
Potential Biomarkers of miR-371–373 Gene Cluster in Tumorigenesis


Abstract
:1. Background
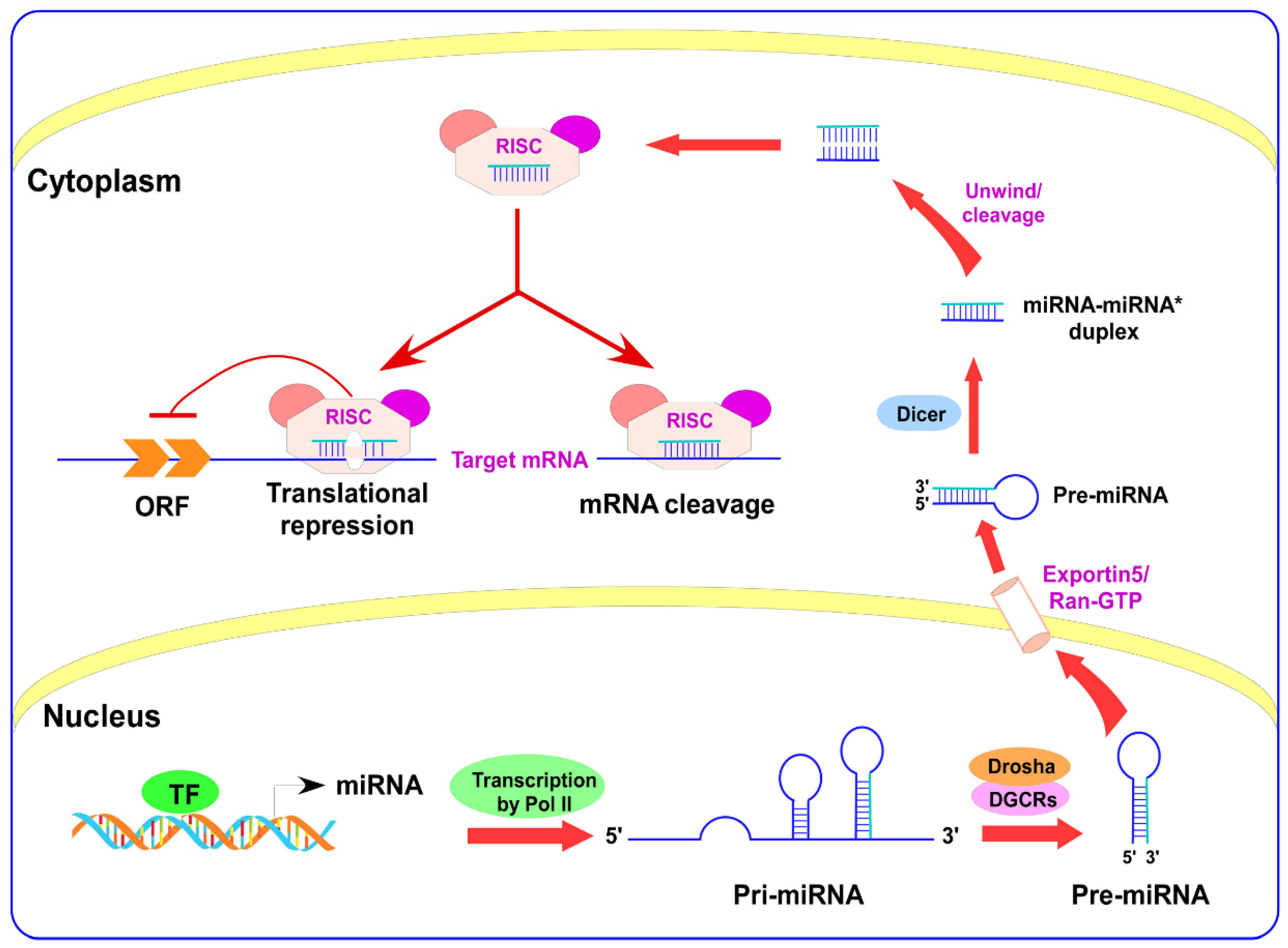
2. miRNAs Biogenesis
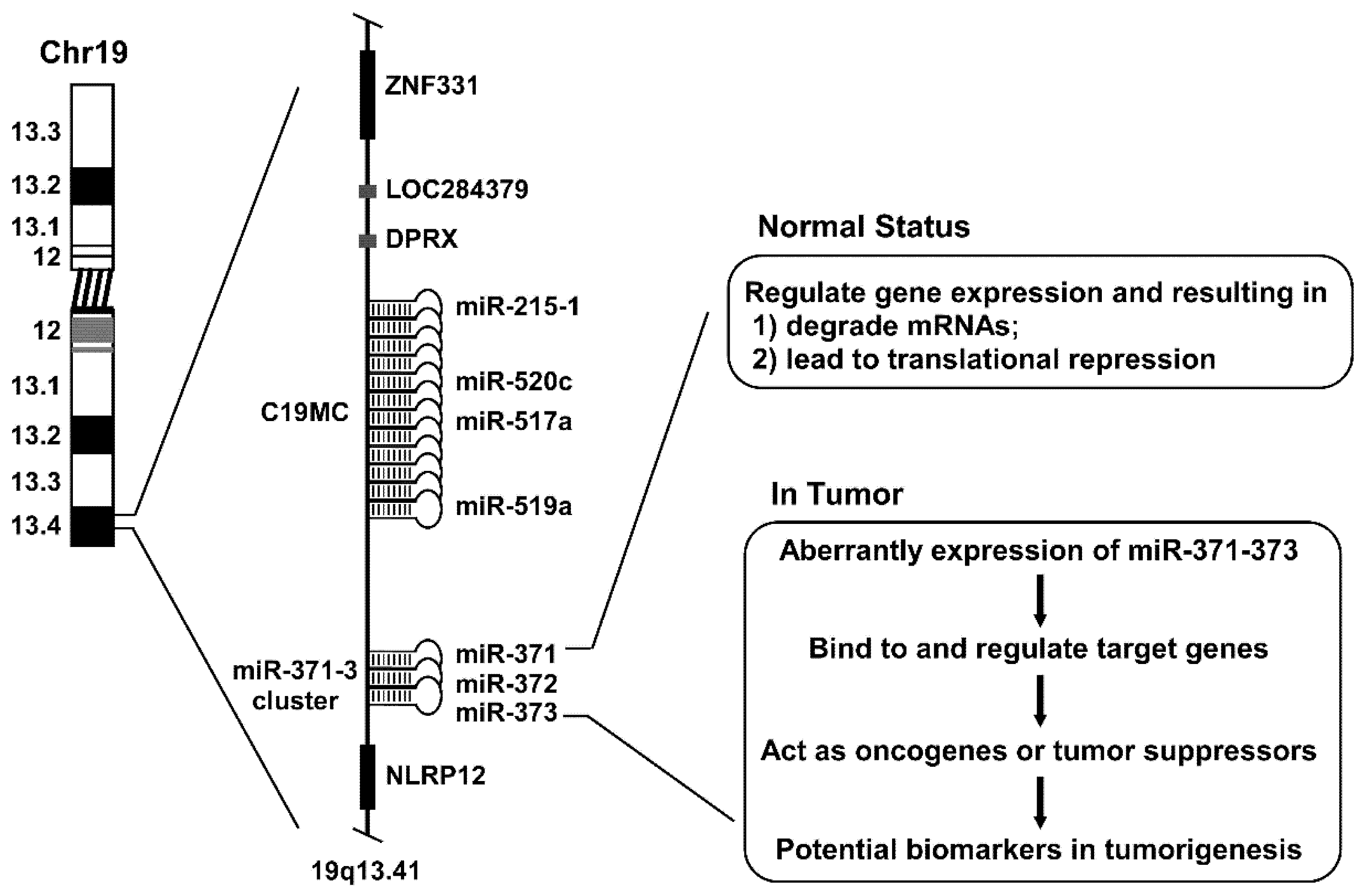
3. miR-371–373 Gene Cluster
4. Functions of miR-371–373 in Tumorigenesis
4.1. Expression Status of miR-371–373 in Different Cancer Cells
4.2. Role of miR-371–373 on Cancer Stem Cells
4.3. miR-371–373 Serves as Oncogenes in Human Cancers
4.4. miR-371–373 Serves as Tumor Suppressors in Human Cancers
4.5. miR-371–373 Targets Cell Cycle-Related Genes
4.6. miR-371–373 Gene Cluster as an Epigenetic Regulator in Cancer
4.7. miR-371–373 as the Target of New Chemotherapeutic Drugs
5. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| miR-371–373 | microRNA-371–373genecluster |
| 3′-UTR | 3′-untranslatedregion |
| ESCs | embryonicstemcells |
| DGCR8 | DiGeorgesyndromecriticalregion8 |
| pri-miRNAs | primarymiRNAs |
| pre-miRNAs | precursormiRNAs |
| RISC | RNA-inducedsilencingcomplex |
| HNSCC | headandnecksquamouscellcarcinoma |
| ATAD2 | ATPasefamilyAAAdomain-containingprotein2 |
| PCa | prostatecancers |
| HPAC | pancreaticadenocarcinoma |
| ULK1 | atumorproteinUNC51-likekinase1 |
| CSCs | cancerstemcells |
| PGCLCs | primordialgermcell-likecells |
| TGFBR2 | TGFbreceptor2 |
| LATS2 | largetumorsuppressorkinase2 |
| TGCTs | testiculargermcelltumors |
| CDK | cyclin |
| FGF9 | fibroblastgrowthfactor9 |
| LSCC | lungsquamouscellcarcinoma |
| EOC | epithelialovariancancer |
| LNSC | lungnon-small-cellcancer |
| RCC | renalcellcarcinoma |
| AKT1 | AKTserine/threoninekinase1 |
| IGF2BP1 | insulin-likegrowthfactor2mRNA-bindingprotein1 |
| FXYD6 | FXYDdomain-containingiontransportregulator6 |
| HDAC | histonedeacetylase |
| TSA | trichostatinA |
| GCTs | germcelltumors |
| ORF | openreadingframe |
| TF | transcriptionfactor. |
References
- Wightman, B.; Ha, I.; Ruvkun, G. Posttranscriptional regulation of the heterochronic gene lin-14 by lin-4 mediates temporal pattern formation in C. elegans. Cell 1993, 75, 855–862. [Google Scholar] [CrossRef]
- Lee, R.C.; Feinbaum, R.L.; Ambros, V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef]
- Lau, N.C.; Lim, L.P.; Weinstein, E.G.; Bartel, D.P. An Abundant Class of Tiny RNAs with Probable Regulatory Roles in Caenorhabditis elegans. Science 2001, 294, 858–862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, R.C.; Ambros, V. An extensive class of small RNAs in Caenorhabditis elegans. Science 2001, 294, 862–864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lagos-Quintana, M.; Rauhut, R.; Lendeckel, W.; Tuschl, T. Identification of novel genes coding for small expressed RNAs. Science 2001, 294, 853–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jansson, M.D.; Lund, A.H. MicroRNA and cancer. Mol. Oncol. 2012, 6, 590–610. [Google Scholar] [CrossRef]
- Lee, Y.; Kim, M.; Han, J.; Yeom, K.H.; Lee, S.; Baek, S.H. MicroRNA genes are transcribed by RNA polymerase II. EMBO J. 2004, 23, 4051–4060. [Google Scholar] [CrossRef]
- Cai, X.; Hagedorn, C.H.; Cullen, B.R. Human microRNAs are processed from capped, polyadenylated transcripts that can also function as mRNAs. RNA 2004, 10, 1957–1966. [Google Scholar] [CrossRef] [Green Version]
- Bracht, J.; Hunter, S.; Eachus, R.; Weeks, P.; Pasquinelli, A.E. Trans-splicing and polyadenylation of let-7 microRNA primary transcripts. RNA 2004, 10, 1586–1594. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.; Ahn, C.; Han, J.; Choi, H.; Kim, J.; Yim, J. The nuclear RNase III Drosha initiates microRNA processing. Nature 2003, 425, 415–419. [Google Scholar] [CrossRef]
- Denli, A.M.; Tops, B.; Plasterk, R.H.A.; Ketting, R.F.; Hannon, G.J. Processing of primary microRNAs by the Microprocessor complex. Nature 2004, 432, 231–235. [Google Scholar] [CrossRef]
- Gregory, R.I.; Yan, K.P.; Amuthan, G.; Chendrimada, T.; Doratotaj, B.; Cooch, N. The Microprocessor complex mediates the genesis of microRNAs. Nature 2004, 432, 235–240. [Google Scholar] [CrossRef]
- Nicholson. A.W. Ribonuclease III mechanisms of double-stranded RNA cleavage. Wiley Interdiscip. Rev. RNA 2014, 5, 31–48. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.A.; Jo, M.H.; Choi, Y.G.; Park, J.; Kwon, S.C.; Hohng, S. Functional anatomy of the human Microprocessor. Cell 2015, 161, 1374–1387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, S.C.; Nguyen, T.A.; Choi, Y.G.; Jo, M.H.; Hohng, S.; Kim, V.N. Structure of human DROSHA. Cell 2015, 164, 81–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okada, C.; Yamashita, E.; Lee, S.J.; Shibata, S.; Katahira, J.; Nakagawa, A.Y. A high-resolution structure of the pre-microRNA nuclear export machinery. Science 2009, 326, 1275–1279. [Google Scholar] [CrossRef]
- Bernstein, E.; Caudy, A.; Hammond, S.M.; Hannon, G.J. Role for a bidentate ribonuclease in the initiation step of RNA interference. Nat. Cell Biol. 2001, 409, 363–366. [Google Scholar] [CrossRef] [PubMed]
- Grishok, A.; Pasquinelli, A.E.; Conte, D.; Li, N.; Parrish, S.; Ha, I. Genes and mechanisms related to RNA interference regulate expression of the small temporal RNAs that control C. elegans developmental timing. Cell 2001, 106, 23–34. [Google Scholar] [CrossRef] [Green Version]
- Hutvágner, G.; McLachlan, J.; Pasquinelli, A.E.; Bálint, E.; Tuschl, T.; Zamore, P.D. A cellular function for the RNA-interference enzyme Dicer in the maturation of the let-7 small temporal RNA. Science 2001, 293, 834–838. [Google Scholar] [CrossRef] [Green Version]
- Lau, P.-W.; Guiley, K.Z.; De, N.; Potter, C.S.; Carragher, B.; Macrae, I.J. The molecular architecture of human Dicer. Nat. Struct. Mol. Biol. 2012, 19, 436–440. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; A Kolb, F.; Jaskiewicz, L.; Westhof, E.; Filipowicz, W. Single Processing Center Models for Human Dicer and Bacterial RNase III. Cell 2004, 118, 57–68. [Google Scholar] [CrossRef] [Green Version]
- Achard, P.; Herr, A.; Baulcombe, D.; Harberd, N.P. Modulation of floral development by a gibberellin-regulated microRNA. Development 2004, 131, 3357–3365. [Google Scholar] [CrossRef] [Green Version]
- Gregory, R.I.; Chendrimada, T.P.; Shiekhattar, R. MicroRNA biogenesis: isolation and characterization of the microprocessor complex. Methods Mol. Biol. 2006, 342, 33–47. [Google Scholar]
- Yu, Y.-H.; Kuo, H.-K.; Chang, K.-W. The Evolving Transcriptome of Head and Neck Squamous Cell Carcinoma: A Systematic Review. PLoS ONE 2008, 3, e3215. [Google Scholar] [CrossRef]
- Voorhoeve, P.M.; le Sage, C.; Schrier, M.; Gillis, A.J.; Stoop, H.; Nagel, R.; Liu, Y.P.; Van Duijse, J.; Drost, J.; Griekspoor, A.; et al. A Genetic Screen Implicates miRNA-372 and miRNA-373 As Oncogenes in Testicular Germ Cell Tumors. Cell 2006, 124, 1169–1181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suh, M.R.; Lee, Y.; Kim, J.Y.; Kim, S.K.; Moon, S.H.; Lee, J.Y. Human embryonic stem cells express a unique set of microRNAs. Dev. Biol. 2004, 270, 488–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, G.; Liu, H.; He, H.; Wang, Y.; Lu, X.; Yu, Y. miR-372 down-regulates the oncogene ATAD2 to influence hepatocellular carcinoma proliferation and metastasis. BMC Cancer 2014, 14, 107. [Google Scholar] [CrossRef] [Green Version]
- Huang, Q.; Gumireddy, K.; Schrier, M.; le Sage, C.; Nagel, R.; Nair, S. The microRNAs miR-373 and miR-520c promote tumour invasion and metastasis. Nat. Cell Biol. 2008, 10, 202–210. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Wilson, M.J. miR-520c and miR-373 upregulate MMP9 expression by targeting mTOR and SIRT1, and activate the Ras/Raf/MEK/Erk signaling pathway and NF-kappaB factor in human fibrosarcoma cells. J. Cell Physiol. 2012, 227, 867–876. [Google Scholar] [CrossRef] [Green Version]
- Keklikoglou, I.; Koerner, C.; Schmidt, C.; Zhang, J.D.; Heckmann, D.; Shavinskaya, A. MicroRNA-520/373 family functions as a tumor suppressor in estrogen receptor negative breast cancer by targeting NF-kappaB and TGF-beta signaling pathways. Oncogene 2012, 31, 4150–4163. [Google Scholar] [CrossRef]
- Wu, S.; Aksoy, M.; Shi, J.; Houbaviy, H.S. Evolution of the miR-290–295/miR-371-373 Cluster Family Seed Repertoire. PLoS ONE 2014, 9, e108519. [Google Scholar] [CrossRef]
- Houbaviy, H.B.; Dennis, L.; Jaenisch, R.; Sharp, P.A. Characterization of a highly variable eutherian microRNA gene. RNA 2005, 11, 1245–1257. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Lu, G.; Ke, X.; Lu, X.; Wang, X.; Li, H. miR-488 acts as a tumor suppressor gene in gastric cancer. Tumor Biol. 2016, 37, 8691–8698. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Hu, Y.; Ju, J.; Hou, L.; Li, Z.; Xiao, D. Downregulation of miR-522 suppresses proliferation and metastasis of non-small cell lung cancer cells by directly targeting DENN/MADD domain containing 2D. Sci Rep. 2016, 6, 19346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Song, G.; Tan, W.; Qi, M.; Zhang, L.; Chan, J. miR-573 inhibits prostate cancer metastasis by regulating epithelial-mesenchymal transition. Oncotarget 2015, 6, 35978–35990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, X.; Qian, X.; Duan, L.; Liu, H.; Zhu, Y.; Qi, J. microRNA-372 Suppresses Migration and Invasion by Targeting p65 in Human Prostate Cancer Cells. DNA Cell Biol. 2016, 35, 828–835. [Google Scholar] [CrossRef] [Green Version]
- Nakata, K.; Ohuchida, K.; Mizumoto, K.; Aishima, S.; Oda, Y.; Nagai, E.; Tanaka, M. Micro RNA-373 is Down-regulated in Pancreatic Cancer and Inhibits Cancer Cell Invasion. Ann. Surg. Oncol. 2014, 21, 564–574. [Google Scholar] [CrossRef]
- Cho, W.J.; Shin, J.M.; Kim, J.S.; Lee, M.R.; Hong, K.S.; Lee, J.H. miR-372 regulates cell cycle and apoptosis of ags human gastric cancer cell line through direct regulation of LATS2. Mol. Cells 2009, 28, 521–527. [Google Scholar] [CrossRef]
- Feber, A.; Xi, L.; Luketich, J.D.; Pennathur, A.; Landreneau, R.J.; Wu, M. MicroRNA expression profiles of esophageal cancer. J. Thorac. Cardiovasc. Surg. 2008, 135, 255–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghasemi, M.; Samaei, N.M.; Mowla, S.J.; Shafiee, M.; Vasei, M.; Ghasemian, N. Upregulation of miR-371-373 cluster, a human embryonic stem cell specific microRNA cluster, in esophageal squamous cell carcinoma. J. Can. Res. Ther. 2018, 14, S132–S137. [Google Scholar]
- Chen, H.; Zhang, Z.; Lu, Y.; Song, K.; Liu, X.; Xia, F. Downregulation of ULK1 by microRNA-372 inhibits the survival of human pancreatic adenocarcinoma cells. Cancer Sci. 2017, 108, 1811–1819. [Google Scholar] [CrossRef] [Green Version]
- Qu, H.-W.; Jin, Y.; Cui, Z.-L.; Jin, X.-B. MicroRNA-373-3p inhibits prostate cancer progression by targeting AKT1. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 6252–6259. [Google Scholar]
- Vincent, K.; Pichler, M.; Lee, G.W.; Ling, H. MicroRNAs, genomic instability and cancer. Int. J. Mol. Sci. 2014, 15, 14475–14491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, L.; Thomson, J.M.; Hemann, M.T.; Hernando-Monge, E.; Mu, D. A microRNA polycistron as a potential human oncogene. Nature 2005, 435, 828–833. [Google Scholar] [CrossRef]
- Takamizawa, J.; Konishi, H.; Yanagisawa, K.; Tomida, S.; Osada, H.; Endoh, H. Reduced expression of the let-7 microRNAs in human lung cancers in association with shortened postoperative survival. Cancer Res. 2004, 64, 3753–3756. [Google Scholar] [CrossRef] [Green Version]
- Yekta, S.; Shih, I.H.; Bartel, D.P. MicroRNA-directed cleavage of HOXB8 mRNA. Science 2004, 304, 594–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, L.P.; Lau, N.C.; Garrett-Engele, P.; Grimson, A.; Schelter, J.M.; Castle, J. Microarray analysis shows that some microRNAs downregulate large numbers of target mRNAs. Nature 2005, 433, 769–773. [Google Scholar] [CrossRef] [PubMed]
- PillaI, R.S.; Bhattacharyya, S.N.; Artus, C.G.; Zoller, T.; Cougot, N.; Basyuk, E. Inhibition of translational initiation by Let-7 microRNA in human cells. Science 2005, 309, 1573–1576. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Teruya-Feldstein, J.; Weinberg, R.A. Tumour invasion and metastasis initiated by microRNA-10b in breast cancer. Nature 2007, 449, 682–688. [Google Scholar] [CrossRef]
- Place, R.F.; Li, L.-C.; Pookot, D.; Noonan, E.J.; Dahiya, R. MicroRNA-373 induces expression of genes with complementary promoter sequences. Proc. Natl. Acad. Sci. USA 2008, 105, 1608–1613. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.; Pestell, T.G.; Lisanti, N.P.; Pestell, R.G. Cancer stem cells. Int. J. Biochem. Cell. Biol. 2012, 44, 2144–2151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, J.; Getz, G.; Miska, E.A.; Alvarez-Saavedra, E.; Lamb, J.; Peck, D. MicroRNA expression profiles classify human cancers. Nature 2005, 435, 834–838. [Google Scholar] [CrossRef] [PubMed]
- DeSano, J.T.; Xu, L. MicroRNA Regulation of Cancer Stem Cells and Therapeutic Implications. AAPS J. 2009, 11, 682–692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, M.; Kaufman, M. Establishment in culture of pluripotent cells from mouse embryos. Nature 1981, 292, 154–156. [Google Scholar] [CrossRef] [PubMed]
- Martin, G. Isolation of a pluripotent cell line from early mouse embryos cultured in medium conditioned by teratocarcinoma stem cells. Proc. Natl. Acad. Sci. USA 1981, 78, 7634–7638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, A. Embryonic Stem Cells. In Stem Cell Biology; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2001. [Google Scholar]
- Houbaviy, H.B.; Murray, M.F.; Sharp, P.A. Embryonic stem cell-specific MicroRNAs. Dev. Cell 2003, 5, 351–358. [Google Scholar] [CrossRef]
- Wong, D.J.; Segal, E.; Chang, H.Y. Stemness, cancer and cancer stem cells. Cell Cycle 2008, 7, 3622–3624. [Google Scholar] [CrossRef]
- Landgraf, P.; Rusu, M.; Sheridan, R.; Sewer, A.; Iovino, N.; Aravin, A. A mammalian microRNAexpression atlas based on small RNA librarysequencing. Cell 2007, 129, 1401–1414. [Google Scholar] [CrossRef] [Green Version]
- Wienholds, E.; Kloosterman, W.P.; Miska, E.; AlvarezSaavedra, E.; Berezikov, E.; de Bruijn, E. MicroRNAexpression in zebrafish embryonic development. Science 2005, 309, 310–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivey, K.N.; Srivastava, D. MicroRNAs as regulators of differentiation and cell fate decisions. Cell Stem Cell 2010, 7, 36–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, A.D.; Diao, L.T.; Xu, H.; Xiao, Z.D.; Li, J.H.; Zhou, H. β-Catenin/LEF1 transactivates the microRNA-371-373 cluster that modulates the Wnt/s-catenin-signaling pathway. Oncogene 2012, 31, 2968–2978. [Google Scholar] [CrossRef] [Green Version]
- Reya, T.; Morrison, S.J.; Clarke, M.F.; Weissman, I.L. Stem cells, cancer, and cancer stem cells. Nature 2001, 414, 105–111. [Google Scholar] [CrossRef] [Green Version]
- Zhao, R.C.; Zhu, Y.S.; Shi, Y. New hope for cancer treatment: Exploring the distinction between normal adult stem cells and cancer stem cells. Pharmacol Ther. 2008, 119, 74–82. [Google Scholar] [CrossRef]
- Luu, H.H.; Zhang, R.; Haydon, R.C.; Rayburn, E.; Kang, Q.; Si, W. Wnt/beta-catenin signaling pathway as a novel cancer drug target. Curr. Cancer Drug Targets 2004, 4, 653–671. [Google Scholar] [CrossRef]
- Wang, L.Q.; Yu, P.; Li, B.; Guo, Y.H.; Liang, Z.R.; Zheng, L.L.Y. miR-372 and miR-373 enhance the stemness of colorectal cancer cells by repressing differentiation signaling pathways. Mol. Oncol. 2018, 12, 1949–1964. [Google Scholar] [CrossRef] [PubMed]
- Qi, J.; Yu, J.Y.; Shcherbata, H.R.; Mathieu, J.; Wang, A.J.; Seal, S. microRNAs regulate human embryonic stem cell division. Cell Cycle 2009, 8, 3729–3741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, N.D.; Kissner, M.; Subramanyam, D.; Parchem, R.J.; Laird, D.J.; Blelloch, R.H. A miR-372/let-7 Axis Regulates Human Germ Versus Somatic Cell Fates. Stem Cells 2016, 34, 1985–1991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, S.-Y.; Xu, P.-F.; Gao, T.-T. MiR-372-3p inhibits the growth and metastasis of osteosarcoma cells by targeting FXYD6. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 62–69. [Google Scholar]
- Calin, G.A.; Liu, C.G.; Sevignani, C.; Ferracin, M.; Felli, N.; Dumitru, C.D. MicroRNA profiling reveals distinct signatures in B cell chronic lymphocytic leukemias. Proc. Natl. Acad. Sci. USA 2004, 101, 11755–11760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volinia, S.; Calin, G.A.; Liu, C.G.; Ambs, S.; Cimmino, A.; Petrocca, F. A microRNA expression signature of human solid tumors defines cancer gene targets. Proc. Natl. Acad. Sci. USA 2006, 103, 2257–2261. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.H.; Goan, Y.G.; Hsiao, M.; Lee, C.H.; Jian, S.H.; Lin, J.T. MicroRNA-373 (miR-373) post-transcriptionally regulates large tumor suppressor, homolog 2 (LATS2) and stimulates proliferation in human esophageal cancer. Exp. Cell Res. 2009, 315, 2529–2538. [Google Scholar] [CrossRef]
- Guo, Y.; Chen, Z.; Zhang, L.; Zhou, F.; Shi, S.; Feng, X. Distinctive microRNA profiles relating to patient survival in esophageal squamous cell carcinoma. Cancer Res. 2008, 68, 26–33. [Google Scholar] [CrossRef] [Green Version]
- Gillis, A.J.; Stoop, H.J.; Hersmus, R.; Oosterhuis, J.W.; Sun, Y.; Chen, C. High-throughput microRNAome analysis in human germcell tumours. J. Pathol. 2007, 213, 319–328. [Google Scholar] [CrossRef] [PubMed]
- Maesawa, C.; Tamura, G.; Suzuki, Y.; Ogasawara, S.; Ishida, K.; Saito, K. Aberrations of tumor-suppressor genes (p53, apc, mcc and Rb) in esophageal squamous-cell carcinoma. Int. J. Cancer. 1994, 57, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yang, J.; Cui, X.; Chen, Y.; Zhu, V.F.; Hagan, J.P. A novel epigenetic CREB-miR-373 axis mediates ZIP4-induced pancreatic cancer growth. EMBO Mol. Med. 2013, 5, 1322–1334. [Google Scholar] [CrossRef]
- Zhou, C.; Li, X.; Zhang, X.; Liu, X.; Tan, Z.; Yang, C. microRNA-372 maintains oncogene characteristics bytargeting TNFAIP1 and affects NFκB signaling in humangastric carcinoma cells. Int. J. Oncol. 2013, 42, 635–642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marek, L.; Ware, K.E.; Fritzsche, A.; Hercule, P.; Helton, W.R.; Smith, J.E. Fibroblast growth factor (FGF) and FGF receptor-mediated autocrine signaling in non-small-cell lung cancer cells. Mol. Pharmacol. 2009, 75, 196–207. [Google Scholar] [CrossRef] [PubMed]
- Ohgino, K.; Soejima, K.; Yasuda, H.; Hayashi, Y.; Hamamoto, J.; Naoki, K. Expression of fibroblast growth factor 9 is associated with poor prognosis in patients with resected non-small cell lung cancer. Lung Cancer 2014, 83, 90–96. [Google Scholar] [CrossRef]
- Li, Z.G.; Mathew, P.; Yang, J.; Starbuck, M.W.; Zurita, A.J.; Liu, J. Androgen receptor-negative human prostate cancer cells induce osteogenesis in mice through FGF9-mediated mechanisms. J. Clin. Investig. 2008, 118, 2697–2710. [Google Scholar] [CrossRef] [Green Version]
- Hendrix, N.D.; Wu, R.; Kuick, R.; Schwartz, D.R.; Fearon, E.R.; Cho, K.R. Fibroblast growth factor 9 has oncogenic activity and is a downstream target of Wnt signaling in ovarian endometrioid adenocarcinomas. Cancer Res. 2006, 66, 1354–1362. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Liu, S.; Zhao, X.; Wang, Y.; Tian, D.; Jiang, W. miR-372-3p promotes cell growth and metastasis by targeting FGF9 in lung squamous cell carcinoma. Cancer Med. 2017, 6, 1323–1330. [Google Scholar] [CrossRef]
- Yang, K.; Handorean, A.M.; Iczkowski, K.A. MicroRNAs 373 and 520c Are Downregulated in Prostate Cancer, Suppress CD44 Translation and Enhance Invasion of Prostate Cancer Cells in vitro. Int. J. Clin. Exp. Pathol. 2008, 2, 361–369. [Google Scholar]
- Zhang, Y.; Zhao, F.J.; Chen, L.L.; Wang, L.Q.; Nephew, K.P.; Wu, Y.L. miR-373 targeting of the Rab22a oncogene suppresses tumor invasion and metastasis in ovarian cancer. Oncotarget 2014, 5, 12291–12303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harel, S.A.; Ben-Moshe, N.B.; Aylon, Y.; Bublik, D.R.; Moskovits, N.; Toperoff, G. Reactivation of epigenetically silenced miR-512 and miR-373 sensitizes lung cancer cells to cisplatin and restricts tumor growth. Cell Death Differ. 2015, 22, 1328–1340. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Huang, M.; Kong, L.; Li, Y. miR-372 suppresses tumour proliferation and invasion by targeting IGF2BP1 in renal cell carcinoma. Cell Prolif. 2015, 48, 593–599. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The Next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, W.; Sun, M.; Zou, G.-M.; Chen, J. MicroRNA and cancer: Current status and prospective. Int. J. Cancer 2006, 120, 953–960. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.-K.; Tan, E.-L.; Gan, S.-Y. Elucidating the roles of miR-372 in cell proliferation and apoptosis of nasopharyngeal carcinoma TW01 cells. Exp. Oncol. 2014, 36, 170–173. [Google Scholar]
- Seol, H.S.; Akiyama, Y.; Shimada, S.; Lee, H.J.; Kim, T.I.; Chun, S.M. Epigenetic silencing of microRNA-373 to epithelial-mesenchymal transition in non-small cell lung cancer through IRAK2 and LAMP1 axes. Cancer Lett. 2014, 353, 232–241. [Google Scholar] [CrossRef] [PubMed]
- Gutschner, T.; Hammerle, M.; Pazaitis, N.; Bley, N.; Fiskin, E.; Uckelmann, H. Insulin-like growth factor 2 mRNA-binding protein 1 (IGF2BP1) is an important protumorigenic factor in hepatocellular carcinoma. Hepatology 2014, 59, 1900–1911. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Hannon, G.J. MicroRNAs: small RNAs with a big role in gene regulation. Nat. Rev. Genet. 2004, 5, 522–531. [Google Scholar] [CrossRef] [PubMed]
- Radtke, A.; Cremers, J.F.; Kliesch, S.; Riek, S.; Junker, K.; Mohamed, S.A. Can germ cell neoplasia in situ be diagnosed by measuring serum levels of microRNA371a-3p? J. Cancer Res. Clin. Oncol. 2017, 143, 2383–2392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, W.; He, X.; Kong, J.; Ye, B. miR-373 affects human lung cancer cells’ growth and its E-cadherin expression. Oncol Res. 2012, 20, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Ding, M.; Lu, X.; Wang, C.; Zhao, Q.; Ge, J.; Xia, Q. The E2F1–miR-520/372/373–SPOP Axis Modulates Progression of Renal Carcinoma. Cancer Res. 2018, 78, 6771–6784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ullmann, P.; Rodriguez, F.; Schmitz, M.; Meurer, S.K.; Qureshi-Baig, K.; Felten, P. The miR-371-373 cluster represses colon cancer initiation and metastatic colonization by inhibiting the TGFBR2/ID1 signaling Axis. Cancer Res. 2018, 78, 3793–3808. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Yu, H.; Liu, Y.; Liu, X.; Zheng, J.; Ma, J. Long Non-Coding RNA HOXA-AS2 Regulates Malignant Glioma Behaviors and Vasculogenic Mimicry Formation via the miR-373/EGFR Axis. Cell. Physiol. Biochem. 2018, 45, 131–147. [Google Scholar] [CrossRef]
- Feng, L.; Ma, Y.; Sun, J.; Shen, Q.; Liu, L.; Lu, H. YY1-miR-372-SQSTM1 regulatory axis in autophagy. Autophagy 2014, 10, 1442–1453. [Google Scholar] [CrossRef] [Green Version]
- Yeh, L.-Y.; Liu, C.-J.; Wong, Y.-K.; Chang, C.; Lin, S.-C.; Chang, K.-W. miR-372 inhibits p62 in head and neck squamous cell carcinoma in vitro and in vivo. Oncotarget 2015, 6, 6062–6075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Hao, B.; Han, G.; Liu, Y.; Dai, D.; Li, Y. miR-372 regulates glioma cell proliferation and invasion by directly targeting PHLPP2. J. Cell. Biochem. 2015, 116, 225–232. [Google Scholar] [CrossRef]
- Chen, X.; Sun, M.; Hu, Y.; Zhang, H.; Wang, Z.; Zhou, N. FXYD6 is a new biomarker of cholangiocarcinoma. Oncol Lett. 2014, 7, 393–398. [Google Scholar] [CrossRef] [Green Version]
- Tian, R.Q.; Wang, X.H.; Hou, L.J.; Jia, W.H.; Yang, Q.; Li, Y.X. MicroRNA-372 is down-regulated and targets cyclin-dependent kinase 2 (CDK2) and cyclin A1 in human cervical cancer, which may contribute to tumorigenesis. J. Biol. Chem. 2011, 286, 25556–25563. [Google Scholar] [CrossRef] [Green Version]
- Nakano, H.; Miyazawa, T.; Kinoshita, K.; Yamada, Y.; Yoshida, T. Functional screening identifies a microRNA, miR-491 that induces apoptosis by targeting Bcl-X (L) in colorectal cancer cells. Int. J. Cancer. 2010, 127, 1072–1080. [Google Scholar] [CrossRef] [PubMed]
- Cairo, S.; Wang, Y.; de Reynies, A.; Duroure, K.; Dahan, J.; Redon, M.-J. Stem cell-like micro-RNA signature driven by Myc in aggressive liver cancer. Proc. Natl. Acad. Sci. USA 2010, 107, 20471–20476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.-L.; Sun, K.-X.; Zong, Z.-H.; Chen, S.; Zhao, Y. MicroRNA-372 inhibits endometrial carcinoma development by targeting the expression of the Ras homolog gene family member C (RhoC). Oncotarget 2015, 7, 6649–6664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holliday, R. The inheritance of epigenetic defects. Science 1987, 238, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Holliday, R. Epigenetics: An overview. Dev. Genet. 1994, 15, 453–457. [Google Scholar] [CrossRef]
- Wu, C.; Morris, J.R. Genes, genetics, and epigenetics: A correspondence. Science 2001, 293, 1103–1105. [Google Scholar] [CrossRef] [Green Version]
- Mersfelder, E.L.; Parthun, M.R. The tale beyond the tail: histone core domain modifications and the regulation of chromatin structure. Nucleic Acids Res. 2006, 34, 2653–2662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, J.; Wang, F.; Cai, Y.; Jin, J. The Functional Analysis of Histone Acetyltransferase MOF in Tumorigenesis. Int. J. Mol. Sci. 2016, 17, 99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernstein, E.; Allis, C.D. Rna meets chromatin. Genes Dev. 2005, 19, 1635–1655. [Google Scholar] [CrossRef] [Green Version]
- Peng, Y.; Croce, C.M. The role of MicroRNAs in human cancer. Signal Transduct. Target. Ther. 2016, 1, 15004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lovat, F.; Valeri, N.; Croce, C.M. MicroRNAs in the Pathogenesis of Cancer. Semin. Oncol. 2011, 38, 724–733. [Google Scholar] [CrossRef]
- Di Leva, G.; Garofalo, M.; Croce, C.M. MicroRNAs in cancer. Annu. Rev. Pathol. 2014, 9, 287–314. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.; Kelly, T.K.; Jones, P.A. Epigenetics in cancer. Carcinogenesis 2010, 31, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Sandoval, J.; Esteller, M. Cancer epigenomics: beyond genomics. Curr. Opin. Genet. Dev. 2012, 22, 50–55. [Google Scholar] [CrossRef]
- Pfister, S.X.; Ahrabi, S.; Zalmas, L.P.; Sarkar, S.; Aymard, F.; Bachrati, C.Z. SETD2-dependent histone H3K36 trimethylation is required for homologous recombination repair and genome stability. Cell Rep. 2014, 7, 2006–2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, W.; Jiang, K.; Shen, M.; Qian, Y.; Peng, Y. SIRT2 suppresses non-small cell lung cancer growth by targeting JMJD2A. Biol. Chem. 2015, 396, 929–936. [Google Scholar] [CrossRef] [PubMed]
- An, J.; Xu, J.; Li, J.; Jia, S.; Li, X.; Lu, Y. HistoneH3 demethylase JMJD2A promotes growth of liver cancercells through up-regulating miR372. Oncotarget 2017, 8, 49093–49109. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Gao, W.; Luo, J.; Tian, R.; Sun, H.; Zou, S. Methyl-CpG binding protein MBD2 is implicated in methylation-mediated suppression of miR-373 in hilar cholangiocarcinoma. Oncol Rep. 2011, 25, 443–451. [Google Scholar] [CrossRef]
- Chen, Y.; Luo, J.; Tian, R.; Sun, H.; Zou, S. miR-373 negatively regulates methyl-CpGbinding domain protein 2 (MBD2) in hilar cholangiocarcinoma. Dig. Dis. Sci. 2011, 56, 1693–1701. [Google Scholar] [CrossRef]
- Lujambio, A.; Ropero, S.; Ballestar, E.; Fraga, M.F.; Cerrato, C.; Setien, F. Genetic unmasking of an epigenetically silenced microRNA in human cancer cells. Cancer Res. 2007, 67, 1424–1429. [Google Scholar] [CrossRef] [Green Version]
- Anfossi, S.; Babayan, A.; Pantel, K.; Calin, G.A. Clinical utility of circulating non-coding RNAs- an update. Nat. Rev. Clin. Oncol. 2018, 15, 541–563. [Google Scholar] [CrossRef]
- Xi, X.; Li, T.; Huang, Y.; Sun, J.; Zhu, Y.; Yang, Y. RNA Biomarkers: Frontier of Precision Medicine for Cancer. Non-Coding RNA 2017, 3, 9. [Google Scholar] [CrossRef]
- Ramalho-Carvalho, J.; Fromm, B.; Henrique, R.; Jerónimo, C. Deciphering the function of non-coding RNAs in prostate cancer. Cancer Metastasis Rev. 2016, 35, 235–262. [Google Scholar] [CrossRef] [PubMed]
- Murray, M.J.; Halsall, D.J.; Hook, C.E.; Williams, D.M.; Nicholson, J.C.; Coleman, N.; Sweet, W.; Duh, Y.-J.; Greenfield, L.; Tarco, E.; et al. Identification of MicroRNAs From the miR-371∼373 and miR-302 Clusters as Potential Serum Biomarkers of Malignant Germ Cell Tumors. Am. J. Clin. Pathol. 2011, 135, 119–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belge, G.; Dieckmann, K.P.; Spiekermann, M.; Balks, T.; Bullerdiek, J. Serum levels of microRNAs miR-371-3: A novel class of serum biomarkers for testicular germ cell tumors? Eur. Urol. 2012, 61, 1068–1069. [Google Scholar] [CrossRef] [PubMed]
- Syring, I.; Bartels, J.; Holdenrieder, S.; Kristiansen, G.; Muller, S.C.; Ellinger, J. Circulating serum miRNA (miR-367-3p, miR-371a-3p, miR-372-3p and miR-373-3p) as biomarkers in patients with testicular germ cell cancer. J. Urol. 2015, 193, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Domínguez-Vigil, I.G.; Moreno-Martínez, A.K.; Wang, J.Y.; Roehrl, M.H.A.; Barrera-Saldaña, H.A. The dawn of the liquid biopsy in the fight against cancer. Oncotarget 2017, 9, 2912–2922. [Google Scholar] [CrossRef] [Green Version]
- Dieckmann, K.P.; Radtke, A.; Spiekermann, M.; Balks, T.; Matthies, C.; Becker, P. Serum levels of microRNA miR-371a-3p: a sensitive and specific new biomarker for germ cell tumours. Eur. Urol. 2017, 71, 213–220. [Google Scholar] [CrossRef] [Green Version]
- Henrique, R.; Jeronimo, C. Testicular Germ Cell Tumors Go Epigenetics: Will miR-371a-3p Replace Classical Serum Biomarkers? Eur. Urol. 2017, 71, 221–222. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| miRNAs | Tumor Tissues or Cancer Cells | Target Genes | Exp. of Targets | Consequent | Ref |
|---|---|---|---|---|---|
| mir-372 | Prostate cancer | p65 | Down | Acts as a tumor suppressor | [30,85] |
| Liver carcinoma | ATAD2 | Down | Acts as a tumor suppressor | [32] | |
| Gastric carcinoma, ESCC | LATS2 | Down | Acts as an oncogene | [38,72] | |
| Colorectal cancer | NFkB, MAPK/Erk, and VDR | Down | Enhances the stemness | [65] | |
| Osteosarcoma tissues | FXYD6 | Down | Acts as a tumor suppressor | [68] | |
| TGCT | LATS2 | Down | Acts as an oncogene | [73,74,75] | |
| Gastric cancer | TNFAIP1 | Down | Acts as an oncogene | [77] | |
| Lung carcinoma | FGF9 | Up | Acts as an oncogene | [82] | |
| SCC | p62 | Up | Acts as a tumor suppressor | [86] | |
| Glioma tissues | PHLPP2 | Up | Acts as an oncogene | [87] | |
| Cervical, liver cancers, EC, HeLa, germ cell, | CDK2,Cyclin A1 | Down | Acts as a tumor suppressor | [88,89] | |
| Renal cell carcinoma | MBD2 | Down | Acts as a tumor suppressor | [90] | |
| mir-373 | Fibrosarcoma | mTOR, SIRT1 | Down | Acts as an oncogene | [29] |
| Breast cancer | TGFB, NFkB | Up | Acts as a tumor suppressor | [30] | |
| Esophageal cancer | LATS2 | Up | Acts as an oncogene | [71] | |
| Testicular germ cell tumors | LATS2 | Down | Acts as an oncogene | [73,74,75] | |
| Pancreatic cancer | ZIP4 | Up | Acts as an oncogene | [76] | |
| Prostate cancer | CSDC2 | Up | Acts as a tumor suppressor | [83] | |
| Prostate cancer tissue | CD44 | Down | Acts as an oncogene | [83] | |
| Ovarian cancer | Rab22A | Down | Acts as a tumor suppressor | [84] | |
| Prostate cancerA549 cell | E-cadherin | Up | Acts as a tumor suppressor | [83,84] | |
| Urinary bladder cancer | EGFR | Up | Acts as a tumor suppressor | [91] | |
| Hilar cholangiocarcinoma | MBD2 | Down | Acts as a tumor suppressor | [90] | |
| Lung cancer | IRAK2, LAMP1 | Down | Acts as a tumor suppressor | [90,92] | |
| Lung cancer | RelA, PIK3CA | Down | Acts as a tumor suppressor | [85] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shah, J.A.; Khattak, S.; Rauf, M.A.; Cai, Y.; Jin, J. Potential Biomarkers of miR-371–373 Gene Cluster in Tumorigenesis. Life 2021, 11, 984. https://doi.org/10.3390/life11090984
Shah JA, Khattak S, Rauf MA, Cai Y, Jin J. Potential Biomarkers of miR-371–373 Gene Cluster in Tumorigenesis. Life. 2021; 11(9):984. https://doi.org/10.3390/life11090984
Chicago/Turabian StyleShah, Junaid Ali, Saadullah Khattak, Mohd Ahmar Rauf, Yong Cai, and Jingji Jin. 2021. "Potential Biomarkers of miR-371–373 Gene Cluster in Tumorigenesis" Life 11, no. 9: 984. https://doi.org/10.3390/life11090984
APA StyleShah, J. A., Khattak, S., Rauf, M. A., Cai, Y., & Jin, J. (2021). Potential Biomarkers of miR-371–373 Gene Cluster in Tumorigenesis. Life, 11(9), 984. https://doi.org/10.3390/life11090984