
Entamoeba histolytica Trophozoites Interact with the c-Met Receptor at the Surface of Liver Origin Cells through the Gal/GalNAc Amoebic Lectin

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Cocultures of Entamoeba histolytica: HepG2
2.3. Immunoprecipitation
2.4. Western Blot
2.5. Hematoxylin and Eosin Staining
2.6. Immunofluorescence Assay
3. Results
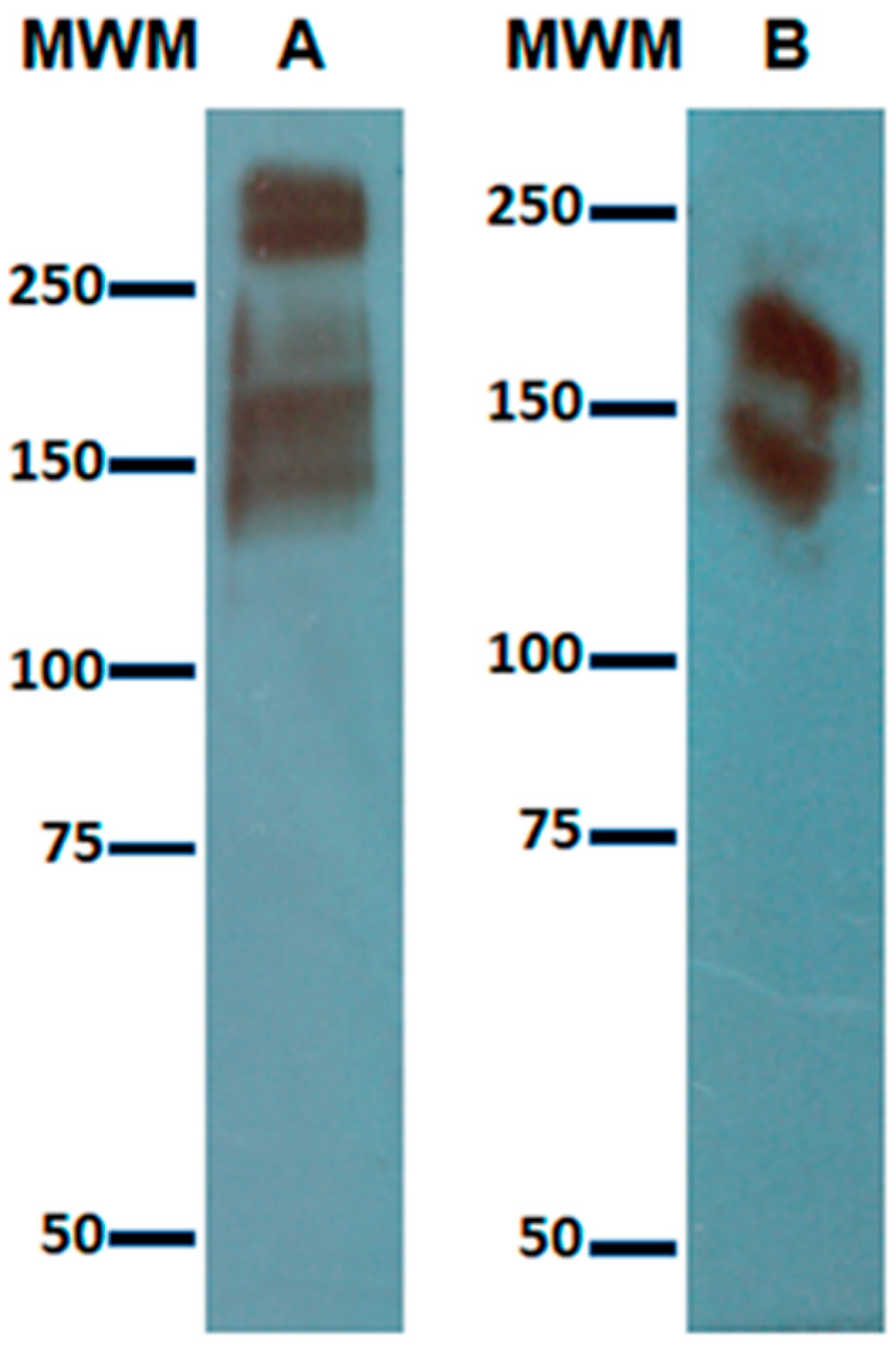
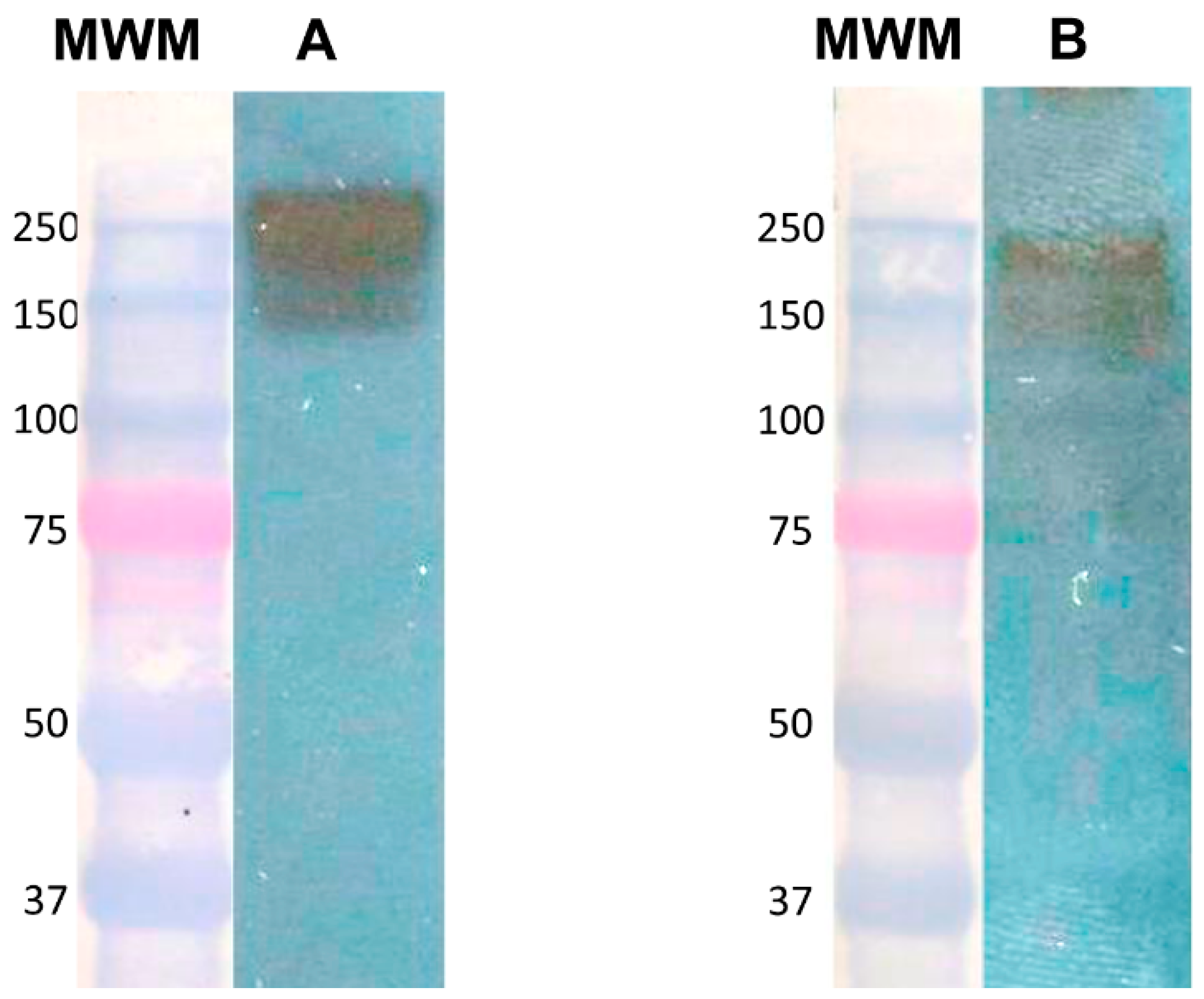
3.1. Expression of Gal Lectin in Amoebic Trophozoites and c-Met in HepG2 Cells
3.2. Identification of the Gal Lectin by Anti-HGF Antibody
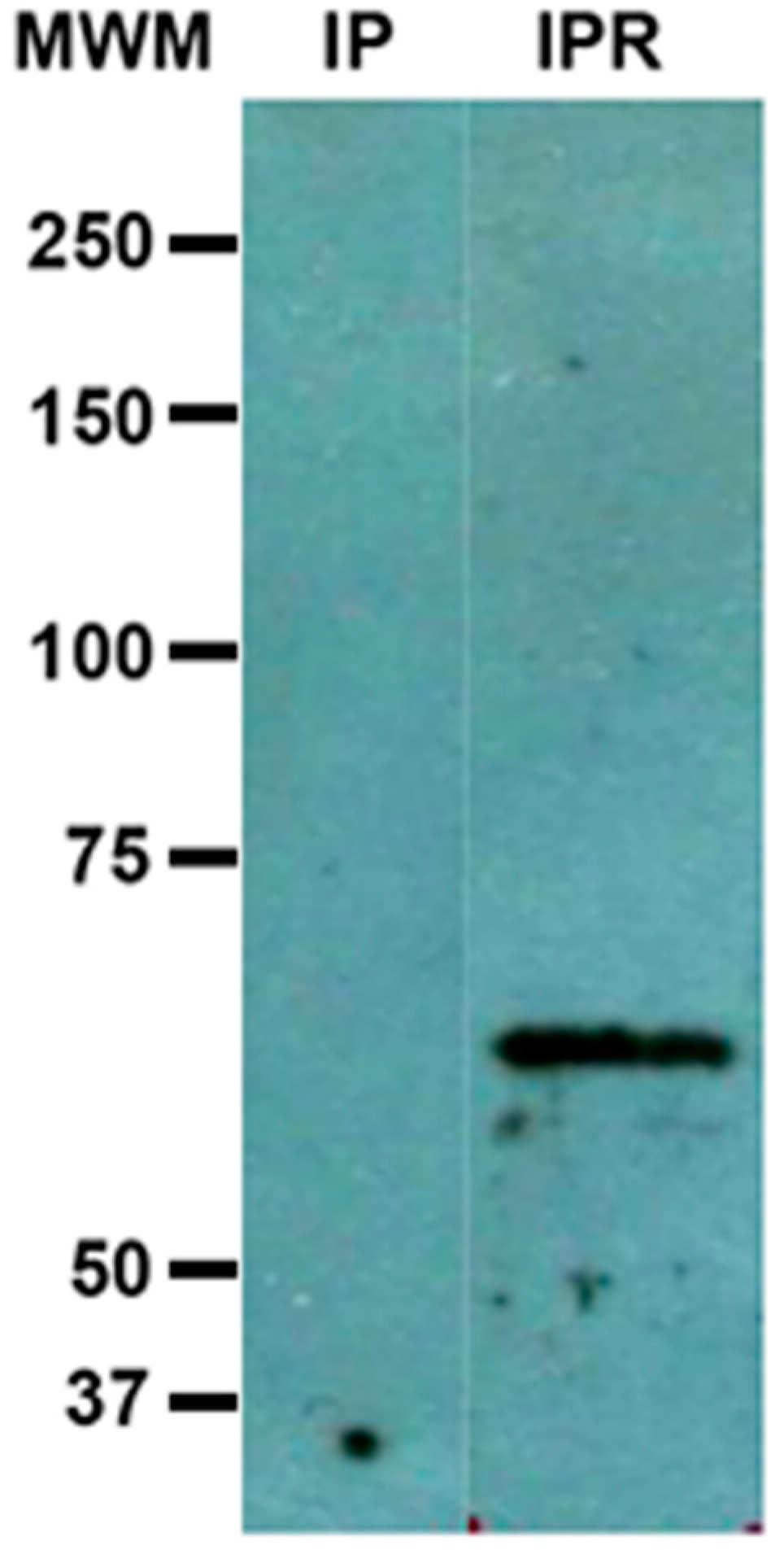
3.3. Immunoprecipitation: Interaction of c-Met and Gal Lectin
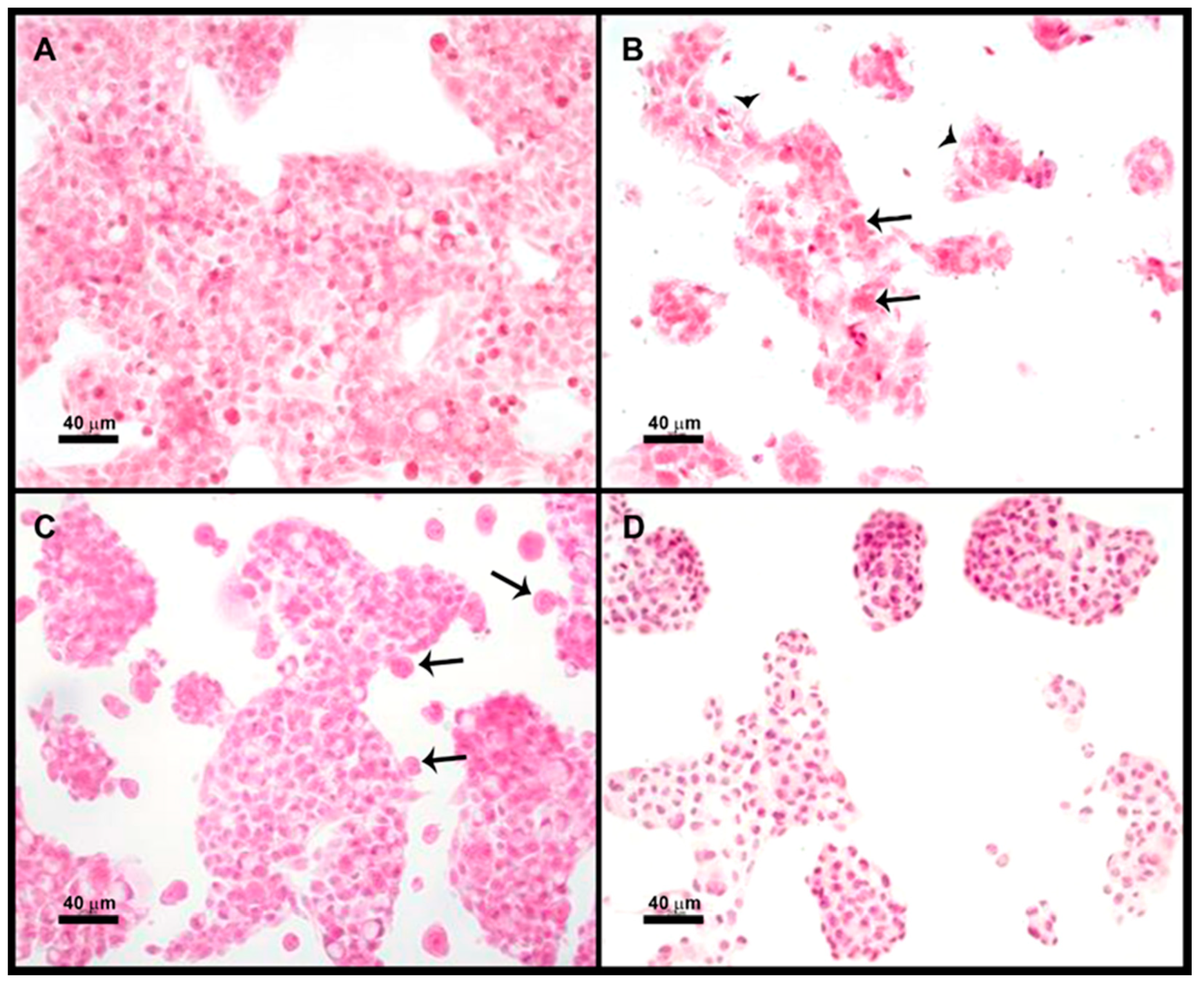
3.4. Optical Microscopy of the E. histolytica Trophozoites: HepG2 Cocultures
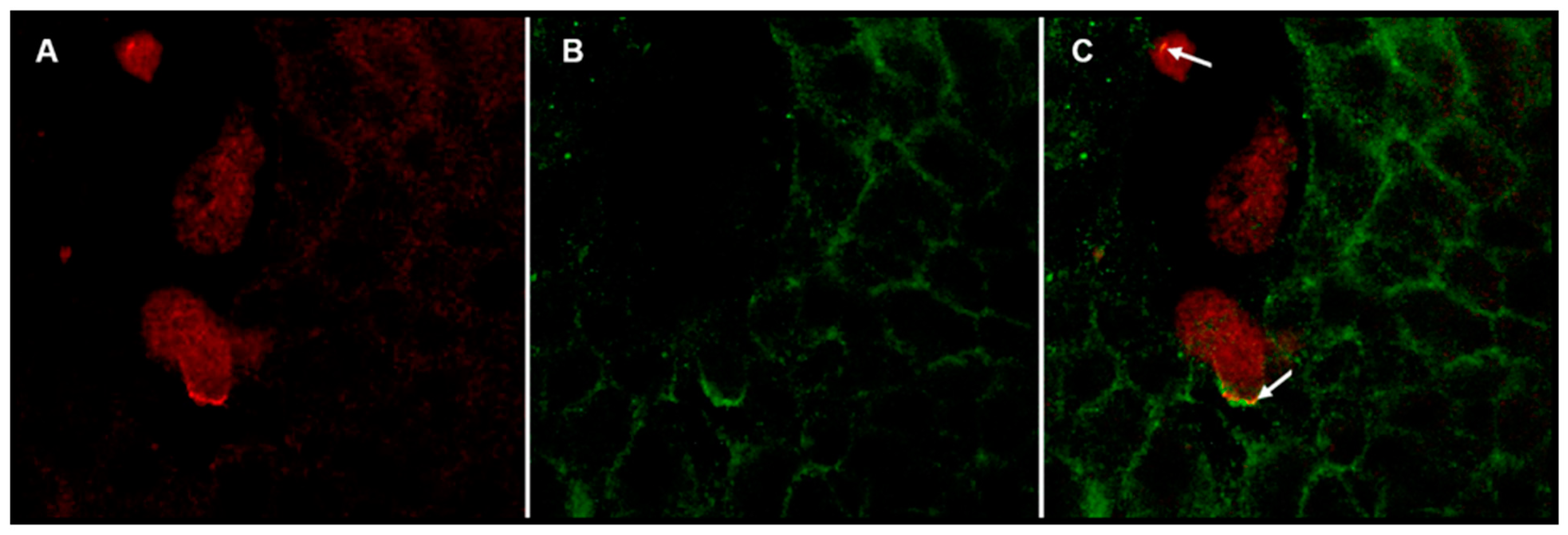
3.5. Fluorescence Confocal Microscopy Study: Colocalization of Gal Lectin and c-Met Receptor
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Petri, W.A.; Haque, R.; Mann, B.J. The bittersweet interface of parasite and host:lectin-carbohydrate interactions during human invasion by the parasite Entamoeba histolytica. Annu. Rev. Microbiol. 2002, 56, 39–64. [Google Scholar] [CrossRef] [PubMed]
- Dodson, J.M.; Lenkowski, P.W.; Eubanks, A.C.; Jackson, T.F.; Napodano, J.; Lyerly, D.M.; Lockhart, L.A.; Mann, B.J.; Petri, W.A. Infection and immunity mediated by the carbohydrate recognition domain of the Entamoeba histolytica Gal/GalNAc lectin. J. Infect. 1999, 179, 460–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ralston, K.S.; Petri, W.A. The ways of a killer: How does Entamoeba histolytica elicit host cell death. Essays Biochem. 2011, 51, 193–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kammanadiminti, S.J.; Mann, B.J.; Dutil, L.; Chadee, K. Regulation of Toll-like receptor-2 expression by the Gal-lectin of Entamoeba histolytica. FASEB J. 2004, 18, 155–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galván, J.M.; Domínguez, M.C.; Meza, I. Pathogenic bacteria prime the induction of Toll-like receptor signalling in human colonic cells by the Gal/GalNAc lectin Carbohydrate Recognition Domain of Entamoeba histolytica. Int. J. Parasitol. 2011, 41, 1101–1112. [Google Scholar] [CrossRef] [PubMed]
- Diamond, L.S.; Harlow, D.R.; Cunic, C.C. A new medium for the axenic cultivation of Entamoeba histolytica and other Entamoeba. Trans. R. Soc. Trop. Med. Hyg. 1978, 72, 431–432. [Google Scholar] [CrossRef]
- López, V.R.; Arreguín, E.R.; González, C.A.; Néquiz, A.M.; García de León, M.C.; Olivos, G.A.; López, V.D.; Pérez, R. Entamoeba histolytica: Expression and localization of Gal/GalNAc lectin in virulent and non-virulent variants from HM1:IMSS strain. Exp. Parasitol. 2010, 125, 244–250. [Google Scholar] [CrossRef] [PubMed]
- Petri, W.A.; Chapman, M.D.; Snodgrass, T.B.; Mann, J.; Broman, J.; Ravdin, J. Subunit structure of the galactose and N-acetyl-D-galactosamine-inhibitable adherence lectin in Entamoeba histolytica. J. Biol. Chem. 1989, 264, 3007–3012. [Google Scholar] [CrossRef]
- Cheng, X.J.; Tsukamoto, H.; Kaneda, Y.; Tachibana, H. Identification of the 150 kDa surface antigen of Entamoeba histolytica as a galactose and N-acetyl-D-galactosamine-inhibitable lectin. Parasitol. Res. 1998, 84, 632–639. [Google Scholar] [CrossRef]
- Welter, B.H.; Goldston, A.M.; Temesvari, L.A. Localisation to lipid rafts correlates with increased function of the Gal/GalNAc lectin in the human protozoan parasite, Entamoeba histolytica. Int. J. Parasitol. 2011, 42, 1409–1419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giordano, S.; Di Renzo, M.F.; Narsimhan, R.P.; Cooper, C.S.; Rosa, C.; Comoglio, P.M. Biosynthesis of the protein encoded by the c-met proto-oncogene. Oncogene 1989, 4, 1383–1388. [Google Scholar] [PubMed]
- López, V.R.; Montfort, I.; Pérez, R. Galactose-specific adhesin and cytotoxicity of Entamoeba histolytica. Parasitol. Res. 2000, 86, 226–231. [Google Scholar] [CrossRef]
- Ravdin, J.I.; Guerrant, R.L. Role of adherence in cytopathogenic mechanisms of Entamoeba histolytica. Study with mammalian tissue culture cells and human erythrocytes. J. Clin. Investig. 1981, 68, 1305–1313. [Google Scholar] [CrossRef] [PubMed]
- Saffer, L.D.; Petri, W.A. Role of the galactose lectin of Entamoeba histolytica in adherence-dependent killing of mammalian cells. Infect. Immun. 1991, 59, 4681–4683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baxt, L.A.; Baker, R.P.; Singh, U.; Urban, S. An Entamoeba histolytica rhomboid protease with atypical specificity cleaves a surface lectin involved in phagocytosis and immune evasion. Genes. Dev. 2008, 22, 1636–1646. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez-Hernández, J.; Retana-González, C.; Ramos-Martínez, E.; Cruz-Colín, J.; Saralegui-Amaro, A.; Baltazar-Rosario, G.; Gutiérrez-Ruíz, C.; Aristi-Urista, G.; López-Vancell, R. Entamoeba histolytica Trophozoites Interact with the c-Met Receptor at the Surface of Liver Origin Cells through the Gal/GalNAc Amoebic Lectin. Life 2021, 11, 923. https://doi.org/10.3390/life11090923
Pérez-Hernández J, Retana-González C, Ramos-Martínez E, Cruz-Colín J, Saralegui-Amaro A, Baltazar-Rosario G, Gutiérrez-Ruíz C, Aristi-Urista G, López-Vancell R. Entamoeba histolytica Trophozoites Interact with the c-Met Receptor at the Surface of Liver Origin Cells through the Gal/GalNAc Amoebic Lectin. Life. 2021; 11(9):923. https://doi.org/10.3390/life11090923
Chicago/Turabian StylePérez-Hernández, Jesus, Clarisa Retana-González, Espiridión Ramos-Martínez, José Cruz-Colín, Andrés Saralegui-Amaro, Gabriela Baltazar-Rosario, Concepción Gutiérrez-Ruíz, Gerardo Aristi-Urista, and Rosario López-Vancell. 2021. "Entamoeba histolytica Trophozoites Interact with the c-Met Receptor at the Surface of Liver Origin Cells through the Gal/GalNAc Amoebic Lectin" Life 11, no. 9: 923. https://doi.org/10.3390/life11090923
APA StylePérez-Hernández, J., Retana-González, C., Ramos-Martínez, E., Cruz-Colín, J., Saralegui-Amaro, A., Baltazar-Rosario, G., Gutiérrez-Ruíz, C., Aristi-Urista, G., & López-Vancell, R. (2021). Entamoeba histolytica Trophozoites Interact with the c-Met Receptor at the Surface of Liver Origin Cells through the Gal/GalNAc Amoebic Lectin. Life, 11(9), 923. https://doi.org/10.3390/life11090923