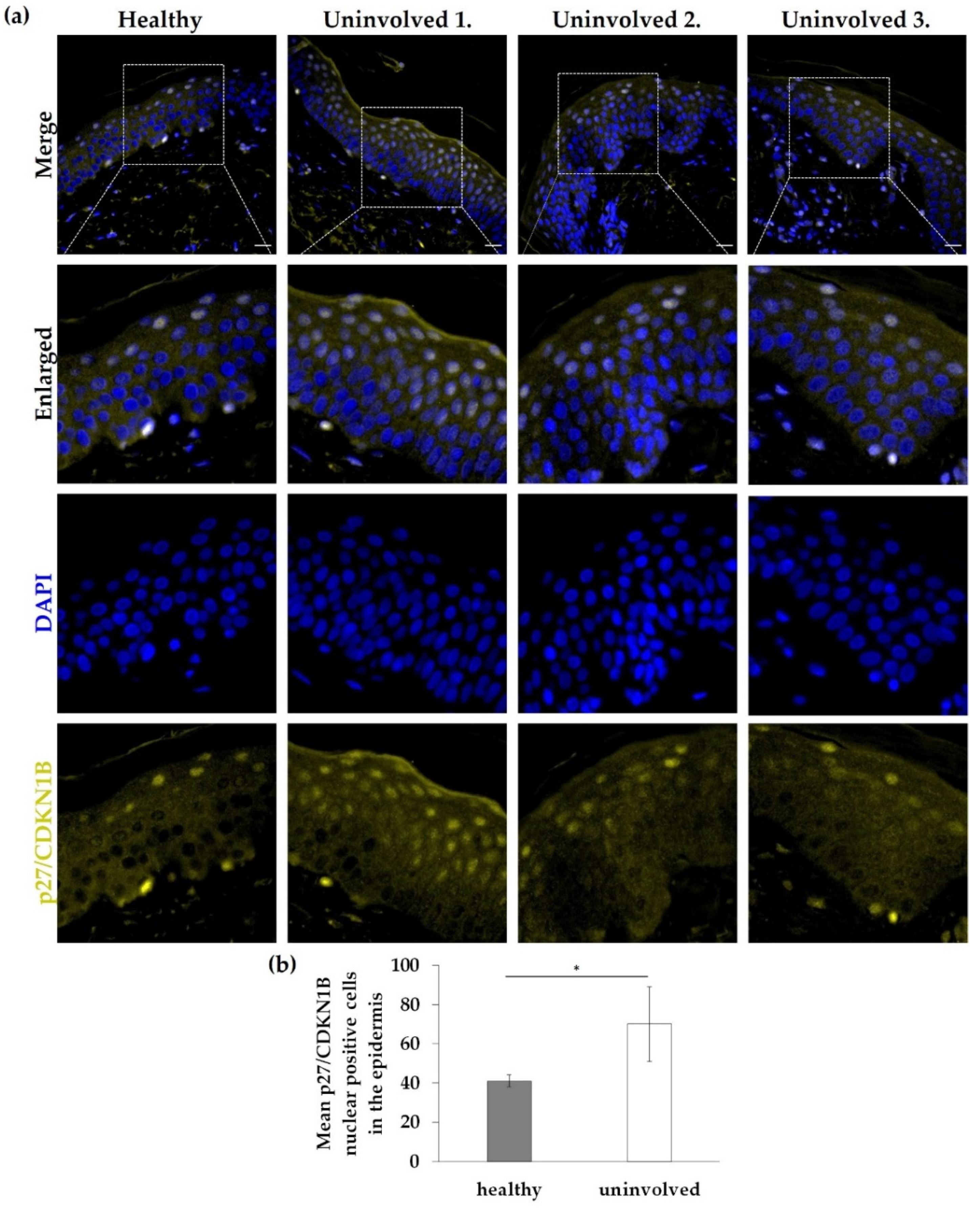
3.4. Nuclear Expression of p27/CDKN1B in the Psoriatic Uninvolved Epidermis Is Increased
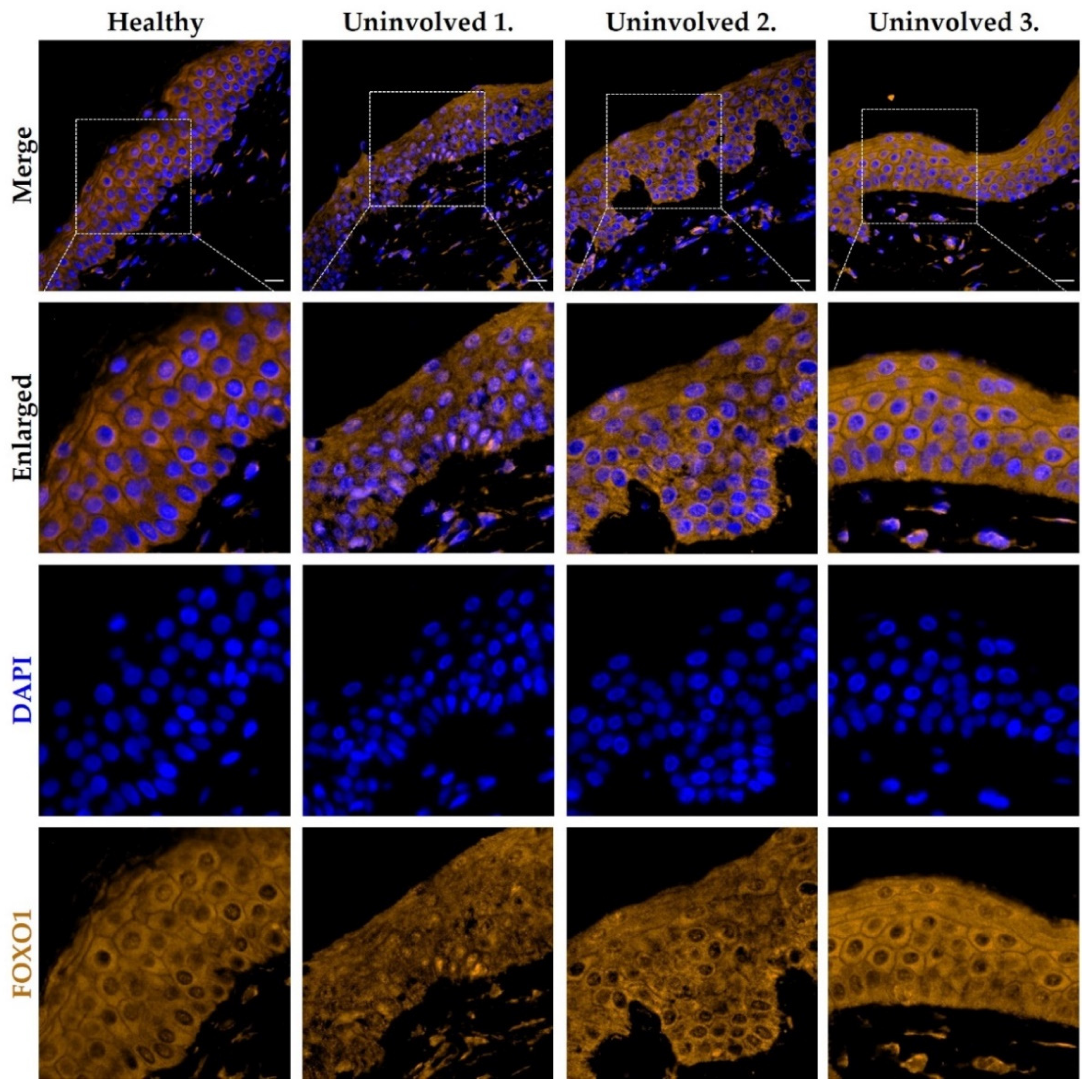
3.5. The FOXO1 Expression Pattern in the Psoriatic Uninvolved Skin Is Altered Compared to Normal Skin
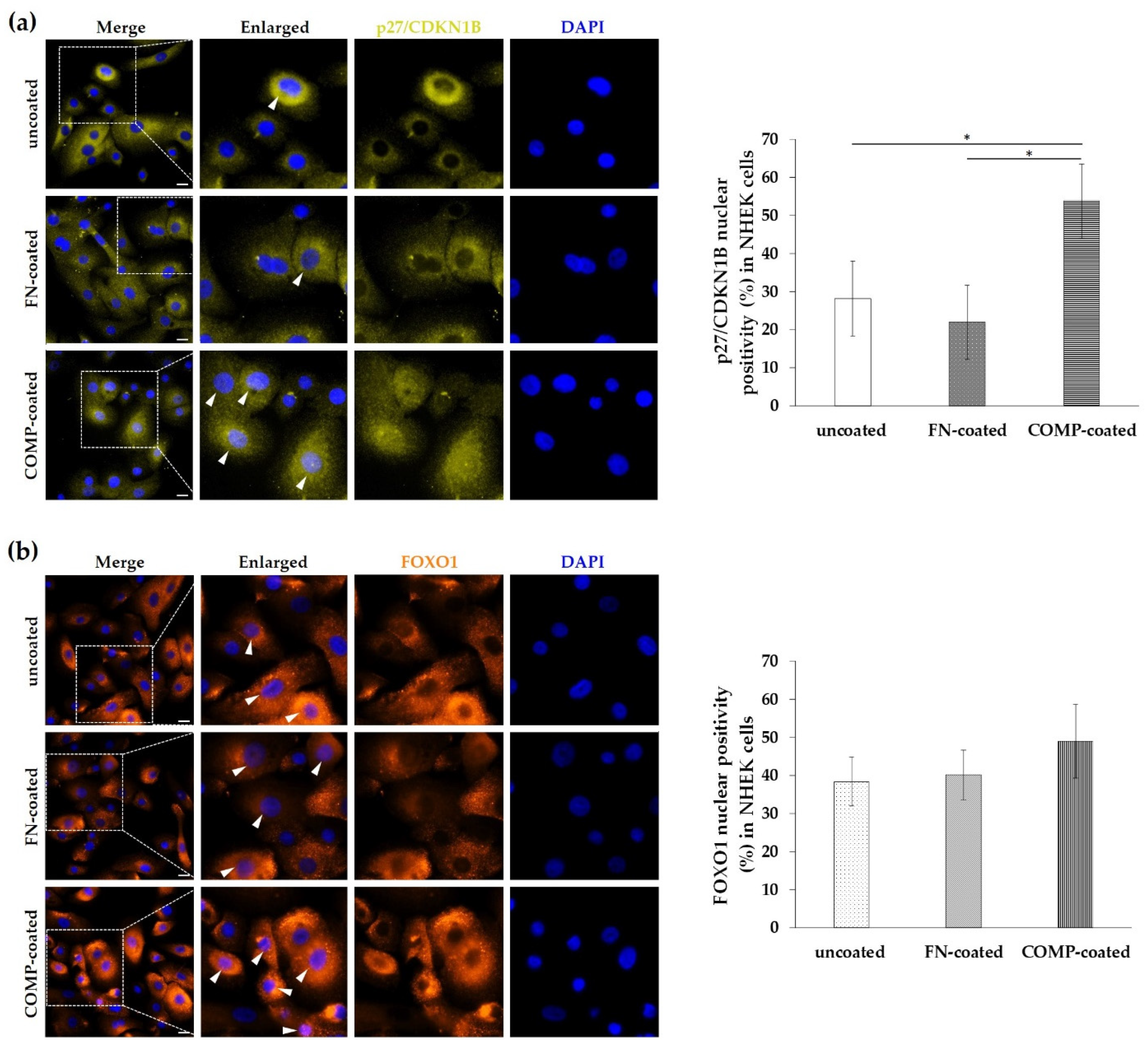
3.6. Nuclear Localization of P27/CDKN1B and FOXO1 Was Elevated in Normal Human Epidermal Keratinocytes (NHEKs) Cultured On COMP-Coated Surfaces
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Amoruso, G.F.; Nisticò, S.P.; Iannone, L.; Russo, E.; Rago, G.; Patruno, C.; Bennardo, L. Ixekizumab may improve renal function in psoriasis. Healthcare 2021, 9, 543. [Google Scholar] [CrossRef]
- Dattola, A.; Silvestri, M.; Bennardo, L.; Passante, M.; Rizzuto, F.; Dastoli, S.; Patruno, C.; Bianchi, L.; Nisticò, S.P. A novel vehicle for the treatment of psoriasis. Dermatol. Ther. 2020, 33, e13185. [Google Scholar] [CrossRef] [Green Version]
- Griffiths, C.E.M.; Armstrong, A.W.; Gudjonsson, J.E.; Barker, J.N.W.N. Psoriasis. Lancet Lond. Engl. 2021, 397, 1301–1315. [Google Scholar] [CrossRef]
- Bozó, R.; Flink, L.B.; Belső, N.; Gubán, B.; Széll, M.; Kemény, L.; Bata-Csörgő, Z. Could basement membrane alterations, resembling micro-wounds at the dermo-epidermal junction in psoriatic non-lesional skin, make the skin susceptible to lesion formation? Exp. Dermatol. 2020. [Google Scholar] [CrossRef]
- Mondello, M.R.; Magaudda, L.; Pergolizzi, S.; Santoro, A.; Vaccaro, M.; Califano, L.; Cannavò, S.P.; Guarneri, B. Behaviour of laminin 1 and type IV collagen in uninvolved psoriatic skin. Immunohistochemical study using confocal laser scanning microscopy. Arch. Dermatol. Res. 1996, 288, 527–531. [Google Scholar] [CrossRef] [PubMed]
- Vaccaro, M.; Magaudda, L.; Cutroneo, G.; Trimarchi, F.; Barbuzza, O.; Guarneri, F.; Guarneri, B. Changes in the distribution of laminin alpha1 chain in psoriatic skin: Immunohistochemical study using confocal laser scanning microscopy. Br. J. Dermatol. 2002, 146, 392–398. [Google Scholar] [CrossRef]
- Fleischmajer, R.; Kuroda, K.; Hazan, R.; Gordon, R.E.; Lebwohl, M.G.; Sapadin, A.N.; Unda, F.; Iehara, N.; Yamada, Y. Basement membrane alterations in psoriasis are accompanied by epidermal overexpression of MMP-2 and its inhibitor TIMP-2. J. Investig. Dermatol. 2000, 115, 771–777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pellegrini, G.; De Luca, M.; Orecchia, G.; Balzac, F.; Cremona, O.; Savoia, P.; Cancedda, R.; Marchisio, P.C. Expression, topography, and function of integrin receptors are severely altered in keratinocytes from involved and uninvolved psoriatic skin. J. Clin. Investig. 1992, 89, 1783–1795. [Google Scholar] [CrossRef] [Green Version]
- Ting, K.M.; Rothaupt, D.; McCormick, T.S.; Hammerberg, C.; Chen, G.; Gilliam, A.C.; Stevens, S.; Culp, L.; Cooper, K.D. Overexpression of the Oncofetal Fn Variant Containing the EDA Splice-in Segment in the Dermal–Epidermal Junction of Psoriatic uninvolved skin. J. Investig. Dermatol. 2000, 114, 706–711. [Google Scholar] [CrossRef]
- Széll, M.; Bata-Csörgo, Z.; Koreck, A.; Pivarcsi, A.; Polyánka, H.; Szeg, C.; Gaál, M.; Dobozy, A.; Kemény, L. Proliferating keratinocytes are putative sources of the psoriasis susceptibility-related EDA+ (extra domain A of fibronectin) oncofetal fibronectin. J. Investig. Dermatol. 2004, 123, 537–546. [Google Scholar] [CrossRef]
- Gubán, B.; Vas, K.; Balog, Z.; Manczinger, M.; Bebes, A.; Groma, G.; Széll, M.; Kemény, L.; Bata-Csörgő, Z. Abnormal regulation of fibronectin production by fibroblasts in psoriasis. Br. J. Dermatol. 2016, 174, 533–541. [Google Scholar] [CrossRef] [Green Version]
- Bozó, R.; Szél, E.; Danis, J.; Gubán, B.; Bata-Csörgő, Z.; Szabó, K.; Kemény, L.; Groma, G. Cartilage oligomeric matrix protein negatively influences keratinocyte proliferation via α5β1-Integrin: Potential relevance of altered cartilage oligomeric matrix protein expression in psoriasis. J. Investig. Dermatol. 2020, 140, 1733–1742.e7. [Google Scholar] [CrossRef]
- Bata-Csorgo, Z.; Hammerberg, C.; Voorhees, J.J.; Cooper, K.D. Kinetics and regulation of human keratinocyte stem cell growth in short-term primary ex vivo culture. Cooperative growth factors from psoriatic lesional T lymphocytes stimulate proliferation among psoriatic uninvolved, but not normal, stem keratinocytes. J. Clin. Investig. 1995, 95, 317–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, G.; McCormick, T.S.; Hammerberg, C.; Ryder-Diggs, S.; Stevens, S.R.; Cooper, K.D. Basal keratinocytes from uninvolved psoriatic skin exhibit accelerated spreading and focal adhesion kinase responsiveness to fibronectin. J. Investig. Dermatol. 2001, 117, 1538–1545. [Google Scholar] [CrossRef] [Green Version]
- Szabó, K.; Bata-Csörgő, Z.; Dallos, A.; Bebes, A.; Francziszti, L.; Dobozy, A.; Kemény, L.; Széll, M. Regulatory networks contributing to psoriasis susceptibility. Acta Derm. Venereol. 2014, 94, 380–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiss, G.; Shemer, A.; Trau, H. The Koebner phenomenon: Review of the literature. J. Eur. Acad. Dermatol. Venereol. 2002, 16, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Qiao, P.; Guo, W.; Ke, Y.; Fang, H.; Zhuang, Y.; Jiang, M.; Zhang, J.; Shen, S.; Qiao, H.; Dang, E.; et al. Mechanical stretch exacerbates psoriasis by stimulating keratinocyte proliferation and cytokine production. J. Investig. Dermatol. 2019, 139, 1470–1479. [Google Scholar] [CrossRef] [Green Version]
- Hatta, N.; Takata, M.; Kawara, S.; Hirone, T.; Takehara, K. Tape stripping induces marked epidermal proliferation and altered TGF-alpha expression in non-lesional psoriatic skin. J. Dermatol. Sci. 1997, 14, 154–161. [Google Scholar] [CrossRef]
- Sonkoly, E.; Bata-Csorgo, Z.; Pivarcsi, A.; Polyanka, H.; Kenderessy-Szabo, A.; Molnar, G.; Szentpali, K.; Bari, L.; Megyeri, K.; Mandi, Y.; et al. Identification and characterization of a novel, psoriasis susceptibility-related noncoding RNA gene, PRINS. J. Biol. Chem. 2005, 280, 24159–24167. [Google Scholar] [CrossRef] [Green Version]
- Széll, M.; Danis, J.; Bata-Csörgő, Z.; Kemény, L. PRINS, a primate-specific long non-coding RNA, plays a role in the keratinocyte stress response and psoriasis pathogenesis. Pflugers Arch. 2016, 468, 935–943. [Google Scholar] [CrossRef] [Green Version]
- Danis, J.; Göblös, A.; Bata-Csörgő, Z.; Kemény, L.; Széll, M. PRINS non-coding RNA regulates nucleic acid-induced innate immune responses of human keratinocytes. Front. Immunol. 2017, 8, 1053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szabad, G.; Kormos, B.; Pivarcsi, A.; Széll, M.; Kis, K.; Kenderessy Szabó, A.; Dobozy, A.; Kemény, L.; Bata-Csörgo, Z. Human adult epidermal melanocytes cultured without chemical mitogens express the EGF receptor and respond to EGF. Arch. Dermatol. Res. 2007, 299, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Burgering, B.M.T. A brief introduction to FOXOlogy. Oncogene 2008, 27, 2258–2262. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, M.A.; Zhu, Q.; Hammond, J.M. FoxO1a can alter cell cycle progression by regulating the nuclear localization of p27kip in granulosa cells. Mol. Endocrinol. 2004, 18, 1756–1767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Luo, W.; Chen, S. Comparison of gene expression profiles reveals aberrant expression of FOXO1, Aurora A/B and EZH2 in lesional psoriatic skins. Mol. Biol. Rep. 2011, 38, 4219–4224. [Google Scholar] [CrossRef]
- Therianou, A.; Vasiadi, M.; Delivanis, D.A.; Petrakopoulou, T.; Katsarou-Katsari, A.; Antoniou, C.; Stratigos, A.; Tsilioni, I.; Katsambas, A.; Rigopoulos, D.; et al. Mitochondrial dysfunction in affected skin and increased mitochondrial DNA in serum from patients with psoriasis. Exp. Dermatol. 2019, 28, 72–75. [Google Scholar] [CrossRef] [PubMed]
- Kunz, M.; Simon, J.C.; Saalbach, A. Psoriasis: Obesity and fatty acids. Front. Immunol. 2019, 10, 1807. [Google Scholar] [CrossRef]
- Tadini, G.; Cerri, A.; Crosti, L.; Cattoretti, G.; Berti, E. P53 and oncogenes expression in psoriasis. Acta Derm. Venereol. Suppl. (Stockh.) 1989, 146, 33–35. [Google Scholar]
- Nakayama, K.I.; Hatakeyama, S.; Nakayama, K. Regulation of the cell cycle at the G1-S transition by proteolysis of cyclin E and p27Kip1. Biochem. Biophys. Res. Commun. 2001, 282, 853–860. [Google Scholar] [CrossRef]
- Granata, M.; Skarmoutsou, E.; Gangemi, P.; Mazzarino, M.C.; D’Amico, F. S100A7, Jab1, and p27kip1 expression in psoriasis and S100A7 CRISPR-activated human keratinocyte cell line. J. Cell. Biochem. 2019, 120, 3384–3392. [Google Scholar] [CrossRef]
- Henri, P.; Prevel, C.; Pellerano, M.; Lacotte, J.; Stoebner, P.E.; Morris, M.C.; Meunier, L. Psoriatic epidermis is associated with upregulation of CDK2 and inhibition of CDK4 activity. Br. J. Dermatol. 2020, 182, 678–689. [Google Scholar] [CrossRef] [PubMed]
- Kadunce, D.P.; Krueger, G.G. Pathogenesis of psoriasis. Dermatol. Clin. 1995, 13, 723–737. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bozó, R.; Danis, J.; Flink, L.B.; Vidács, D.L.; Kemény, L.; Bata-Csörgő, Z. Stress-Related Regulation Is Abnormal in the Psoriatic Uninvolved Skin. Life 2021, 11, 599. https://doi.org/10.3390/life11070599
Bozó R, Danis J, Flink LB, Vidács DL, Kemény L, Bata-Csörgő Z. Stress-Related Regulation Is Abnormal in the Psoriatic Uninvolved Skin. Life. 2021; 11(7):599. https://doi.org/10.3390/life11070599
Chicago/Turabian StyleBozó, Renáta, Judit Danis, Lili Borbála Flink, Dániel László Vidács, Lajos Kemény, and Zsuzsanna Bata-Csörgő. 2021. "Stress-Related Regulation Is Abnormal in the Psoriatic Uninvolved Skin" Life 11, no. 7: 599. https://doi.org/10.3390/life11070599
APA StyleBozó, R., Danis, J., Flink, L. B., Vidács, D. L., Kemény, L., & Bata-Csörgő, Z. (2021). Stress-Related Regulation Is Abnormal in the Psoriatic Uninvolved Skin. Life, 11(7), 599. https://doi.org/10.3390/life11070599